Introduction
Hepatocellular carcinoma (HCC) is the fifth most
frequent cancer and the third most common cause of
cancer-associated mortality worldwide, with >600,000 mortalities
reported annually (1,2). Over the last few decades, the incidence
of HCC has increased in eastern Asia and sub-Saharan Africa
(3,4);
the estimated number of new cases diagnosed annually increased from
437,000 to 564,000 between 1990 and 2000 (1,4). Due to
the highly aggressive nature of the tumor, and because tumors are
highly resistant to traditional treatments, such as chemotherapy
and radiation, the 5-year survival rate of HCC is poor, with an
overall survival rate of <16% (5).
In addition, surgical resection and liver transplantation are
restricted, and are suitable only for patients diagnosed with early
stage disease (6). It is generally
recognized that exploring the underlying molecular mechanisms of
HCC initiation and progression in order to search for functional
molecular targets may provide a new approach for HCC treatment.
MicroRNAs (miRNAs) are small, endogenous, non-coding
RNAs, which act as key post-transcriptional regulators of
target-gene expression. They bind primarily to 3′-untranslated
regions (3′-UTRs) of target gene mRNAs. This process leads to
translational repression or mRNA cleavage (7,8). Recent
studies have reported that >1,000 human miRNAs have been
identified that regulate ~1/3 of the coding genes in the human
genome (9). Many of these miRNAs act
as tumor suppressors and/or oncogenes, and are involved in cell
activities that include development, differentiation,
proliferation, apoptosis, metabolism and immunity (7,10,11). Recently, an increasing number of
studies have revealed that microRNA-497 (miR-497) levels are
decreased in tumors, and that it functions as a tumor suppressor in
a number of types of human cancer, including colorectal, gastric,
cervical and breast cancers, adrenocortical carcinoma and melanoma
(12–20). Similarly, a study by Furuta et
al (21) indicated that miR-497
targets multiple cell cycle regulators and suppresses cell cycle
progression in vitro. However, whether miR-497 regulates
other target genes in HCC is unknown.
Insulin-like growth factor-1 receptor (IGF-1R) is a
member of the receptor tyrosine kinase family and contains two
extracellular α subunits (including the ligand-binding site) and
two β subunits (with intracellular tyrosine kinase activity)
(22,23). IGF-1R has an important role in
malignant cell growth and survival, and is highly expressed in
malignant tumors in the nervous system, liver, thymus, adrenal
cortex, gallbladder, colon, pancreas and lung (24–28).
Furthermore, IGF-1R is crucial for activating the phosphoinositide
3-kinase (PI3K)/Akt pathway (29,30), which
promotes cell proliferation and survival, and is activated by
numerous growth factor receptor tyrosine kinases (31–33).
IGF-1R mRNA and protein expression are increased in HCC and are
closely associated with the progression of malignant tumors
(34,35). To date, studies have demonstrated that
miR-497 targets IGF-1R and has a tumor suppressive role in human
cervical cancer (20) and in
colorectal cancer (12). However,
whether miR-497 functions as a tumor suppressor by directly
targeting IGF-1R in HCC remains unclear.
In the current study, the expression levels of
miR-497 and IGF-1R were examined in HCC cells and tumor samples.
miR-497 overexpression was found to inhibit cell growth, reduce
IGF-1R expression and decrease PI3K/Akt pathway activation.
Although downregulation of miR-497 contributed to malignant
behavior in HCC cells, it increased IGF-1R expression and elevated
activation of PI3K/Akt signaling. These results suggest that
miR-497 functions as tumor suppressor by targeting IGF-1R in
HCC.
Materials and methods
Ethics statement
The Institutional Animal Care and Use Committee at
Nanjing Medical University (Nanjing, China) approved the study
protocols for experiments involving human tissue and animals.
Tissue specimens
A total of 60 paired HCC and adjacent non-tumor
tissues were evaluated for the expression of miR-497 and IGF-1R.
Adjacent non-tumor tissues were ≥2 cm away from the edge of the
tumors. All tissues were obtained from patients that had undergone
partial hepatectomy at the First Affiliated Hospital of Nanjing
Medical University between December 2011 and February 2014. The
patient cohort included 49 males and 11 females, with a median age
of 58 years (range, 36–66 years). Of the 60 patients, 18 patients
exhibited stage A disease, 28 patients exhibited stage B and 14
patients exhibited stage C disease, according to the Barcelona
Clinic Liver Cancer staging system (36). A total of 35 cases exhibited an HCC
mass >5 cm in diameter, while 25 cases exhibited an HCC mass
with a diameter of <5 cm. Patients that had undergone treatment
prior to surgery were excluded from the study. HCC specimens and
adjacent non-tumor tissues were confirmed by pathological
examination, and immediately stored in liquid nitrogen post
surgery. Written informed consent was obtained from either the
patient or the families of the patients.
Liver cancer cell lines
Human HCC cell lines (YY-8103, HepG2, Hep3B,
SMMC-7721 and MHCC-97H) and normal human liver cells (L02) were
obtained from the Department of Liver Transplantation Center at the
First Affiliated Hospital of Nanjing Medical University (Nanjing,
China). Cell lines were cultured in Dulbecco's modified Eagle's
medium (DMEM) containing 10% fetal bovine serum (FBS) (both
HyClone™; GE Healthcare Life Sciences, Logan, UT, USA), penicillin
(50 U/ml; (Invitrogen™; Thermo Fisher Scientific, Inc., Carlsbad,
CA, USA) and streptomycin (50 µg/ml; Invitrogen™; Thermo Fisher
Scientific, Inc.) and propagated in 5% CO2 in a 37°C
humidified incubator.
Reverse transcription
(RT)-quantitative polymerase chain reaction (qPCR)
Total RNA from liver tissue samples and HCC cell
lines were extracted using Invitrogen™ TRIzol reagent (Thermo
Fisher Scientific, Inc.), following the manufacturer's
instructions. To determine miRNA expression, total RNA (1
µl/sample) was reverse-transcribed using miRNA-specific stem-loop
RT primers, reverse transcriptase, RT buffer, dNTPs and an RNase
inhibitor, according to the manufacturer's instructions (TaqMan®
MicroRNA Reverse Transcription Kit; Applied Biosystems™; Thermo
Fisher Scientific, Inc.). qPCR was performed using an Applied
Biosystems™ StepOnePlus™ Real-Time PCR System (Thermo Fisher
Scientific, Inc.). The 20-µl reaction system contained the
corresponding cDNA (2 µl), miRNA-specific TaqMan® primers (1 µl),
TaqMan® Universal PCR Master Mix (10 µl) and ddH2O (7
µl) (Applied Biosystems™; Thermo Fisher Scientific, Inc.). The PCR
conditions were 95°C for 10 min, followed by 50 cycles at 95°C for
15 sec and 60°C for 1 min. RNU6B was used as an endogenous
housekeeping control for data normalization of miRNA levels. The
comparative threshold cycle (Ct) method was used to measure the
relative changes in expression (37);
2−∆∆Ct represents the fold-change of expression.
Immunohistochemical staining
All tissues were paraffin-embedded and obtained from
the Department of Pathology at the First Affiliated Hospital of
Nanjing Medical University. Paraffin-embedded tissues were cut into
4-µm sections, and incubated with the rabbit anti-human IGF-1R
polyclonal antibody (cat. no. ab39398; Abcam, Cambridge, MA, USA;
dilution, 1:100) overnight at 4°C. SP-9000 Histostain™-Plus kits
(ZSGB-BIO, Beijing, China) were used according to the
manufacturer's instructions. Scoring was measured according to the
cell cytoplasm staining pattern: 0, no cytoplasmic staining; 1,
weak cytoplasmic staining; 2, moderate cytoplasmic staining; and 3,
strong cytoplasmic staining.
Cell transfection
The miR-497 mimic, miR-497 inhibitor, miRNA mimic
negative control (NC) (a miRNA mimic) and miRNA inhibitor NC were
purchased from Shanghai GenePharma Co., Ltd. (Shanghai, China) and
were transfected using Lipofectamine® 2000 transfection reagent
(Invitrogen™; Thermo Fisher Scientific) according to the
manufacturer's instructions. The sequences were as follows: miR-497
mimic, 5′-CAGCAGCACACUGUGGUUUGU-3′; NC,
5′-UUCUCCGAACGUGUCACGUTT-3′; miR-497 inhibitor,
5′-ACAAACCACAGUGUGCUGCUG-3′; and miRNA inhibitor NC,
5′-CAGUACUUUUGUGUAGUACAA-3′.
Cell proliferation assays
Cells were seeded at a density of 2,000–5,000
cells/well in 96-well plates in 100 µl complete media. Cell
Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) was
used to measure cell viability according to the manufacturers
instructions. Briefly, cells were seeded at a density of
2,000–5,000 cells/well in 96-well plates in 100 µl DMEM containing
10% FBS (HyClone™; GE Healthcare Life Sciences), penicillin (50
U/l; Invitrogen™; Thermo Fisher Scientific, Inc.) and streptomycin
(50 µg/ml; Invitrogen™; Thermo Fisher Scientific, Inc.) and
cultured for 6 days in 5% CO2 in a 37°C humidified
incubator. Next, 10 µl CCK-8 solution was added to each well and
incubated at 37°C for 1 h. The absorbance was then calculated at a
wavelength of 450 nm using a microplate reader (ELX808; BioTek
Instruments, Inc., Winooski, VT, USA). Each experiment was repeated
at least three times.
Colony formation assays
To examine the effect of upregulated or
downregulated miR-497 expression on the proliferation of HCC cell
lines, cells transfected with miR-497 mimics, miR-497 inhibitor, NC
and miRNA inhibitor NC were used for colony formation assays. Each
cell type was seeded into 6-well plates (500 cells/well) and
cultured for 3 weeks. Cultures were stained with 0.4% crystal
violet (Beyotime Biotech, Jiangsu, China). Colonies of >2 mm
were counted and the mean number of colonies per well was
calculated from three wells for each experiment. Each experiment
was repeated at least three times.
Soft agar colony formation assays
Cells were transfected with miR-497 mimics, miR-497
inhibitor, NC and miRNA inhibitor NC, suspended in 0.5 ml of 1% low
melting point agarose with complete culture media, and layered on
top of 0.5 ml of 2% low melting point agarose (Department of Liver
Transplantation Center, The First Affiliated Hospital of Nanjing
Medical University) in 24-well plates. Cell counts varied from
2,000 to 5,000 cells depending on the cell line. Plates were
incubated for 2 weeks in a 37°C humidified incubator with 5%
CO2. Colonies in at least 6 random microscopic fields
were counted (Nikon Eclipse 50i; Nikon Corporation, Tokyo, Japan)
and photographed (AxioCam MRc5; Carl Zeiss Shanghai Co., Ltd.,
Shanghai, China). All experiments were repeated three times.
Tumorigenicity assays in nude
mice
Male BALB/c nude mice (aged 3–4 weeks) were
purchased from the Department of Laboratory Animal Center of
Nanjing Medical University. Cells with differential miR-497
expression were injected subcutaneously into the lateral root of
the anterior limb of the nude mice (5.00×106
cells/mouse; 6 mice/experimental group). Tumor size was measured
every third day following injection. At 3 weeks after injection,
mice were sacrificed via cervical dislocation and tumors were
excised and photographed. The weights of the tumors were also
recorded. Experimental animals were maintained in accordance with
Institutional Animal Care and Use Committee guidelines (38).
Western blot analyses
Cell lysates were prepared using cold lysis buffer
containing 25 mM Tris-Cl (pH 7.5), 5 mM EDTA, 1% sodium dodecyl
sulfate (SDS), and a protease inhibitor cocktail (Sigma-Aldrich,
St. Louis, MO, USA). Protein concentration was subsequently
determined using a bicinchoninic acid assay kit (Thermo Fisher
Scientific, Rockford, IL, USA). After boiling for 5 min, samples
were subjected to 10% SDS-polyacrylamide gel electrophoresis and
transferred to polyvinylidene difluoride membranes (EMD Millipore,
Billerica, MA, USA). Membranes were blocked for 1 h at room
temperature with 5% blocking buffer (Inner Mongolia Yili Industrial
Group Co., Ltd., Inner Mongolia, China), washed three times with
Tris-buffered saline containing 0.1% Tween-20, and incubated
overnight at 4°C with rabbit anti-human polyclonal primary
antibodies against IGF-1R (cat. no. ab39398; dilution, 1:100;
Abcam), p21 (cat. no. ab109199; dilution, 1:1,000; Abcam), p27
(cat. no. ab32034; dilution, 1:1,000; Abcam), phospho- (p-) Ser473
Akt (cat. no. ab126433; dilution, 1:1,000; Abcam), p-glycogen
synthase kinase 3 β (GSK3β; cat. no. ab75745; dilution, 1:1,000;
Abcam) and β-actin (cat. no. ab119716; dilution, 1:1,000; Abcam).
After the membranes were washed, they were incubated for 1 h at
room temperature with the mouse anti-rabbit IgG secondary antibody
(cat. no. bs-0295M; dilution, 1:1,000; Beijing Biosynthesis
Biotechnology Co., Ltd., Beijing, China). Proteins were detected by
enhanced chemiluminescence (ECL) using a Pierce™ ECL Western
Blotting detection system (Thermo Fisher Scientific, Inc.,
Rockford, IL, USA). β-actin was used as the internal control.
Statistical analyses
Statistical analyses were performed using SPSS
version 18.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA). Quantitative
data are presented as the mean ± standard deviation. Differences
between two groups were assessed using a Students t-test
(two-tailed). P<0.05 was considered to indicate statistically
significant differences.
Results
HCC exhibits decreased miR-497
expression and increased IGF-1R expression
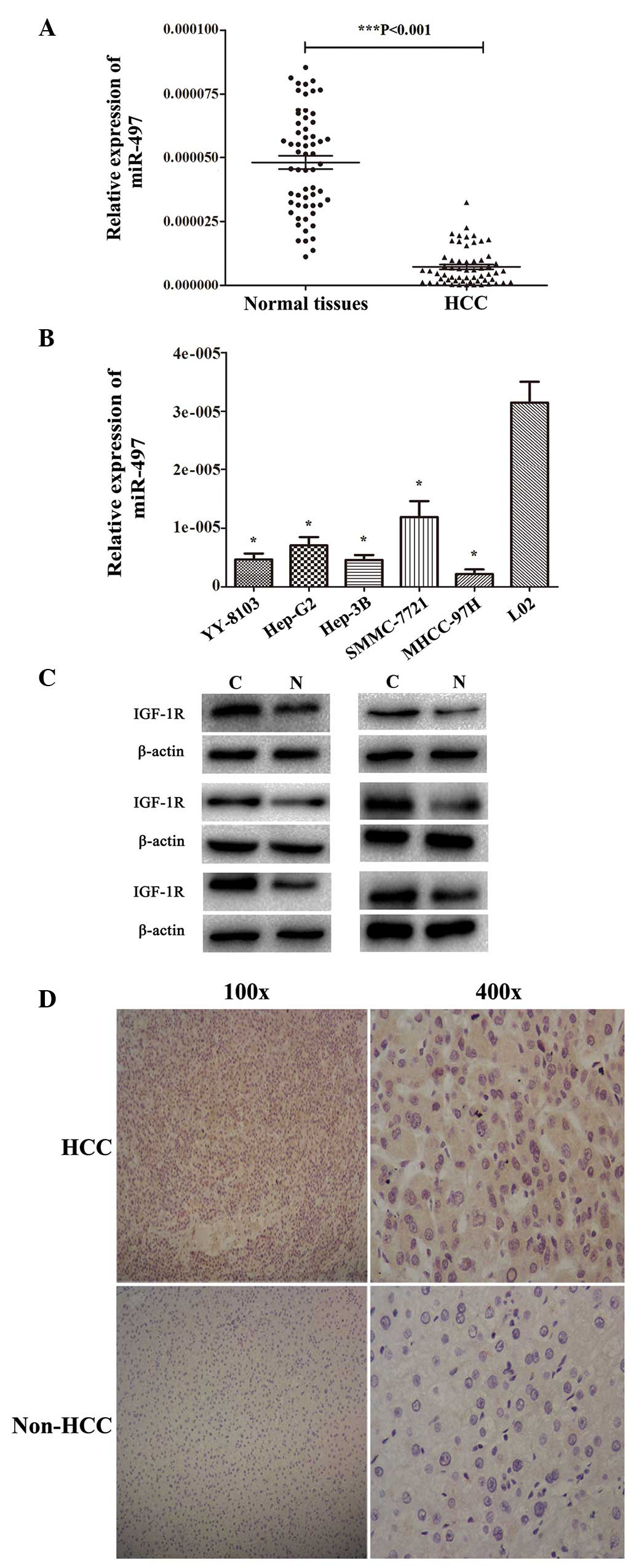
qPCR was performed to measure miR-497 expression
levels in HCC and adjacent non-cancerous liver tissues from 60
patients. miR-497 expression was observed to be decreased compared
with matched normal liver tissues in 42/60 (70.0%) HCC specimens
(P<0.001; Fig. 1A). Next, the
expression of miR-497 was evaluated in five HCC-derived cell lines
using qPCR. miR-497 expression was significantly reduced in all
HCC-derived cell lines (YY-8103, HepG2, Hep3B, SMMC-7721 and
MHCC-97H) compared with L02 normal human liver cells (Fig. 1B). Of the five HCC-derived cell lines,
MHCC-97H cells exhibited the lowest level of miR-497 expression,
whereas SMMC-7721 exhibited the highest level of miR-497 expression
(Fig. 1B). One of the predicted
common targets of miR-497 is IGF-1R (http://mirtarbase.mbc.nctu.edu.tw/php/search.php?q=search_exact&searchword=hsa-miR-497-5p).
Therefore, western blot analysis was conducted to measure IGF-1R
protein levels in HCC and adjacent non-cancer tissue. The results
indicated that IGF-1R protein expression was increased in HCC
specimens compared with matched normal liver tissues (Fig. 1C). Immunohistochemistry was also used
to evaluate IGF-1R protein expression in HCC specimens and paired
normal tissues in the same 60 matched samples. Of these specimens,
24/60 (40.0%) cancerous specimens exhibited no or weak positive
staining, whereas 45/60 (75.0%) non-HCC tissues showed no or weak
positive staining (Fig. 1D).
Collectively, these data indicate that miR-497 expression is
decreased and IGF-1R expression is increased in HCC tissues.
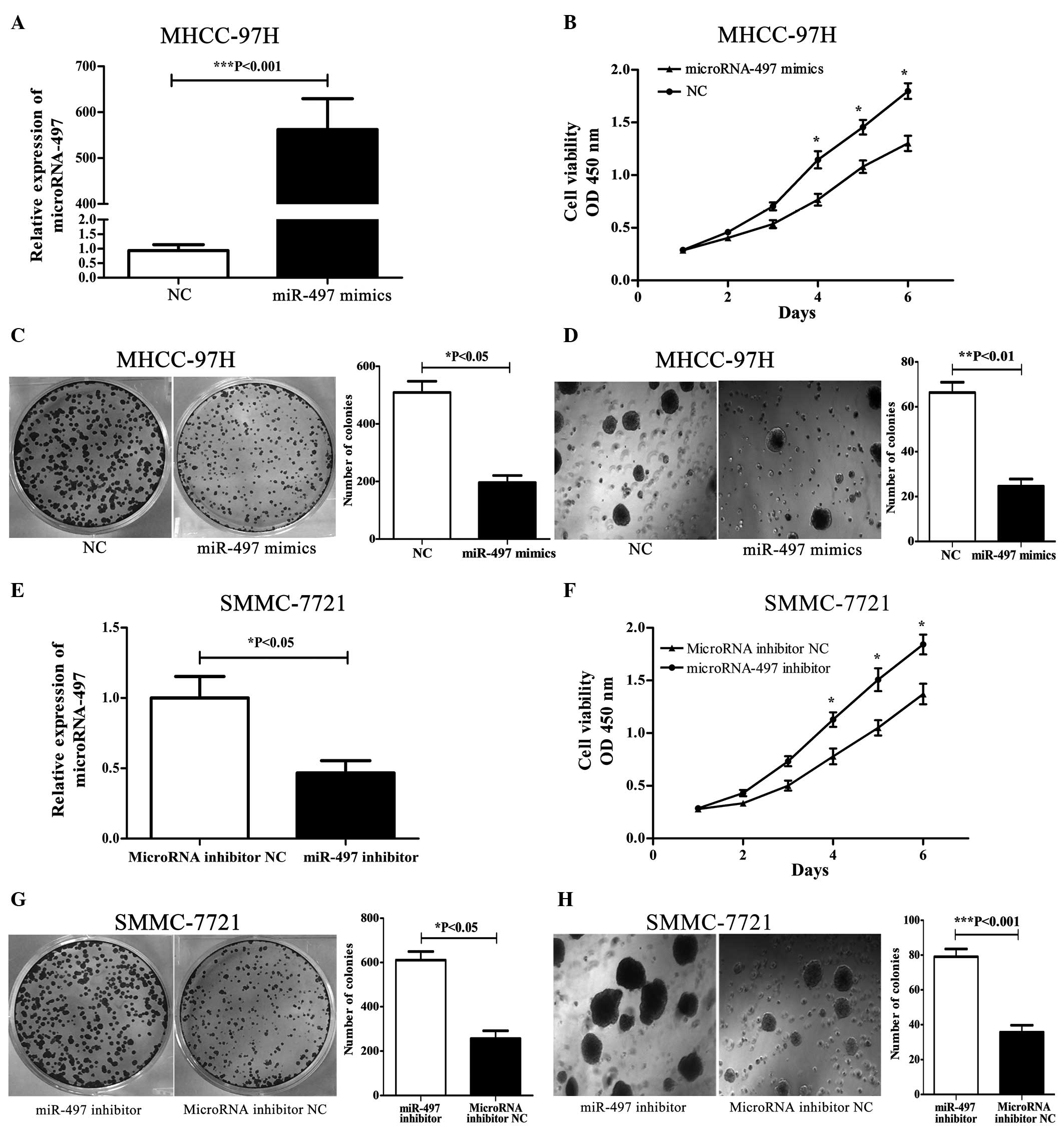
miR-497 overexpression inhibits
proliferation and colony formation in MHCC-97H cells
To overexpress miR-497, miR-497 mimics were
transfected into MHCC-97H cells, which exhibited the lowest level
of miR-497 expression among the 5 HCC-derived cell lines. qPCR was
used to evaluate miR-497 expression in transfected cells at 24 h
post transfection. In miR-497 mimic-transfected cells, miR-497
expression was significantly higher compared with that of miR-497
NC-transfected cells (Fig. 2A). To
investigate the effects on proliferation in miR-497
mimic-transfected cells, cell growth was monitored for 6 days.
miR-497 mimic-transfected MHCC-97H cells exhibited significantly
reduced cell proliferation compared with that of miR-497
NC-transfected cells (Fig. 2B)
(P<0.05). MHCC-97H cells with upregulated miR-497 expression
were subjected to colony formation assays. As shown in Fig. 2C, miR-497 overexpression in MHCC-97H
cells significantly inhibited colony formation relative to MHCC-97H
cells transfected with miR-497 NC (P<0.05); furthermore, the
majority of clones were smaller than those of control cells. Next,
soft agar assays were utilized to assess colony formation; these
are the most stringent assays for detecting the proliferative
ability of cells (39). Reduced
colony formation was observed in soft agar (Fig. 2D) that had been seeded with MHCC-97H
cells transfected with miR-497 mimics, compared with that seeded
with miR-497 NC-transfected cells (P<0.01). These results
indicate that miR-497 inhibits tumor cell growth in
vitro.
miR-497 knockdown promotes
proliferation and colony formation in SMMC-7721 cells
To knockdown miR-497, a miR-497 inhibitor was
transfected into SMMC-7721 cells, which exhibited the highest level
of miR-497 expression among the 5 HCC-derived cell lines. qPCR was
performed to assess the efficiency of miR-497 knockdown in these
cells, confirming that miR-497 expression in miR-497
inhibitor-transfected cells was significantly lower compared to
miRNA inhibitor NC-transfected cells (Fig. 2E). miR-497 inhibitor-transfected
SMMC-7721 cells (Fig. 2F) exhibited
enhanced cell proliferation compared with miRNA inhibitor
NC-transfected cells (P<0.05). Next, SMMC-7721 cells with
downregulated miR-497 expression were subjected to colony formation
assays. As shown in Fig. 2G,
decreased miR-497 expression in SMMC-7721 cells significantly
promoted colony formation relative to cells transfected with the
miRNA inhibitor NC (P<0.05). Enhanced colony formation in soft
agar (Fig. 2H) was also observed in
SMMC-7721 cells transfected with the miR-497 inhibitor compared
with miRNA inhibitor NC-transfected cells (P<0.001).
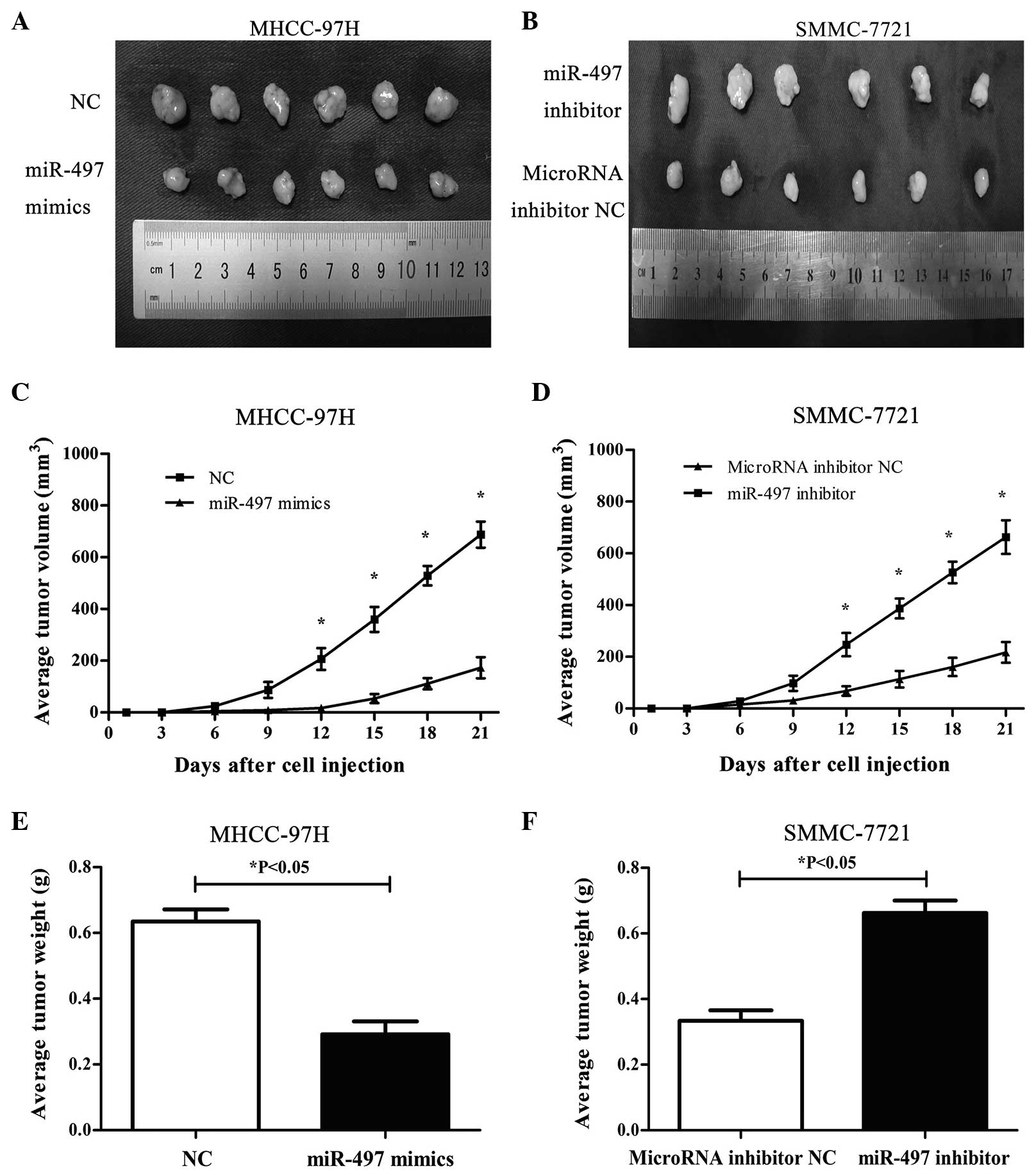
Differential expression of miR-497
affects tumorigenesis in nude mice
The effects of differential miR-497 expression on
the tumorigenic potential of HCC cells were investigated in
vivo. MHCC-97H cells with upregulated miR-497 expression and
SMMC-7721 cells with downregulated miR-497 expression were injected
subcutaneously into BALB/c nude mice. Tumor size was measured on
every third day following injection. After 3 weeks, mice were
sacrificed and the excised tumors were photographed and weighed.
Results are shown in Fig. 3. Compared
to mice injected with MHCC-97H cells transfected with miR-497 NC,
mice injected with MHCC-97H cells overexpressing miR-497 exhibited
smaller tumors during the same time period, and the mean tumor
volumes and weights were significantly lower than the control group
(P<0.05) (Fig. 3A, C, and E).
Compared with mice injected with miRNA inhibitor NC-transfected
SMMC-7721 cells, mice injected with miR-497-underexpressing
SMMC-7721 cells exhibited an increased capacity for tumorigenesis
(P<0.05) (Fig. 3B, D and F). Taken
together, these results strongly suggest that miR-497 inhibits
tumor cell growth and tumorigenicity in vivo.
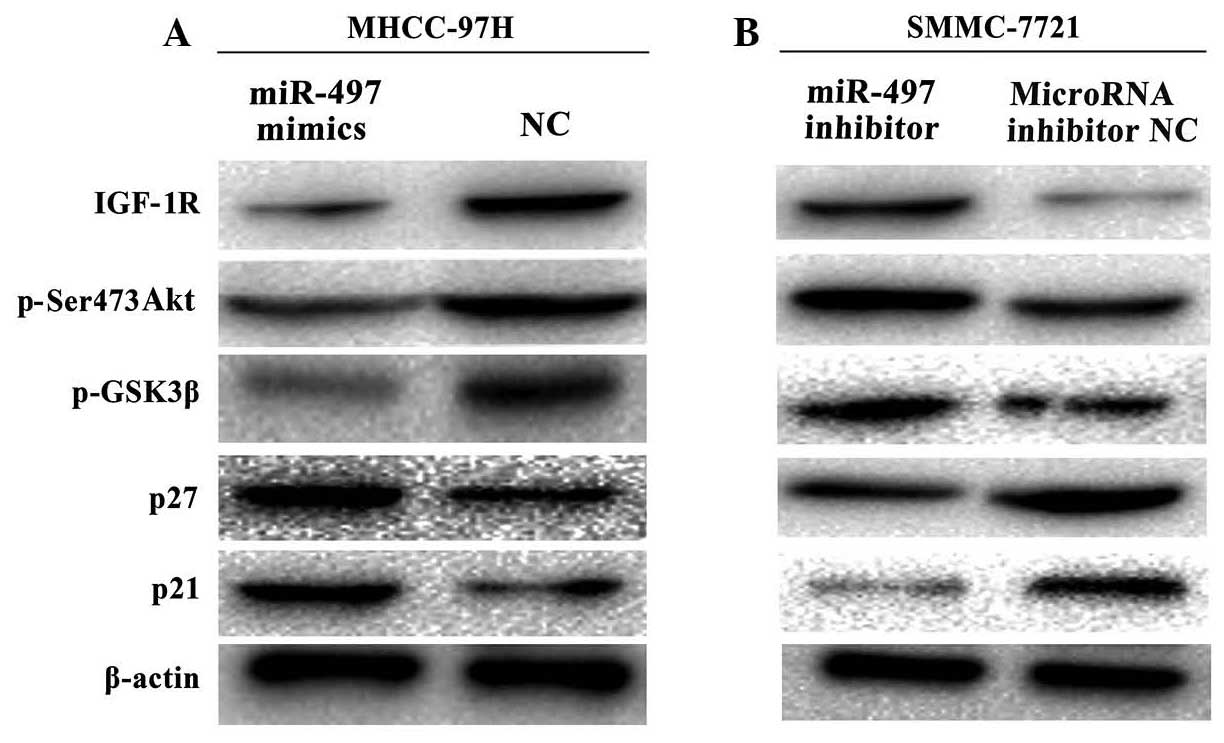
miR-497 expression affects PI3K/Akt
signaling and the expression of cyclin pathway-related
proteins
To determine the mechanism(s) by which miR-497
regulates tumor growth and progression, potential miR-497-regulated
molecules were examined in HCC cell lines with upregulated or
downregulated miR-497 expression. Protein expression data revealed
upregulation of p21 and p27, and downregulation of IGF-1R, p-Ser473
Akt and p-GSK3β in miR-497-overexpressing MHCC-97H cells (Fig. 4A). Conversely, miR-497 silencing by
the miR-497 inhibitor in SMMC-7721 cells led to downregulation of
p21 and p27, and upregulation of IGF-1R, p-Ser-473 Akt and p-GSK3β
(Fig. 4B).
Discussion
HCC is the fifth most frequent cancer worldwide
(40). A series of risk factors
contribute to HCC occurrence, including infection with hepatitis B
and C viruses, cirrhosis, chemical exposure, radiation and type 2
diabetes (41,42). Because surgical resection is only
suitable for patients diagnosed with early stage disease, systemic
chemotherapy remains an indispensable treatment option despite
serious adverse reactions (43).
Recently, increasing evidence suggests that miRNAs are involved in
hepatocarcinogenesis, thus opening new avenues of investigation
into the underlying molecular mechanism(s) of HCC, as well as
providing potential new therapeutic targets (44).
The current study focused on miRNA-497, which was
previously demonstrated to exhibit decreased expression in numerous
tumor types, and which may function as a tumor suppressor (12–20). The
results confirmed that the expression levels of miRNA-497 are
decreased in HCC tumor tissues or HCC-derived cell lines compared
with adjacent non-cancerous tissues or normal human L02
hepatocytes. Previously, Furuta et al (21) reported that miR-497 suppressed cell
growth by targeting multiple cell-cycle regulators in HCC (21). These results were consistent with
those of the current in vitro experiments in which exogenous
overexpression of miRNA-497 was observed to inhibit MHCC-97H colony
formation and tumor growth.
IGF-1R, a target gene of miR-497, was also selected
for investigation; overexpression of this gene has been reported in
human cervical cancer and colorectal cancer (12,20).
Furthermore, several online databases, including miRanda
(http://www.microrna.org/microrna/getMrna.do?gene=3480&utr=31402&organism=9606&matureName=hsa-miR-497)
and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/php/search.php?q=search_exact&searchword=hsa-miR-497-5p),
indicated that IGF-1R mRNA contains miR-497 binding sites. IGF-1R
belongs to the receptor tyrosine kinase family, and is activated by
insulin-like growth factor (IGF)-1 and IGF-2 with high affinities.
IGF-1R is a crucial component of the IGF axis that promotes cell
proliferation, migration, and transformation (45,46).
Previous studies have reported that IGF-1R expression is increased
in HCC and is closely associated with tumor progression (34,35). In
the current study, western blot and immunohistochemical analyses
revealed that IGF-1R was upregulated in HCC samples. As the
PI3K/Akt signaling pathway is often activated in HCC, and is
partially activated by IGF-1R (29,30,47),
western blot analyses were employed to investigate components of
this pathway. The results revealed that miR-497 overexpression
reduced IGF-1R expression and decreased PI3K/Akt pathway
activation. Changes were detected in downstream targets, including
decreased phosphorylation of Akt (Ser473) and GSK3β, and increased
expression of p27 and p21. Conversely, miR-497 silencing resulted
in increased IGF-1R expression and elevated activation of PI3K/Akt
signalling. These results indicate that miR-497 regulates PI3K/Akt
pathway activation, consistently with results from a previous study
on human colorectal cancer (12).
Notably, both miRNA-497 and p53 cluster at 17p13.1 (21), and wild-type p53 inhibits IGF-1R
expression, whereas mutant p53 increases IGF-1R expression
(48,49). These findings suggest that abnormal
transcription or translation of the chromosome 17p13.1 fragment may
play a crucial role in hepatocarcinogenesis.
In conclusion, the current results verify that
miRNA-497 downregulation occurs frequently during
hepatocarcinogenesis. miRNA-497 overexpression may suppress
cellular growth by targeting IGF-1R and inhibiting activation of
the PI3K/Akt pathway in HCC-derived cell lines. Currently, a series
of antibodies and small molecule inhibitors targeting the IGF axis
may provide an alternative strategy in the management of HCC
(50). Therefore, artificial
upregulation of miRNA-497 may also provide a new therapeutic option
for HCC.
Acknowledgements
This study was supported by a grant from the Natural
Science Foundation of China (no. 81270483).
References
|
1
|
Bosch FX, Ribes J, Díaz M and Cléries R:
Primary liver cancer Worldwide incidence and trends.
Gastroenterology. 127((Suppl 1)): S5–S16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cabrera R and Nelson DR: Review article:
The management of hepatocellular carcinoma. Aliment Pharmacol Ther.
31:461–476. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Caldwell S and Park SH: The epidemiology
of hepatocellular cancer: From the perspectives of public health
problem to tumor biology. J Gastroenterol. 44(Suppl 19): 96–101.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bosch FX, Ribes J and Borràs J:
Epidemiology of primary liver cancer. Semin Liver Dis. 19:271–285.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tsuchiya N, Sawada Y, Endo I, et al:
Biomarkers for the early diagnosis of hepatocellular carcinoma.
World J Gastroenterol. 21:10573–10583. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bruix J and Sherman M: Practice Guidelines
Committee, American Association for the Study of Liver Diseases:
Management of hepatocellular carcinoma. Hepatology. 42:1208–1236.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics biogenesis,
mechanism, and function. Cell. 116:281–297. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity, microRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: T arget recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vasudevan S, Tong Y and Steitz JA:
Switching from repression to activation, microRNAs can up-regulate
translation. Science. 318:1931–1934. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo ST, Jiang CC, Wang GP, Li YP, Wang CY,
Guo XY, Yang RH, Feng Y, Wang FH, Tseng HY, et al: MicroRNA-497
targets insulin-like growth factor 1 receptor and has a tumour
suppressive role in human colorectal cancer. Oncogene.
32:1910–1920. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo Q, Li X, Gao Y, Long Y, Chen L, Huang
Y and Fang L: MiRNA-497 regulates cell growth and invasion by
targeting cyclin E1 in breast cancer. Cancer Cell Int. 13:952013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Poell JB, van Haastert RJ, de Gunst T,
Schultz IJ, Gommans WM, Verheul M, Cerisoli F, van Noort PI,
Prevost GP, Schaapveld RQ, et al: A functional screen identifies
specific microRNAs capable of inhibiting human melanoma cell
viability. PLoS One. 7:e435692012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu W, Zhu D, Lu S, Wang T, Wang J, Jiang
B, Shu Y and Liu P: miR-497 modulates multidrug resistance of human
cancer cell lines by targeting BCL2. Med Oncol. 29:384–391. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen L, Li J, Xu L, Ma J, Li H, Xiao X,
Zhao J and Fang L: miR-497 induces apoptosis of breast cancer cells
by targeting Bcl-w. Exp Ther Med. 3:475–480. 2012.PubMed/NCBI
|
|
17
|
Lehmann U, Streichert T, Otto B, Albat C,
Hasemeier B, Christgen H, Schipper E, Hille U, Kreipe HH and Länger
F: Identification of differentially expressed microRNAs in human
male breast cancer. BMC Cancer. 10:1092010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li D, Zhao Y, Liu C, Chen X, Qi Y, Jiang
Y, Zou C, Zhang X, Liu S, Wang X, et al: Analysis of MiR-195 and
MiR-497 expression, regulation and role in breast cancer. Clin
Cancer Res. 17:1722–1730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Özata DM, Caramuta S, Velázquez-Fernández
D, Akçakaya P, Xie H, Höög A, Zedenius J, Bäckdahl M, Larsson C and
Lui WO: The role of microRNA deregulation in the pathogenesis of
adrenocortical carcinoma. Endocr Relat Cancer. 18:643–655. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luo M, Shen D, Zhou X, Chen X and Wang W:
MicroRNA-497 is a potential prognostic marker in human cervical
cancer and functions as a tumor suppressor by targeting the
insulin-like growth factor 1 receptor. Surgery. 153:836–847. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Furuta M, Kozaki K, Tanimoto K, Tanaka S,
Arii S, Shimamura T, Niida A, Miyano S and Inazawa J: The
tumor-suppressive miR-497-195 cluster targets multiple cell-cycle
regulators in hepatocellular carcinoma. PLoS One. 8:e601552013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
LeRoith D and Helman L: The new kid on the
block(ade) of the IGF-1 receptor. Cancer Cell. 5:201–202. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pollak M: Insulin and insulin-like growth
factor signalling in neoplasia. Nat Rev Cancer. 8:915–928. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Morrione A, DeAngelis T and Baserga R:
Failure of the bovine papillomavirus to transform mouse embryo
fibroblasts with a targeted disruption of the insulin-like growth
factor I receptor genes. J Virol. 69:5300–5303. 1995.PubMed/NCBI
|
|
25
|
Valentinis B and Baserga R: IGF-I receptor
signalling in transformation and differentiation. Mol Pathol.
54:133–137. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hellawell GO, Turner GD, Davies DR,
Poulsom R, Brewster SF and Macaulay VM: Expression of the type 1
insulin-like growth factor receptor is up-regulated in primary
prostate cancer and commonly persists in metastatic disease. Cancer
Res. 62:2942–2950. 2002.PubMed/NCBI
|
|
27
|
Tomizawa M, Shinozaki F, Sugiyama T,
Yamamoto S, Sueishi M and Yoshida T: Insulin-like growth factor-I
receptor in proliferation and motility of pancreatic cancer. World
J Gastroenterol. 16:1854–1858. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kornprat P, Rehak P, Rüschoff J and
Langner C: Expression of IGF-I, IGF-II, and IGF-IR in gallbladder
carcinoma. Histopathology. J Clin Pathol. 59:202–206. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Weber MM, Fottner C, Liu SB, Jung MC,
Engelhardt D and Baretton GB: Overexpression of the insulin-like
growth factor I receptor in human colon carcinomas. Cancer.
95:2086–2095. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sekharam M, Zhao H, Sun M, Fang Q, Zhang
Q, Yuan Z, Dan HC, Boulware D, Cheng JQ and Coppola D: Insulin-like
growth factor 1 receptor enhances invasion and induces resistance
to apoptosis of colon cancer cells through the Akt/Bcl-x(L)
pathway. Cancer Res. 63:7708–7716. 2003.PubMed/NCBI
|
|
31
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Markman B, Atzori F, Pérez-García J,
Tabernero J and Baselga J: Status of PI3K inhibition and biomarker
development in cancer therapeutics. Ann Oncol. 21:683–691. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Scharf JG, Schmidt-Sandte W, Pahernik SA,
Ramadori G, Braulke T and Hartmann H: Characterization of the
insulin-like growth factor axis in a human hepatoma cell line
(PLC). Carcinogenesis. 19:2121–2128. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang YC, Wang XP, Zhang LY, Song AL, Kou
ZM and Li XS: Effect of blocking IGF-I receptor on growth of human
hepatocellular carcinoma cells. World J Gastroenterol.
12:3977–3982. 2006.PubMed/NCBI
|
|
36
|
Llovet JM, Brú C and Bruix J: Prognosis of
hepatocellular carcinoma, The BCLC staging classification. Semin
Liver Dis. 19:329–338. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
National Research Council (US) Committee
for the Update of the Guide for the Care Use of Laboratory Animals
Guide for the Care and Use of Laboratory Animals (8th). Washington,
WA: National Academies Press. 2011.
|
|
39
|
Ni QF, Tian Y, Kong LL, Lu YT, Ding WZ and
Kong LB: Latexin exhibits tumor suppressor potential in
hepatocellular carcinoma. Oncol Rep. 31:1364–1372. 2014.PubMed/NCBI
|
|
40
|
Anwar SL and Lehmann U: MicroRNAs:
Emerging novel clinical biomarkers for hepatocellular carcinomas. J
Clin Med. 4:1631–1650. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Llovet JM and Bruix J: Molecular targeted
therapies in hepatocellular carcinoma. Hepatology. 48:1312–1327.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vigneri P, Frasca F, Sciacca L, Pandini G
and Vigneri R: Diabetes and cancer. Endocr Relat Cancer.
16:1103–1123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Laca L, Dedinska I, Miklusica J, Janik J,
Palkoci B and Pindura M: Surgical treatment of hepatocellular
carcinoma. Bratisl Lek Listy. 116:539–541. 2015.PubMed/NCBI
|
|
44
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics monitoring and therapeutics.
Histopathology. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pollak MN, Schernhammer ES and Hankinson
SE: Insulin-like growth factors and neoplasia. Nat Rev Cancer.
4:505–518. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Khandwala HM, McCutcheon IE, Flyvbjerg A
and Friend KE: The effects of insulin-like growth factors on
tumorigenesis and neoplastic growth. Endocr Rev. 21:215–244. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xu X, Sakon M, Nagano H, Hiraoka N,
Yamamoto H, Hayashi N, Dono K, Nakamori S, Umeshita K, Ito Y, et
al: Akt2 expression correlates with prognosis of human
hepatocellular carcinoma. Oncol Rep. 11:25–32. 2004.PubMed/NCBI
|
|
48
|
LeRoith D and Roberts CT Jr: The
insulin-like growth factor system and cancer. Cancer Lett.
195:127–137. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sjögren K, Liu JL, Blad K, Skrtic S, Vidal
O, Wallenius V, LeRoith D, Törnell J, Isaksson OG, Jansson JO, et
al: Liver-derived insulin-like growth factor I (IGF-I) is the
principal source of IGF-I in blood but is not required for
postnatal body growth in mice. Proc Natl Acad Sci USA.
96:7088–7092. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wu J and Zhu AX: Targeting insulin-like
growth factor axis in hepatocellular carcinoma. J Hematol Oncol.
4:302011. View Article : Google Scholar : PubMed/NCBI
|