Introduction
Glioma is a high-grade malignant primary
intracranial tumor, with a high morbidity and mortality rate
(1). Glioblastoma is a type of glioma
characterized by a relatively high degree of malignancy.
Early-stage gliobastoma may lead to severe intracranial
hypertension, and focal symptoms and signs (1). Surgery constitutes the most commonly
used treatment modality in early-stage glioblastoma (1). Combined therapy including radiotherapy,
chemotherapy, gene therapy, immunotherapy and targeted therapy are
employed in later-stage glioblastoma (1). However, the clinical effects of these
therapies and patient prognosis are far from ideal due to the
unmanageable invasiveness and unclear pathogenesis of the disease.
It has been found that the expression of microRNA (miR) is closely
associated with the formation of glioma (2,3). Janus
kinase 2/signal transducer and activator of transcription 3
(JAK2/STAT3) signaling pathways are important processes for the
formation and transmission of tumors (4).
The aim of the present study was to examine the
expression of miR-184 in JAK2/STAT3 signaling pathways in the
formation of glioblastoma to provide a new basis for the
development of the mechanism of glioblastoma.
Materials and methods
Materials
The LN28 glioblastoma cell line was purchased from
the Chinese Academy of Sciences Cell Bank. The following
experimental instruments were used: Micropipette tip (Rainin
Instrument LLC, Oakland, CA, USA), optical microscope (Olympus
Corp., Tokyo, Japan), polymerase chain reaction (PCR) TC-XP (Bioer
Technology Co., Ltd., Hangzhou, China), constant temperature
incubator (Changzhou Huapuda Instrument Co. Ltd., Changzhou,
China), paraffin slicing machine (Leica, Mannheim, Germany) and
tissue embedder (Leica). TRIzol reagent, RNase-free, reverse
transcription (RT)-PCR primers, RT kit, quantitative PCR (qPCR)
kit, Express SYBR-GreenER miR qPCR kits, NCode VILO miR cDNA
(Invitrogen Life Technologies, Carlsbad, CA, USA) and the miR
extraction kit (Roche Diagnostics GmbH, Mannheim, Germany) were
employed. The rabbit polyclonal primary antibodies p-JAK2 (Cat. no.
GTX101132, 1:500) and p-STAT3 (Cat. no. GTX110587, 1:500) were
purchased from Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA.
Experimental process
The cell lines were thawed by removing the cells
from the vial and heated over a water bath for 1 min at 37°C. The
cells were centrifuged for 4 min at 1,000 × g, and the supernatant
was subsequently removed and supplemented with 10% fetal bovine
serum in DMEM culture medium. The cells were cultured at a constant
temperature of 37°C in an incubator with saturated humidity and 5%
CO2. The cells were grown as monolayers and passaged.
The medium was replaced with DMEM (10% FBS), the cells were washed
with PBS for 2 times, and 0.25% pancreatin consisting of EDTA was
added. The cells were then observed under an inverted microscope
(Olympus Corp.), micropippetted and agitated until the cells were
completely removed from the flask. When the surface of the flask
was transparent and without frizz, it indicated that cells were
completely detached from the flask wall. The supernatant was
removed by centrifugation at 8,000 × g for 5 min and the cells were
again cultured and passaged as described above. miR-184 mimics and
their inhibitor were transfected into glioma cells. The LN18 cells
were collected in the logarithmic phase, and the concentration of
cell suspension was adjusted to 3×106/ml. The cells were
cultured with DMEM (10% FBS) in 6-well plates at 37°C containing 5%
CO2 for 12 h until the cell density fused to 70–80%.
Then, 5 µl Lipofectamine® 2000 was added together with 200 µl
serum-free culture medium and the cells were kept at a room
temperature for incubation for 15 min.
Observation index qPCR detection
method
Pancreatin (Boster, Wuhan, China) was used to digest
the glioma cells in the logarithmic phase. The number of cells
counted was approximately 5×106. Phosphate-buffered
saline (PBS) was used to wash the cells, followed by centrifugation
at 1,500 × g for 5 min. TRIzol reagent (1 ml) was subsequently
added, and the cells were micropippetted and agitated prior to cell
suspension. Chloroform (0.2 ml) was added and mixed, and the cells
were again suspended prior to centrifugation. The cells were then
carefully transferred to the upper aqueous phase into a new
RNAase-free centrifuge tube. Without disturbing the white muddy
layer, the same volume of isopropanol was added, and the mixture
was suspended. The cells were again centrifuged, the supernatant
was removed, and the white or transparent precipitation yielded was
RNA. Subsequently, the RNA was washed in 1 ml of 75% ethyl alcohol
and precipitated. The cells were again centrifuged, and the
supernatant was removed. The procedure was repeated, after which
RNA pellet was air-dried and dissolved in DEPC (Boster).
Pre-hybridization was performed according to the instructions
provided with the kit (Boster) prior to the addition of the miR-184
probe to produce the reaction. The probe was preheated with 0.2X
SSC buffer (Amresco, Solon, OH, USA), rinsed, and blocked using
non-specific staining. Biotinylated mouse anti-digoxin polyclonal
antibody (1:200) was added and incubated at room temperature. The
cells were washed with PBS and coloration was developed using DAB,
with the degree of color development occurring under a microscope.
The cells were then washed with tap water to terminate
reaction.
MTT cell proliferation assay
Glioma cells in the logarithmic phase were
collected, and seeded at a density of 5×103 cells in a
96-well plate with RPMI-1640 culture solution supplemented with 10%
fetal calf serum. After culturing for 24 h the supernatant was
removed. Subsequently, 20 µl sterile MTT (5 mg/ml; Sigma, St.
Louis, MO, USA) was added every 24 h into the well to be measured.
Three ventral orifices were set at each time point and monitored
for 6 days consecutively prior to culturing for 4 h in a cell
incubator. The supernatant was completely removed, and DMS0 at 150
µl/hole was added and mixed for 10 min on a shaker (Bio-Rad,
Hercules, CA, USA) until the purple crystal was completely
dissolved. Using a microplate reader (Bio-Rad), absorbance was
measured at a wavelength of 579 nm. The IC50 of the endophytic
fungi extract in in the two types kinds of self healing was
measured and the experiment was repeated three times. The cell
growth inhibiting rate was calculated as: (1-average value in
experimental group/average value in control group) × 100%.
Clone formation
Glioma cells were collected in the logarithmic phase
using regular 0.25% trypsin digestion and passaging for cell
suspension. Cell suspension was diluted to a gradient, and the
cells were inoculated in the culture dish with a certain density.
This was followed by micropippetting and agitation, as well as
dilution and inoculation. The cells were cultured for 2–3 weeks in
a 37°C environment with 5% CO2, and saturated. The
solution was replaced with fresh culture solution according to the
change of pH. Termination of the culture occurred following
identification of clone formation via visual observance. The
culture solution was removed, and the cells were washed twice with
PBS solution and air-dried. This process was followed by fixation
with methyl alcohol for 15 min, after which the methyl alcohol was
removed and the cells were air-dried. Cells were stained with the
appropriate amount of Giemsa for 10–30 min, after which the
staining was washed with water prior to air drying. The plate was
inverted and a grid overlay of a piece of transparent film was
prepared. Cell clones >50 were counted and the colony forming
efficiency was calculated as: (%) = (colony number/inoculation cell
number) × 100. The experiment was repeated three times.
Cell scratch and invasion assays
A line was drawn on the back of pour plate using a
marker pen. Cells were added from the experimental group until each
pour plate had ~5×105 cells. Using a sterile pipette, a
spearhead cut was made along the bottom of the culture pour plate
in a ‘―’ shape. The cells were cultured with DMEM culture solution
comprising 5% calf serum and the cell migration in the scratch area
was was observed. To coat the basilar membrane 50 mg/l artificial
reconstructed basement membrane with Matrigel (dilution, 1:5) was
used to coat the upper surface of 20 membranes (BD Biosciences,
Franklin Lakes, NJ, USA) at the bottom of the Transwell chamber,
and air-dried. For hydration 50 µl serum-free culture solution
supplemented with 10 g/l bovine serum albumin in each plate was
used. The cells were inoculated by adding 500 µl culture medium
outside the chamber, and 100 µl tumor cells within the chamber.
Serum-free culture medium served as the culture solution, followed
by fixing and staining. Briefly, the cells were washed with PBS,
and any cells on the upper layer of the microporous membrane
removed using a cotton swab. The cells were then fixed with
absolute alcohol and the solution was stained with crystal violet.
The cells were visualized under a microscopic. The number of cells
that transferred to the lower layer of the microporous membrane
were counted under an inverted microscope. For each sample 10
visual fields were counted. The procedure was repeated at least
three times and the average number of cells was obtained.
Western blotting
SDS-PAGE electrophoresis on 30 µg total protein and
electrotransferred onto a nitrocellulose membrane (NC). The NC
membrane was placed in Tris-buffered saline and Tween-20 (TBST)
containing 50 g/l skim milk powder at room temperature. As per the
pre-stained Marker, the NC membrane was cut into different strips
based on molecular weight and inoculated in TBST solution
containing p-JAK2 and p-STAT3 primary antibodies at 4°C overnight.
The following day, the membrane was washed with TBST on a shaker
and the strips were inoculated with 1:5,000 corresponding mouse
anti-rabbit monoclonal secondary antibody. The membrane was placed
into the electrophoresis gel imaging (Bio-Rad) and the images
developed using ECT luminous solution. Gray value of the strips
were subsequently calculated.
Statistical analysis
SPSS 19.0 software (SPSS, Inc., Chicago, IL, USA)
was applied to record and process the data. Measurement data were
presented as means ± standard deviation. One-way variance analysis
was applied to determine comparisons between groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Comparison of the expression of
miR-184, cell proliferation, clone and invasion ability
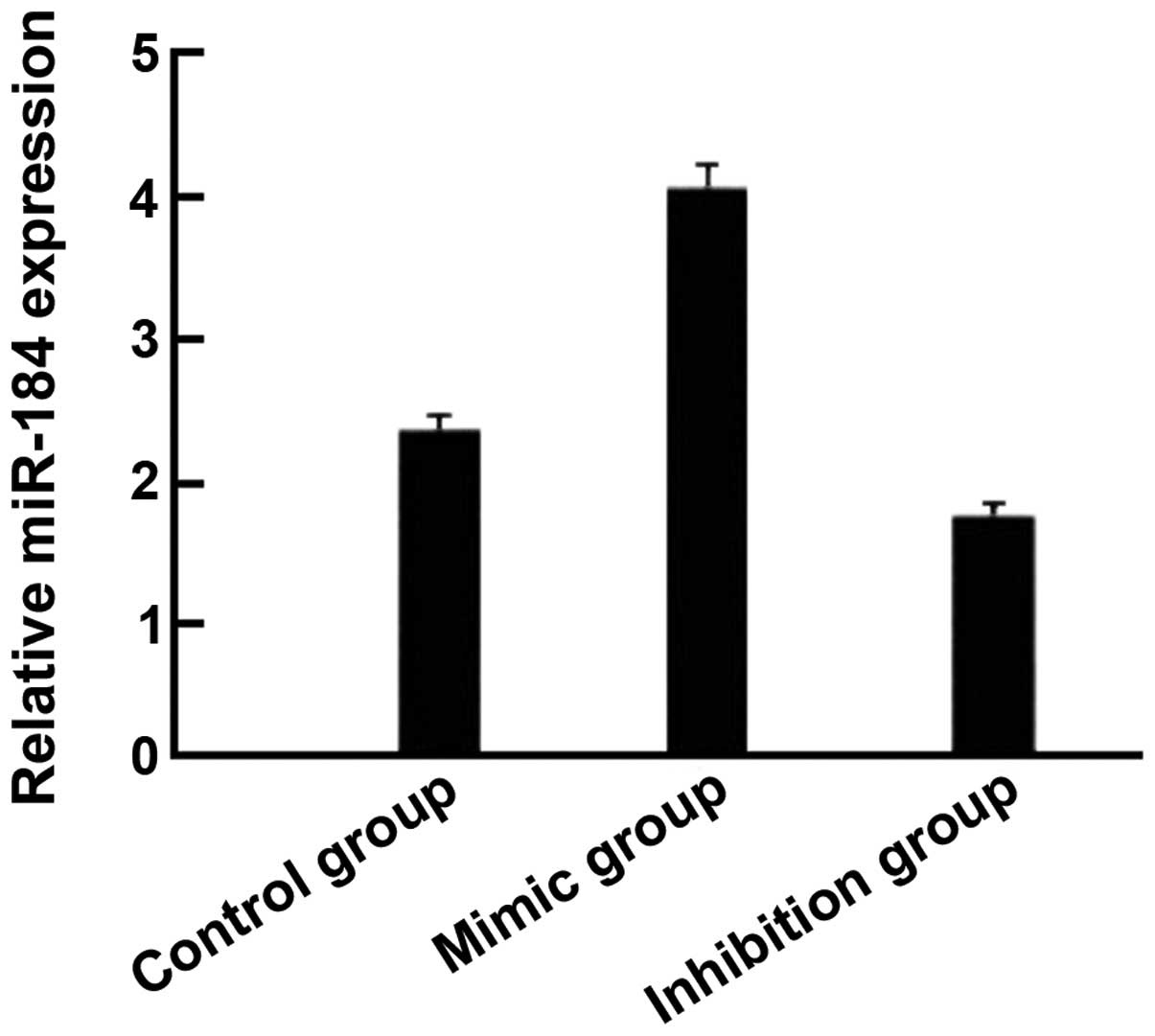
Compared with the control group, the expression of
miR-184 in the miR-184 mimic group was significantly increased
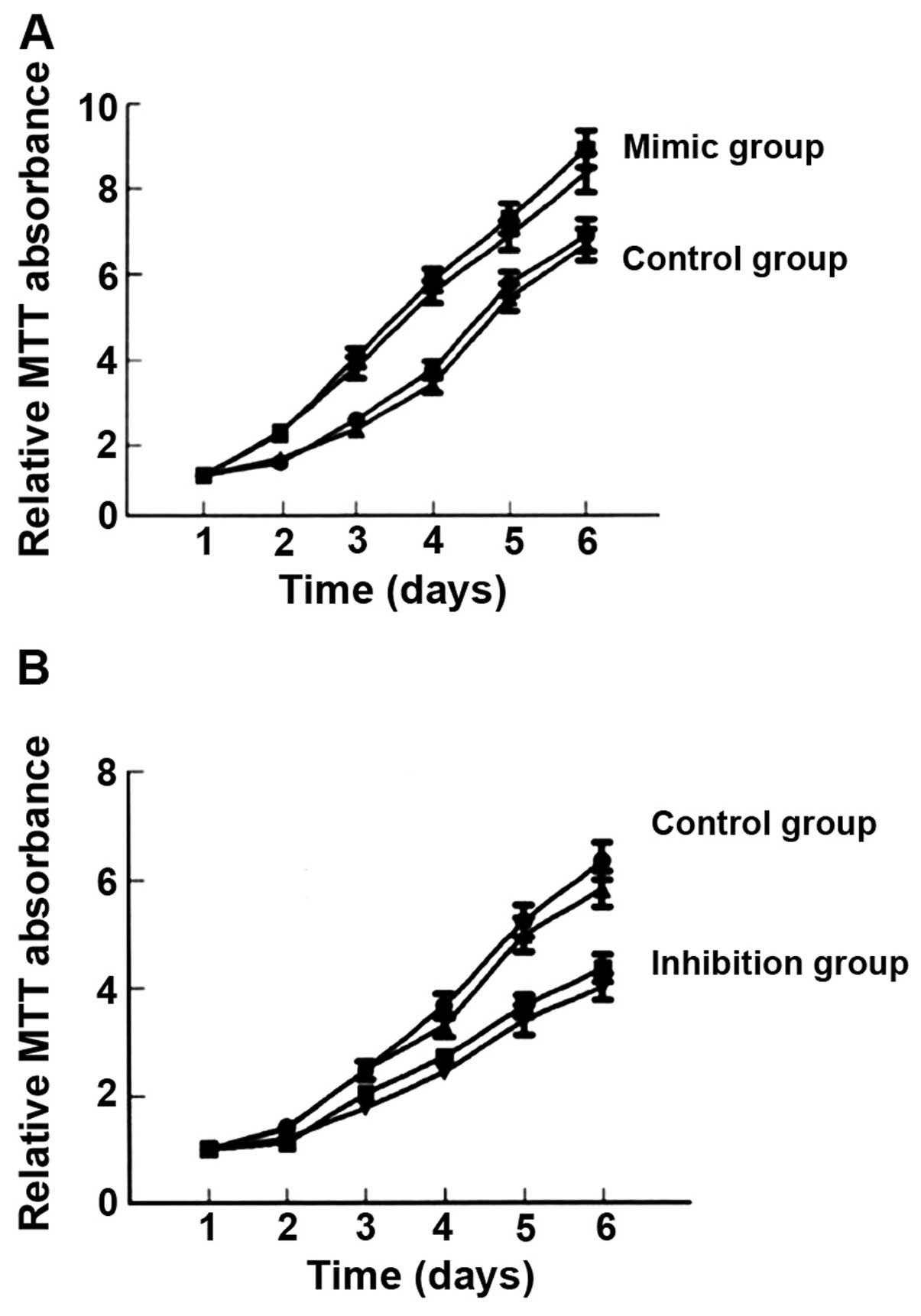
(Fig. 1) and cell proliferation
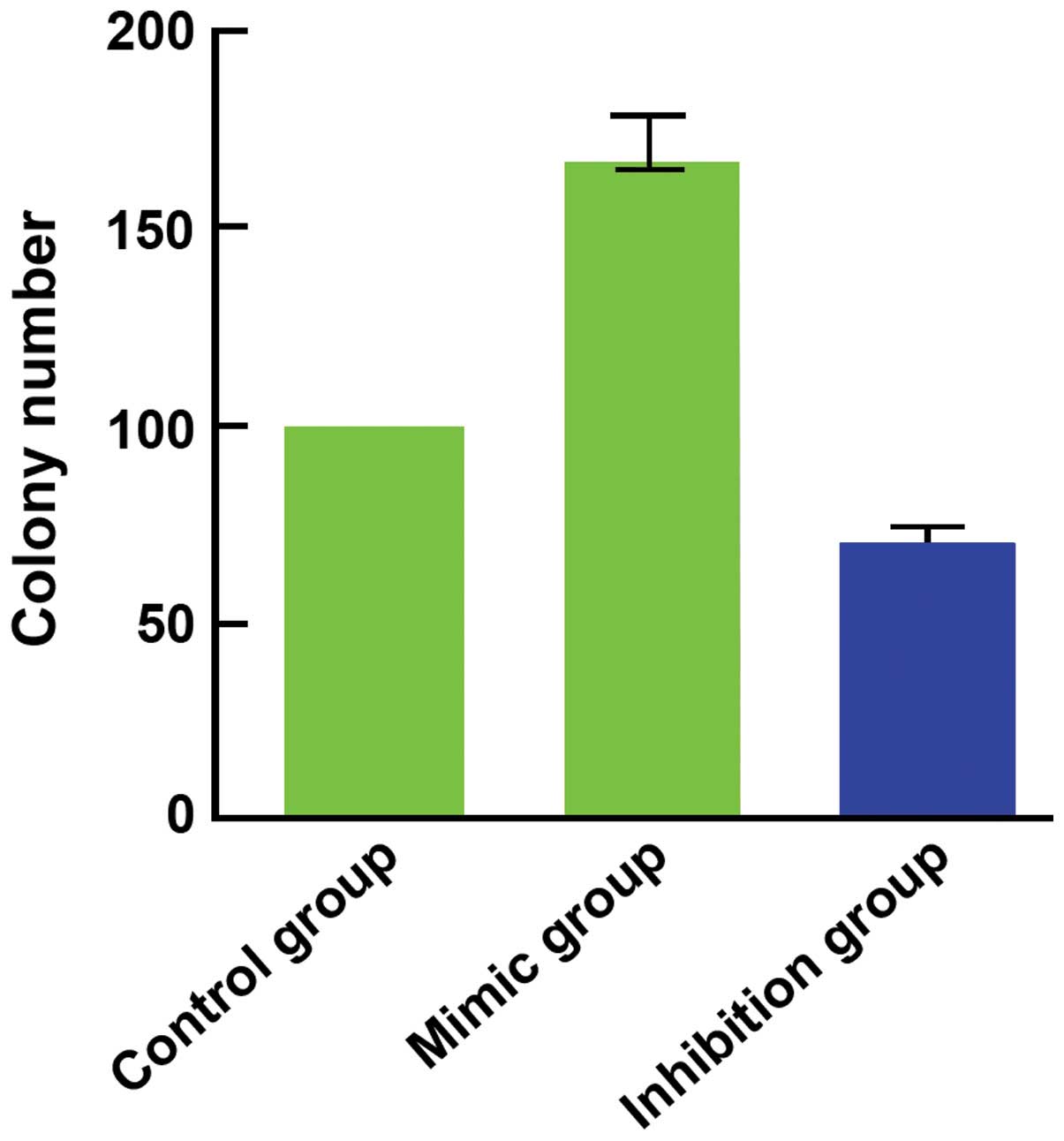
(Fig. 2), as well as clone (Fig. 3) and invasion ability (Fig. 4) were enhanced. By contrast, the
expression of miR-184 in the miR-184 inhibition group was reduced
compared with the control group (Fig.
1) and cell proliferation (Fig.
2B), as well as clone (Fig. 3)
and invasion ability (Fig. 4) were
decreased. Differences were statistically significant (P<0.05).
The number of cells permeating the septum in the control group was
98.3±12.4; in the miR-184 mimic, 115.6±15.7; and in the inhibition
group, 85.7±11.3. Comparisons of the number of cells permeating the
septum between groups were statistically significant (F=5.126,
P<0.001) (Figs. 1–4).
Comparison on the expression level of
p-JAK2 and p-STAT3
Compared with the control group (256.7±43.6,
243.9±34.2), the expression level of p-JAK2 (345.8±49.7) and
p-STAT3 (315.2±32.1) in the mimic group was significantly higher
whereas that in the inhibition group (194.2±36.9, 167.8±39.5) was
significantly lower. The differences were statistically significant
(P<0.05) (Table I).
 | Table I.Comparison on the expression level of
p-JAK2 and p-STAT3. |
Table I.
Comparison on the expression level of
p-JAK2 and p-STAT3.
| Groups | p-JAK2 (%) | p-STAT3 (%) |
|---|
| Control group | 256.7±43.6 | 243.9±34.2 |
| Mimic group | 345.8±49.7 | 315.2±32.1 |
| Inhibition group | 194.2±36.9 | 167.8±39.5 |
| F | 5.627 | 6.105 |
| P-value | <0.001 | <0.001 |
Discussion
MicroRNA is a type of non-protein coding
micromolecule short-chain RNA that has a regulatory function and is
widely identified in animals and plants. miR is closely associated
with the occurrence and development of glioma (5). miR affects sequence specificity,
regulates gene expression and inhibits gene expression following
transcription through post-transcriptional control, thereby
participating in the regulation of cell growth, proliferation,
differentiation, metabolism and apotosis (6). Although a number of miR have been
identifed, not all have an open reading frame. miRs may be present
in eukaryotic genomes yet not encode protein. Their lengths are
similar at approximately 18–22 nt. miRs belong to short sequence
micromolecule RNA (7). Previous
findings (8,9) have shown that the expression of several
miR in glioma was different from those in normal cerebral tissue,
for example, the expression of miR-21 and miR-9-2 was upregulated
while that of miR-128-1, miR-181a, miR-181b, and miR-181c was
downregulated. Other findings have shown that the expression of 11
types of miR was downregulated, including miR-10b and miR-21
(2). Certain miR were able to promote
glioma invasion and their expression in glioma was significantly
upregulated. Typical examples included miR-10-b, miR-21 and
miR-221/222 (10). By contrast,
certain miR were able to inhibit glioma invasion. Therefore, the
role miR plays as a target for tumor diagnosis and treatment is
crucial (11).
The expression of miR-184 is abnormal in various
tumors, including hepatic carcinoma, lung carcinoma, nasopharyngeal
carcinoma, and adrenal gland neoplasms. Additionally, miR-184 may
regulate the formation mechanism of tumor through the JAK2/STAT3
signaling pathway. JAK2/STAT3 is an important signaling pathway of
the JAK/STAT family. The JAK2/STAT3 channel is closely associated
with inflammatory reaction, oxidative stress, cell injury and
apoptosis. Lung cancer tolerance was reversed in patients
undergoing radiotherapy by adopting niclosamide to destroy STAT3
phosphorylation (12). Previous
findings on hematological malignancy have shown that leukemia
partially resulted from overexpression or continuous activation of
a number of growth factor receptors and their downstream enzymes,
which were incurred from gene mutation or chromosome translocation.
The joint effects of multiple channels, including JAK/STAT would
lead the cells to proliferate uncontrollably and finally resulted
in leukemia (13).
The results of the present study have shown that
compared with the control group, the expression of miR-184 in the
miR-184 mimic group was increased, and the properties of cell
proliferation, clone and invasion ability were enhanced. The number
of cells penetrating the septum, as well as the expression of
p-JAK2 and p-STAT3 protein were increased and the differences were
statistically significant. By contrast, compared with the control
group, the expression of miR-184 in the miR-184 inhibition group
was decreased, and cell proliferation, as well as clone and
invasion ability were reduced. The number of cells penetrating the
septum, as well as the expression of p-JAK2 and p-STAT3 protein
were decreased and differences were statistically significant. In
conclusion, miR-184 may be involved in the formation of
glioblastoma and influence the expression of the JAK2/STAT3
signaling pathway.
References
|
1
|
Waghmare I, Roebke A, Minata M,
Kango-Singh M and Nakano I: Intercellular cooperation and
competition in brain cancers: Lessons from Drosophila and human
Studies. Stem Cells Transl Med. 3:1262–1268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li R and Li X, Ning S, Ye J, Han L, Kang C
and Li X: Identification of a core miRNA-pathway regulatory network
in glioma by therapeutically targeting miR-181d, miR-21, miR-23b,
β-Catenin, CBP, and STAT3. PLoS One. 9:el019032014.
|
|
3
|
Northcott PA, Fernandez-L A, Hagan JP,
Ellison DW, Grajkowska W, Gillespie Y, Grundy R, Van Meter T, Rutka
JT, Croce CM, et al: The miR-17/92 polycistron is up-regulated in
sonic hedgehog-driven medulloblastomas and induced by N-myc in
sonic hedgehog-treated cerebellar neural precursors. Cancer Res.
69:3249–3255. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duan W, Yang Y, Yan J, Yu S, Liu J, Zhou
J, Zhang J, Jin Z and Yi D: The effects of curcumin post-treatment
against myocardial ischemia and reperfusion by activation of the
JAK2/STAT3 signaling pathway. Basic Res Cardiol. 107:2632012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li T, Li D, Sha J, Sun P and Huang Y:
MicroRNA-21 directly targets MARCKS and promotes apoptosis
resistance and invasion in prostate cancer cells. Biochem Biophys
Res Commun. 383:280–285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xia H, Qi Y, Ng SS, Chen X, Li D, Chen S,
Ge R, Jiang S, Li G, Chen Y, et al: microRNA-146b inhibits glioma
cell migration and invasion by targeting MMPs. Brain Res.
1269:158–165. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Garajová I, Le Large TY, Frampton AE,
Rolfo C, Voortman J and Giovannetti E: Molecular mechanisms
underlying the role of microRNAs in the chemoresistance of
pancreatic cancer. BioMed Res Int. 2014:6784012014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saadatian Z, Masotti A, Saleh Nariman Fam
Z, et al: Single-nucleotide polymorphisms within micrornas
sequences and their 3′ utr target sites may regulate gene
expression in gastrointestinal tract cancers. Iran Red Crescent Med
J. 16:el66592014. View Article : Google Scholar
|
|
9
|
Orang AV and Barzegari A: MicroRNAs in
colorectal cancer: From diagnosis to targeted therapy. Asian Pac J
Cancer Prev. 15:6989–6999. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pang JC, Kwok WK, Chen Z and Ng HK:
Oncogenic role of microRNAs in brain tumors. Acta Neuropathol.
117:599–611. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang Y, Duan W, Jin Z, Yi W, Yan J, Zhang
S, Wang N, Liang Z, Li Y, Chen W, et al: JAK2/STAT3 activation by
melatonin attenuates the mitochondrial oxidative damage induced by
myocardial ischemia/reperfusion injury. J Pineal Res. 55:275–286.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
You S, Li R, Park D, Xie M, Sica GL, Cao
Y, Xiao ZQ and Deng X: Disruption of STAT3 by niclosamide reverses
radioresistance of human lung cancer. Mol Cancer Ther. 13:606–616.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Steelman LS, Abrams SL, Whelan J, Bertrand
FE, Ludwig DE, Bäsecke J, Libra M, Stivala F, Milella M, Tafuri A,
et al: Contributions of the Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and
Jak/STAT pathways to leukemia. Leukemia. 22:686–707. 2008.
View Article : Google Scholar : PubMed/NCBI
|