Introduction
Breast cancer has become the leading cause of
mortality in females and it is a significant threat to the health
of females (1). It is believed that
breast carcinoma is a heterogeneous disease with genetic and
non-genetic etiology (2). To date, it
has been confirmed that breast cancer susceptibility genes include
breast cancer 1 (BRCA1) and BRCA2. However, these genes only
account for 5 to 10% of cases with inherited mutations, and the
majority of breast cancers are sporadic (3,4). Previous
genome-wide association studies (GWAS) involving a large number of
samples have revealed novel single-nucleotide polymorphisms (SNPs)
of breast cancer. They are respectively located on genes FGFR2,
TNRC9/TOX3, MAP3K1, LSP1 and CASP8 (5). Among these, TOX high mobility group box
family member 3 (TOX3) has been further confirmed to have a
stronger association with the risk of breast cancer by GWAS studies
in different people (6–13).
TOX3 is a member of a high mobility group (HMG) of
box proteins that is associated with the regulation of thymocyte
selection (14), and is also called
trinucleotide repeat-containing gene 9 (TNRC9) or CAG trinucleotide
repeat-containing gene F9 (CAGF9) (15). Structurally, TOX3 is a nucleoprotein
which includes a nuclear localization signal in the N-terminal
domain, a HMG box domain which is able to change the chromatin
structure by bending and unwinding DNA, and a C-terminal
polyglutamine stretch. It is located on chromosome 16q12.1
(16). Its structure suggests that it
may be closely associated with transcription. TOX3 is identified as
a novel regulator of calcium-dependent transcription and interacts
with the CREB-CBP complex to regulate CRE-mediated transcription in
rat neuronal cells (17). Dittmer
et al observed that overexpression of TOX3 protects neuronal
cells from cell death by inducting anti-apoptotic transcripts and
inhibiting pro-apoptotic transcripts; it depends on the
phosphorylated CREB or CITED1 within the transcriptionally active
complex interacting with the native CREB and inducing the
CREB-responsive BCL-2 promoter (18).
Furthermore, there are certain studies demonstrating that TOX3 is
correlated with other carcinomas (19–21). A
study by Birkenkamp-Demtroder et al revealed that TOX3
overexpression in bladder cancer cells reduces cell proliferation
and affects the interferon signaling pathway (15). In addition, TOX3 expression was
observed to be notably upregulated in lung adenocarcinoma compared
with control tissues (20). However,
there is increasing evidence demonstrating that TOX3 is closely
correlated with the risk of breast cancer. Fasching et al
reported that TOX3 was associated with overall survival in breast
carcinoma (22).
The expression level of TOX3 in breast cancer
remains unclear. There is evidence that high mRNA expression levels
of TOX3 occur in patients with shorter overall survival, and a
positive correlation has been observed between the mRNA expression
level of TOX3 and breast carcinomas with metastasis (23). However, a study by Riaz et al
indicated that the risk alleles (rs3803662 and rs12443621) were
associated with lower expression of TOX3 mRNA and suggested a tumor
suppressor role of TOX3 (24).
Additionally, susceptibility loci in TOX3 had a stronger
association with ER-positive breast cancer than ER-negative breast
cancer (8,25). However, the mRNA expression level does
not represent protein function. It is unclear whether TOX3 is
involved in breast cancer tumorigenesis or ER-positive breast
cancer, and therefore it is critical to understand the expression
levels of TOX3 protein in human breast carcinoma and controls. Few
studies have investigated TOX3 protein expression in a large number
of samples in relation to clinicopathological characteristics. The
aim of the present study was to measure the expression of TOX3
protein in breast cancer, controls and ER-positive or negative
carcinoma, to check whether TOX3 demonstrated an association with
clinicopathological characteristics of patients and tumors, and to
provide a comprehensive evaluation for TOX3 in breast cancer
tumorigenesis.
Materials and methods
Human tissue samples
A breast cancer tissue microarray purchased from US
Biomax (Rockville, MD, USA) were used to assess TOX3 protein
expression with immunohistochemical staining. It contained 267
human breast cancer tissue samples (comprising 217 invasive ductal
carcinomas, 45 invasive lobular carcinoma, 2 medullary carcinoma, 2
mucinous carcinomas and 1 invasive papillary carcinoma) and 25
healthy controls. The age of tumor patients ranged from 27 to 82
years with a mean age of 49.3 years, and the age of controls ranged
from 15 to 50 years with an average age of 30.2 years. The
pathological information of patients is shown in Table I. The expression of ER was assessed in
66 tumor patients, and there were noted to be 22 ER-positive
patients and 44 ER-negative cases. Additionally, three fresh breast
cancer tissues and matched controls used to detect TOX3 expression
by western blot analysis and quantitative polymerase chain reaction
(qPCR) were obtained from the Affiliated Hospital of Qiqihar
Medical University, China. The use of these samples for this study
was approved by the ethics committee of Qiqihar Medical University,
and the written informed consent was obtained from the
subjects.
 | Table I.Pathological information of
patients. |
Table I.
Pathological information of
patients.
| Pathological
variables | Number of
patients |
|---|
| Normal breast
tissues | 25 |
| Breast cancer
tissues | 267 |
| TNM grading |
|
|
T1/T2/T3/T4 | 32/190/27/18 |
| Lymph node
metastasis |
|
|
N0/N1/N2/N3 | 210/48/8/1 |
| TNM stage |
|
|
I/II/III/IV | 31/209/24/3 |
| Estrogen
receptor |
|
|
ER+/ER− | 22/44 |
Cell culture
Human breast cancer cell lines (ZR-75-1, MDA-MB-231,
MCF-7 and Bcap-37) with ER-positive or negative expression were
purchased from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). ZR-75-1 and Bcap-37 cells were maintained in
Roswell Park Memorial Institute-1640 (RPMI-1640) medium, and
MDA-MB-231 cells were cultured in Leibovitz's-15 (L15) medium.
MCF-7 cells were maintained in minimum essential medium (MEM). The
media were supplemented with 10% fetal bovine serum (FBS),
penicillin (100 IU/ml) and streptomycin (100 µg/ml). In addition to
the aforementioned items, the RPMI-1640 medium used for ZR-75-1
cells included glucose (2.5 g/l) and sodium pyruvate (0.11 g/l) and
MEM medium used for MCF-7 cells contained sodium pyruvate (0.11
g/l) and bovine insulin (0.01 mg/ml). ZR-75-1, MCF-7 and Bcap-37
cells were incubated in a 5% CO2 humidified incubator at
37°C and MDA-MB-231 cells were cultured at 37°C in a humidified
atmosphere of 100% air.
Immunohistochemistry
Immunohistochemistry was performed to analyze the
protein expression of TOX3. The tissue microarray section was
pretreated at 60°C for 1 h, then dewaxed and rehydrated by xylene
and graded alcohol. Antigen retrieval was facilitated in sodium
citrate buffer for 2 min in an autoclave at 121°C. Endogenous
peroxidase activity was blocked with 3% hydrogen peroxide/methanol
for 10 min at room temperature. Thereafter, the section was
incubated with goat serum for 10 min at room temperature and then
with TOX3 rabbit polyclonal antibody diluted at 1:80 (Abgent, San
Diego, CA, USA) at 4°C overnight. Staining was implemented with an
UltraSensitiveTM SP IHC kit (Fuzhou Maixin Biotech, Fujian, China)
according to the manufacturer's instructions. Finally, the section
was stained with 3,3′-diaminobenzidine as the chromogen and
counterstained with hematoxylin.
TOX3 expression was estimated semiquantitatively
according to the TOX3-immunopositive cell percentage in tumor cells
(0, negative staining; 1, 0–10% positive; 2, 11–50% positive; 3,
51–80% positive; 4, 80–100% positive) (26). Secondly, the staining intensity of
positive cells was scored as follows: 0, negative; 1, faint; 2,
moderate; 3, strong. On the basis of these data, the results were
further scored 0–3 (the percentage of positive cells multiplied by
the staining intensity score, resulting in a score from 0 to 12; a
score of 0 or 1 was considered as negative, and 2–3 was considered
as 1+, 4–7 as 2+, and 8–12 as 3+) (27). The results were analyzed by two
pathologists respectively who were blinded to the
clinicopathological information.
qPCR
Total RNA was extracted from cells or fresh tumor
tissues with RNAiso reagent (Takara Biotechnology, Dalian, China).
cDNA was synthesized using a PrimeScript™ RT reagent kit (Takara
Biotechnology) according to the manufacturer's instructions. The
qPCR reaction was performed with a SYBR Premix Ex Taq™ (Tli RNaseH
Plus) kit (Takara Biotechnology) in a final reaction volume of 50
µl containing 2X SYBR Premix Ex Taq™ (Tli RNaseH Plus) (Takara
Biotechnology), ROX reference dye and the corresponding primers.
The PCR conditions were as follows: 1 cycle of 30 sec at 95°C, 40
cycles of 5 sec at 95°C, and 31 sec at 60°C. The PCR reaction was
implemented in an Applied Biosystems 7300 Real-Time PCR system
(Applied Biosystems, Foster City, CA, USA). β-actin was used as the
internal control. The sequences of following primers were used for
the qPCR reaction: 5′-CTGGGACGACATGGAGAAAA-3′ (sense) and
5′-AAGGAAGGCTGGAAGAGTGC-3′ (antisense) for the β-actin gene;
5′-TATGCCTCACACATCTCCTTCA-3′ (sense) and 5′-ATGGCTCTGTTGGCTTCATC-3′
(antisense) for the TOX3 gene. The qPCR analysis was performed
using the 2-ΔΔCt method (28). The
experiments were implemented in triplicate to ensure
reproducibility.
Western blot
Western blot analysis was performed to detect the
protein expression of TOX3 in breast tumor, corresponding control
tissue and breast cancer cell lines. Proteins were isolated with
RIPA lysis buffer (Beyotime Biotechnology, Haimen, China) from
tissue specimens. The protein concentration was detected
quantitatively using a bicinchoninic acid protein assay kit
(Beijing ComWin Biotech, Beijing, China). Protein supernatant was
added to lane marker loading buffer and boiled at 100°C for 5 min.
Furthermore, the proteins were separated by 10% sodium dodecyl
sulfate polyacrylamide gel electrophoresis and then transferred to
nitrocellulose membrane by electrophoresis, and nonspecific protein
bindings were blocked with blocking buffer at 4°C overnight.
Subsequently, the membranes were incubated with TOX3 rabbit
monoclonal antibody (1:1200; Abcam, Cambridge, UK) and GAPDH mouse
polyclonal antibody (1:3000; Beijing ComWin Biotech, Beijing,
China) for 3 h at room temperature and washed with Tris-buffered
saline with 0.1% Tween-20 (TBST) three times. Next, they were
incubated with IgG secondary antibody (1:3000; Cell Signaling
Technology, MA, USA) for 2 h at room temperature and washed again
with TBST three times. Finally, the blots were developed using a
SuperSignal West Pico substrate kit (Thermo Fisher Scientific, MA,
USA). GAPDH was used as the internal control to measure the
relative expression of TOX3. Band intensities were determined using
ImageJ2× software version 2.1.4.7 (National Institutes of Health,
MD, USA).
Statistical analysis
The results from qPCR and western blot analysis were
expressed as the means ± standard deviation and assessed using
Student's t-test. Pearson's χ2 test and the Wilcoxon rank test were
used to analyze categorical associations. P<0.05 was considered
to indicate a statistically significant difference. All the
analyses were performed using SPSS 17.0 software (SPSS Inc.,
Chicago, IL, USA).
Results
Expression of TOX3 is increased in
breast tumor tissue compared with controls
In this study, immunohistochemistry was used to
detect the protein expression of TOX3 in tissue microarray samples
consisting of 25 control breast tissues and 267 breast carcinoma
tissue specimens. The mean immunoreactive score (IRS) of TOX3 in
breast carcinoma tissue was notably higher than that in controls
(Table II). Additionally, the levels
of TOX3 expression in breast carcinoma demonstrated a positive
correlation with the grading and staging of breast tumors. High
TOX3 expression levels were noted in advanced stages (T3 and T4).
In addition, the mean IRS of TOX3 in stage T3 and T4 was
significantly increased compared with that in stage T1 (Table II). Furthermore, compared with T1
stage, the percentage of TOX3 high expression (IRS=3+) in T3 and T4
stages was notably increased (Table
III, Fig. 1). The levels of TOX3
expression in tumors with lymph node metastasis (N2 stage) were
significantly higher than those without regional lymph node
metastasis (N0 stage), but there was no statistical significance in
N3 stage (Table III). Moreover,
high TOX3 expression in stage III and IV was also observed
(Table III). To further confirm the
results of immunohistochemistry, qPCR and western blot analysis
were implemented to detect the expression of TOX3 mRNA and protein
in the fresh tissues. Total RNA was obtained from breast carcinoma
tissues and matched control breast tissue of three patients. As
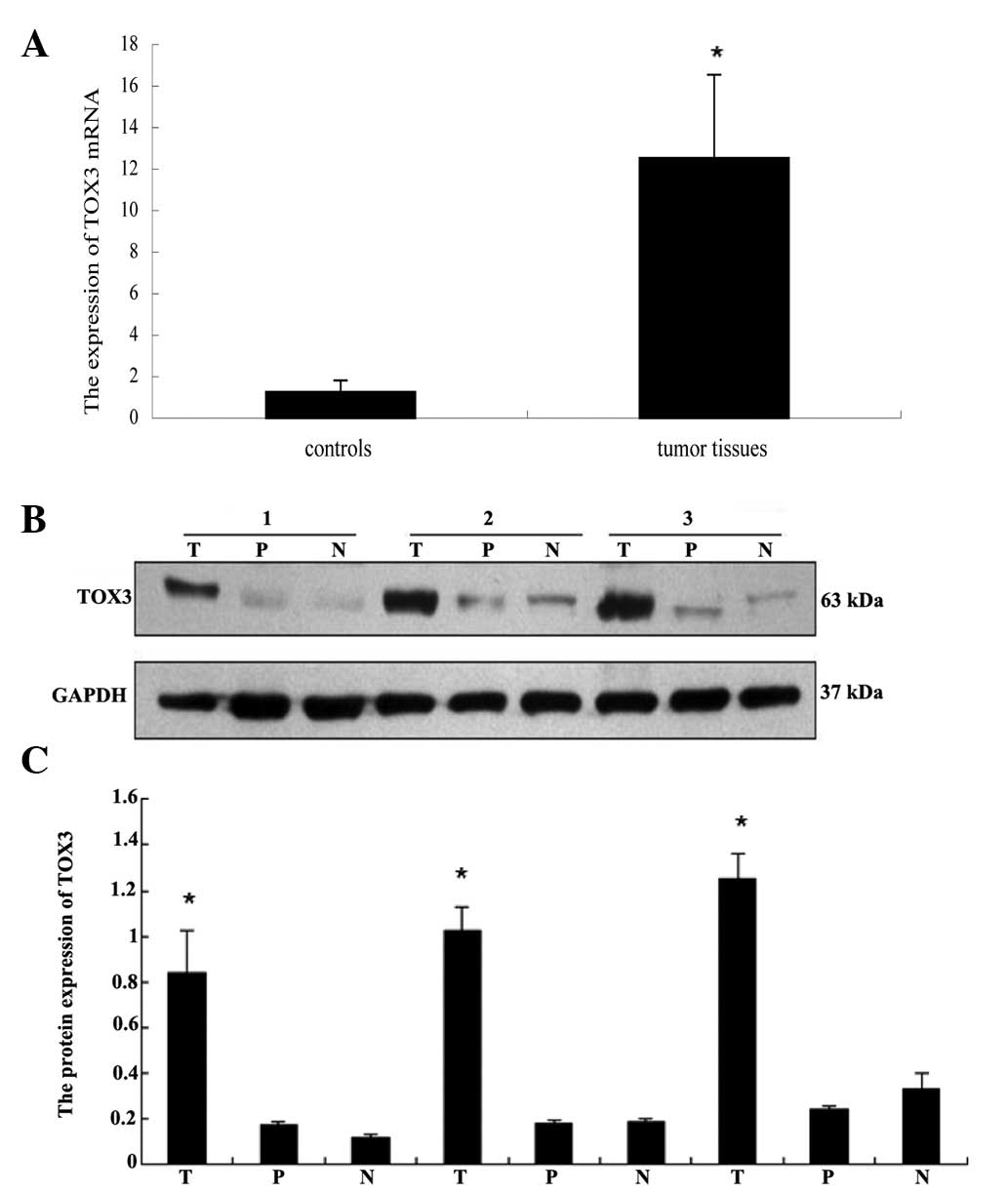
shown in Fig. 2, compared with the
matched adjacent control breast tissue, the expression levels of
TOX3 mRNA were upregulated significantly in breast cancer specimens
(P<0.05; Fig. 2A). The western
blot analysis results further confirmed the increased expression of
TOX3 in breast malignancies compared with controls (P<0.05;
Fig. 2B and C). These results were
consistent with the immunohistochemistry results.
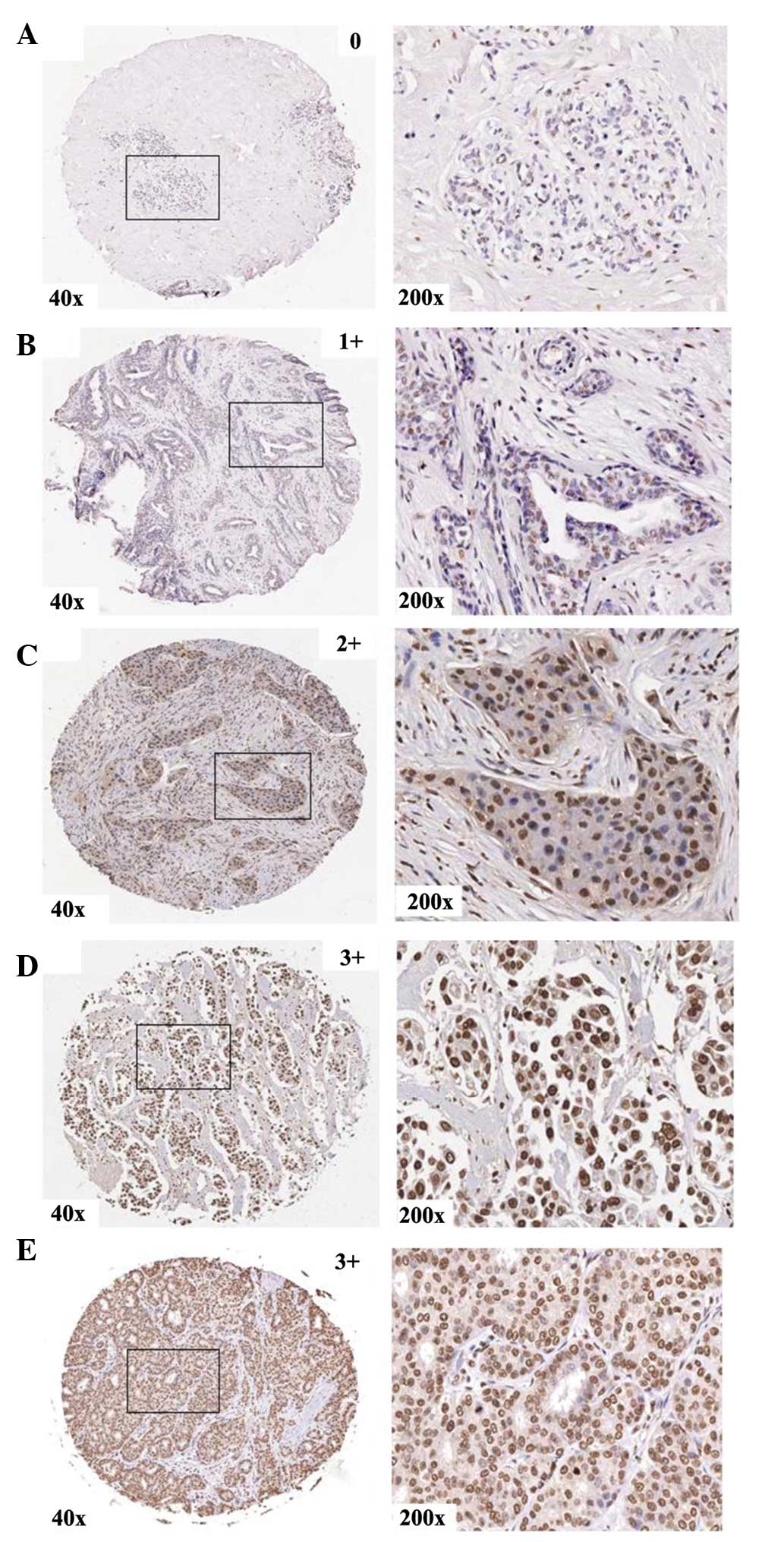 | Figure 1.Protein expression of TOX3 in human
control breast tissues and breast carcinomas in association with
pathological characteristics. The representative tissue scores for
control breast tissues (A, score 0) and tumor-node-metastasis
grading of breast carcinoma (B, T1 stage, 1+score; C, T2 stage,
score 2+; D, T3 stage, score 3+; E, T4 stage, score 3+) are given.
The left panels reveal the immunohistochemistry staining at 40×
magnification and the right panels at 200× magnification. As shown,
the protein expression of TOX3 was upregulated in breast carcinoma,
and was particularly higher in T3 and T4 stage than in T1
stage. |
| Table II.Expression of TOX3 protein in human
breast tumors and controls. |
Table II.
Expression of TOX3 protein in human
breast tumors and controls.
| Tumor pathology | Number of patients
(n) | Immunoreactive score
(mean) | P-value |
|---|
| Normal breast
tissues | 25 | 0.88 |
|
| Breast cancer
tissues | 267 | 1.98 | <0.010 |
| TNM grading |
|
|
|
| T1 | 32 | 1.69 |
|
| T2 | 190 | 1.93 |
0.059 |
| T3 | 27 | 2.30 |
0.020 |
| T4 | 18 | 2.56 | <0.010 |
| Lymph node
metastasis |
|
|
|
| N0 | 210 | 1.94 |
|
| N1 | 48 | 2.10 |
0.143 |
| N2 |
8 | 2.63 |
0.006 |
| N3 |
1 | 3.00 |
0.132 |
| TNM stage |
|
|
|
| I | 31 | 1.68 |
|
| II | 209 | 1.97 |
0.030 |
|
III | 24 | 2.54 | <0.010 |
| IV |
3 | 2.67 |
0.049 |
| Table III.TOX3 expression in breast carcinoma
with respect to various pathological characteristics. |
Table III.
TOX3 expression in breast carcinoma
with respect to various pathological characteristics.
|
| TOX3
expression |
|
|
|---|
|
|
|
|
|
|---|
| Tumor
pathology | 1+ [n (%)] | 2+ [n (%)] | 3+ [n (%)] | Total (n) | P-value |
|---|
| TNM grading |
|
|
|
| <0.01 |
| T1 | 14 (43.75) | 14
(43.75) | 4 (12.5) | 32 |
|
| T2 | 51 (26.84) | 101 (53.16) | 38 (20.00) | 190 |
|
| T3 | 4
(14.82) | 11
(40.74) | 12 (44.44) | 27 |
|
| T4 | 0 (0.00) |
8 (44.44) | 10 (55.56) | 18 |
|
| Lymph node
metastasis |
|
|
|
|
0.03 |
| N0 | 56 (26.67) | 111 (52.86) | 43 (20.48) | 210 |
|
| N1 | 11 (22.92) | 21
(43.75) | 16 (33.33) | 48 |
|
| N2 | 0 (0.00) |
3 (37.50) | 5
(62.50) |
8 |
|
| N3 | 0 (0.00) | 0
(0.00) |
1 (100.00) |
1 |
|
| TNM stage |
|
|
|
| <0.01 |
| I | 14 (45.16) | 13
(41.94) | 4
(12.90) | 31 |
|
| II | 54 (25.84) | 108 (51.67) | 47 (22.49) | 209 |
|
|
III | 0 (0.00) | 11
(45.83) | 13 (54.17) | 24 |
|
| IV | 0 (0.00) |
1 (33.33) | 2
(66.67) |
3 |
|
Protein expression of TOX3 in
ER-positive tumor and breast cancer cells
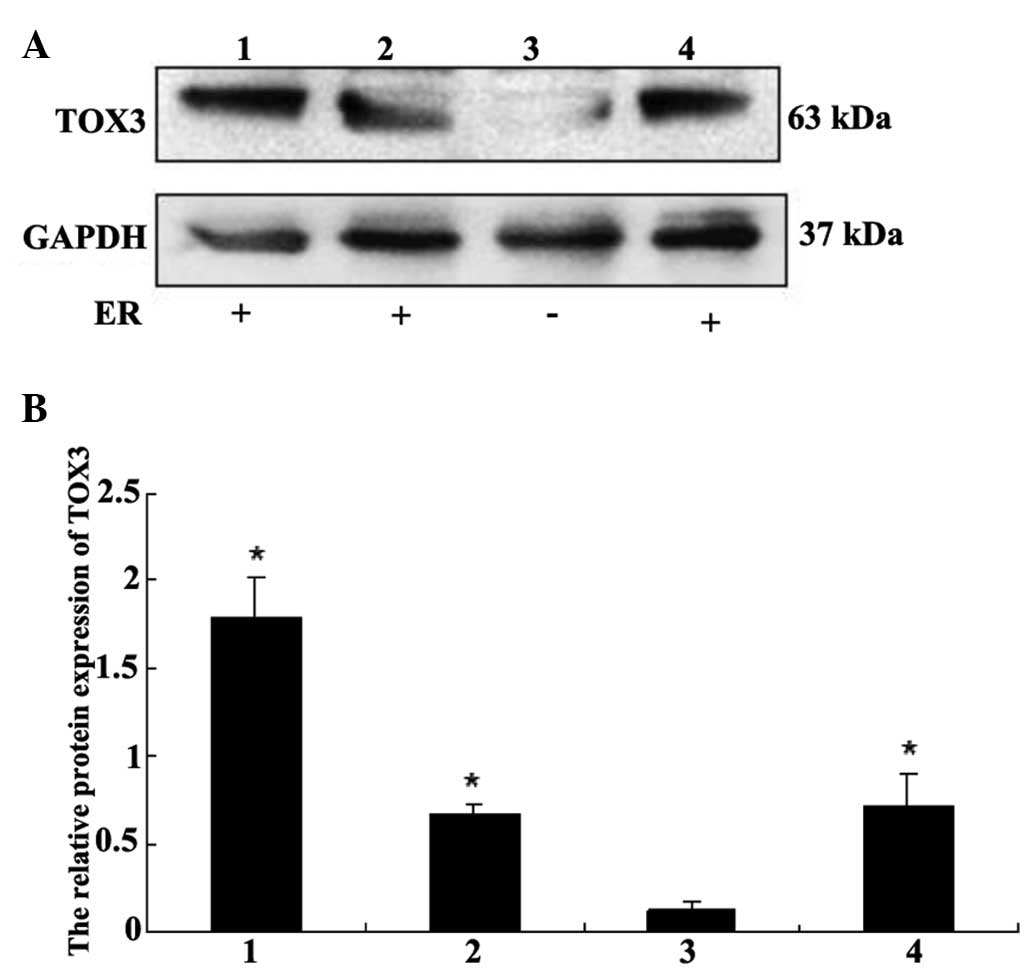
As shown from the immunohistochemical staining
results, the IRS of TOX3 protein expression was higher in
ER-positive tumor than in ER-negative tissue, but there was no
statistical significance (Table IV).
Conversely, a high degree of diversity of TOX3 protein expression
was noted in breast cancer cells, which was intense in ZR-75-1,
MCF-7 and Bcap-37 cell lines compared with MDA-MB-231 breast cell
lines (Fig. 3). Notably, the ER
demonstrated positive expression in ZR-75-1, MCF-7 and Bcap-37 cell
lines, while the MDA-MB-231 cell line had ER-negative
expression.
| Table IV.Protein expression of TOX3 in
estrogen receptor-positive tumors. |
Table IV.
Protein expression of TOX3 in
estrogen receptor-positive tumors.
| Estrogen
receptor | Number of patients
(n) | Immunoreactive
score (mean) | P-value |
|---|
| ER− | 44 | 2.07 |
|
| ER+ | 22 | 2.36 | 0.157 |
Discussion
In recent years, evidence from GWAS has demonstrated
that TOX3 has a strong association with the risk of breast cancer.
Certain studies suggest that the TOX3 gene is associated with
mammographic density (29), and that
breast cancer patients who express high TOX3 mRNA levels have
shorter overall survival and distant metastasis-free survival times
(23). One study reveals an
association between the risk alleles (rs3803662 and rs12443621) and
lower expression of TOX3 mRNA (24),
but another study demonstrates high mRNA expression levels of TOX3
in patients with affected lymph nodes (23). It remains unclear whether these
diverse results were caused by differences in the individuals
involved or solely by methodical differences. Differential TOX3
expression may indicate a diverse progression step during tumor
transformation, and the expression of mRNA may not be in agreement
with that of protein. Therefore, given the lack of data on the
protein expression of TOX3 in malignant breast tumors, in the
present study we assessed the protein expression levels of TOX3 in
breast tumor samples and controls using immunohistochemistry.
Moreover, western blot analysis and qPCR were performed to further
confirm the immunohistochemistry results in three breast tumor
tissues and matched controls in order to demonstrate the clinical
pathological value of TOX3.
Our data suggest that the protein expression levels
of TOX3 in breast carcinoma specimens were significantly higher
than those in controls. Western blot analysis and qPCR results from
fresh tissues further confirmed these results. Moreover,
differential protein expression levels of TOX3 were noted depending
on the TNM stage and the presence of lymph node metastases. TOX3
was upregulated significantly in T3 and T4 stages compared with T1
stage. Compared with patients with no lymph node metastases (N0),
high expression of TOX3 was observed in patients with lymph node
metastases (N2) and stage III disease, but there was no statistical
significance in N3 stage patients. This may be due to the fact that
there was only one case in the N3 stage. Analysis of the literature
in 32 tumor samples also revealed that TOX3 expression was higher
in advanced breast carcinoma (T3 and T4 stages) than in local
breast cancer (T1 and T2 stages) (30), but this study did not demonstrate the
association of TOX3 with lymph node metastases and degree of
differentiation. In addition, another study revealed that high mRNA
expression levels of TOX3 were observed in lymph node patients and
that this was associated with survival (23). These results are compatible with those
of Smid et al, who reported that the TOX3 gene was
implicated in breast carcinoma metastasis to the bone (31). Results from an analysis by Shan et
al demonstrated increased expression of TOX3 in MDA-MB-231
cells, which conferred a stronger capability for invasion and
metastasis. The study also proposed that the TOX3 gene promoted
breast cancer aggressiveness and that TOX3 gene amplification is
significantly associated with a reduction in disease-free and
metastasis-free survival rates (30).
Data from the present study indicate that TOX3 is associated with
clinical and pathological characteristics of the resulting invasive
breast malignancy. It may also be involved in the progression of
breast cancer and be associated with poor prognosis in breast
cancer patients.
In addition, certain researchers suggested that TOX3
was strongly associated with ER-positive cancers by GWAS (6,8,24,25). These
studies are consistent with the finding that, compared with
ER-negative tumors, mRNA expression levels of TOX3 are
significantly higher in ER-positive carcinomas (23). Additionally, there are studies
confirming that the TOX3 transcript was increased in luminal A and
luminal B breast cancer subtypes, but was downregulated in
basal-like breast cancer (32). It is
of note that luminal A and luminal B breast cancers are ER-positive
carcinomas, while the basal-like breast cancer subtype is an
ER-negative tumor (33). However,
there are no data on the protein expression of TOX3 in ER-positive
carcinomas. In the present study, high protein expression levels of
TOX3 were noted in ER-positive cancerous specimens compared with
ER-negative carcinomas, but there was no obvious statistical
significance. This is may be due to the limited number of cases in
the analyses, and it is necessary to increase the number of cases
to further investigate the association between TOX3 expression and
ER expression in breast carcinoma. Conversely, the protein
expression level of TOX3 is significantly reduced in MDA-MB-231
cells, while it is intense in ZR-75-1, MCF-7 and Bcap-37 cells. It
is notable that ER expression is negative in the MDA-MB-231 cell
line, but positive in ZR-75-1, MCF-7 and Bcap-37 cells. This study
demonstrates that TOX3 may be correlated with the risk of breast
cancer, particularly ER-positive cancers. Our study further
supports the link between TOX3 and ER-dependent transcription.
However, it is necessary to further study this subject to confirm
these findings.
In conclusion, our study provides an insight into
the protein expression levels of TOX3 in breast carcinoma and
controls. It reveals that protein expression of TOX3 is upregulated
in breast carcinoma and differs depending on various pathological
characteristics. It may be correlated with the risk of breast
cancer; specifically, TOX3 expression may be correlated with
ER-positive tumors. Our data may lay the foundation for gaining
further insight into the potential regulation and function of TOX3
during breast cancer development.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (no. 81374021 and 81202084), the Key
Project Research Fund of Qiqihar Medical University (no.
QY2013ZD-02) and the Science and Technology Program of Qiqihar
(SFGG-201201).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008. GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dapic V, Carvalho MA and Monteiro AN:
Breast cancer susceptibility and the DNA damage response. Cancer
Control. 12:127–136. 2005.PubMed/NCBI
|
|
3
|
Di LJ, Fernandez AG, De Siervi A, Longo DL
and Gardner K: Transcriptional regulation of BRCA1 expression by a
metabolic switch. Nat Struct Mol Biol. 17:1406–1413. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Prosperi MC, Ingham SL, Howell A, Lalloo
F, Buchan IE and Evans DG: Can multiple SNP testing in BRCA2 and
BRCA1 female carriers be used to improve risk prediction models in
conjunction with clinical assessment? BMC Med Inform Decis Mak.
14:872014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Easton DF, Pooley KA, Dunning AM, Pharoah
PD, Thompson D, Ballinger DG, Struewing JP, Morrison J, Field H,
Luben R, et al: Genome-wide association study identifies novel
breast cancer susceptibility loci. Nature. 447:1087–1093. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stacey SN, Manolescu A, Sulem P, Rafnar T,
Gudmundsson J, Gudjonsson SA, Masson G, Jakobsdottir M, Thorlacius
S, Helgason A, et al: Common variants on chromosomes 2q35 and 16q12
confer susceptibility to estrogen receptor-positive breast cancer.
Nat Genet. 39:865–869. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ruiz-Narváez EA, Rosenberg L, Cozier YC,
Cupples LA, Adams-Campbell LL and Palmer JR: Polymorphisms in the
TOX3/LOC643714 locus and risk of breast cancer in African-American
women. Cancer Epidemiol Biomarkers Prev. 19:1320–1327. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liang J, Chen P, Hu Z and Shen H, Wang F,
Chen L, Li M, Tang J, Wang H and Shen H: Genetic variants in
trinucleotide repeat-containing 9 (TNRC9) are associated with risk
of estrogen receptor positive breast cancer in a Chinese
population. Breast Cancer Res Treat. 124:237–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen MB, Wu XY, Shen W, Wei MX, Li C, Cai
B, Tao GQ and Lu PH: Association between polymorphisms of
trinucleotide repeat containing 9 gene and breast cancer risk:
evidence from 62,005 subjects. Breast Cancer Res Treat.
126:177–183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han W, Woo JH, Yu JH, Lee MJ, Moon HG,
Kang D and Noh DY: Common genetic variants associated with breast
cancer in Korean women and differential susceptibility according to
intrinsic subtype. Cancer Epidemiol Biomarkers Prev. 20:793–798.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Elematore I, Gonzalez-Hormazabal P, Reyes
JM, Blanco R, Bravo T, Peralta O, Gomez F, Waugh E, Margarit S,
Ibañez G, et al: Association of genetic variants at TOX3, 2q35 and
8q24 with the risk of familial and early-onset breast cancer in a
South-American population. Mol Biol Rep. 41:3715–3722. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Long J, Cai Q, Shu XO, Qu S, Li C, Zheng
Y, Gu K, Wang W, Xiang YB, Cheng J, et al: Identification of a
functional genetic variant at 16q12.1 for breast cancer risk:
results from the Asia Breast Cancer Consortium. PLoS Genet.
6:e10010022010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He X, Yao G, Li F, Li M and Yang X:
Risk-association of five SNPs in TOX3/LOC643714 with breast cancer
in southern China. Int J Mol Sci. 15:2130–2141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wilkinson B, Chen JY, Han P, Rufner KM,
Goularte OD and Kaye J: TOX an HMG box protein implicated in the
regulation of thymocyte selection. Nat Immunol. 3:272–280. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Birkenkamp-Demtroder K, Mansilla F,
Dyrskjøt L, Thorsen K, Fristrup N, Brems-Eskildsen AS, Munksgaard
PP, Sørensen KD, Borre M and Ørntoft TF: TOX3 (TNRC9) over
expression in bladder cancer cells decreases cellular proliferation
and triggers an interferon-like response. J Mol Biomark Diagn.
4:22013.
|
|
16
|
O'Flaherty E and Kaye J: TOX defines a
conserved subfamily of HMG-box proteins. BMC Genomics. 4:132003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan SH, Qiu Z and Ghosh A: TOX3 regulates
calcium-dependent transcription in neurons. Proc Natl Acad Sci USA.
106:2909–2914. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dittmer S, Kovacs Z, Yuan SH, Siszler G,
Kögl M, Summer H, Geerts A, Golz S, Shioda T and Methner A: TOX3 is
a neuronal survival factor that induces transcription depending on
the presence of CITED1 or phosphorylated CREB in the
transcriptionally active complex. J Cell Sci. 124:252–260. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xi L, Feber A, Gupta V, Wu M, Bergemann
AD, Landreneau RJ, Litle VR, Pennathur A, Luketich JD and Godfrey
TE: Whole genome exon arrays identify differential expression of
alternatively spliced, cancer-related genes in lung cancer. Nucleic
Acids Res. 36:6535–6547. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tesserma M, Yingling CM, Grimes MJ, Thomas
CL, Liu Y, Leng S, Joste N and Belinsky SA: Differential epigenetic
regulation of TOX subfamily high mobility group box genes in lung
and breast cancers. PLoS One. 7:e348502012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang X, Zhu H, Wu X, Wang M, Gu D, Gong
W, Xu Z, Tan Y, Gong Y, Zhou J, et al: A genetic polymorphism in
TOX3 is associated with survival of gastric cancer in a Chinese
population. PLoS One. 8:e721862013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fasching PA, Pharoah PD, Cox A, Nevanlinna
H, Bojesen SE, Karn T, Broeks A, van Leeuwen FE, van't Veer LJ, Udo
R, et al: The role of genetic breast cancer susceptibility variants
as prognostic factors. Hum Mol Genet. 21:3926–3939. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gudmundsdottir ET, Barkardottir RB, Arason
A, Gunnarsson H, Amundadottir LT, Agnarsson BA, Johannsson OT and
Reynisdottir I: The risk allele of SNP rs3803662 and the mRNA level
of its closest genes TOX3 and LOC643714 predict adverse outcome for
breast cancer patients. BMC Cancer. 12:6212012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Riaz M, Berns EM, Sieuwerts AM,
Ruigrok-Ritstier K, de Weerd V, Groenewoud A, Uitterlinden AG, Look
MP, Klijn JG, Sleijfer S, et al: Correlation of breast cancer
susceptibility loci with patient characteristics, metastasis-free
survival, and mRNA expression of the nearest genes. Breast Cancer
Res Treat. 133:843–851. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Garcia-Closas M and Chanock S: Genetic
susceptibility loci for breast cancer by estrogen receptor status.
Clin Cancer Res. 14:8000–8009. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Reeh M, Bockhorn M, Görgens D, Vieth M,
Hoffmann T, Simon R, Izbicki JR, Sauter G, Schumacher U and Anders
M: Presence of the coxsackievirus and adenovirus receptor (CAR) in
human neoplasms: a multitumour array analysis. Br J Cancer.
109:1848–1858. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lien HC, Lu YS, Cheng AL, Chang WC, Jeng
YM, Kuo YH, Huang CS, Chang KJ and Yao YT: Differential expression
of glucocorticoid receptor in human breast tissues and related
neoplasms. J Pathol. 209:317–327. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Arocho A, Chen B, Ladanyi M and Pan Q:
Validation of the 2-DeltaDeltaCt calculation as an alternate method
of data analysis for quantitative PCR of BCR-ABL P210 transcripts.
Diagn Mol Pathol. 15:56–61. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fernandez-Navarro P, Pita G, Santamariña
C, Moreno MP, Vidal C, Miranda-García J, Ascunce N, Casanova F,
Collado-García F, Herráez B, et al: Association analysis between
breast cancer genetic variants and mammographic density in a large
population-based study (Determinants of Density in Mammographies in
Spain) identifies susceptibility loci in TOX3 gene. Eur J Cancer.
49:474–481. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shan J, Dsouza SP, Bakhru S, Al-Azwani EK,
Ascierto ML, Sastry KS, Bedri S, Kizhakayil D, Aigha II, Malek J,
et al: TNRC9 downregulates BRCA1 expression and promotes breast
cancer aggressiveness. Cancer Res. 73:2840–2849. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Smid M, Wang Y, Klijn JG, Sieuwerts AM,
Zhang Y, Atkins D, Martens JW and Foekens JA: Genes associated with
breast cancer metastatic to bone. J Clin Oncol. 24:2261–2267. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nordard SH, Johansen FE, Alnaes GI, Naume
B, Børresen-Dale AL and Kristensen VN: Genes harbouring
susceptibility SNPs are differentially expressed in the breast
cancer subtypes. Breast Cancer Res. 9:1132007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
O'Brien KM, Cole SR, Engel LS, Bensen JT,
Poole C, Herring AH and Millikan RC: Breast cancer subtypes and
previously established genetic risk factors: a Bayesian approach.
Cancer Epidemiol Biomarkers Prev. 23:84–97. 2014. View Article : Google Scholar : PubMed/NCBI
|