Introduction
Head and neck squamous cell carcinoma (HNSCC) is one
of the most prevalent types of human cancer, with ~500,000 novel
cases being diagnosed worldwide every year (1). HNSCC is the sixth leading cause of
cancer-associated mortality (2).
Although the molecular pathogenesis of HNSCC is not yet fully
understood, it has been reported that aberrant cell division is one
of the main mechanisms of carcinogenesis (3). Undergoing chromosome condensation,
nuclear envelope breakdown, separation, bipolar-spindle assemblage,
chromosome segregation and cytokinesis, the cells separate and
replicated genetic material is split into two daughter cells
(3). A complex regulatory network of
kinase and phosphatase regulate this process aiding the accuracy of
cell division.
There are three homologues of Aurora kinase (A, B
and C) (4). Aurora kinase A (AURKA)
has been implicated in numerous types of cancer, including colonic,
breast, liver, gastric, uterine, ovarian, non-small cell lung,
pancreatic and esophageal cancer (5–10).
Furthermore, a correlation has been observed between the
overexpression of AURKA mRNA and tumor progression and shortened
survival in patients with HNSCC (11). Despite AURKA mRNA and protein being
frequently overexpressed in various types of cancer, they are not
always correlated with the gene amplification (12). The overexpression of AURKA has also
been reported in gastric (13),
breast (14) and ovarian (15) cancer. Therefore, besides gene
amplification, other mechanisms, including transcriptional
activation (16), suppression of
protein degradation (17) and
activation of certain signaling pathways (18), may also serve essential roles.
Focal adhesion kinase (FAK), a non-receptor tyrosine
kinase, is a key regulator of cell proliferation, migration and
invasion, and is involved in the development and progression of
cancer. It has previously been established that FAK serves a vital
role in the mediation of signal transduction pathways, involved in
cell attachment, migration, invasion, proliferation and survival,
which are crucial for cancer development and metastasis (19,20).
Numerous studies have described increased expression of FAK protein
in a variety of human cancers, including sarcomas, astrocytomas,
and carcinomas of the breast, colon, thyroid, prostate, oral
cavity, liver, stomach and ovary (20). Furthermore, Canel et al
(21) demonstrated that FAK
expression may be employed as an effective index for cervical lymph
node metastases in patients with laryngeal squamous cell carcinoma.
Additionally, Akt may be activated as result of AURKA
overexpression. It has been demonstrated that an AURKA inhibitor
may overcome AURKA-induced chemoresistance in various types of
cancer (22).
The present study aimed to validate the hypothesis
that AURKA activates FAK through the AURKA/Akt/FAK signaling
pathway, and subsequently promotes the cell migration and invasion
of HNSCC cells. The current study may provide a basis for the
future development of inhibitors of the AURAK/Akt/FAK signaling
pathway, with the aim to alleviate and eventually treat HNSCCs.
Materials and methods
Cell lines and cell culture
Human HNSCC cell lines (FaDu and Hep2) were obtained
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China). The cells were maintained at 37°C with 5% CO2 in
Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS) and 1
µg/ml penicillin/streptomycin.
Furthermore, the FaDu and Hep2 cells were treated
with 75 nM of the AURKA inhibitor, VX-680 (Selleck Chemicals,
Houston, TX, USA), for 24 and 48 h (23), with 100 µM of the FAK inhibitor,
TAE226 (Selleck Chemicals), for 12 and 24 h (24), or with 5 µM of the Akt inhibitor,
triciribine (Selleck Chemicals), for 6 and 12 h (25), respectively. Subsequent to the
treatment of each inhibitor, for the aforementioned specific times,
the cells were harvested for the following experiments.
Transwell migration and invasion
assay
The cell migration capability was determined using
the previously described methods (26). A total of 600 µl medium containing 20%
FBS was added to the lower chamber, whilst a total of
3×104 cells in 150 µl serum-free medium were added to
the upper chamber. The Transwell chambers (8 µm; 24-well format;
Corning Incorporated, Corning, NY, USA) were incubated at 37°C
overnight. The cells were treated with the various inhibitors for
the indicated times. Following the scraping of non-migrating cells
from the upper surface of the membrane with cotton swabs, crystal
violet was used to stain the cells that had migrated to and invaded
the bottom chamber. The cells were then counted under a microscope
(3 fields at random with ×100 magnification; U-ULS100HG; Olympus
Optical Co. Ltd., Tokyo, Japan). For the invasion assay, the insert
membrane was coated with diluted Matrigel Basement Membrane Matrix
(BD Biosciences, Franklin Lakes, NJ), and the assay was conducted
in a similar manner to the aforementioned assay.
Western blot analysis
Following the treatment of the cells with the
various inhibitors, the cells were then dissolved using Pierce
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc.) containing proteinase inhibitors (2.5 µg/ml leupeptin, 1
µg/ml aprotinin and 1 mM phenylmethanesulfonyl fluoride).
Subsequently, the total protein within the lysates was quantified
using a protein assay kit obtained from Bio-Rad Laboratories Inc.
(Hercules, CA, USA). Furthermore, 50 µg protein was lysed and
separated by sodium dodecyl sulfate polyacrylamide gel
electrophoresis. The membrane was blotted with antibodies against
phosphorylated (p)-AURKA (dilution, 1:3,000; catalog no. 3079P),
AURKA (dilution, 1:1,000; catalog no. 3092S), p-Fak (Y397;
dilution, 1:1000; catalog no. 3283), p-Fak (Y925; dilution,
1:1,000; catalog no. 3284P), Fak (dilution, 1:1,000; catalog no.
3285P), p-Akt473 (dilution, 1:1,000; catalog no. 9271) and Akt
(dilution, 1:1,000; catalog no. 9272), all monoclonal Anti-rabbit
IgG antibodies, obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA), and p-Fak (Y861; dilution, 1:1,000; monoclonal,
Anti-rabbit IgG; catalog no. ab81293) obtained from Abcam
(Cambridge, UK). The antibodies were added for 1 h at room
temperature, and were incubated with horseradish
peroxidase-conjugated secondary antibody for 1 h. During the
procedure, the glyceraldehyde 3-phosphate dehydrogenase level was
regarded as the loading control.
Statistical analysis
The data from all experiments were analyzed with
GraphPad Prism software (version 6; GraphPad Software, Inc., La
Jolla, CA, USA) and shown as the mean±standard deviation. The
Student's t-test was performed to assess the difference
between the experimental and control groups. Values of P<0.05,
indicated by *, were considered to indicate a statistically
significant difference and values of P<0.01, indicated by **,
were considered to indicate a highly statistically significant
difference.
Results
Migration of FaDu and Hep2 cells
decreases following treatment with AURKA, Akt and FAK
inhibitors
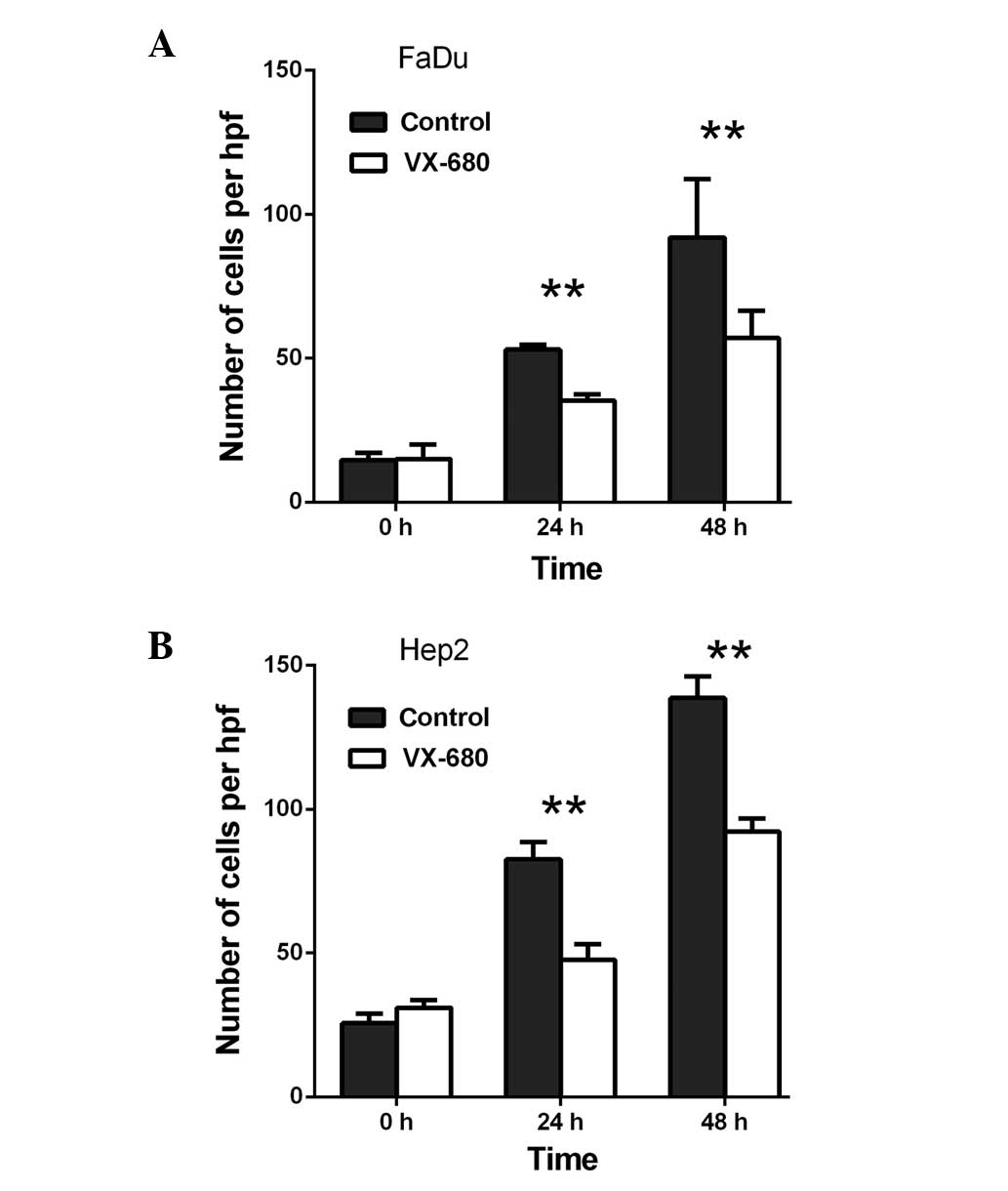
Transwell migration assays were performed in order
to investigate the role of AURKA in the migration of FaDu and Hep2
cells. Initially, when compared with the control, VX-680 decreased
the migration of the FaDu cells to 66.6% at 24 h (P<0.01) and to
62.0% at 48 h (P<0.01) (Fig. 1A).
Meanwhile, in the Hep2 cells, VX-680 decreased migration to 57.6%
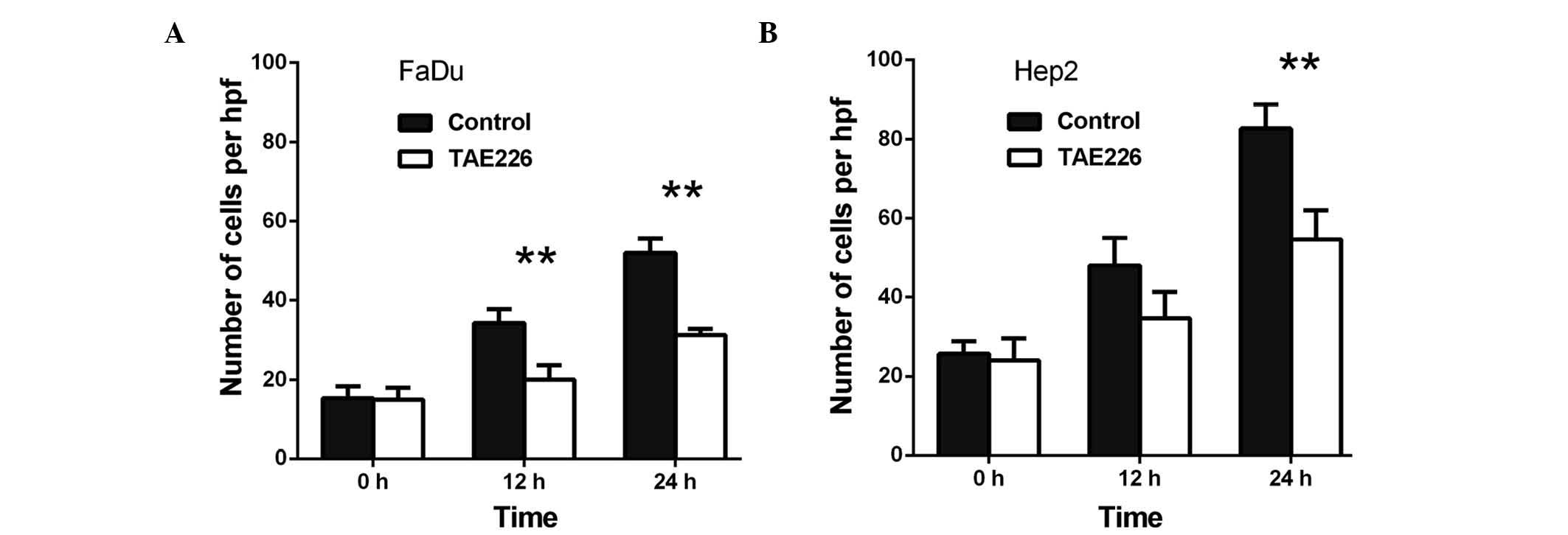
at 24 h (P<0.01) and 66.5% at 48 h (P<0.01) (Fig. 1B). Secondly, when compared with the
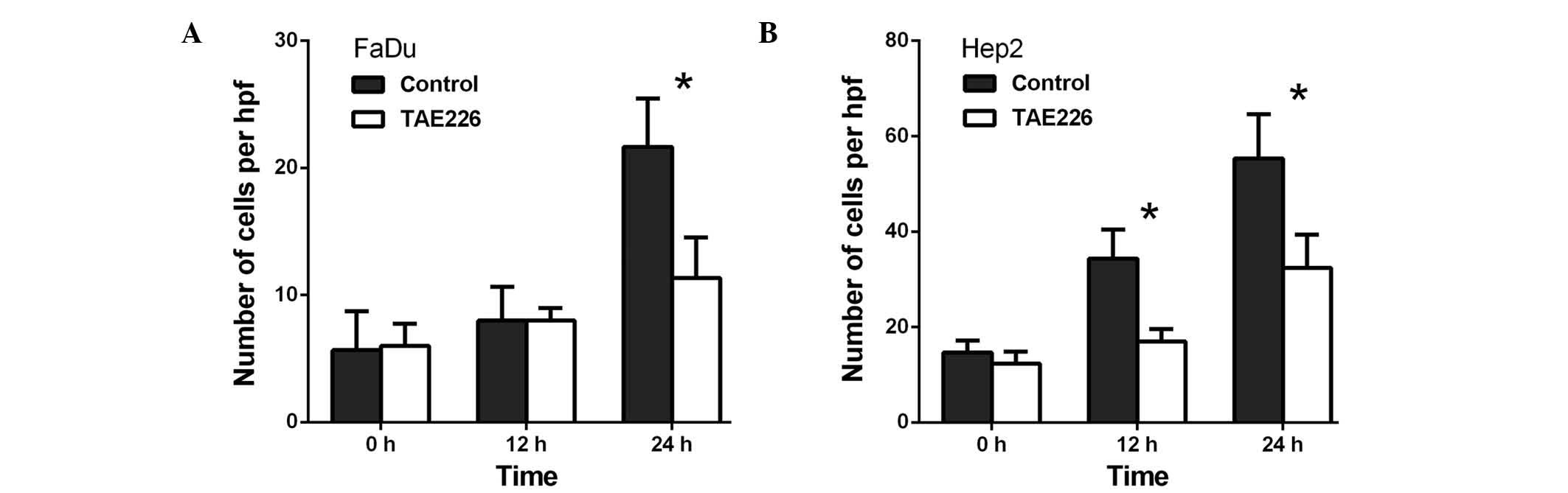
control, TAE226 was observed to decrease the migration of the FaDu
cells to 58.3% at 12 h (P<0.01) and to 60.3% at 24 h
(P<0.01), respectively (Fig. 2A).
Regarding the Hep2 cells, TAE226 was observed to decrease migration
to 72.0% at 12 h (P<0.01) and to 66.1% at 24 h (P<0.01)
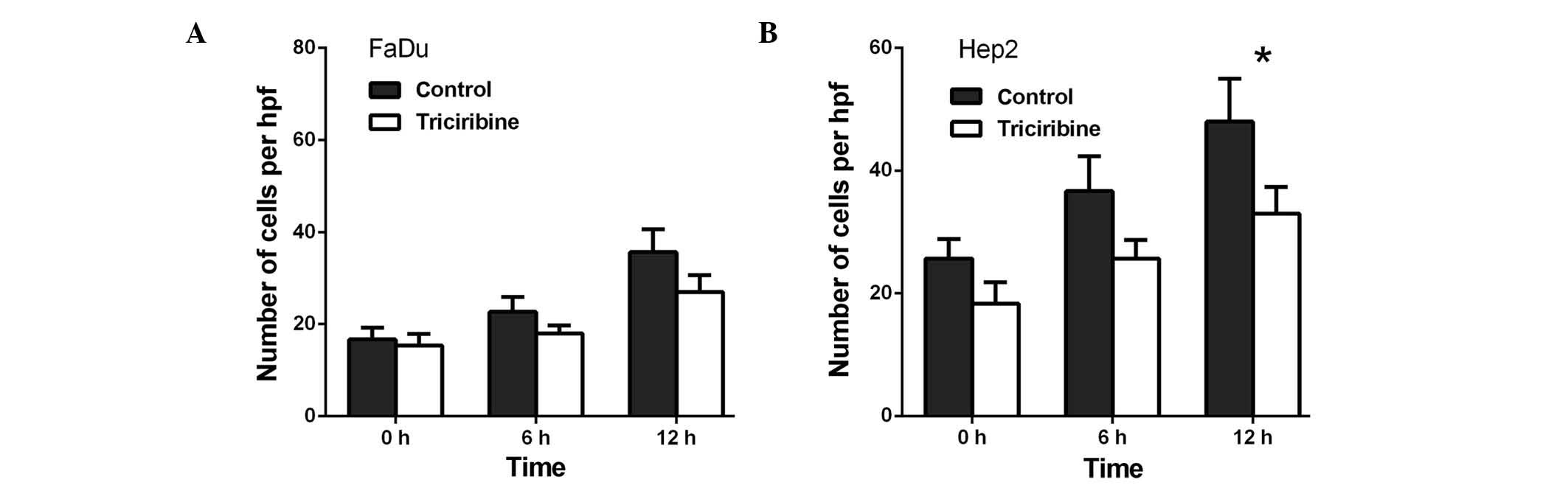
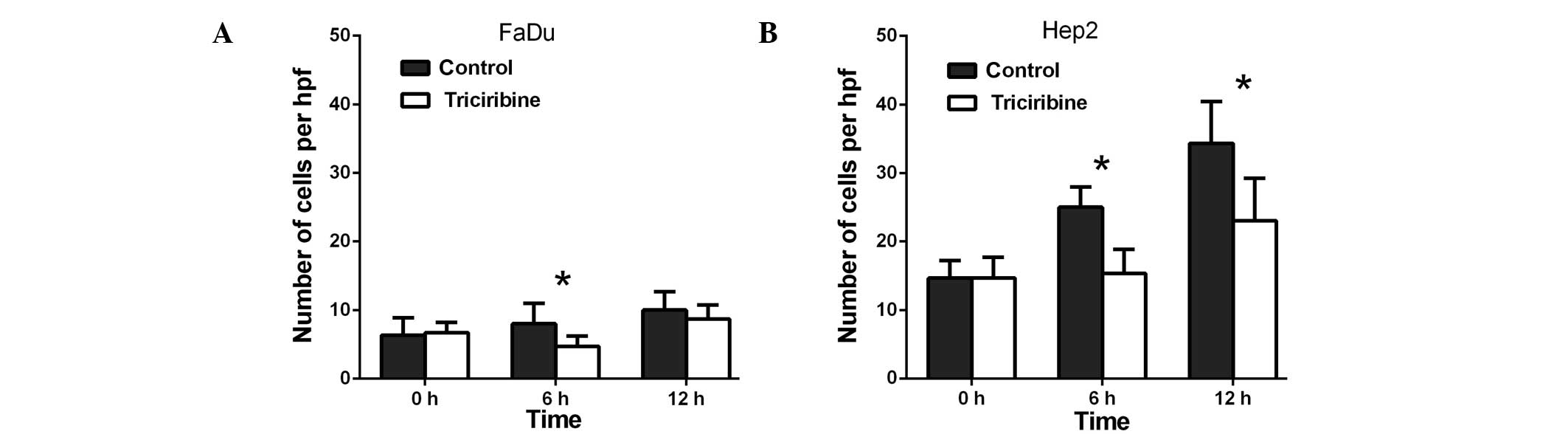
(Fig. 2B). Lastly, triciribine
decreased the migration of the FaDu cells to 79.3% at 6 h
(P>0.05) and to 75.6% at 12 h (P>0.05; Fig. 3A), whereas in the Hep2 cells,
triciribine decreased the migration to 71.8% at 6 h (P>0.05) and
68.8% at 12 h (P<0.05) (Fig.
3B).
Capability of FaDu and Hep2 cell
invasion is decreased following the treatment with AURKA, Akt and
FAK inhibitors
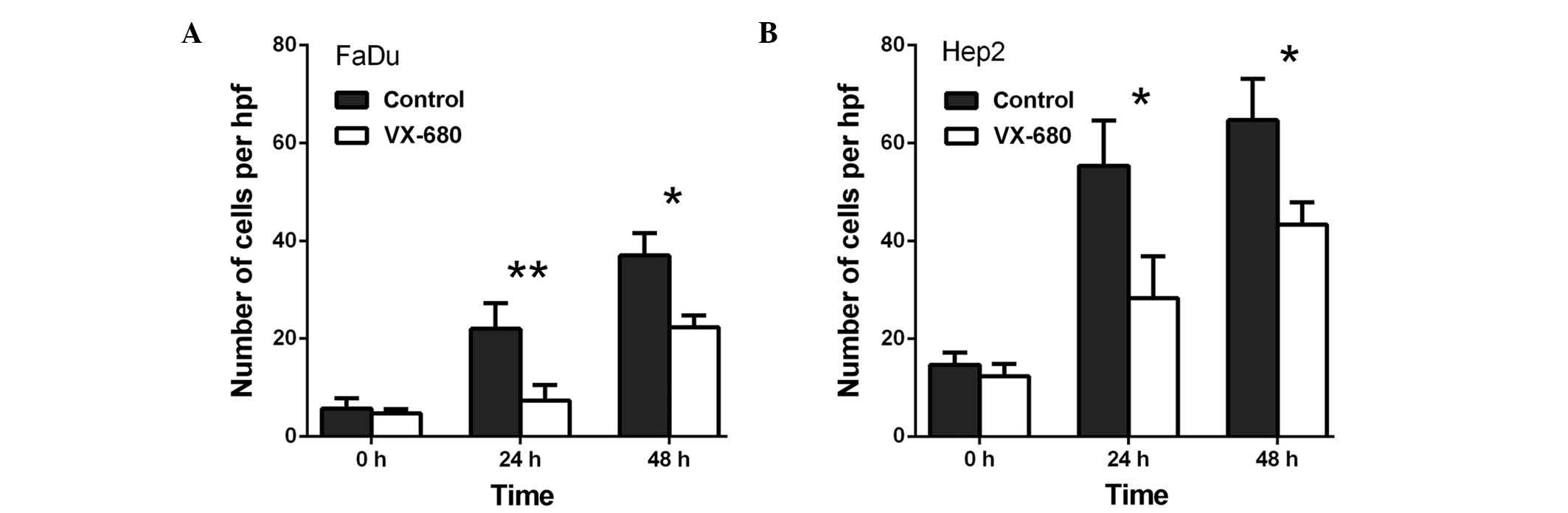
Transwell invasion assays were performed to
investigate the role of AURKA in the invasion of the FaDu and Hep2
cells. As presented in Fig. 4, VX-680
decreased the invasion of the FaDu cells to 33.3% at 24 h
(P<0.01) and to 60.4% at 48 h (P<0.05) (Fig. 4A), whereas VX-680 was observed to
decrease the invasion of the Hep2 cells to 51.2% at 24 h
(P<0.05) and to 67.0% at 48 h (P<0.05) (Fig. 4B). Secondly, TAE226 was observed to
decrease the invasion of the FaDu cells to 52.1% at 24 h
(P<0.05; Fig. 5A), whereas in the
Hep2 cells, TAE226 decreased invasion to 49.6% at 12 h (P<0.05)
and to 58.5% at 24 h (P<0.05) (Fig.
5B). Lastly, triciribine decreased the invasion of the FaDu
cells to 58.8% at 6 h (P<0.05) and to 86.7% at 12 h (P<0.05)
(Fig. 6A), whilst in the Hep2 cells,
triciribine decreased invasion to 61.3% at 6 h (P<0.05) and to
67.1% at 12 h (P<0.05) (Fig.
6B).
Suppression of AURKA inhibits
p-FAKY397 phosphorylation
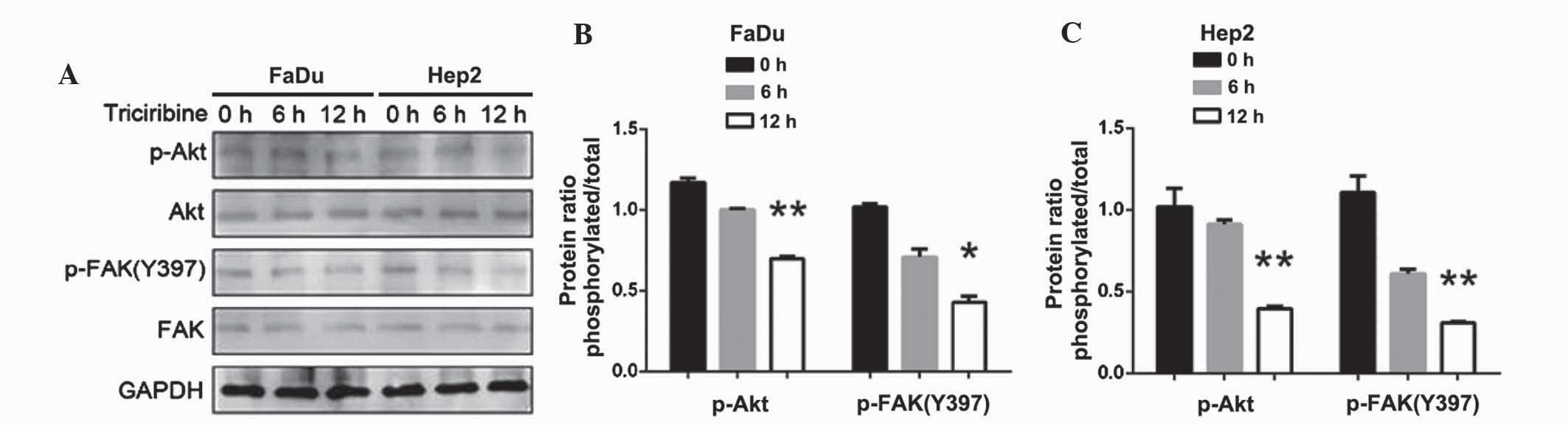
The FaDu and Hep2 cells were treated with 75 nM
VX-680, with western blot analysis then used with the p-FAK
antibody to examine the effect of AURKA suppression on Y397
phosphorylation. Following the downregulation of AURKA, p-AURKA and
p-FAK Y397 expression decreased (P<0.05); however, the
expression of p-FAK Y861 and p-FAK Y925 did not change (Fig. 7). The results also demonstrated that
the treatment with VX-680 at 24 h did not significantly decrease
p-FAK Y397 phosphorylation (P<0.05), but did completely block
Y397-phosphorylation in the Hep2 cells at 48 h (P<0.01; Fig. 7). The expression of p-Akt was also
slightly decreased (P<0.01 in FaDu cells; P<0.05 in Hep2
cells; Fig. 7).
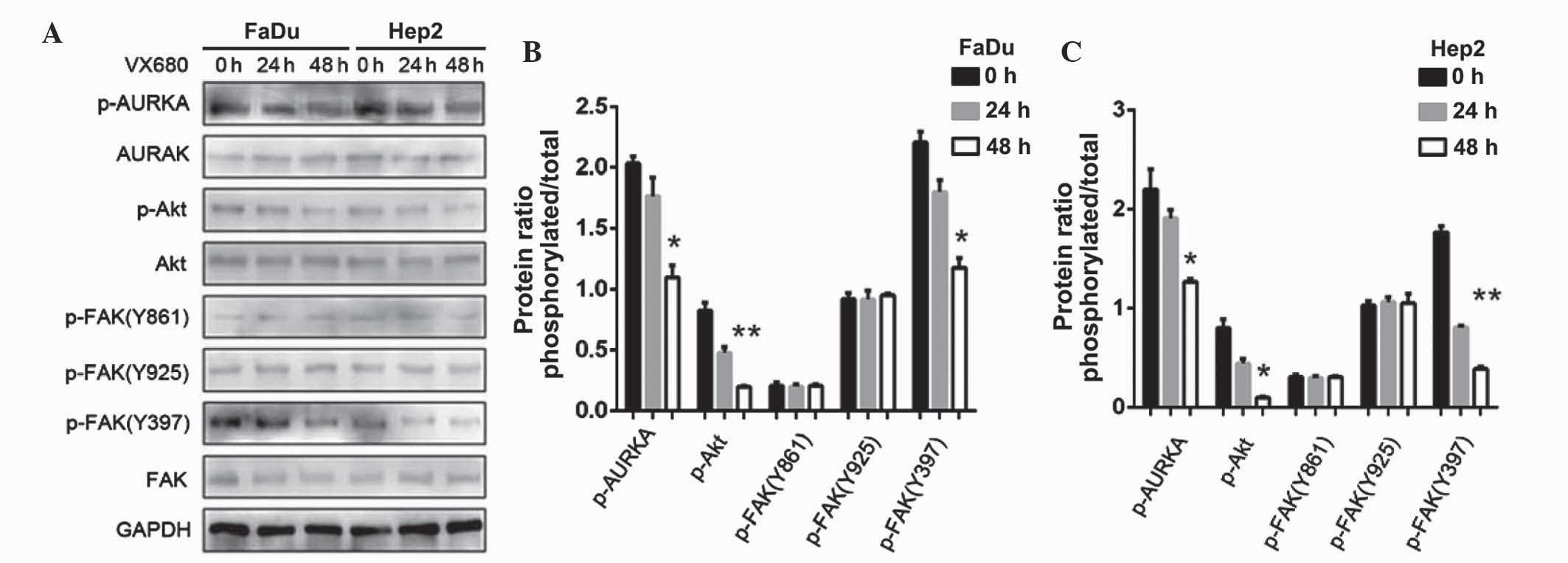 | Figure 7.(A) Effects of downregulation of AURKA
on FAK and Akt phosphorylation in FaDu and Hep2 cells. p-FAK Y397
and p-Akt were extremely decreased. (B) The expression levels of
p-AURKA, p-Akt, p-FAK(Y861), p-FAK(Y925) and p-FAK(Y397) in FaDu
cells were normalized to GAPDH. (C) The expression levels of
p-AURKA, p-Akt, p-FAK(Y861), p-FAK(Y925) and p-FAK(Y397) in Hep2
cells were normalized to GAPDH. The bar represents the mean ±
standard deviation of results from three separate experiments.
*P<0.05; **P<0.01. AURKA, Aurora kinase A; p, phosphorylated;
Akt, protein kinase B; FAK, focal adhesion kinase; GAPDH,
glyceraldehyde 3-phosphate dehydrogenase. |
AURKA promotes migration and invasion
through the Akt-FAK signaling pathway
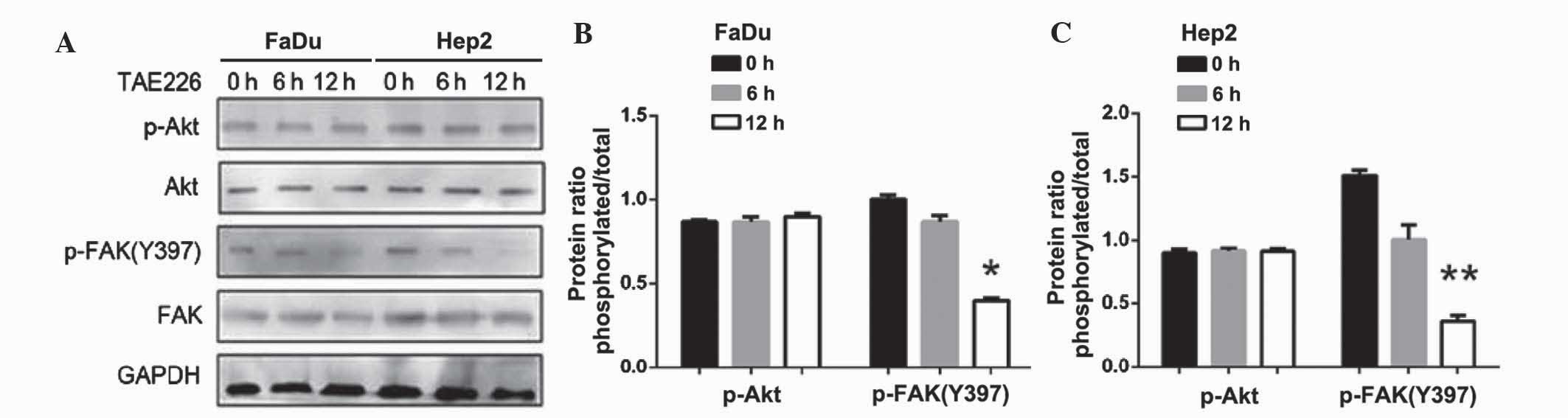
Following the inhibition of Akt, the expression of
p-Akt (P<0.01) and p-FAK (Y397) were decreased (P<0.05;
Fig. 8). By contrast, following the
inhibition of FAK, the expression of p-FAK (Y397) decreased, but
p-Akt expression did not change (P<0.05 in FaDu cells; P<0.01
in Hep2 cells; Fig. 9). Thus, the
downregulation of AURKA suppressed p-FAK via the inhibition of
p-Akt.
Discussion
The role of AURKA as an oncogene has been supported
by a number of previous studies (3,27). Zhang
et al demonstrated that the suppression of AURKA expression
inhibits the growth and invasiveness of laryngeal squamous cell
carcinoma cells, in vitro and in vivo (28). AURKA has been observed to positively
regulate the G2 to M phase of the cell cycle, and
activation of AURKA in late G2 is inhibited by DNA
damage (3). Furthermore, high
expression levels of AURKA have been associated with late clinical
stages and regional metastasis in HNSCC (29). With research progression, a greater
number of trials have been performed to investigate the treatment
of HNSCC with Aurora kinase inhibitors (30). Li et al demonstrated that Akt
promotes cell survival through its ability to phosphorylate and
activate several pro-apoptotic targets. Subsequent consequences of
this include the phosphorylation of Fak and the activation of
signal transduction pathways, ultimately contributing to
AURKA-mediated tumorigenesis (31).
However, the pathway by which the migration and invasion of HNSCC
was enhanced was not determined. Chemotherapy, radiotherapy and
surgery have made great advances, however, the rate of morbidity
remains high in HNSCC (2). In the
present study, it was demonstrated that AURKA upregulated FAK via
the activation of the AURKA/Akt/FAK signaling pathway. This
subsequently led to the promotion of cell migration and invasion in
HNSCC, indicating a strong association with the overexpression of
AURKA and the AURKA/Akt/FAK signaling pathway.
The current study provided evidence that Aurora
kinase inhibitors, which are implicated in the AURKA/Akt/FAK
signaling pathway, should not only be considered in clinical trials
for the treatment of HNSCC patients, but should also be tested in
combination with other therapeutic drugs. During the process of
future clinical trials, it may be beneficial for pre-clinical
investigators to examine the various doses and schedules. Rational
emergency treatment and advisable drug pharmacodynamics should be
taken into account in such trials with volunteers. Finally, further
investigation may determine the potential of using aurora kinase
inhibitors as a target therapy for the treatment of other types of
cancer.
In conclusion, a better understanding of the role
that AURKA may serve in recurrence and metastasis, alongside the
association between AURKA and Akt pathways in HNSCC, provides a
novel insight and rationale for the possibility of further combined
molecular targeting therapy in HNSCC and other types of cancer.
Acknowledgements
This study was supported by a grant from the
Research Project of Shanghai Science and Technology Commission (no.
12ZR1418700).
References
|
1
|
Jemal A, Murray T, Samuels A, Ghafoor A,
Ward E and Thun MJ: Cancer statistics, 2003. CA Cancer J Clin.
53:5–26. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hunter KD, Parkinson EK and Harrison PR:
Profiling early head and neck cancer. Nat Rev Cancer. 5:127–135.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marumoto T, Zhang D and Saya H: Aurora-A -
a guardian of poles. Nat Rev Cancer. 5:42–50. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stenoien DL, Sen S, Mancini MA and
Brinkley BR: Dynamic association of a tumor amplified kinase,
Aurora-A, with the centrosome and mitotic spindle. Cell Motil
Cytoskeleton. 55:134–146. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baba Y, Nosho K, Shima K, Irahara N, Kure
S, Toyoda S, Kirkner GJ, Goel A, Fuchs CS and Ogino S: Aurora-A
expression is independently associated with chromosomal instability
in colorectal cancer. Neoplasia. 11:418–425. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kitzen JJ, de Jonge MJ and Verweij J:
Aurora-A kinase inhibitors. Crit Rev Oncol Hematol. 73:99–110.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Milam MR, Gu J, Yang H, Celestino J, Wu W,
Horwitz IB, Lacour RA, Westin SN, Gershenson DM, Wu X and Lu KH:
STK15 F31I polymorphism is associated with increased uterine cancer
risk: A pilot study. Gynecol Oncol. 107:71–74. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ogawa E, Takenaka K, Katakura H, Adachi M,
Otake Y, Toda Y, Kotani H, Manabe T, Wada H and Tanaka F:
Perimembrane Aurora-A expression is a significant prognostic factor
in correlation with proliferative activity in non-small-cell lung
cancer (NSCLC). Ann Surg Oncol. 15:547–554. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang R, Wang JH, Chu XY, Geng HC and Chen
LB: Expression of STK15 mRNA in hepatocellular carcinoma and its
prognostic significance. Clin Biochem. 42:641–647. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang SB, Zhou XB, Zhu HX, Quan LP, Bai JF,
He J, Gao YN, Cheng SJ and Xu NZ: Amplification and overexpression
of Aurora-A in esophageal squamous cell carcinoma. Oncol Rep.
17:1083–1088. 2007.PubMed/NCBI
|
|
11
|
Zhang H, Chen X, Jin Y, Liu B and Zhou L:
Overexpression of Aurora-A promotes laryngeal cancer progression by
enhancing invasive ability and chromosomal instability. Eur Arch
Otorhinolaryngol. 269:607–614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang LH, Xiang J, Yan M, Zhang Y, Zhao Y,
Yue CF, Xu J, Zheng FM, Chen JN, Kang Z, et al: The mitotic kinase
Aurora-A induces mammary cell migration and breast cancer
metastasis by activating the Cofilin-F-actin pathway. Cancer Res.
70:9118–9128. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sakakura C, Hagiwara A, Yasuoka R, Fujita
Y, Nakanishi M, Masuda K, Shimomura K, Nakamura Y, Inazawa J, Abe T
and Yamagishi H: Tumour-amplified kinase BTAK is amplified and
overexpressed in gastric cancers with possible involvement in
aneuploid formation. Br J Cancer. 84:824–831. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou H, Kuang J, Zhong L, Kuo WL, Gray JW,
Sahin A, Brinkley BR and Sen S: Tumour amplified kinase STK15/BTAK
induces centrosome amplification, aneuploidy and transformation.
Nat Genet. 20:189–193. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gritsko TM, Coppola D, Paciga JE, Yang L,
Sun M, Shelley SA, Fiorica JV, Nicosia SV and Cheng JQ: Activation
and overexpression of centrosome kinase BTAK/Aurora-A in human
ovarian cancer. Clin Cancer Res. 9:1420–1426. 2003.PubMed/NCBI
|
|
16
|
Weaver KL, Alves-Guerra MC, Jin K, Wang Z,
Han X, et al: NACK is an integral component of the Notch
transcriptional activation complex and is critical for development
and tumorigenesis. Cancer Res. 74:4741–4751. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Archewa P, Pata S, Chotjumlong P,
Supanchart C, Krisanaprakornkit S and Iamaroon A: Akt2 and p-Akt
overexpression in oral cancer cells is due to a reduced rate of
protein degradation. J Investig Clin Dent. Epub ahead of print.
2015.
|
|
18
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gabarra-Niecko V, Schaller MD and Dunty
JM: FAK regulates biological processes important for the
pathogenesis of cancer. Cancer Metastasis Rev. 22:359–374. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McLean GW, Carragher NO, Avizienyte E,
Evans J, Brunton VG and Frame MC: The role of focal-adhesion kinase
in cancer - a new therapeutic opportunity. Nat Rev Cancer.
5:505–515. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Canel M, Secades P, Rodrigo JP, Cabanillas
R, Herrero A, Suarez C and Chiara MD: Overexpression of focal
adhesion kinase in head and neck squamous cell carcinoma is
independent of fak gene copy number. Clin Cancer Res. 12:3272–3279.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang F, Guo X, Yang G, Rosen DG and Liu J:
AURKA and BRCA2 expression highly correlate with prognosis of
endometrioid ovarian carcinoma. Mod Pathol. 24:836–845. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guan Z, Wang XR, Zhu XF, Huang XF, Xu J,
Wang LH, Wan XB, Long ZJ, Liu JN, Feng GK, et al: Aurora-A, a
negative prognostic marker, increases migration and decreases
radiosensitivity in cancer cells. Cancer Res. 67:10436–10444. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hochwald SN, Nyberg C, Zheng M, Zheng D,
Wood C, Massoll NA, Magis A, Ostrov D, Cance WG and Golubovskaya
VM: A novel small molecule inhibitor of FAK decreases growth of
human pancreatic cancer. Cell Cycle. 8:2435–2443. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang L, Dan HC, Sun M, Liu Q, Sun XM,
Feldman RI, Hamilton AD, Polokoff M, Nicosia SV, Herlyn M, et al:
Akt/protein kinase B signaling inhibitor-2, a selective small
molecule inhibitor of Akt signaling with antitumor activity in
cancer cells overexpressing Akt. Cancer Res. 64:4394–4399. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng R, Chen X, Yu Y, Su L, Yu B, Li J,
Cai Q, Yan M, Liu B and Zhu Z: miR-126 functions as a tumour
suppressor in human gastric cancer. Cancer Lett. 298:50–63. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Giet R, Petretti C and Prigent C: Aurora
kinases, aneuploidy and cancer, a coincidence or a real link.
TRENDS in Cell Biology. 15:241–250. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang H, Chen X, Liu B and Zhou L: Effects
of stable knockdown of Aurora kinase A on proliferation, migration,
chromosomal instability, and expression of focal adhesion kinase
and matrix metalloproteinase-2 in HEp-2 cells. Mol Cell Biochem.
357:95–106. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Reiter R, Gais P, Jütting U, Steuer-Vogt
MK, Pickhard A, Bink K, Rauser S, Lassmann S, Höfler H, Werner M
and Walch A: Aurora kinase A messenger RNA overexpression is
correlated with tumor progression and shortened survival in head
and neck squamous cell carcinoma. Clin Cancer Res. 12:5136–5141.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Naruganahalli KS, Lakshmanan M, Dastidar
SG and Ray A: Therapeutic potential of Aurora kinase inhibitors in
cancer. Curr Opin Investig Drugs. 7:1044–1051. 2006.PubMed/NCBI
|
|
31
|
Li DW, Sun YJ, Sun ZF and Dong P:
Involvement of focal adhesion kinase in cellular proliferation,
apoptosis and prognosis of laryngeal squamous cell carcinoma. J
Laryngol Otol. 126:1127–1133. 2012. View Article : Google Scholar : PubMed/NCBI
|