Introduction
Ovarian carcinoma is one of the most common types of
cancer and is among the leading causes of mortality from
gynecological malignancies in the world (1). It is the fifth leading cause of
cancer-related mortality among females in Western countries
(2). No nationwide statistical data
on ovarian cancer have been reported in China as yet. It was
estimated that there were 19,300 new cases of ovarian cancers per
year in China during the 1980s, and an upward trend in incidence
has been observed in recent years (3). However, the molecular determinants of
ovarian tumorigenesis are still under investigation, while it is
conceivable that ovarian cancer, like most other cancers, arises as
a result of acquired alterations in gene expression due to specific
signal transduction pathways (4).
The transcriptional factor FOXC1, also known as
FREAC3 and FKHL7 (5,6), is a member of the forkhead box
(FOX)/winged-helix transcription factor family, which consists of
almost 100 members, with orthologs expressed in an array of species
ranging from yeast to humans (7).
These transcriptional factors share a highly conserved DNA-binding
forkhead domain (FHD) of 110 amino acids, consisting of three
α-helices and two large loops that form ‘wing’ structures. It is
through the FHD that FOX proteins are able to interact with DNA.
The function of FOX genes has become better understood in recent
years. FOX is a group of critical transcription factors which
control a variety of processes, including regulation of
embryogenesis and maintenance of differentiated cell states, major
organ systems and tissues from all three germ layers in the brain,
cardiovascular system, lung and kidney (8–11).
Mutation studies in mice, zebrafish and Drosophila have
revealed a diverse range of key roles of these genes during
embryonic development, including node formation and
anterior-posterior and left-right axis patterning (12–14). In
addition to their vital roles in normal development processes, a
number of FOX genes also participate in tumorigenesis (12).
The FOXC1 transcript has been detected in multiple
human organs using northern blot analysis (5,6,15). It is located on human chromosome 6p25
(6), encoding a 553 amino acid
protein (16,17). The FOXC1 coding sequence contains no
introns and comprises a 1659-bp open reading frame that contains
the FHD. The molecular weight of the FOXC1 protein is 56,789
Da.
Studies of animal models have demonstrated the
significance of FOXC1 as a key transcription factor in development.
Recombinant FOXC1 null mice die peri- or postnatally with massive
skeletal, cardiac, ocular and urogenital anomalies (18–20).
Mutations of the FOXC1 gene in humans result in various
glaucoma-related phenotypes, including Axenfeld-Rieger anomaly and
cardiac valve abnormalities (5,6,21,22). A
previous study also revealed that FOXC1 might be involved in
several types of genital tumorigenesis, including human prostate,
endometrial and ovarian cancers (23,24). The
FOXC1 gene may exert a negative regulation of cell proliferation in
several gynecological cancer cell lines (24).
However, it is unclear whether the FOXC1 protein
exists in ovarian tumor cell lines and tissues. In addition, the
clinical significance and molecular mechanism of the FOXC1 protein
in ovarian carcinoma remain poorly understood. This study was
designed to clarify the issue and explore the association of FOXC1
protein expression with clinicopathological factors and outcome of
the disease.
Materials and methods
Cell culture
Two human serous ovarian cystadenocarcinoma cell
lines were used in the present study: SKOV-3 was a gift from the
Ultrasound Institute of Chongqing Medical University, China, and
HO-8910 was purchased from Nanjing KeyGen Biotechnology Company,
China. SKOV-3 and HO-8910 cells were maintained in RPMI-1640 medium
(Gibco-Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal
calf serum (Gibco-Invitrogen) and 1% penicillin/streptomycin at
37°C in the presence of 5% CO2.
Tissue specimens
Twenty-five samples of serous ovarian cystoadenoma,
15 samples of ovarian borderline serous cystoadenoma and 40 samples
of serous ovarian cystadenocarcinoma, all paraffin-embedded, were
retrieved from case files at the Department of Pathology of the
Second Affiliated Hospital of Chongqing Medical University, China,
between February 2004 and February 2009. The diagnosis of serous
ovarian tumors was based on typical light microscopic findings. The
study was approved by the ethics committee of the Second Affiliated
Hospital of Chongqing Medical University.
Reverse transcription-quantitative
polymerase chain reaction RT-qPCR)
Total RNA was isolated from cultured SKOV-3 and
HO-8910 cells with TRIzol reagent (Takara Bio Inc., Otsu, Japan).
Total RNA (500 ng) was used as a template for RT using an RNA RT
kit from Takara Bio Inc. The RT reaction was set up in a 10 µl
mixture containing 2 µl 5X PrimeScript buffer (for qPCR), 0.5 µl
PrimeScript RT enzyme mix, 0.5 µl random 6 mers (100 µM) and 0.5 µl
oligo-dT primer (50 µM). Incubation was performed in an ABI 9700
DNA thermal cycler (Applied Biosystems, Foster City, CA, USA) for
15 min at 37°C, followed by 5 sec at 85°C. qPCR was performed on an
ABI 7300 real-time fluorescence quantitative PCR thermocycler
(Applied Biosystems) using an SYBR PrimeScript real-time PCR kit
(Takara Bio Inc.). Thermal cycler conditions involved holds for 10
sec at 94°C, followed by 40 cycles of 5 sec at 94°C and 30 sec at
60°C. The relative amount of mRNA was calculated using the
comparative threshold cycle method. The housekeeping gene β-actin
served as an internal parameter. The amplification efficiencies of
the target and reference were demonstrated to be approximately
equal with a slope of log input amount to threshold cycle <0.1.
The following oligonucleotide primers were used: FOXC1 forward
5′-AGCATCCGCCACAACCTC-3′, reverse 5′-GCCTGTCCTTCTCCTCCTT-3′; and
β-actin forward 5′-TGGCACCCAGCACAATGAA-3′, reverse
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′. Primers were designed using
Primer3 software, available from the Primer3 v. 0.4.0 website
(http://frodo.wi.mit.edu/primer3/,
Whitehead Institute for Biomedical Research, Cambridge, MA,
USA).
Western blot analysis
SKOV-3 and HO-8910 cells in the exponential phase of
growth were pooled and centrifuged at 250 × g at 4°C for 10 min,
and the packed cell volume was estimated. The deposition was
incubated on ice for 30 min in 200 µl lysis buffer A (10 mmol/l
HEPES, pH=7.9, 10 mmol/l KCl, 1.5 mmol/l MgCl2, 0.1
mmol/l ethylenedinitrolotetraacetic acid, 0.05% Nonidet P-40, 1
mmol/l dithiothreitol, 1 mmol/l phenylmethyl sulfonyl fluoride, 1
mg/l leupeptin), and centrifuged at 250 × g at 4°C for 10 min. The
supernatant from this spin was used as the cytoplastic extract. The
precipitum was then incubated on ice for 30 min in 100 µl lysis
buffer B (20 mmol/l HEPES, pH=7.9, 420 mmol/l NaCl, 1.5 mmol/l
MgCl2, 0.2 mmol/l ethylenedinitrolotetraacetic acid, 1
mmol/l dithiothreitol and phenylmethyl sulfonylfluoride, 1 µg/ml
leupeptin) before centrifuge at 14,000 × g at 4°C for 15 min. The
supernatant was the nuclear extract. Identical amounts of protein
from the extract were denatured and then subjected to
electrophoresis on a 5% stacking and 12% separating sodium dodecyl
sulfate (SDS) polyacrylamide gel using a Mini PROTEAN apparatus
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Electrophoretic
transfer to nitrocellulose was accomplished at 80 V for 2 h in 25
mmol/l Tris, pH 8.3, 192 mmol/l glycine, 0.1% SDS and 20% methanol.
The membrane was then blocked with 5% skimmed milk overnight at
4°C, followed by two 5-min washes in phosphate-buffered saline/1%
Tween-20 (PBST). The membrane was incubated with primary FOXC1
antibody (goat anti-human polyclonal antibody, sc-21396, Santa Cruz
Biotechnology Inc., Santa Cruz, CA, USA; dilution 1:200) at room
temperature for 2 h in antibody dilution buffer (1% bovine serum
albumin in PBST) followed by three 5-min washes in PBST. Secondary
antibody (donkey anti-goat antibody, Santa Cruz Biotechnology,
Inc.; dilution 1:5,000) was diluted in antibody dilution buffer,
and then added to the membrane for 2 h at room temperature,
followed by three 5-min washes in PBST. Finally, detection
procedures were performed using an enhanced chemiluminescence
western blotting detection kit (Beyotime Institute of
Biotechnology, Inc.).
Immunohistochemistry
The tissue samples were fixed by immersion in 10%
buffered formalin and subsequently embedded in paraffin according
to standard protocols. Sections for immunohistochemistry were cut
at 5 µm, mounted on Superfrost Plus glass slides (Thermo Fisher
Scientific, Inc., Waltham, MA, USA), baked for 30 min at 70°C and
left to dry overnight at 37°C. Subsequently, the sections were
deparaffinized and rehydrated by passing through xylene and a
graded series of ethanol solutions. Before the primary antibody was
applied, antigen retrieval was performed by boiling the sections in
10 mmol/l sodium citrate buffer (pH 6.0) at 98°C in a microwave
oven at 750 W for a total of 30 min (three cycles of 10 min each).
Slides were left to cool down to room temperature in the antigen
retrieval solution for 30 min. To block endogenous peroxidase
activity, the sections were incubated with 3% hydrogen peroxide in
methanol for 10 min. After blocking for 10 min in rabbit serum, the
sections were incubated with a specific primary antibody that
recognized FOXC1 (goat polyclonal antibody, sc-21396, Santa Cruz
Biotechnology Inc.; dilution 1:200) at 4°C overnight. Then, the
standard streptavidin-biotin complex immunoperoxidase technique
(goat streptavidin-peroxidase kit, Maixin Biotechnology Co.,
Fuzhou, China) was used according to the manufacturer's
instructions. All the sections were counterstained with
hematoxylin. Known positive internal controls (normal endometrium
tissues) (24) and negative controls
(sections in which the primary antibody was substituted with PBS)
were also stained in each run. Staining was assessed without any
knowledge of the clinical data by two observers.
SKOV-3 cells and HO-8910 cells were harvested in the
exponential phase of growth and were seeded into six-well plates
containing sterile coverslips at a concentration of
1×106 cells per well. After 48 h, cells were washed with
PBS and fixed with 4% paraformaldehyde at room temperature for 30
min. After rinsing in washing buffer, the cells were incubated with
3% hydrogen peroxide in methanol for 10 min and permeabilized using
0.05% Triton X-100/PBS for 10 min. The following steps were
performed in the same way as for immunohistochemistry of tumor
tissue.
Statistical analysis
The correlations between FOXCI expression and
clinicopathological parameters were examined by the Chi-square test
or Fisher's exact test. Statistical calculations were performed
using SAS software for Windows (Microsoft, Redmond, WA, USA).
P≤0.05 was considered to indicate a statistically significant
difference.
Results
Identification of FOXC1 expression in
serous ovarian carcinoma cell lines
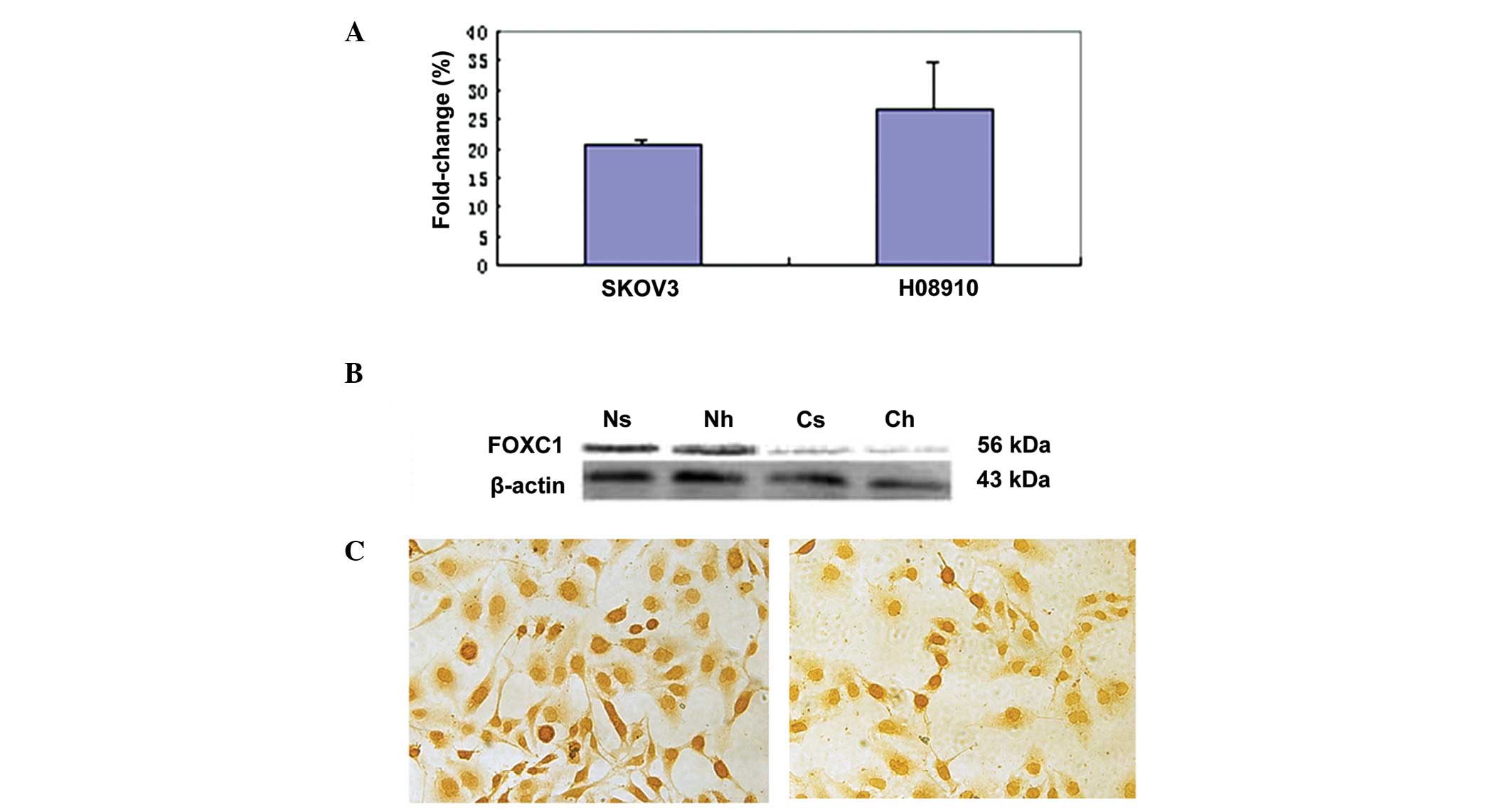
To study FOXC1 gene expression in ovarian tumors,
serous ovarian carcinoma cell lines HO-8910 and SKOV-3 were
selected. RT-qPCR was employed to evaluate FOXC1 mRNA expression in
the two cell lines. Western blot analysis and immunohistochemistry
with the anti-FOXC1 antibody were performed to confirm FOXC1
protein expression. There was no significant difference in FOXC1
mRNA and protein levels between the two cell lines (Fig. 1A–C). A single immunoreactive band of
56 kDa was detected, corresponding to the molecular weight
predicted for FOXC1, and no non-specific bands were detected with
this antibody. However, FOXC1 immunohistochemistry assay revealed
dichotomous immunoreactivity in the nucleus and cytoplasm of the
two cell lines. FOXC1 was localized mainly in the nucleus of SKOV-3
and HO-8910 cells, while less staining was observed in the
cytoplasm, which was in accordance with the results of western blot
analysis.
FOXC1 protein expression in serous
ovarian tumors and its clinicopathological significance
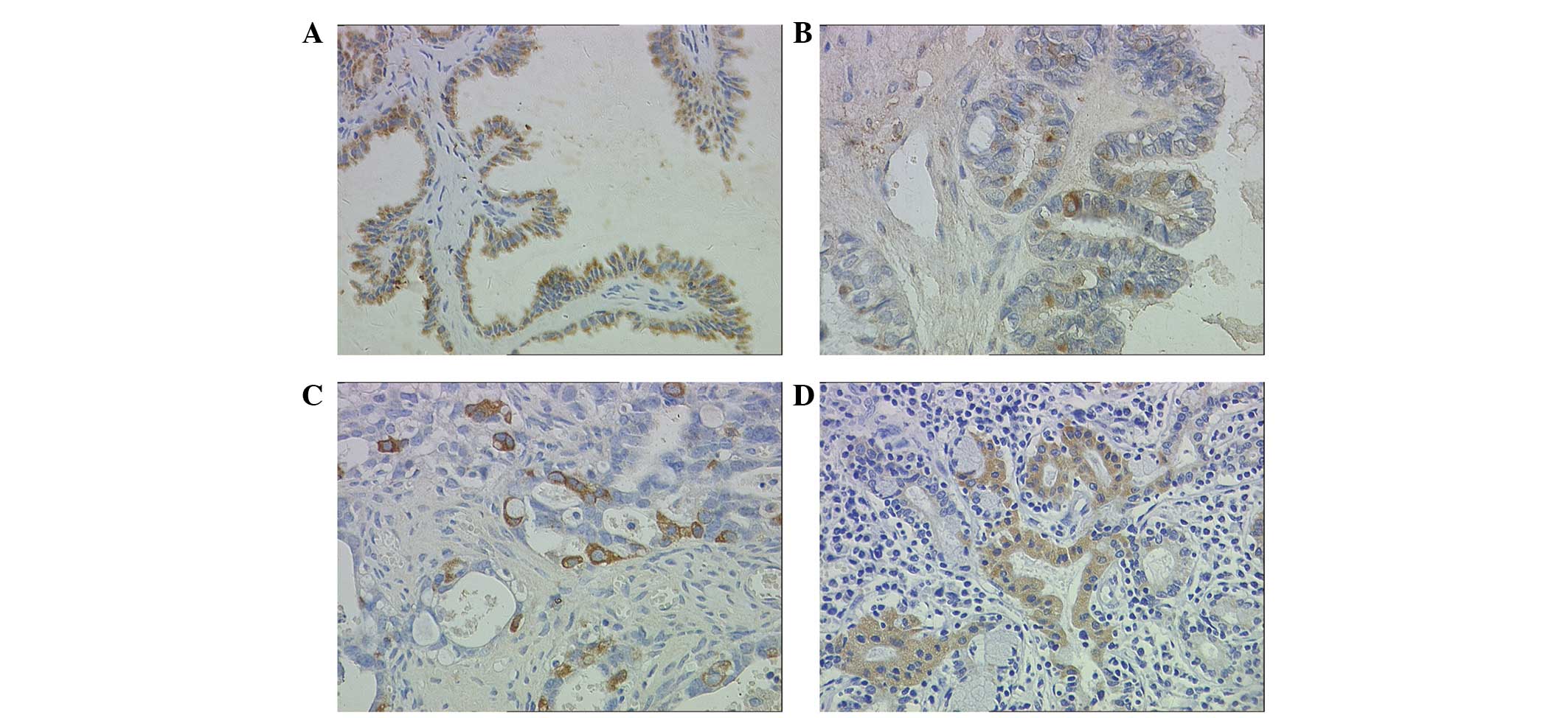
Staining patterns on immunohistochemistry were
consistent within each tissue. A sample was considered to
demonstrate positive immunoreactivity if >30% of the cells
exhibited intracellular staining following standard immunostaining.
Intercellular structure and inflammatory cells did not express
FOXC1 (Fig. 2A–D). In contrast with
the observation in cell lines (Fig.
1C), no nuclear FOXC1 immunoreactivity was detected in any of
serous ovarian tumor tissues. Notably, positive immunostaining for
FOXC1 was observed exclusively in the cytoplasm in 46 (57.5%) cases
of serous ovarian tumors.
Correlations between the incidence of cytoplasmic
FOXC1 immunoreactivity and clinicopathological features of serous
ovarian tumors are shown in Table I.
A retrospective investigation was conducted to clarify the
correlations of FOXC1 protein with clinicopathological variables of
serous ovarian tumors. A total of 80 patients with a medium (±
standard deviation) age of 48 years (range, 25 to 74 years) were
included in this study. According to the World Health Organization
(1999) criteria, the 80 serous ovarian tumors were classified
histologically as benign (serous ovarian cystoadenoma), borderline
(ovarian borderline serous cystoadenoma or low malignant potential
tumor) or malignant (serous ovarian cystadenocarcinoma) (25). Serous ovarian cystadenocarcinomas were
graded as well, moderately or poorly differentiated. Patients were
assigned a clinical stage in accordance with the International
Federation of Gynecology and Obstetrics (FIGO) 2000 standards
(26). Among the 80 cases of serous
ovarian tumor, 25 were benign, 15 borderline and 40 malignant.
Among the 40 cases of serous ovarian cystadenocarcinoma, 7 were
well differentiated, 21 moderately differentiated and 12 poorly
differentiated. Twelve cases were in stage I–II and 28 were in
stage III–IV. The volume of ascites in cystadenocarcinoma was
defined as small (≤500 ml), medium (500–2,000 ml) or large
(>2,000 ml). There were 7 cases with no ascites, 12 with a small
amount, 8 with a medium amount and 13 with a large amount. None of
the serous ovarian cystadenocarcinoma patients had received
preoperative chemotherapy or radiotherapy. FOXC1 positive
reactivity was observed in 21 (84%) cases of serous ovarian
cystoadenoma, 10 (66.7%) serous ovarian borderline cystoadenomas
and 15 (37.5%) serous ovarian cystadenocarcinomas. In malignant
tumors, FOXC1-positive immunohistochemistry was observed in 4
well-differentiated cases (57.1%), 9 moderately-differentiated
cases (42.9%) and 2 poorly-differentiated cases (16.7%) based on
the histological criteria used. According to the FIGO criteria, 9
cases were in stage I and II (75%) and 6 were in stage III and IV
(25%). Four cases were observed with no ascites (57.1%), 6 with a
small amount (50%), 3 with a medium amount (37.5%) and 2 with a
large amount (15.4%). A significant correlation between positive
FOXC1 immunoreactivity and pathological subtypes of serous ovarian
tumor was identified, and FOXC1 protein significantly decreased
with advancing FIGO stage (P<0.01). No significant association
was revealed between FOXC1 protein expression and the
clinicopathological factors of age, histological grade and volume
of ascites (P>0.05).
 | Table I.Correlation of FOXC1 protein with
clinicopathological features in serous ovarian tumors. |
Table I.
Correlation of FOXC1 protein with
clinicopathological features in serous ovarian tumors.
|
|
| Immunohistochemical
results [n (%)] |
|
|---|
|
|
|
|
|
|---|
| Variables | No. of cases | (+) | (−) | P-value |
|---|
| Age
groupa |
|
|
| 0.1500 |
|
<48 | 38 | 25 (65.8) | 13 (34.2) |
|
| ≥48 | 40 | 20 (50.0) | 20 (50.0) |
|
| Pathological
subtype |
|
|
| <0.0010 |
|
Benign | 25 | 21 (84.0) | 4
(16.0) |
|
|
Borderline | 15 | 10 (66.7) | 5
(33.3) |
|
|
Malignant | 40 | 15 (37.5) | 25 (62.5) |
|
| Histological grade of
cystadenocarcinoma |
|
|
| 0.1600 |
|
Well-differentiated | 7 | 4
(57.1) | 3
(42.9) |
|
|
Moderately differentiated | 21 | 9
(42.9) | 12 (57.1) |
|
| Poorly
differentiated | 12 | 2
(16.7) | 10 (83.3) |
|
| FIGO stages of
cystadenocarcinoma |
|
|
| 0.0035 |
| I–II | 12 | 9
(75.0) | 3
(25.0) |
|
|
III–IV | 28 | 6
(21.4) | 22 (78.6) |
|
| Volume of
cystadenocarcinoma ascites |
|
|
| 0.1900 |
| None | 7 | 4
(57.1) | 3
(42.9) |
|
| Small
amount | 12 | 6
(50.0) | 6
(50.0) |
|
| Moderate
amount | 8 | 3
(37.5) | 5
(62.5) |
|
| Large
amount | 13 | 2
(15.4) | 11 (84.6) |
|
Discussion
To our knowledge, the present study is the first to
conduct a FOXC1 immunohistochemical examination in serous ovarian
tumors. In the study, endogenous expression of FOXC1 mRNA and
protein was confirmed in two serous ovarian cystadenocarcinoma cell
lines SKOV-3 and HO-8910 with RT-qPCR and western blot analysis.
The subcellular location of FOXC1 protein was immunohistochemically
detected in the two cell lines and in 57.5% (46/80 cases) of serous
ovarian tumors. FOXC1 protein was present in 84% of serous ovarian
cystadenomas and 66.7% of borderline cystadenomas, whereas its
expression was observed in only 37.5% of adenocarcinomas. Our
retrospective study demonstrated that positive immunostaining for
FOXC1 protein significantly decreased with advancing FIGO stage
(I–II vs. III–IV) as well as pathological subtypes from benign to
borderline and malignant tumors (Table
I).
As demonstrated in the study, the differential FOXC1
staining in these three major categories of serous ovarian tumors
may provide evidence for a different pathogenetic basis for benign,
borderline and malignant serous ovarian tumors. The observation
sparked our interest in the potential role of FOXC1 gene in ovarian
tumorigenesis and the correlation between FOXC1 protein and
clinicopathological characteristics. Notably, exclusively
cytoplasmic immunostaining for FOXC1 protein was observed in serous
ovarian tumor tissues and normal endometrial tissues used as
positive controls, whereas both nuclear and cytoplasmic
localization of FOXC1 protein was observed in the two cell lines,
with expression levels in the nucleus markedly higher than those in
the cytoplasm (Fig. 1C). This result
was in discordance with previous studies reporting that the
wild-type full-length FOXC1 protein was localized exclusively in
the nucleus in almost all cells, due to two independent nuclear
localization signals within FOXC1 FHD (27–29). FOXC1
has been identified as a phosphoprotein, and the activity of the
transcriptional inhibitory domain located at amino acids 215–366
may be regulated by phosphorylation (30). In previous studies, FOXO1 (FKHRL1)
protein, which is also a FOX family member, phosphorylated by AKT,
was observed to be located in the cytoplasm, whereas
unphosphorylated FOXO1 protein was observed in the nucleus and
acted as a transcription factor for various genes, including,
presumably, FAS ligand (tumor necrosis factor superfamily member 6)
(31). However, it is unclear whether
there are similar mechanisms of FOXC1 protein in ovarian tumors.
One plausible explanation for this phenomenon may be
phosphorylation, which accounts for the protein shuttling between
the nucleus and cytoplasm in the two cell lines and tumor tissues.
FOXC1 expression in the cytoplasm may act as a tumor suppressor
when unphosphorylated.
The present study has limitations. Phosphorylated
FOXC1 antibody is no longer available commercially. Therefore it is
now impossible for us to verify whether FOXC1 protein in the
nucleus is phosphorylated or not. Fifteen out of 40 cases of serous
ovarian cystadenocarcinoma (37.5%) revealed FOXC1-positive
expression, which was correlated with poorer histological grade and
increased volume of ascites. However, no significant associations
were observed between FOXC1 protein expression and
clinicopathological factors including histological grades and
volume of ascites. A possible explanation might be that the total
number of malignant cases was not large enough to analyze clinical
significance. For further studies, a larger number of malignant
tumor cases should be recruited to better understand the trend
between FOXC1 protein expression and the clinicopathological
factors of histological grade and volume of ascites. In addition,
the mechanism of subcellular mislocation of FOXC1 protein should be
investigated to determine whether these interactions are regulated
through phosphorylation.
In conclusion, high expression of FOXC1 protein may
be correlated with a benign pathological subtype of serous ovarian
tumors and a trend towards good prognosis. The molecular mechanism
underlying FOXC1 protein in ovarian tumorigenesis remains to be
explored.
Acknowledgements
The authors thank Professors Liping Zhang, Xiaoqiu
Xiao and Xiaoni Zhong for their support, as well as Professors
Youde Cao and Ying Ma for their expertise and assistance in the
immunohistochemical analysis of serous ovarian tumors. The authors
also acknowledge Mr. Zhengyi Wang in the Institute of Life Science
of Chongqing Medical University.
Financial support was provided by the Natural
Science Fund Programmes of Chongqing Science and Technology
Committee and Scientific Research Projects of Chongqing Health
Bureau (CSTC. 2006BB5279).
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Landis SH, Murray T, Bolden S and Wingo
PA: Cancer statistics, 1999. CA Cancer J Clin. 49:8–31. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu A: Epidemiological analysis of
gynecological malignant carcinoma. Bulletin of Chinese Cancer.
6:3–5. 1997.(In Chinese).
|
|
4
|
Weinberg RA: Tumor suppressor genes.
Science. 254:1138–1146. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mears AJ, Jordan T, Mirzayans F, Dubois S,
Kume T, Parlee M, Ritch R, Koop B, Kuo WL, Collins C, et al:
Mutations of the forkhead/winged-helix gene, FKHL7, in patients
with Axenfeld-Rieger anomaly. Am J Hum Genet. 63:1316–1328. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nishimura DY, Swiderski RE, Alward WL,
Searby CC, Patil SR, Bennet SR, Kanis AB, Gastier JM, Stone EM and
Sheffield VC: The forkhead transcription factor gene FKHL7 is
responsible for glaucoma phenotypes which map to 6p25. Nat Genet.
19:140–147. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lai E, Clark KL, Burley SK and Darnell JE
Jr: Hepatocyte nuclear factor 3/fork head or ‘winged helix’
proteins. A family of transcription factors of diverse biologic
function. Proc Natl Acad Sci USA. 90:10421–10423. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xuan S, Baptista CA, Balas G, Tao W,
Soares VC and Lai E: Winged helix transcription factor BF-1 is
essential for the development of the cerebral hemispheres. Neuron.
14:1141–1152. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kume T, Jiang H, Topczewska JM and Hogan
BL: The murine winged helix transcription factors, Foxc1 and Foxc2,
are both required for cardiovascular development and somitogenesis.
Genes Dev. 15:2470–2482. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shu W, Yang H, Zhang L, Lu MM and Morrisey
EE: Characterization of a new subfamily of winged-helix/forkhead
(Fox) genes that are expressed in the lung and act as
transcriptional repressors. J Biol Chem. 276:27488–27497. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hatini V, Huh SO, Herzlinger D, Soares VC
and Lai E: Essential role of stromal mesenchyme in kidney
morphogenesis revealed by targeted disruption of Winged Helix
transcription factor BF-2. Genes Dev. 10:1467–1478. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lehmann OJ, Sowden JC, Carlsson P, Jordan
T and Bhattacharya SS: Fox's in development and disease. Trends
Genet. 19:339–344. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ang SL and Rossant J: HNF-3 beta is
essential for node and notochord formation in mouse development.
Cell. 78:561–574. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brody SL, Yan XH, Wuerffel MK, Song SK and
Shapiro SD: Ciliogenesis and left-right axis defects in forkhead
factor HFH-4-null mice. Am J Respir Cell Mol Biol. 23:45–51. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pierrou S, Hellqvist M, Samuelsson L,
Enerbäck S and Carlsson P: Cloning and characterization of seven
human forkhead proteins, binding site specificity and DNA bending.
Embo J. 13:5002–5012. 1994.PubMed/NCBI
|
|
16
|
Gould DB, Mears AJ, Pearce WG and Walter
MA: Autosomal dominant Axenfeld-Rieger anomaly maps to 6p25. Am J
Hum Genet. 61:765–768. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mirzayans F, Mears AJ, Guo SW, Pearce WG
and Walter MA: Identification of the human chromosomal region
containing the iridogoniodysgenesis anomaly locus by
genomic-mismatch scanning. Am J Hum Genet. 61:111–119. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kume T, Deng KY, Winfrey V, Gould DB,
Walter MA and Hogan BL: The forkhead/winged helix gene Mf1 is
disrupted in the pleiotropic mouse mutation congenital
hydrocephalus. Cell. 93:985–996. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Winnier GE, Kume T, Deng K, Rogers R,
Bundy J, Raines C, Walter MA, Hogan BL and Conway SJ: Roles for the
winged helix transcription factors MF1 and MFH1 in cardiovascular
development revealed by nonallelic noncomplementation of null
alleles. Dev Biol. 213:418–431. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Carlsson P and Mahlapuu M: Forkhead
transcription factors: Key players in development and metabolism.
Dev Biol. 250:1–23. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Swiderski RE, Reiter RS, Nishimura DY,
Alward WL, Kalenak JW, Searby CS, Stone EM, Sheffield VC and Lin
JJ: Expression of the Mf1 gene in developing mouse hearts,
Implication in the development of human congenital heart defects.
Dev Dyn. 216:16–27. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lehmann OJ, Ebenezer ND, Jordan T, Fox M,
Ocaka L, Payne A, Leroy BP, Clark BJ, Hitchings RA, Povey S, et al:
Chromosomal duplication involving the forkhead transcription factor
gene FOXC1 causes iris hypoplasia and glaucoma. Am J Hum Genet.
67:1129–1135. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
van der Heul-Nieuwenhuijsen L, Dits NF and
Jenster G: Gene expression of forkhead transcription factors in the
normal and diseased human prostate. Bju Int. 103:1574–1580. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou Y, Kato H, Asanoma K, Kondo H, Arima
T, Kato K, Matsuda T and Wake N: Identification of FOXC1 as a
TGF-beta1 responsive gene and its involvement in negative
regulation of cell growth. Genomics. 80:465–472. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kaku T, Ogawa S, Kawano Y, Ohishi Y,
Kobayashi H, Hirakawa T and Nakano H: Histological classification
of ovarian cancer. Med Electron Microsc. 36:9–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Benedet JL, Bender H, Jones H III, Ngan HY
and Pecorelli S: FIGO staging classifications and clinical practice
guidelines in the management of gynecologic cancers. FIGO Committee
on Gynecologic Oncology. Int J Gynaecol Obstet. 70:209–262. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saleem RA, Banerjee-Basu S, Berry FB,
Baxevanis AD and Walter MA: Analyses of the effects that
disease-causing missense mutations have on the structure and
function of the winged-helix protein FOXC1. Am J Hum Genet.
68:627–641. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saleem RA, Banerjee-Basu S, Murphy TC,
Baxevanis A and Walter MA: Essential structural and functional
determinants within the forkhead domain of FOXC1. Nucleic Acids
Res. 32:4182–4193. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saleem RA, Banerjee-Basu S, Berry FB,
Baxevanis AD and Walter MA: Structural and functional analyses of
disease-causing missense mutations in the forkhead domain of FOXC1.
Hum Mol Genet. 12:2993–3005. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Berry FB, Saleem RA and Walter MA: FOXC1
transcriptional regulation is mediated by N- and C-terminal
activation domains and contains a phosphorylated transcriptional
inhibitory domain. J Biol Chem. 277:10292–10297. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo
P, Hu LS, Anderson MJ, Arden KC, Blenis J and Greenberg ME: Akt
promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|