Introduction
Phthalates, including butyl benzyl phthalate (BBP),
di-n-butyl phthalate (DBP) and di-2-ethylhexyl phthalate
(DEHP), are utilized as softeners and plasticizers (1,2).
Epidemiological studies have observed that phthalate exposure may
increase the risk of breast cancer (3–6). However,
the effects of phthalate esters in the breast cancer tumor
microenvironment remain to be elucidated.
The tumor microenvironment is known to have a
significant role in the progression of tumors and the development
of chemoresistance to anticancer drugs (7). The tumor microenvironment is comprised
of stromal cells, immune cells [lymphocytes, macrophages and
dendritic cells (DCs)], growth factors, extracellular matrix
constituents, metabolites and cytokines/chemokines (8). As antigen-presenting cells, DCs have
been observed to exhibit significant roles in the initiation and
regulation of the immune response to cancer (9). Tumor-associated DCs (TADCs) have been
observed to contribute to the metastasis of tumors in various
cancers (10,11). Regulated upon activation, normal
T-cell expressed and secreted (RANTES), also known as C-C chemokine
ligand 5, is a cytokine consistently observed in increased levels
in breast cancer subtypes (12), and
has been observed to be associated with the progression of breast
cancer and the promotion of metastasis (13–16).
Epidemiological studies have provided evidence that
a high dietary intake of flavonoids via fruits and vegetables may
be associated with reduced cancer rates in humans (17–20).
Flavonoids are a class of phenolic compounds that are widely
distributed throughout the plant kingdom; they display diverse
biological activities, including the inhibition of tumor
progression and the prevention of cancer initiation (21,22).
Didymin, a dietary flavonoid glycoside present in citrus fruits,
demonstrates antioxidant and anticancer properties (23–28).
The present study evaluated the effects of phthalate
esters in the breast cancer tumor microenvironment and investigated
didymin, a dietary flavonoid glycoside present in citrus fruits, as
a possible antidote for phthalate ester-associated cancer
aggravation.
Materials and methods
Chemicals
Didymin was obtained from Extrasynthese (Genay,
France), and was dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich, St. Louis, MO, USA) and stored at −20°C. Control
cultures received the carrier solvent (0.1% DMSO). All other
chemicals utilized were in the purest form available
commercially.
Cell culture and conditioned
medium
Human breast adenocarcinoma MDA-MB-231 cells
(American Type Culture Collection, Manassas, VA, USA) were cultured
in α-minimum essential medium (α-MEM; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with non-essential amino
acids, 0.1 mmol/l sodium pyruvate, 1% antibiotic/anti-mycotic
solution and 10% fetal bovine serum (FBS) (all Thermo Fisher
Scientific, Inc.). In order to obtain the various conditioned media
(CM), the MDA-MB-231 cells (2×106/100-mm dish) were
treated with or without BBP, DBP or DEHP (all Sigma-Aldrich) at
identical concentrations of 1 µM for 6 h. Following washing and
culturing for 24 h, the CM of phthalate ester-treated MDA-MB-231
cells (BBP-, DBP- or DEHP-MDA-CM) were harvested (Fig. 1A).
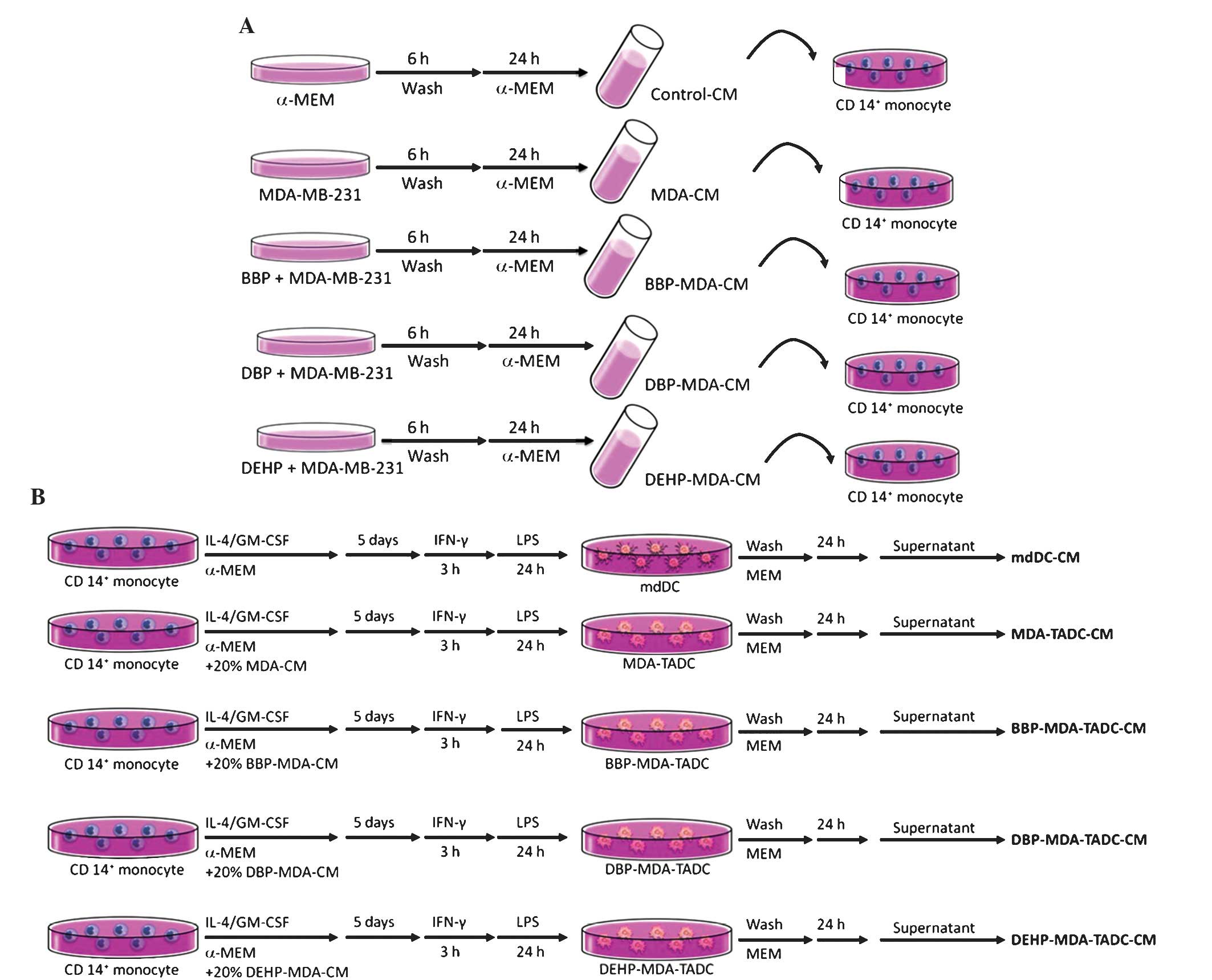 | Figure 1.Flow chart of the production of
various CM. (A) Flow chart of the production of control-CM, MDA-CM,
BBP-MDA-CM, DBP-MDA-CM and DEHP-MDA-CM. (B) Flow chart of the
production of mdDC-CM, MDA-TADC-CM, BBP-MDA-TADC-CM,
DBP-MDA-TADC-CM and DEHP-MDA-TADC-CM. CM, conditioned media; MDA,
MDA-MB-231 cells; BBP, butyl benzyl phthalate; DBP,
di-n-butyl phthalate; DEHP, di-2-ethylhexyl phthalate; mdDC,
monocyte-derived dendritic cells; TADC, tumor-associated mdDC; MEM,
minimum essential medium; IL, interleukin; GM-CSF,
granulocyte-macrophage colony-stimulating factor; IFN, interferon;
LPS, lipopolysaccharide; CD, cluster of differentiation. |
Isolation of cluster of
differentiation (CD)14+ monocytes and differentiation of
monocyte-derived dendritic cells (mdDCs)
Monocytes were purified from peripheral blood
mononuclear cells obtained from healthy consenting donors.
Mononuclear cells were isolated from the blood by Ficoll-Hypaque
gradient (GE Healthcare Life Sciences, Chalfont, UK).
CD14+ monocytes were purified with CD14+
monoclonal antibody-conjugated magnetic beads (MACS MicroBeads;
Miltenyi Biotec GmbH, Bergisch Gladbach, Germany), according to the
manufacturer's protocols. mdDCs were generated by culturing
CD14+ monocytes in α-MEM containing FBS and 20 ng/ml
granulocyte-macrophage colony-stimulating factor (GM-CSF) and 10
ng/ml interleukin (IL)4 (R&D Systems, Inc., Minneapolis, MN,
USA) for 5 days. The medium was replaced with fresh medium
containing GM-CSF and IL4 on day 3. For the maturation of DCs,
immature mdDCs were stimulated with lipopolysaccharide (100 ng/ml;
Sigma-Aldrich) following priming with interferon-γ (EMD Millipore,
Billerica, MA, USA) for 3 h. MDA-MB-231 tumor-associated mdDCs
(MDA-TADCs), BBP-MDA-TADCs, DBP-MDA-TADCs or DEHP-MDA-TADCs were
generated by culturing CD14+ monocytes in α-MEM medium
containing FBS, IL4 and GM-CSF in 20% MDA-CM, BBP-MDA-CM,
DBP-MDA-CM or DEHP-MDA-CM, and subsequently stimulated as
aforementioned. Following washing, the supernatants were collected
and identified as MDA-MB-231-TADC-CM, BBP-MDA-TADC-CM,
DBP-MDA-TADC-CM or DEHP-MDA-TADC-CM (Fig.
1B). The Institutional Review Board (IRB) of Kaohsiung Medical
University Hospital (Kaohsiung, Taiwan) approved the present study
protocol and all participants provided written informed consent in
accordance with the Declaration of Helsinki (IRB numbers:
KMUH-IRB-990174 and KMUH-IRB-20120362).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was utilized for RNA isolation, while
complementary (c)DNA was prepared using an oligo(dT) primer and
reverse transcriptase (Takara Bio, Inc., Otsu, Japan) following
standard protocols (29). RT-qPCR was
performed using SYBR Green on the ABI 7500 Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Each PCR
mixture contained 200 nM of each primer, 10 µl of 2X SYBR Green PCR
Master Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.),
and 5 µl of cDNA and RNase-free water, in a total volume of 20 µl.
The RT-qPCR was performed with a denaturation step at 95°C for 10
min, then for 40 cycles at 95°C for 15 sec and 60°C for 1 min. All
PCRs were performed in triplicate and normalized to internal
control glyceraldehyde-3-phosphate dehydrogenase mRNA. The relative
expression level was presented using the 2−ΔΔCq method.
The primer sequences of target genes in the present study were as
follows: RANTES, F 5′-cgc tgt cat cct cat tgc ta-3′ and R 5′-aca
cac ttg gcg gtt ctt tc-3′; and GAPDH F 5′-GAGTCAACGGATTTGGTCGT-3′
and R 5′-TTGATTTTGGAGGGATCTCG-3′.
Enzyme-linked immunosorbent assay
(ELISA)
RANTES levels were determined using an ELISA-based
kit (R&D Systems Europe Ltd., Abingdon, UK). ELISA was
performed according to the manufacturer's protocols. Depletion of
RANTES from various CMs was performed using a mouse monoclonal
anti-RANTES antibodies (2 µg/ml; Abcam, Cmabridge, UK) and
Sepharose™ Protein A/G beads (PAG50-00-0002, Rockland
Immunochemicals Inc., Gilbertsville, PA, USA) following standard
immunoprecipitation techniques (30).
Cytokine depletion was additionally confirmed using an ELISA-based
kit.
Cell proliferation
The cells were plated in 96-well culture plates.
Following 24 h of incubation, the cells were treated with vehicle
mdDC-CM or specific CM for 72 h. At the conclusion of the assay
period, cell proliferation was measured using a water-soluble
tetrazolium salts (WST)-1 assay. Cell proliferation was determined
using Premixed WST-1 Cell Proliferation Reagent (Clontech
Laboratories, Inc., Mountainview, CA, USA) in accordance with the
manufacturer's protocols.
Cell migration and invasion assay
Cell migration and invasion assays were performed
using QCM™ 24-well Cell Migration and Invasion Assay kits (EMD
Millipore). Briefly, the cells were seeded into the migration
chamber and mdDC-CM or various CM were added to the lower wells for
24 h as a chemoattractant. At the conclusion of the treatment, the
cells were stained using CyQuant GR dye (cat no. 90131) in cell
lysis buffer for 15 min at room temperature. The fluorescence of
the migrated and invaded cells was subsequently measured using a
fluorescence plate reader at excitation/emission wavelengths of
485/520 nm.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistical comparisons of the results were performed using an
analysis of variance. Significant differences between the means of
the test groups were analyzed using Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Breast cancer cells, following
exposure to phthalate esters, affect mdDCs and contribute to breast
cancer progression by enhancing cancer cell proliferation,
migration and invasion
In order to understand whether phthalate esters
exacerbate cancer progression in the breast cancer tumor
microenvironment, the effects of BBP-MDA-TADC-CM, DBP-MDA-TADC-CM
and DEHP-MDA-TADC-CM on breast cancer cell proliferation, migration
and invasion were investigated. MDA-TADC-CM (20%) increased breast
cancer cell proliferation (P=0.01), and this stimulatory effect was
additionally enhanced when MDA-MB-231 cells were pretreated with
BBP, DBP or DEHP (Fig. 2A) (P=0.001,
0.0001 and 0.0007 for BBP, DBP and DEHP, respectively). In
addition, MDA-TADC-CM (20%) induced breast cancer cell migration
and invasion (P=0.002 and 0.008 for migration and invasion
analysis), and this reinforceable effect was worsened when breast
cancer cells were cultured with BBP-MDA-TADC-CM (20%),
DBP-MDA-TADC-CM (20%) or DEHP-MDA-TADC-CM (20%) (Fig. 2B and C).
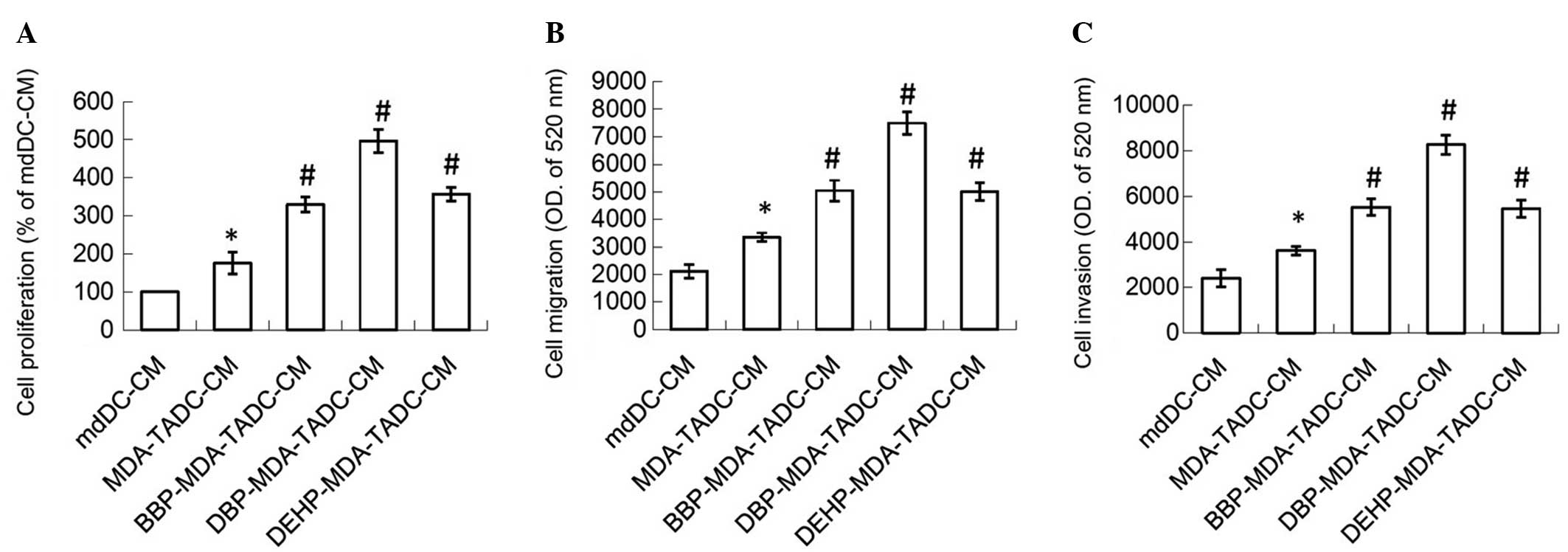 | Figure 2.CM of phthalate ester-treated
MDA-MB-231 cells cause mdDCs to increase breast cancer cell
proliferation, migration and invasion. BBP-, DBP- and DEHP-TADC-CM
increased breast cancer cell (A) proliferation, (B) migration and
(C) invasion. Each value is presented as the mean ± standard
deviation of three independent experiments. *P<0.05 vs. mdDC-CM
treatment. #P<0.05 vs. MDA-TADC-CM treatment. CM,
conditioned media; mdDCs, monocyte-derived dendritic cells; BBP,
butyl benzyl phthalate; DBP, di-n-butyl phthalate; DEHP,
di-2-ethylhexyl phthalate; TADC, tumor-associated mdDC; MEM,
minimum essential medium; OD, optical density. |
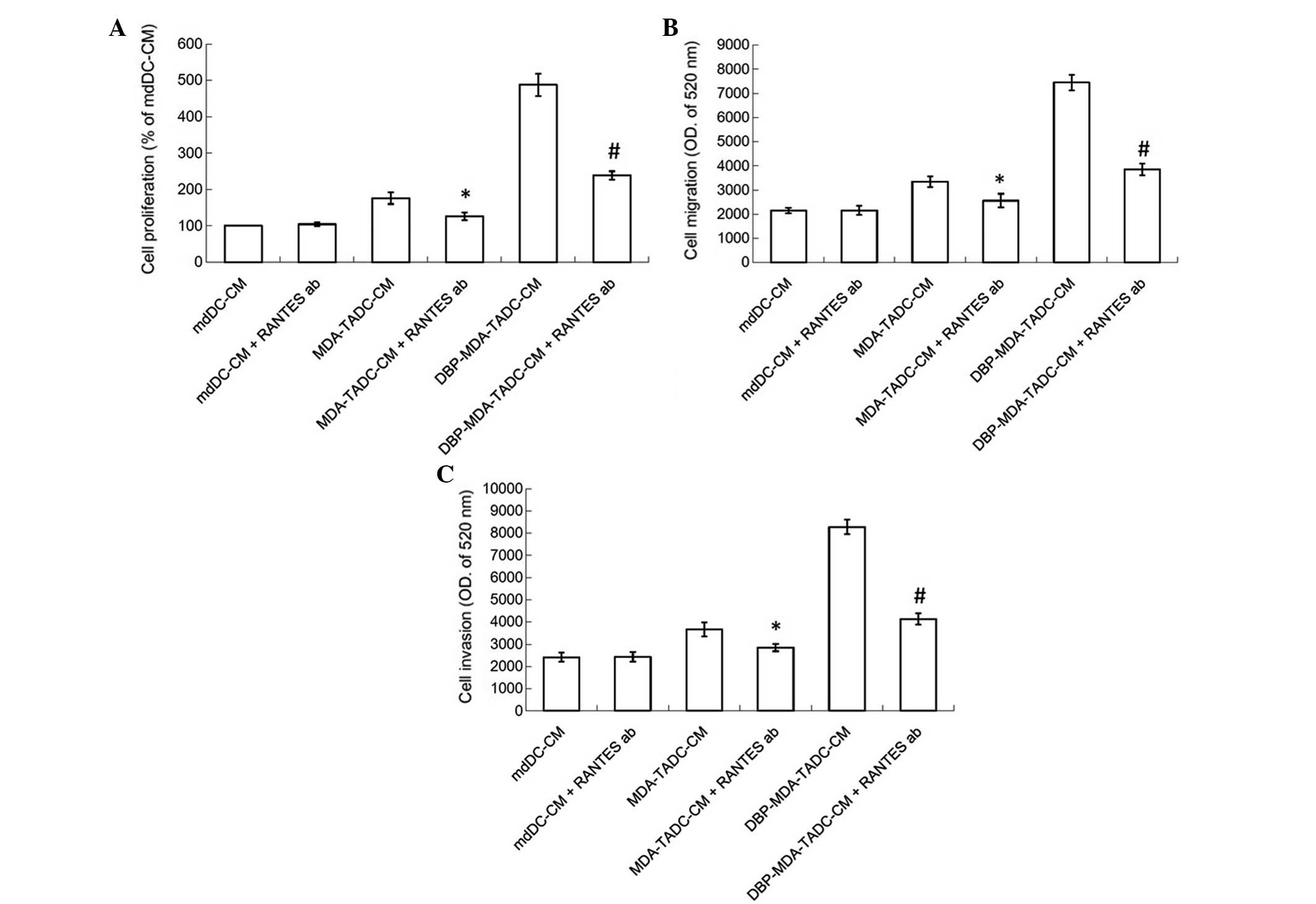
RANTES has a significant role in
TADC-mediated cancer progression
In order to determine the primary factors
contributing to MDA-TADC, BBP-MDA-TADC, DBP-MDA-TADC and
DEHP-MDA-TADC-mediated breast cancer progression, RT-qPCR analysis
revealed that RANTES mRNA levels were increased by 6-, 12-, 17- and
11-fold in MDA-TADCs, BBP-MDA-TADCs, DBP-MDA-TADCs and
DEHP-MDA-TADCs, respectively (Fig.
3A). As demonstrated by ELISA, the protein levels of RANTES
were enhanced in MDA-TADC-CM (P=0.01), BBP-MDA-TADC-CM (P=0.005),
DBP-MDA-TADC-CM (P=0.002) and DEHP-MDA-TADC-CM (Fig. 3B) (P=0.003). The effect of
DBP-MDA-TADC-CM on the induction of breast cancer cell
proliferation, migration and invasion, and the induction of RANTES
was greater than that of BBP-MDA-TADC-CM or DEHP-MDA-TADC-CM
(Figs. 2 and 3). As DBP demonstrated the greatest effect
in these circumstances, it was selected as the model for
investigation of the detailed effects of phthalate ester-associated
cancer aggravation in the breast cancer tumor microenvironment. In
order to understand its role, RANTES was depleted in MDA-TADC-CM
and DBP-MDA-TADC-CM. The effect of MDA-TADC-CM and DBP-MDA-TADC-CM
on cancer progression was subsequently assessed. The successful
depletion of RANTES from MDA-TADC-CM and DBP-MDA-TADC-CM was
confirmed using RANTES ELISA kits (data not shown). As demonstrated
in Fig. 4, RANTES depletion inhibited
the stimulatory effects of MDA-TADC-CM on breast cancer cell
proliferation, migration and invasion (P=0.00002 for cell
proliferation, P<0.0001 for migration and invasion), as well as
blocking the intensified stimulatory effects of DBP-MDA-TADC-CM on
breast cancer cell proliferation, migration and invasion.
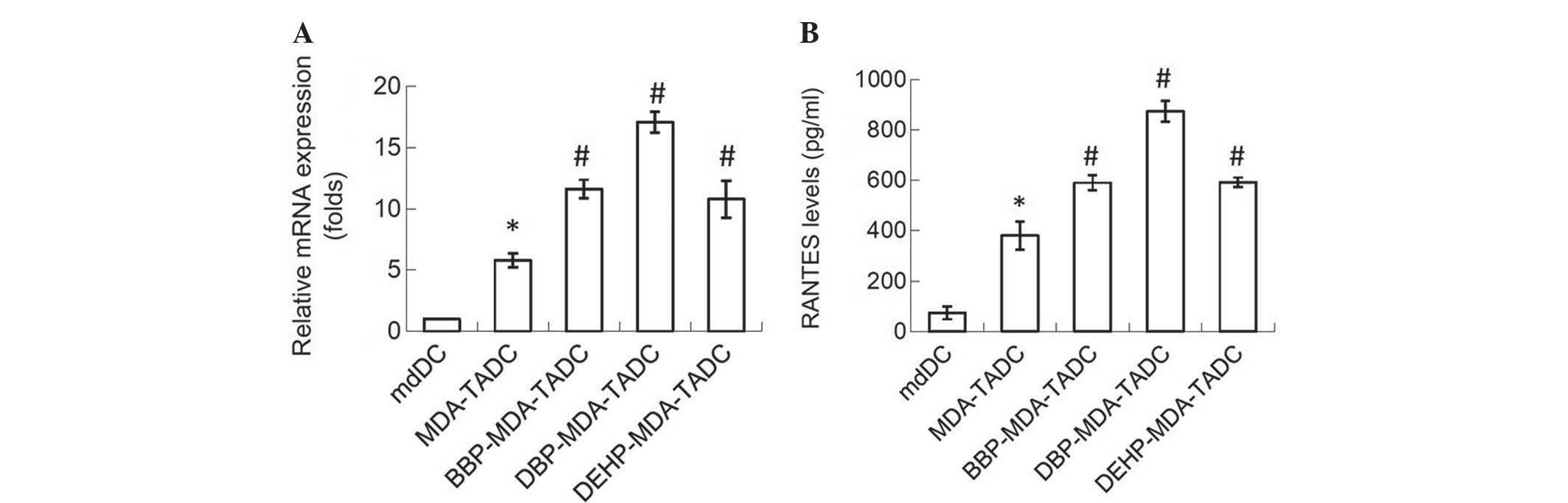 | Figure 3.MDA-CM, BBP-MDA-CM, DBP-MDA-CM and
DEHP-MDA-CM increase the expression of RANTES in mdDCs. (A) MDA-CM,
BBP-MDA-CM, DBP-MDA-CM and DEHP-MDA-CM increased the levels of
RANTES, as assessed by reverse transcription-polymerase chain
reaction. (B) MDA-CM, BBP-MDA-CM, DBP-MDA-CM and DEHP-MDA-CM
increased the RANTES protein levels, as detected by enzyme-linked
immunosorbent assay. Each value is presented as the mean ± standard
deviation of three independent experiments. *P<0.05 vs. mdDC
treatment. #P<0.05 vs. MDA-TADC treatment. MDA,
MDA-MB-231 cells; CM, conditioned media; BBP, butyl benzyl
phthalate; DBP, di-n-butyl phthalate; DEHP, di-2-ethylhexyl
phthalate; RANTES; regulated upon activation, normal T-cell
expressed, and secreted; mdDCs, monocyte-derived dendritic
cells. |
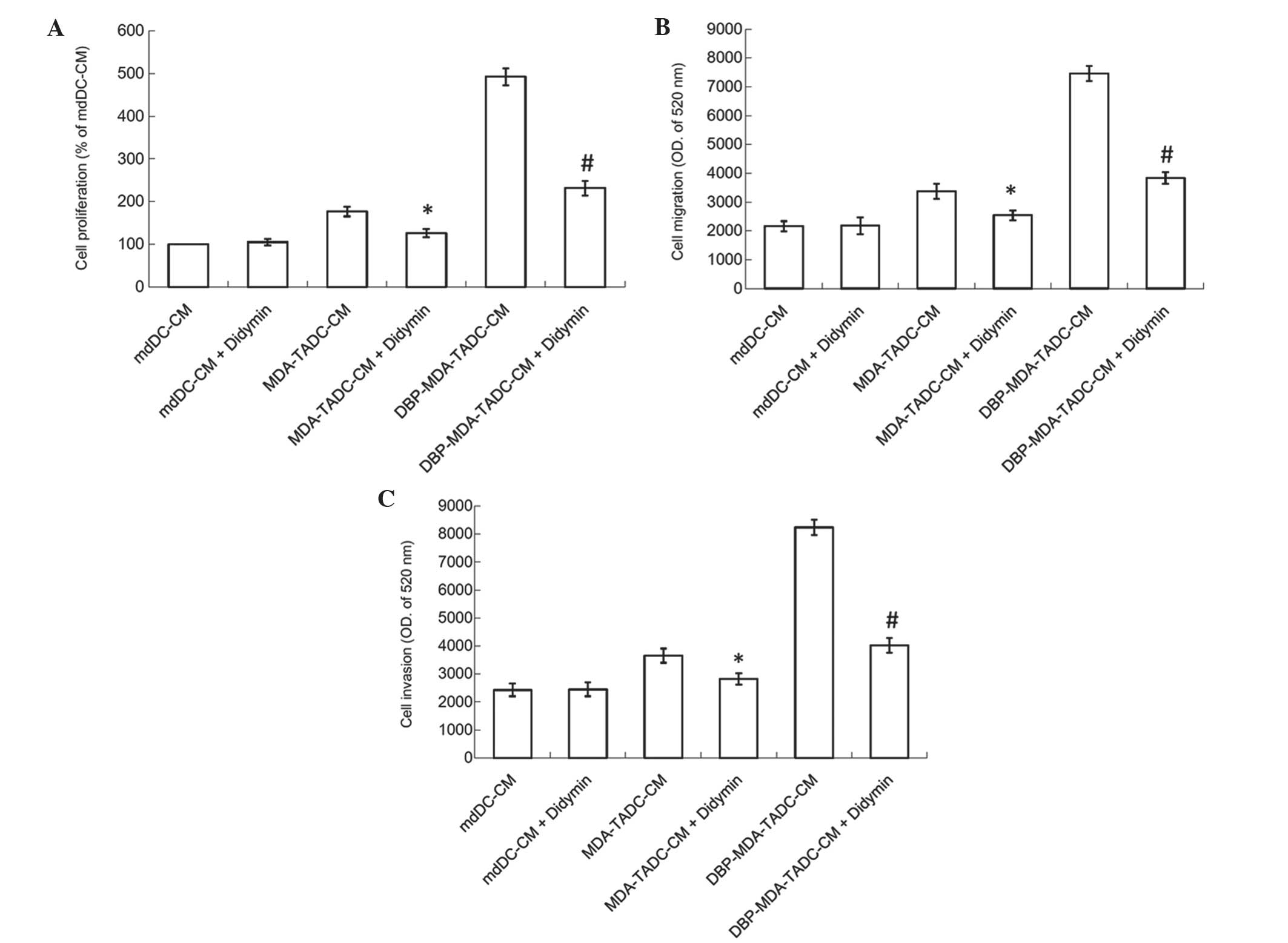
Didymin suppresses
DBP-MDA-TADC-mediated breast cancer aggravation
The tumor microenvironment has a significant role in
the development of chemoresistance to anticancer drugs and tumor
progression (7). TADC has been
demonstrated to promote the progression of cancer by modulating a
number of components in the cancer niche, thereby creating a
supportive and permissive microenvironment for tumor survival,
proliferation and metastasis (11,31). As
phthalate esters stimulate the ability of breast cancer cells to
affect mdDCs and thereby intensified breast cancer cell
proliferation, migration and invasion, the search for a possible
antidote in the fight against phthalate esters-induced cancer
aggravation in the breast cancer tumor microenvironment has become
a matter of importance. The present study therefore assessed the
effect of didymin, a dietary flavonoid glycoside derived from
citrus fruits, on DBP-induced cancer progression. As shown in
Fig. 5A, the MDA-TADC-CM-induced
breast cancer cell proliferation effect was inhibited when cells
were pretreated with didymin, and DBP-MDA-TADC-CM-induced breast
cancer cell proliferation was additionally reversed when the cells
were pretreated with didymin. Similarly, enhancement of breast
cancer cell migration and invasion triggered by DBP-MDA-TADC-CM was
abrogated upon didymin pretreatment (Fig.
5B and C).
Discussion
To the best of our knowledge, the present study is
the first to evaluate the interaction between mdDCs and breast
cancer cells following exposure to phthalate esters. BBP, DBP and
DEHP stimulated the breast cancer cells and subsequently caused the
mdDCs to secrete RANTES, which enhanced the proliferation,
migration and invasion of the human breast cancer cells. To the
best of our knowledge, the present study is additionally the first
to investigate the effects of didymin in interfering with phthalate
ester-mediated breast cancer aggravation in the breast cancer tumor
microenvironment. The results of the present study suggested that
didymin was capable of preventing phthalate ester-associated breast
cancer progression (Fig. 6).
Phthalates, including BBP, DBP and DEHP, are widely
utilized in food wraps and cosmetic products (1–3).
Individuals are exposed to phthalates throughout their entire lives
via ingestion, inhalation and dermal exposure (1,2). Several
phthalates have been demonstrated to promote breast cancer
development by increasing cell proliferation and migration
(32–34). In a previous study, we demonstrated
that phthalate esters were able to induce breast cancer bone
metastasis by targeting parathyroid hormone-related protein
(35). The present study demonstrated
that phthalate esters BBP, DBP and DEHP were able to induce breast
cancer cells to affect mdDCs, thereby intensifying breast cancer
cell proliferation, migration and invasion. The results of the
present study suggested that phthalate esters may increase cancer
progression in the breast cancer tumor microenvironment.
The tumor microenvironment has been observed to
affect cancer progression and generation. The cells surrounding a
tumor provide essential supportive factors that promote progression
(36–39). RANTES has been observed to be a
significant contributor to various chronic inflammatory diseases
and malignancies via the recruitment of inflammatory cells
(15,40,41).
RANTES has additionally been reported to be overexpressed in
certain cancers and is involved in critical steps of cancer spread,
including proliferation, migration, invasion, angiogenesis and
metastatic colonization following activation (42–45).
Furthermore, RANTES has been associated with resistance to
conventional chemotherapeutic drugs, including cisplatin and
tamoxifen (46). The present study
revealed that the phthalate esters BBP, DBP and DEHP were able to
cause breast cancer cells to affect mdDCs to produce the
inflammatory cytokine RANTES, which subsequently enhanced breast
cancer cell proliferation, migration and invasion. Depletion of
RANTES reversed the effects of MDA-TADC-CM- and
DBP-MDA-TADC-CM-mediated breast cancer cell proliferation,
migration and invasion. Elimination of all phthalate exposure may
not be possible, as phthalate esters are extensively used in modern
life (1,2). It is therefore important that strategies
are developed for the prevention of breast cancer progression in
the breast cancer tumor microenvironment. The results of the
present study revealed that didymin, a dietary flavonoid glycoside
obtained from citrus fruits, is able to reverse the negative
actions of DBP-stimulated breast cancer cells, which cause mdDCs to
enhance the proliferation, migration and invasion of human breast
cancer cells.
In conclusion, to the best of our knowledge, the
present study is the first to investigate the interaction between
mdDCs and breast cancer cells following exposure to phthalate
esters. These esters, BBP, DBP and DEHP, were used to stimulate
human breast cancer MDA-MB-231 cells to mediate RANTES upregulation
in mdDCs, thereby inducing breast cancer cell proliferation,
migration and invasion. To the best of our knowledge, this is the
first study to provide evidence that didymin, a dietary flavonoid
glycoside present in citrus fruits, has potential for the
prevention of phthalate ester-associated breast cancer progression
in the breast cancer tumor microenvironment. Therefore, the present
study suggested that didymin may be capable of preventing phthalate
ester-associated cancer aggravation.
Acknowledgements
The present study was supported by grants from the
National Science Council of Taiwan (nos. NSC
102-2628-B-037-002-MY3, NSC 102-2632-B-037-001-MY3, and NSC
102-2314-B-037-035-MY3), the Kaohsiung Medical University ‘Aim for
the Top 500 Universities Grant’ (no. KMU-DT103008), the Kaohsiung
Medical University ‘Aim for the Top Universities Grant’ (no.
KMU-TP103A15), the Kaohsiung Medical University Hospital Research
Foundation (no. KMUH103-3M53) and the Excellence for Cancer
Research Center Grant, the Ministry of Health and Welfare,
Executive Yuan (Taipei, Taiwan; no. MOHW 103-TD-B-111-05).
References
|
1
|
Martino-Andrade AJ and Chahoud I:
Reproductive toxicity of phthalate esters. Mol Nutr Food Res.
54:148–157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ge RS, Chen GR, Tanrikut C and Hardy MP:
Phthalate ester toxicity in Leydig cells, Developmental timing and
dosage considerations. Reprod Toxicol. 23:366–373. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
López-Carrillo L, Hernández-Ramírez RU,
Calafat AM, Torres-Sánchez L, Galván-Portillo M, Needham LL,
Ruiz-Ramos R and Cebrián ME: Exposure to phthalates and breast
cancer risk in northern Mexico. Environ Health Perspect.
118:539–544. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aschengrau A, Coogan PF, Quinn M and
Cashins LJ: Occupational exposure to estrogenic chemicals and the
occurrence of breast cancer, An exploratory analysis. Am J Ind Med.
34:6–14. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carran M and Shaw IC: New Zealand Malayan
war veterans' exposure to dibutylphthalate is associated with an
increased incidence of cryptorchidism hypospadias and breast cancer
in their children. N Z Med J. 125:52–63. 2012.PubMed/NCBI
|
|
6
|
Sprague BL, Trentham-Dietz A, Hedman CJ,
Wang J, Hemming JD, Hampton JM, Buist DS, Bowles Aiello EJ, Sisney
GS and Burnside ES: Circulating serum xenoestrogens and
mammographic breast density. Breast Cancer Res. 15:R452013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Katoh H, Wang D, Daikoku T, Sun H, Dey SK
and Dubois RN: CXCR2-expressing myeloid-derived suppressor cells
are essential to promote colitis-associated tumorigenesis. Cancer
Cell. 24:631–644. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Peltekova VD, Lemire M, Qazi AM, Zaidi SH,
Trinh QM, Bielecki R, Rogers M, Hodgson L, Wang M, D'Souza DJ, et
al: Identification of genes expressed by immune cells of the colon
that are regulated by colorectal cancer-associated variants. Int J
Cancer. 134:2330–2341. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gajewski TF, Schreiber H and Fu YX: Innate
and adaptive immune cells in the tumor microenvironment. Nat
Immunol. 14:1014–1022. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hsu YL, Huang MS, Cheng DE, Hung JY, Yang
CJ, Chou SH and Kuo PL: Lung tumor-associated dendritic
cell-derived amphiregulin increased cancer progression. J Immunol.
187:1733–1744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuo CH, Chen KF, Chou SH, Huang YF, Wu CY,
Cheng DE, Chen YW, Yang CJ, Hung JY and Huang MS: Lung
tumor-associated dendritic cell-derived resistin promoted cancer
progression by increasing Wolf-Hirschhorn syndrome candidate
1/Twist pathway. Carcinogenesis. 34:2600–2609. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gonzalez RM, Daly DS, Tan R, Marks JR and
Zangar RC: Plasma biomarker profiles differ depending on breast
cancer subtype but RANTES is consistently increased. Cancer
Epidemiol Biomarkers Prev. 20:1543–1551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Velasco-Velázquez M and Pestell RG: The
CCL5/CCR5 axis promotes metastasis in basal breast cancer.
Oncoimmunology. 2:e236602013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lv D, Zhang Y, Kim HJ, Zhang L and Ma X:
CCL5 as a potential immunotherapeutic target in triple-negative
breast cancer. Cell Mol Immunol. 10:303–310. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Lv D, Kim HJ, Kurt RA, Bu W, Li Y
and Ma X: A novel role of hematopoietic CCL5 in promoting
triple-negative mammary tumor progression by regulating generation
of myeloid-derived suppressor cells. Cell Res. 23:394–408. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ash SA, Valchev GI, Looney M, Mhathuna Ni
A, Crowley PD, Gallagher HC and Buggy DJ: Xenon decreases cell
migration and secretion of a pro-angiogenesis factor in breast
adenocarcinoma cells, Comparison with sevoflurane. Br J Anaesth.
113((Suppl 1)): i14–i21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takemura H, Sakakibara H, Yamazaki S and
Shimoi K: Breast cancer and flavonoids - a role in prevention. Curr
Pharm Des. 19:6125–6132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Donaldson MS: Nutrition and cancer: A
review of the evidence for an anti-cancer diet. Nutr J. 3:192004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stoner GD and Mukhtar H: Polyphenols as
cancer chemopreventive agents. J Cell Biochem Suppl. 22:169–180.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang CS, Landau JM, Huang MT and Newmark
HL: Inhibition of carcinogenesis by dietary polyphenolic compounds.
Annu Rev Nutr. 21:381–406. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aggarwal BB and Shishodia S: Molecular
targets of dietary agents for prevention and therapy of cancer.
Biochem Pharmacol. 71:1397–1421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ramos AM and Aller P: Quercetin decreases
intracellular GSH content and potentiates the apoptotic action of
the antileukemic drug arsenic trioxide in human leukemia cell
lines. Biochem Pharmacol. 75:1912–1923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Calabrò ML, Galtieri V, Cutroneo P,
Tommasini S, Ficarra P and Ficarra R: Study of the extraction
procedure by experimental design and validation of a LC method for
determination of flavonoids in Citrus bergamia juice. J
Pharm Biomed Anal. 35:349–363. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Finotti E: DiM ajo D: Influence of
solvents on the antioxidant property of flavonoids. Nahrung.
47:186–187. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mouly P, Gaydou EM and Auffray A:
Simultaneous separation of flavanone glycosides and
polymethoxylated flavones in citrus juices using liquid
chromatography. J Chromatogr A. 800:171–179. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ross SA, Ziska DS, Zhao K and ElSohly MA:
Variance of common flavonoids by brand of grapefruit juice.
Fitoterapia. 71:154–161. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hung JY, Hsu YL, Ko YC, Tsai YM, Yang CJ,
Huang MS and Kuo PL: Didymin, a dietary flavonoid glycoside from
citrus fruits, induces Fas-mediated apoptotic pathway in human
non-small-cell lung cancer cells in vitro and in
vivo. Lung Cancer. 68:366–374. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singhal J, Nagaprashantha LD and Vatsyayan
R: Ashutosh,A wasthi S and Singhal SS: Didymin induces apoptosis by
inhibiting N-Myc and upregulating RKIP in neuroblastoma. Cancer
Prev Res (Phila). 5:473–483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zeng F, Xie L, Pang X, Liu W, Nie Q and
Zhang X: Complementary eoxyribonucleic acid cloning of avian G0/G1
switch gene 2, and its expression and association with production
traits in chicken. Poult Sci. 90:1548–1554. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bharadwaj U, Li M, Zhang R, Chen C and Yao
Q: Elevated interleukin-6 and G-CSF in human pancreatic cancer cell
conditioned medium suppress dendritic cell differentiation and
activation. Cancer Res. 67:5479–5488. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Watkins SK, Zhu Z, Riboldi E,
Shafer-Weaver KA, Stagliano KE, Sklavos MM, Ambs S, Yagita H and
Hurwitz AA: FOXO3 programs tumor-associated DCs to become
tolerogenic in human and murine prostate cancer. J Clin Invest.
121:1361–1372. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hsieh TH, Tsai CF, Hsu CY, Kuo PL, Hsi E,
Suen JL, Hung CH, Lee JN, Chai CY, Wang SC and Tsai EM: n-Butyl
benzyl phthalate promotes breast cancer progression by inducing
expression of lymphoid enhancer factor 1. PLoS One. 7:e427502012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hsieh TH, Tsai CF, Hsu CY, Kuo PL, Lee JN,
Chai CY, Wang SC and Tsai EM: Phthalates induce proliferation and
invasiveness of estrogen receptor-negative breast cancer through
the AhR/HDAC6/c-Myc signaling pathway. FASEB J. 26:778–787. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen FP and Chien MH: Lower concentrations
of phthalates induce proliferation in human breast cancer cells.
Climacteric. 17:377–384. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hsu YL, Tsai EM, Hou MF, Wang TN, Hung JY
and Kuo PL: Obtusifolin suppresses phthalate esters-induced breast
cancer bone metastasis by targeting parathyroid hormone-related
protein. J Agric Food Chem. 62:11933–11940. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tsai MJ, Chang WA, Huang MS and Kuo PL:
Tumor microenvironment, A new treatment target for cancer. ISRN
Biochem. 2014:3519592014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Y, Han ZP, Zhang SS, Jing YY, Bu XX,
Wang CY, Sun K, Jiang GC, Zhao X, Li R, et al: Effects of
inflammatory factors on mesenchymal stem cells and their role in
the promotion of tumor angiogenesis in colon cancer. J Biol Chem.
286:25007–25015. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo YP, Zhou H, Krueger J, Kaplan C, Liao
D, Markowitz D, Liu C, Chen T, Chuang TH, Xiang R and Reisfeld RA:
The role of proto-oncogene Fra-1 in remodeling the tumor
microenvironment in support of breast tumor cell invasion and
progression. Oncogene. 29:662–673. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hsu YL, Hou MF, Kuo PL, Huang YF and Tsai
EM: Breast tumor-associated osteoblast-derived CXCL5 increases
cancer progression by ERK/MSK1/Elk-1/snail signaling pathway.
Oncogene. 32:4436–4447. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chang LY, Lin YC, Mahalingam J, Huang CT,
Chen TW, Kang CW, Peng HM, Chu YY, Chiang JM, Dutta A, et al:
Tumor-derived chemokine CCL5 enhances TGF-β-mediated killing of
CD8(+) T cells in colon cancer by T-regulatory cells. Cancer Res.
72:1092–1102. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Soria G, Ofri-Shahak M, Haas I,
Yaal-Hahoshen N, Leider-Trejo L, Leibovich-Rivkin T, Weitzenfeld P,
Meshel T, Shabtai E, Gutman M and Ben-Baruch A: Inflammatory
mediators in breast cancer. Coordinated expression of TNFα &
IL-1β with CCL2 & CCL5 and effects on epithelial-to-mesenchymal
transition. BMC Cancer. 11:1302011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Borczuk AC, Papanikolaou N, Toonkel RL,
Sole M, Gorenstein LA, Ginsburg ME, Sonett JR, Friedman RA and
Powell CA: Lung adenocarcinoma invasion in TGFbetaRII-deficient
cells is mediated by CCL5/RANTES. Oncogene. 27:557–564. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mi Z, Bhattacharya SD, Kim VM, Guo H,
Talbot LJ and Kuo PC: Osteopontin promotes CCL5-mesenchymal stromal
cell-mediated breast cancer metastasis. Carcinogenesis. 32:477–487.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Trellakis S, Bruderek K, Dumitru CA,
Gholaman H, Gu X, Bankfalvi A, Scherag A, Hütte J, Dominas N,
Lehnerdt GF, et al: Polymorphonuclear granulocytes in human head
and neck cancer: Enhanced inflammatory activity, modulation by
cancer cells and expansion in advanced disease. Int J Cancer.
129:2183–2193. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiao X, Katiyar S, Willmarth NE, Liu M, Ma
X, Flomenberg N, Lisanti MP and Pestell RG: c-Jun induces mammary
epithelial cellular invasion and breast cancer stem cell expansion.
J Biol Chem. 285:8218–8226. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yi EH, Lee CS, Lee JK, Lee YJ, Shin MK,
Cho CH, Kang KW, Lee JW, Han W, Noh DY, et al: STAT3-RANTES
autocrine signaling is essential for tamoxifen resistance in human
breast cancer cells. Mol Cancer Res. 11:31–42. 2013. View Article : Google Scholar : PubMed/NCBI
|