Introduction
Glioblastoma is the most common and devastating
primary malignant intracranial tumor in adults (1). The current standard of care (SOC) for
newly diagnosed glioblastomas is surgical resection followed by
radiotherapy (RT) plus concomitant and adjuvant temozolomide (TMZ)
(2). The prognosis remains relatively
poor with a median overall survival of only 14.6 months, median
progression free survival of 6.9 months and 5 year survival rate of
only 9.8% following diagnosis (2,3). The fast
recurrence and multi-drug resistance are some of the key challenges
in combating brain tumors. Glioma stem cells (GSCs) are considered
to be a major source of relapse and chemoresistance (4), and are therefore an important
therapeutic target for glioblastoma.
Accumulating evidence has also demonstrated that
glioblastoma frequently display hyperactivation of the
phosphatidylinositol-3-kinase (PI3K)-Akt pathway (5–7) and
endogenous Akt kinase activity may be activated in response to
clinically relevant concentrations of TMZ (8,9). Akt
activation is correlated with the increased tumorigenicity,
invasiveness and stemness (6) and
overexpression of an active form of Akt increases glioma cell
resistance to TMZ (5,8). It has demonstrated that cancer stem
cells are preferentially sensitive to an inhibitor of Akt (7,10) and
down-regulation of the PI3K/Akt pathway enhances the cytotoxicity
of TMZ (11). At present, the most
effective drug for the treatment of glioblastoma is TMZ, the
primary path leading to glioma cell death is formation of
O-6-methylguanine and apoptotic signalling triggered by O-6-methyl
G:T mispairs, but apoptotic signalling goes through a step mediated
by AMPK (12). However, there are
obvious disadvantages as described above.
Metformin (MET), a first-line treatment for type-2
diabetes, can reduce cancer incidence (13) and mortality (14), and increases the number of breast
carcinoma patients obtaining complete response to neo-adjuvant
therapy (15). Previous studies have
demonstrated that MET can selectively kill cancer stem cells
(16–19) with minor adverse effects (20). Its mechanism of antiproliferative
action is considered to be activation of AMP-activated protein
kinase (AMPK) (17,18) and/or inhibition of Akt activity
(19,21). On the basis of this evidence, a number
of clinical trials are underway (22,23).
Soritau et al (24) proved
that the tumor cells isolated from patients with high-grade gliomas
who were treated with TMZ and MET exhibited a significant decrease
in proliferation rate when compared to those treated with TMZ
alone. But whether MET may potentiate the cytotoxicity of TMZ for
GSCs is unknown.
In the present study, GSCs isolated from human
glioma cell line U87 and Rat glioma cell line C6, were treated with
TMZ either alone or with MET. The combination index (CI) value was
analyzed and calculated by the Chou-Talalay method (25). The present study demonstrated that MET
acts synergistically with TMZ in inhibiting GSCs proliferation and
expansion, showed a significant apoptosis compared to either agent
alone and reinforced the effect on cell cycle arrest. The present
study provides a rationale for why the combination of MET and TMZ
may improve treatment of patients with glioblastoma.
Materials and methods
Cell and gliosphere culture
The human glioma cell line U87 and Rat glioma cell
line C6 were purchased from Nanjing KGI Biotechnology Company
(Nanjing, China). The cells were cultured in Gibco DMEM media
(Thermo Fisher Scientific, Inc., Waltham, USA), supplemented with
Hyclone 10% fetal bovine serum (Thermo Fisher Scientific, Inc.),
Hyclone Penicillin-Streptomycin (100 U/ml), Hyclone glutamine (2 M)
in a humidified atmosphere of 5% CO2 at 37°C. The cells
were dissociated using 0.25% trypsin (Gibco; Thermo Fisher
Scientific, Inc.) and 0.02% EDTA solution and subcultured once in
3–5 days. To generate gliospheres, U87 and C6 glioblastoma cells
were dissociated from DMEM cultures using trypsin-EDTA solution and
cultured in Gibco Neurobasal medium (NBM) supplemented with Gibco
N2 (1x), B27 (1x), glutaMAX (1x), heparin (2 ug/ml), recombinant
human FGF-basic (b-FGF, 20 ng/ml; PeproTech China, Suzhou, China),
recombinant human epidermal growth factor (EGF, 20 ng/ml;
PeproTech), Hyclone Penicillin-Streptomycin (100 U/ml). The
gliospheres were cultured in 6-well plates in 5% CO2
incubator at 37°C with a medium change every 2–3 days. After
gliospheres formed and reached 100–200 cells/sphere, within 10
days, gliospheres were dissociated by accutase (Sigma-Aldrich, St.
Louis, MO, USA) and reseeded at a ratio of 1:2-3.
GSCs identification
(immunofluorescence staining)
Gliospheres were plated onto poly-L-lysine
(Sigma-Aldrich) coated glass cover slips in DMEM with 10% FBS for 8
h. The Gliospheres were washed with cold PBS (0.01 M), fixed with
4% paraformaldehyde (Gibco; Thermo Fisher Scientific, Inc.) for 30
min, permeabilized with 0.1% Triton X-100 for 15 min and blocked in
5% BSA (Sigma-Aldrich) for 1 h at room temperature. Then the
gliospheres were immunostained with rabbit anti-human polyclonal
CD133 (cat. no. ZA-0426; 1:100; ZSGB-BIO; OriGene Technologies,
Inc., Beijing, China), mouse anti-human monoclonal nestin (cat. no.
ZM-0323; 1:40; ZSGB-BIO; OriGene Technologies, Inc.), mouse
anti-human monoclonal β-tubulin III (cat. no. TA500047; 1:100;
ZSGB-BIO; OriGene Technologies, Inc.) and rabbit anit-human
monoclonal glial fibrillary acidic protein (GFAP; cat. no. ZA-0529;
1:100, ZSGB-BIO; OriGene Technologies, Inc.) antibodies at 4°C
overnight. Subsequent visualization was performed with
fluorescein-conjugated AffiniPure goat anti-rabbit IgG (cat. no.
ZF-0311; 1:100) and tetramethylrhodamine-conjugated AffiniPure goat
anti-mouse IgG (cat. no. ZF-0313; 1:100) secondary antibodies
(ZSGB-BIO; OriGene Technologies, Inc.) for 30 min at room
temperature in darkness, and the nuclei were counterstained with
DAPI (0.5 µg/ml; Institute of Biotechnology, Haimen, China).
For immunostaining of differentiated tumor cells,
gliospheres were transferred to DMEM with 10% FBS for another 7
days and immunocytochemistry was performed as described above. The
fluorescent signals were detected and images were captured with a
fluorescence microscope (Olympus IX51, Olympus Corporation, Tokyo,
Japan).
Gliosphere formation and expansion
assay
To test the effect of two agents (TMZ and/or MET) on
gliosphere formation, after primary sphere formation was observed,
gliospheres were dissociated and plated in 96-well plates
(5×103/ml/well) in NBM with B27, N2, glutaMAX, 2 µg/ml
heparin and 20 ng/ml EGF+bFGF in the absence or presence of TMZ,
MET or TMZ plus MET. Cultures were fed 0.02 ml of NBM every 2 days
and images were captured (x200 magnification) after 7 days using
IX51 Olympus microscope. To determine the gliosphere counts, U87
and C6 gliospheres were dissociated and plated in 96-well plates
(5×103/ml/well) in NBM with B27, N2, glutaMAX, 2 µg/ml
heparin and 20 ng/ml EGF+bFGF in the presence of TMZ, MET or TMZ
plus MET for 7 days and the number of gliospheres counted under
IX51 Olympus microscope.
Combination proliferation assay
The cytotoxic effect of TMZ or/and MET on GSCs was
measured by cell counting kit-8 (CCK-8) assay. Briefly, U87 and C6
gliospheres were cultured in 96-well tissue culture plates
(5×103 per 100 µl per well) in NBM with B27, N2,
glutaMAX, 2 µg/ml heparin and 20 ng/ml EGF+bFGF at 37°C for 24 h,
then different concentrations of TMZ (0–3.2 mmol, Sigma-Aldrich) or
MET (0–160 mmol, Sigma-Aldrich) were added and compared with the
DMSO-treated control. The CCK-8 reagent (10 µl/well) was added at
93 h, and the OD was measured at 450 nm after 96 h using a Synergy
HTX Multi-Mode Microplate Reader (BioTek, Winooski, VT, USA). In
order to determine the combination cytotoxic effect on GSCs,
parallel studies were performed in cells treated with TMZ (0–3.2
mmol) plus MET (0–160 mmol) (1:50, TMZ:MET). The combination index
(CI) value was analyzed by CalcuSyn software (version 2; Biosoft,
Cambridge, UK) and calculated by the Chou-Talalay method (25). CI values <1.0, =1, and >1.0
indicates synergistic interaction (more than additive), summation
(additive), and antagonistic interaction (less than additive),
respectively.
Cell cycle analysis
To determine the effect of TMZ in combination with
MET on cell cycle progression, U87 and C6 gliospheres were cultured
in NBM with B27, N2, glutaMAX, 2 µg/ml heparin and 20 ng/ml
EGF+bFGF in the absence or presence of 0.4 mmol TMZ, 20 mmol MET or
0.4 mmol TMZ plus 20 mmol MET in 5% CO2 incubator at
37°C. After 48 h, the GSCs were dissociated by accutase
(Sigma-Aldrich), fixed and permeabilized with 500 µl of 70% cold
ethanol at −20°C overnight. After washing in PBS, the cells were
treated with 100 µg/ml of RNase (Sigma-Aldrich) and incubated at
37°C for 30 min. Then the cells stained with 100 µg/ml of propidium
iodide (PI, Sigma-Aldrich) at 4°C for another 30 min in darkness.
The percentage of cells at different cell cycle stages (G0/G1, G2/M
and S phase) was determined on the basis of DNA content by flow
cytometer using ModFit LT 3.3 software (Verity Software House,
Inc., Topsham, ME, USA).
Apoptosis assay
To determine the effect of TMZ or/and MET on GSCs
apoptosis, U87 and C6 gliospheres were cultured in NBM with B27,
N2, glutaMAX, 2 µg/ml heparin and 20 ng/ml EGF+bFGF in the absence
or presence of 0.4 mmol TMZ, 20 mmol MET or 0.4 mmol TMZ plus 20
mmol MET in 5% CO2 incubator at 37°C. After 48 h, the
GSCs were collected. Apoptotic and necrotic cell death was analyzed
by double staining with fluorescein-isothiocyanate
(FITC)-conjugated annexin V and PI, in which annexin V bound to the
early and late apoptotic cells with membrane-exposed
phosphatidylserine, while PI labeled only the late
apoptotic/necrotic cells with membrane damage. Staining was
performed using the FITC Annexin V Apoptosis Detection Kit (BD
Pharmingen, San Diego, CA, USA), according to the manufacturer's
instructions. The stained cells were analyzed with a BD FACSAria
III flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). The
numbers of viable (annexin V-/PI-), apoptotic (annexin V+/PI-), and
necrotic (annexin V+/PI+) cells were calculated with the FACSDiva
software, version 6.2 (BD Biosciences).
Western blot analysis
To investigate possible molecular determinants of
MET effects, U87 and C6 gliospheres were treated with TMZ (400
µmol), MET (20 mmol) or TMZ+MET (400 µmol+20 mmol) for 72 h were
collected and lysed in RIPA buffer. Equal amounts of proteins (30
µg) were separated by SDS-PAGE gels and were transferred to a PVDF
membrane (Pierce; Thermo Fisher Scientific, Inc.) and blocked for 2
h in 5% nonfat dry milk in PBS-T. The blots were incubated with
primary antibodies, including rabbit anti-human monoclonal
phospho-Akt (cat. no. 13038P; 1:1,000; Cell Signaling Technology,
Inc., Danvers, MA, USA), rabbit anti-human monoclonal Akt (cat. no.
4691P; 1:1,000; Cell Signaling Technology, Inc.) and mouse
anti-human monoclonal β-actin (cat. no. A1978; 1:1,000;
Sigma-Aldrich), at 4°C for 12 h. The membranes were then washed
with PBS-T and incubated with HRP-conjugated goat anti-rabbit IgG
(cat. no. ZB-2301; 1:5,000) or HRP-conjugated goat anti-mouse IgG
(cat. no. ZB-2305; 1:5,000) secondary antibodies (ZSGB-BIO; OriGene
Technologies, Inc.) for 2 h. After subsequent washes, specific
signals were showed using the Pierce ECL Western Blotting Substrate
(Thermo Fisher Scientific, Inc.) and detected by the western blot
automated chemiluminescence image analysis system (Tanon5200;
Shanghai-Day Technology Co., Ltd., Shanghai, China).
Statistical analysis
The experiments were performed ≥3 times and data
were expressed as the mean ± standard deviation. Statistical
comparisons were assessed using the Student's t test. Statistical
analysis was performed with SPSS software, version 19.0 (IBM
SPSS< Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Isolation, characterization and
differentiation of GSCs isolated from U87 and C6 glioma cells
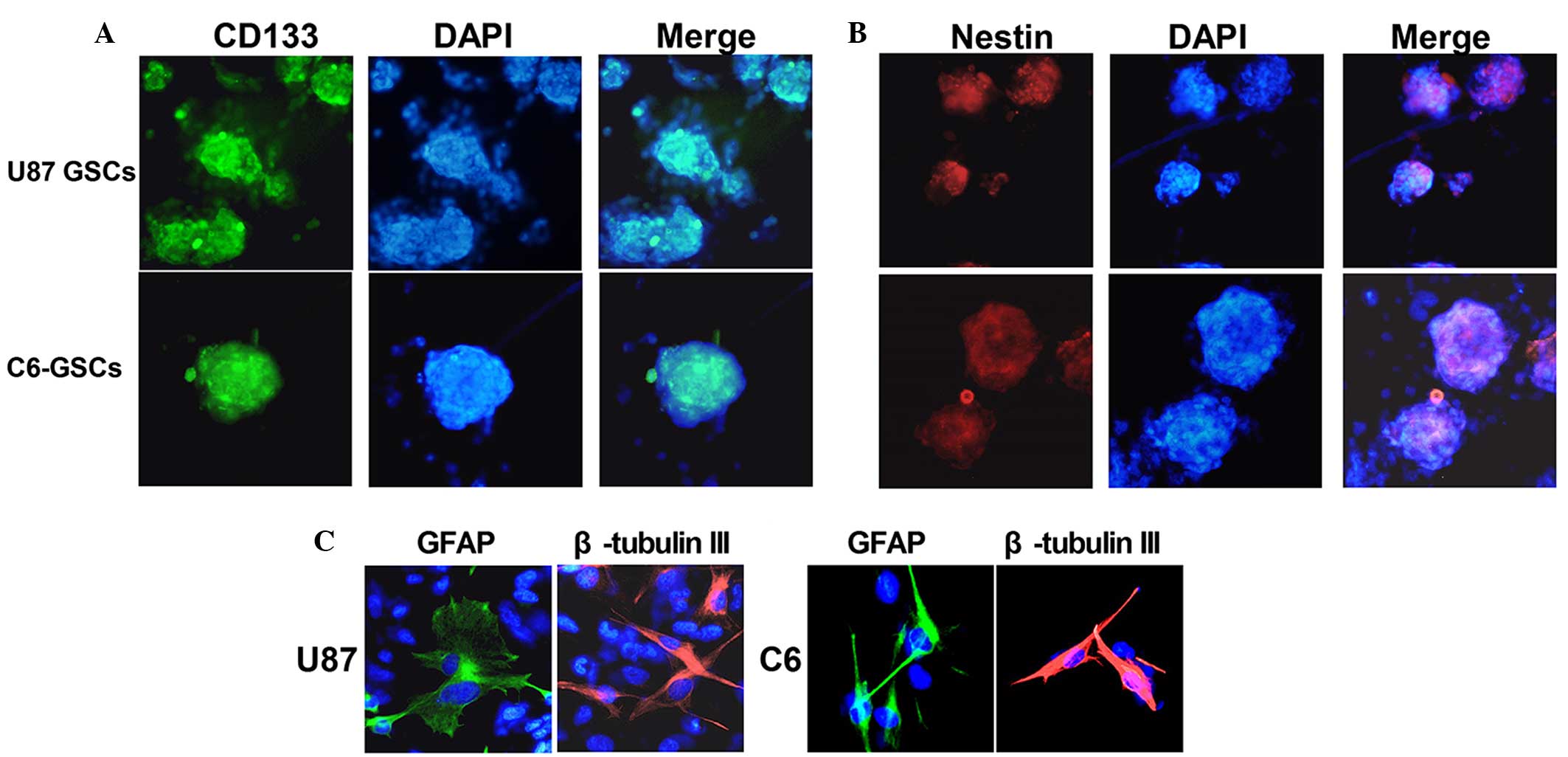
U87 and C6 gliospheres cultures were characterized
for recognized GSC signatures: Self-renewal, neural stem cell
marker expression and differentiation. The cells within the sphere
were positive for neural stem cell markers CD133 (Fig. 1A) and nestin (Fig. 1B), and lack of immunoreactivity for
markers of differentiated neural cell types such as GFAP for
astrocytes and β-tubulin III for neurons. The assay of
multi-lineage differentiation capacity of cells within the sphere
was demonstrated by culturing the cells in differentiation-inducing
culture medium (DMEN+10% FBS) for 7 days. These cells lost
expression of CD133 and nestin when subjected to differentiating
conditions, showed typical morphological differentiation towards
neuronal and astrocytic lineages, identified as β-tubulin-III
positive for neurons and GFAP positive for astrocytes (Fig. 1C). The self-renewing capacity of the
tumor spheres was assayed by dissociation of primary tumor spheres.
When the self-renewal capacity was compared among tumor subtypes at
a plating density of 5×103 cells/well, U87 were observed
to generate a greater mean number of secondary tumor spheres
(151±5) compared with C6 (123±4) (Fig.
2).
TMZ plus MET contribute more
effectively to inhibit gliospheres formation and expansion
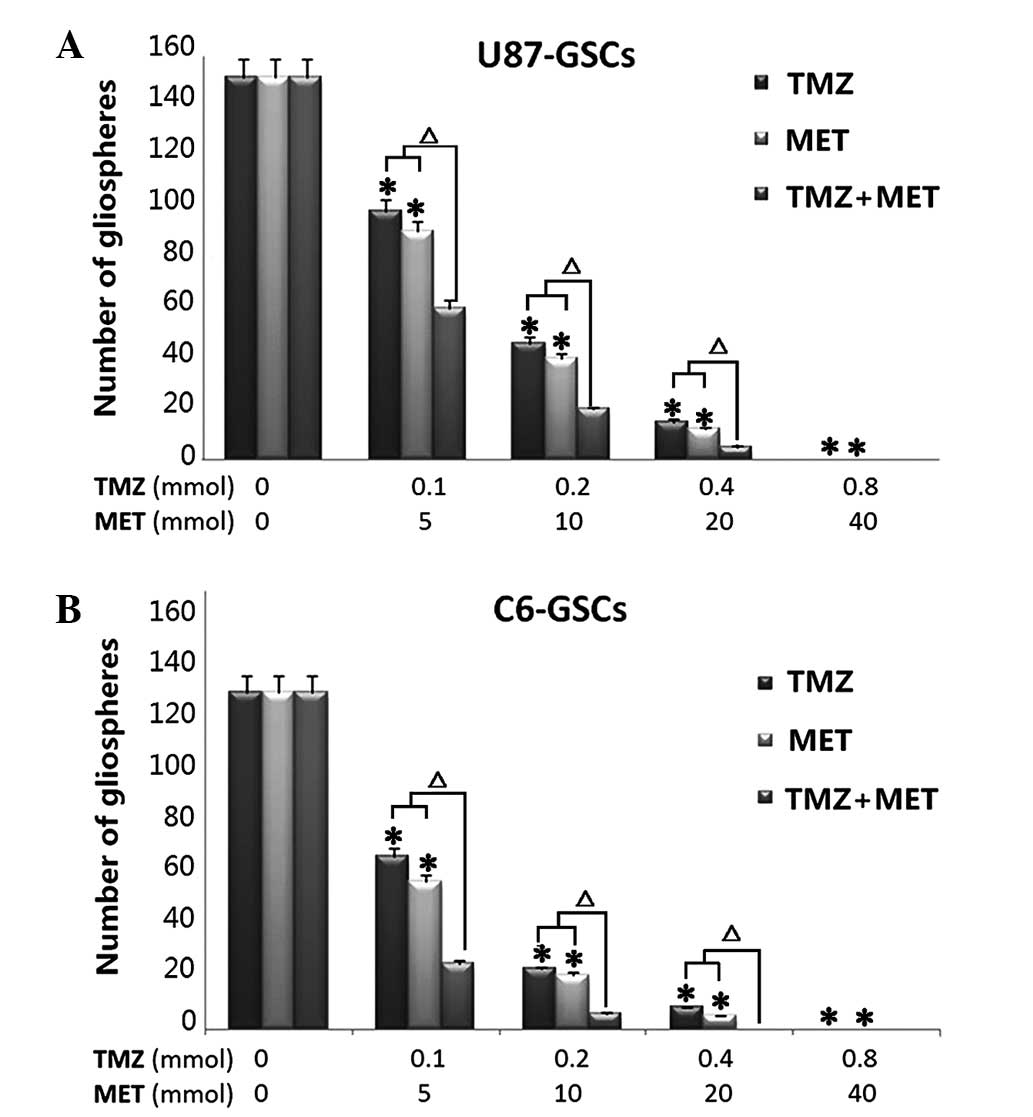
To determine whether TMZ plus MET is more effective
in the inhibition of U87 and C6 gliosphere formation and expansion
compared with either drug alone, culture conditions were
established to generate gliospheres in vitro. U87 and C6
glioma cells were cultured in NBM with B27, N2, glutaMAX, 2 ug/ml
heparin and 20 ng/ml EGF+bFGF, promotes gliosphere formation in 5–7
days and increases in size in 7–10 days. In vitro treatment
with TMZ or MET resulted in a marked reduction in the number and
size of gliospheres generated from U87 and C6 cells (P<0.05).
Notably, when the cells were treated with MTZ plus MET, a
significant decrease was observed in the number and size of
gliospheres compare to either agent alone (P<0.01). Further
analyses demonstrated that in vitro treatment with TMZ, MET
or TMZ plus MET resulted in a dose-dependent decrease in the number
and size of gliospheres (Fig. 2).
These results indicate that TMZ plus MET combination treatment is
more effective at inhibiting U87 and C6 gliosphere formation and
expansion compared with single agent treatment.
TMZ plus MET acts synergistically in
inhibiting GSCs proliferation
In order to determine whether the MET, which is
currently in clinical trials for cancer treatment, augments the
cell proliferation inhibitory effects of TMZ in GSCs, U87 and C6
GSCs were plated with 0–3.2 mmol TMZ either alone or with 0–160
mmol MET for 72 h and the cells were assessed using the CCK-8
assay. Both U87-GSCs and C6-GSCs exhibited reduced proliferation
levels as a dose-dependent response to TMZ and MET. The combination
of TMZ and MET (1:50, TMZ:MET) resulted in a significant shift in
the proliferation inhibition curve compared with treatment with
either drug alone(Fig. 3). The
statistical combination index (CI) was determined for the dual
therapy to determine whether combination therapy was synergistic,
additive, or antagonistic. As shown in Fig. 3 and Table
I, MET acted synergistically (CI<1.0) with TMZ to inhibit
U87-GSCs and C6-GSCs growth at all combination doses tested. As the
concentration of TMZ in combination with MET was increased, the
trend in CI value was reduced, suggesting that increasing the
dosage of TMZ and MET increases the synergistic effect.
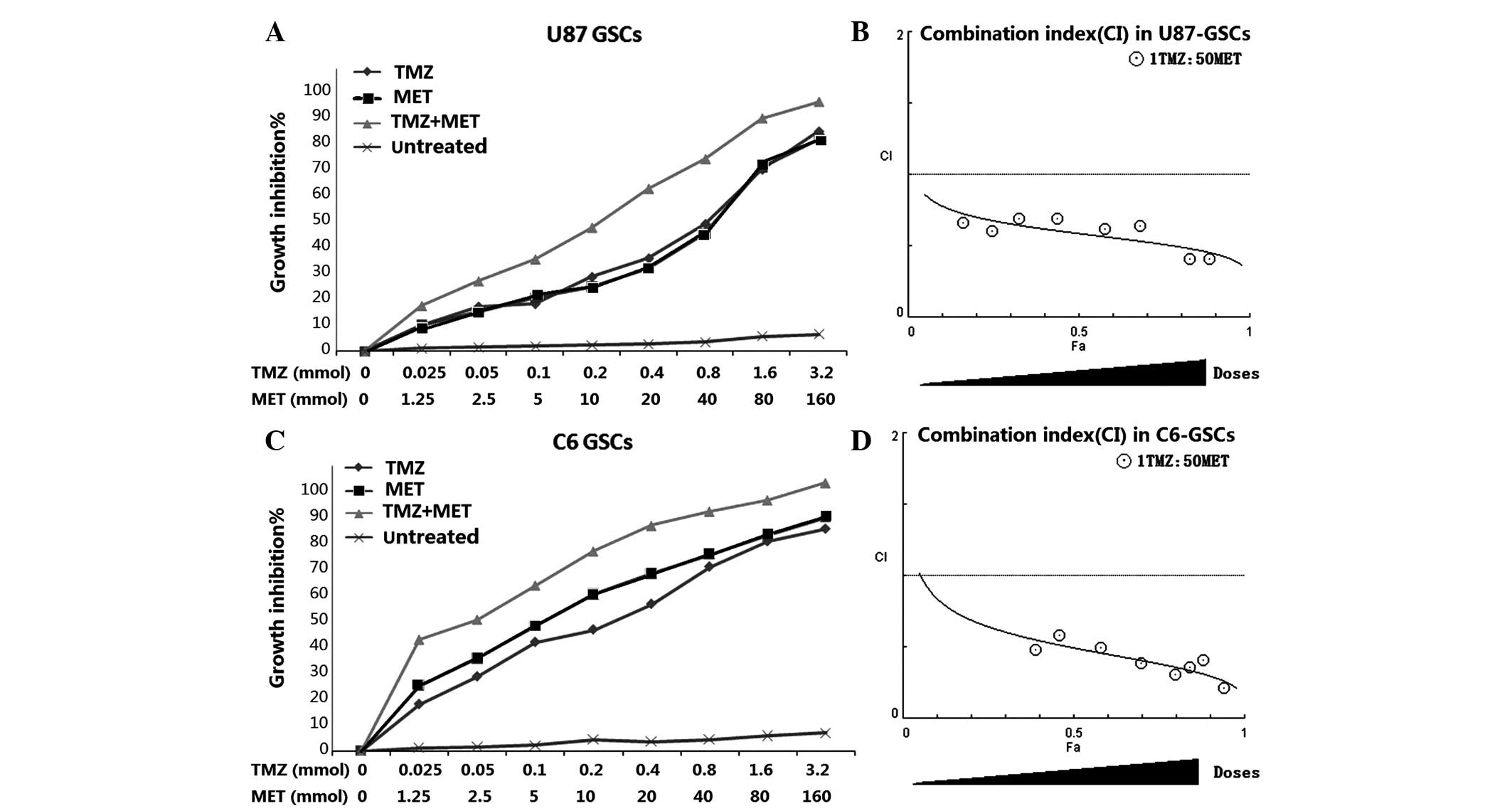 | Figure 3.MET augments the effects of TMZ in
GSCs. (A) U87-GSCs and (C) C6-GSCs were treated with TMZ alone
(0–160 mmol), MET (0–3.2 mmol) or the combination of TMZ and MET
(1:50, TMZ:MET). Calcusyn software was used to ascertain the
combination index (CI) value for each combination ratio to
determine whether combination treatments have an additive or
synergistic effect on (B) U87-GSCs or (D) C6-GSCs. CI values
<1.0, =1, and >1.0 indicates synergistic interaction (more
than additive), summation (additive), and antagonistic interaction
(less than additive), respectively. Results are representative of 3
independent experiments. GSCs, glioma stem cells; TMZ, temozolomid;
MET, metformin; CI, combination index. |
 | Table I.Combination treatments with TMZ and
MET synergistically inhibit cell growth in U87-GSCs and
C6-GSCs. |
Table I.
Combination treatments with TMZ and
MET synergistically inhibit cell growth in U87-GSCs and
C6-GSCs.
|
| TMZ | MET | Combination |
|
|---|
|
|
|
|
|
|
|---|
| Cell line | Dose (mmol) | Growth inhibition
(%) | Dose (mmol) | Growth inhibition
(%) | Ratio | Growth inhibition
(%) | CI value |
|---|
| U87-GSCs |
0.05 | 15.9±0.83 | 2.5 | 14.0±0.75 | 1:50 | 24.8±0.87 | 0.602 |
|
| 0.1 | 16.9±1.22 | 5 | 18.9±1.24 | 1:50 | 32.6±2.65 | 0.691 |
|
| 0.2 | 26.4±1.15 | 10 | 22.7±1.32 | 1:50 | 43.8±1.39 | 0.693 |
|
| 0.4 | 32.8±1.85 | 20 | 29.4±1.58 | 1:50 | 57.6±2.12 | 0.619 |
|
| 0.8 | 45.2±3.21 | 40 | 41.6±2.01 | 1:50 | 68.1±1.85 | 0.643 |
| C6-GSCs |
0.05 | 26.1±0.95 | 2.5 | 32.5±0.98 | 1:50 | 46.3±2.34 | 0.582 |
|
| 0.1 | 38.2±1.30 | 5 | 44.2±2.09 | 1:50 | 58.2±1.08 | 0.493 |
|
| 0.2 | 42.6±1.59 | 10 | 55.1±2.32 | 1:50 | 70.0±3.05 | 0.388 |
|
| 0.4 | 51.6±2.03 | 20 | 62.3±1.68 | 1:50 | 79.7±2.71 | 0.308 |
|
| 0.8 | 64.7±2.08 | 40 | 69.0±2.41 | 1:50 | 84.2±3.02 | 0.358 |
TMZ and/or MET induced cell cycle
arrest in GSCs
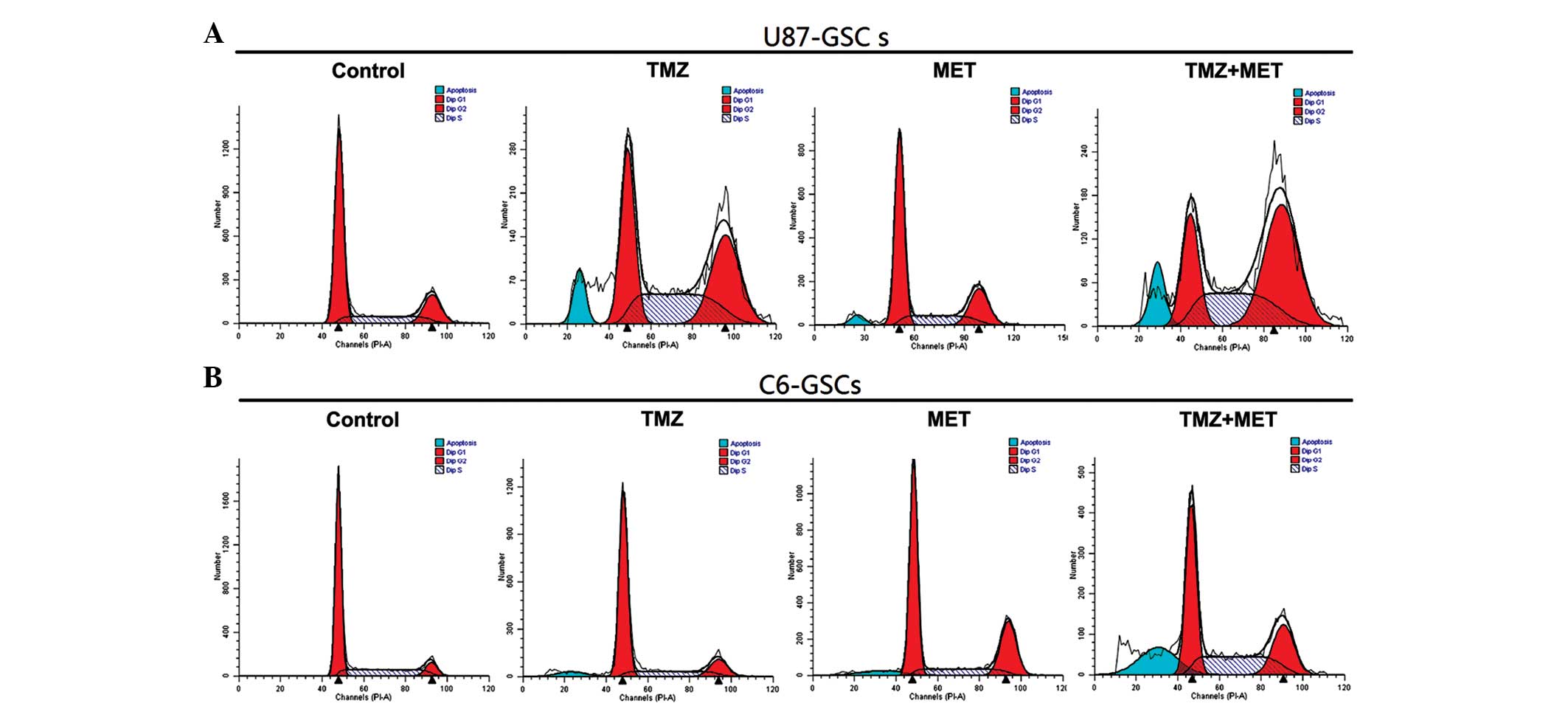
Whether the inhibition of proliferation by TMZ or
MET was due to cell cycle arrest or apoptosis in GSCs was
investigated using flow cytometry. U87-GSCs and C6-GSCs were
treated with TMZ (0.4 mmol), MET (20 mmol) or TMZ (0.4 mmol) plus
MET (20 mmol) for 48 h and the cell cycle phase was determined. As
shown in Fig. 4 and Table II, U87-GSCs cultured in NBM with B27,
N2, glutaMAX, heparin and EGF+bFGF showed 62.79% G0/G1 cells, 18.3%
G2/M cells and 18.91% S-phase cells, addition of TMZ or MET
resulted in a reduction in G0/G1 cells (45.19 and 59.2%,
respectively), a marked increase in G2/M cells (34.49 and 22.19%,
respectively) and an increase in S-phase cells (20.32 and 18.61%,
respectively). However, when U87-GSCs were treated with TMZ plus
MET, a greater reduction in G0/G1 cells (22.67%; P<0.05), and a
greater increase in G2/M and S-phase cells was observed (48.89 and
28.43%, respectively; P<0.05). This result indicates that the
U87-GSC cells were arrested at the G2/M phase by TMZ or MET
treatment, but TMZ plus MET induced cell cycle arrest at both the
G2/M phase and S-phase; the effect was markedly better than
treatment with a single agent, particularly for the G2/M phase
arrest. C6-GSCs, cultured in NBM with B27, N2, glutaMAX, heparin
and EGF+bFGF, were analyzed by flow cytometry and the result
demonstrated that 66.43% cells were in G0/G1, 8.53% were in G2/M
and 25.04% were in S-phase. The addition of TMZ or MET resulted in
a slight decrease in the percentage of G0/G1 cells (60.06 and
56.34%, respectively), an increase in the percentage of G2/M cells
(22.64 and 25.02%, respectively) and a slight decrease in the
percentage of S-phase cells (17.30 and 18.65%, respectively). When
C6-GSCs were treated with TMZ plus MET resulted in a greater
reduction in the percentage of G0/G1 cells (43.09%; P<0.05) and
a marked increase in the percentage of cells in S-phase (31.92%;
P<0.05). These results indicated that the C6-GSCs cycles were
arrested at S phase following treatment with TMZ plus MET; however,
little difference was observed in the percentage of C6-GSCs in G2/M
arrest when treated with TMZ plus MET compared with either single
agent. These results suggest that TMZ plus MET treatment regulates
cell cycle progression of GSCs in culture and using the agents
together enhances the effects.
| Table II.Modulation of cell cycle and
apoptosis by TMZ, MET or TMZ plus MET. |
Table II.
Modulation of cell cycle and
apoptosis by TMZ, MET or TMZ plus MET.
| Cell line | Sub-group | Mean (%) Sub G0
phase | Mean (%) G0/G1
phase | Mean (%) S
phase | Mean (%) G2/M
phase |
|---|
| U87-GSGS | Control | 0.08±0.02 | 62.79±3.10 | 18.91±2.30 | 18.30±1.65 |
|
| TMZ | 6.22±0.20 | 45.19±1.28 | 20.32±1.41 | 34.49±1.69 |
|
| MET | 5.38±0.03 | 59.20±2.36 | 18.61±1.36 | 22.19±2.52 |
|
| TMZ+MET |
14.77±1.09a |
22.67±3.21a |
28.43±2.84a |
48.89±3.21a |
| C6-GSCs | Control | 0.05±0.01 | 66.43±2.89 | 25.04±1.05 | 8.53±0.39 |
|
| TMZ | 5.37±0.05 | 60.06±2.03 | 17.30±0.56 | 22.64±0.58 |
|
| MET | 6.28±0.24 | 56.34±1.38 | 18.65±0.47 | 25.02±0.12 |
|
| TMZ+MET |
19.61±0.31a |
43.09±0.45a |
31.92±0.90a | 24.99±0.14 |
TMZ plus MET significantly induced
apoptosis in GSCs
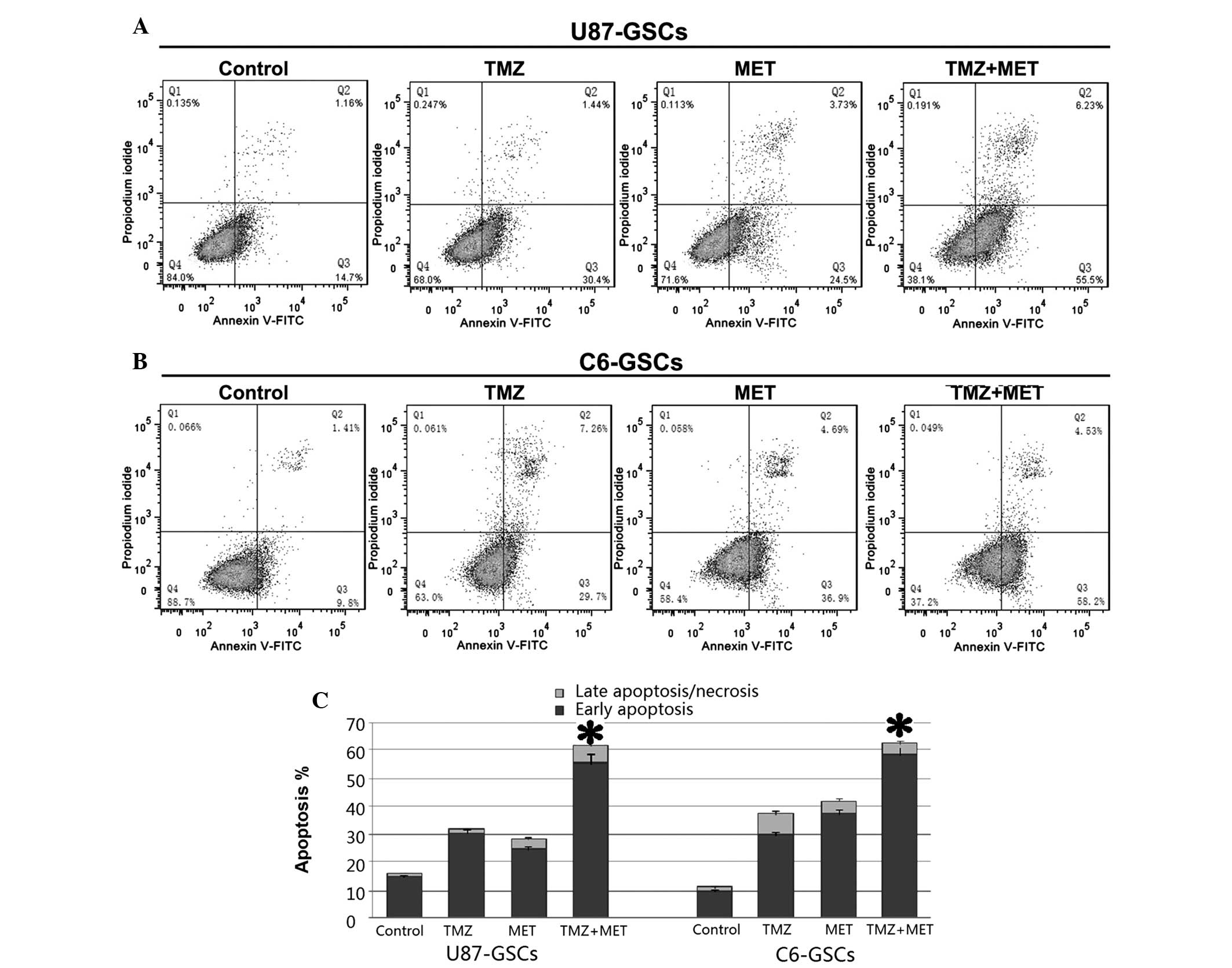
The present study also aimed to determine whether
the TMZ or MET-induced reduction in cell viability is accompanied
by cell apoptosis. Annexin V and PI staining and flow cytometry was
used to detect apoptosis in U87-GSCs and C6-GSCs treated with TMZ
(0.4 mmol), MET (20 mmol) or TMZ (0.4 mmol) plus MET (20 mmol) for
48 h. As shown in Fig. 5, U87-GSCs
and C6-GSCs cultured without TMZ or MET exhibited low apoptotic
cell death. Treatment with TMZ or MET induced cell apoptosis
(P<0.05); however, treatment with TMZ plus MET resulted in a
significant increase in apoptotic cells compared with single agents
(P<0.05). In U87-GSCs, the percentage of apoptotic cells in the
control group was 15.86%, while those of TMZ and MET groups were
31.84 and 28.23%, respectively, and was significantly increased to
61.73% in TMZ plus MET group (P<0.05). In C6-GSCs, the
percentage of apoptotic cells in the control group was 11.21%, and
those of the TMZ and MET groups were 36.96% and 41.59%,
respectively; and was significantly increased to 62.73% in TMZ plus
MET group (P<0.05). The above results demonstrated that TMZ or
MET significantly induced apoptosis in GSCs compared with the
control groups and MET treatment enhances TMZ-induced
apoptosis.
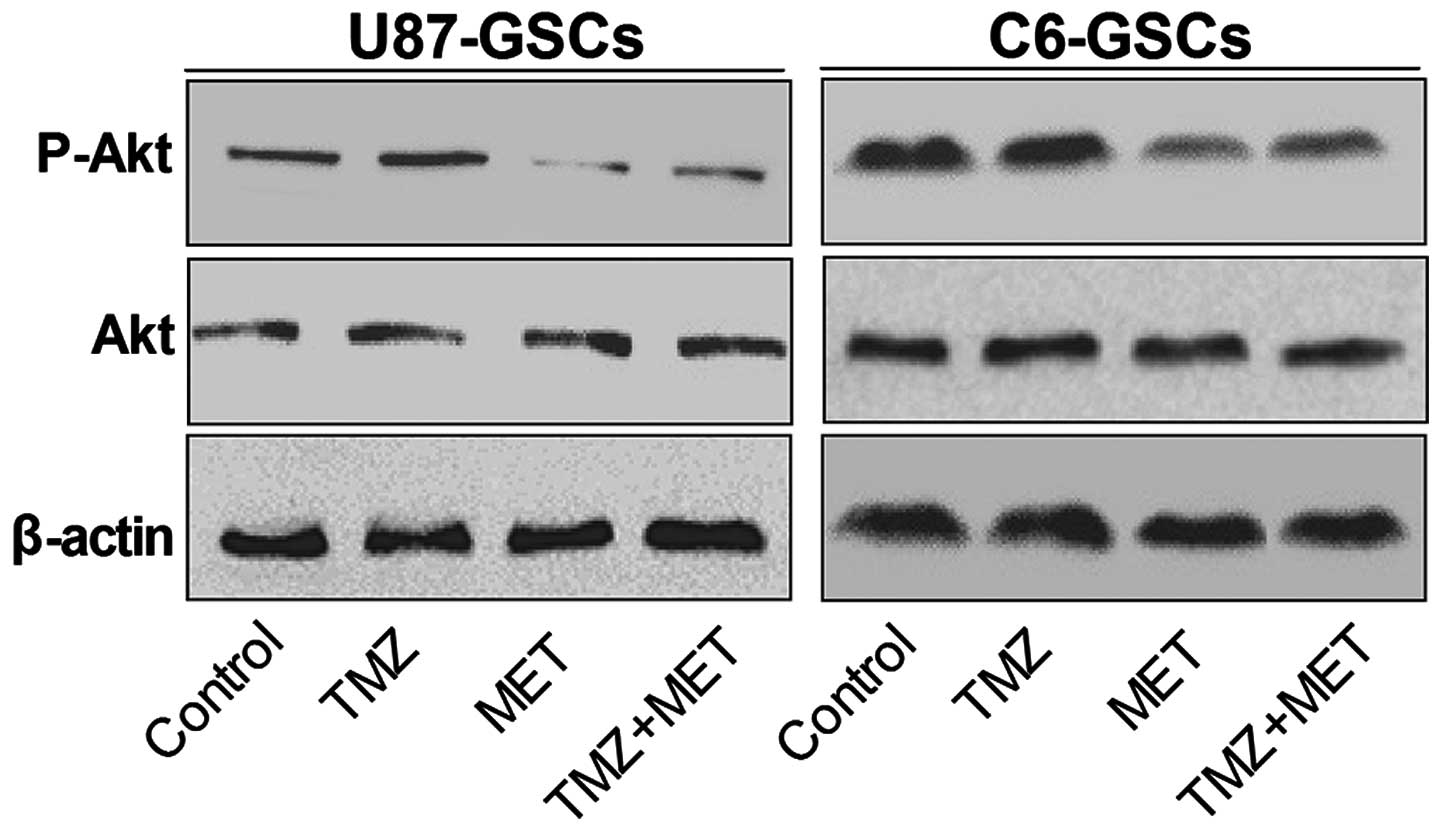
MET potentiates the cytotoxicity of
TMZ for GSCs by inhibition of Akt activation
Fig. 6 demonstrates
that when GSCs were treated with TMZ alone for 72 h, Akt
phosphorylation is slightly enhanced. However, Akt phosphorylation
was inhibited by MET, suggesting that Akt inhibition is required
for MET antiproliferative activity in GSCs. Notably, the
phosphorylated Akt (P-Akt) levels were markedly down-regulated
compared with the control group and TMZ alone groups compared with
treatment with TMZ and MET in combination. These results indicate
that the potential mechanism of MET enhancing TMZ cytotoxicity may
involve down-regulation of P-Akt.
Discussion
Previous studies have indicated that glioblastomas
originate from a pool of stem-like cells that share properties in
common with neuronal stem cells. GSCs are a small population of
cells that have self-renewal, transplantation, recurrence
properties and multidrug resistance (26). The identification of stem-like cells
in tumors and the proposed role for this subpopulation in drug
resistance opened the search for targeted molecular therapies and
novel drugs able to deplete this tumor cell subpopulation, either
by activating differentiation or through selective toxicity that
causes loss of self-renewal (19).
However, in medicine, it is usually difficult to get agents with
high target specificity; only a few of compounds have showed
selective toxicity for cancer stem cells (27,28).
However, certain drugs already used in the clinic that may have
encouraging results, such as MET, may selectively kill cancer stem
cells (16–19) with minor adverse events (20).
To the best of our knowledge, the present report
demonstrates for the first time the ability of MET to function
synergistically with TMZ to reduce U87-GSCs and C6-GSCs
proliferation, although it has been proved previously that MET
improves the survival of cisplatin-treated tumour cells (29).
TMZ is currently the most effective drug for the
treatment of glioblastoma and one mechanism of TMZ-induced
apoptosis involves an obligatory AMP-activated protein kinase
(AMPK) activation step (12);
however, this increases the activity of Akt which increases glioma
cell resistance to TMZ (5,8) and may increase the tumorigenicity,
invasiveness and stemness of tumor cells (6). The anticancer mechanism of MET is
considered to be activation of AMPK (17,18) or
inhibition of the PI3K-Akt pathway (19,21).
Although the present study demonstrated that MET potentiates the
cytotoxicity of TMZ for GSCs and has a synergistic effect, whether
this is achieved through increased AMPK activity and/or inhibition
of AKT activity in GSCs remains unknown.
The potential for MET to target GBM proliferation
was proposed due to the hypothesized synergy with TMZ on AMPK
activation (17); the present authors
are sceptical of this hypothesis and it has not been confirmed.
However, certain studies support this hypothesis. Liu et al
(30), suggested that MET's growth
inhibition in vitro does not require AMPK. Instead, AMPK
activation may be a response mechanism to counter stress induced by
anticancer agents (30). It is
possible that the growth-inhibiting effects are a combination of
the AMPK-independent mTOR-dependent cellular and systemic effects
of MET (30). Gritti et al
(16) determined that MET exhibits
its selective antiproliferative effects in human GSCs via
inhibition of the CLIC1-mediated ion current, although this
requires further studies to confirm this observation.
It has been proposed that the Akt pathway represents
a novel target for the sensitization of Akt overexpression gliomas
to chemotherapeutic methylating agents such as TMZ (5), due to the ability of Akt overexpression
to suppress TMZ-induced mitotic catastrophe as well as TMZ-induced
senescence (5). It has been proved
that TMZ combined with the PI3K inhibitor LY294002 inhibits
melanoma cell growth, survival and invasion (31); and similar results were observed in
glioma cells (11). Akt activation
has been reported to suppress activation of the G2 checkpoint in
human colon carcinoma cells exposed to radiation (32), Akt activation has also been proven to
suppresses TMZ-induced G2 arrest (5).
However, in our trial, MET, through the inhibition of Akt
activation, selectively killed cancer stem cells and acted
synergetic with TMZ, enhancing TMZ-induced G2 arrest for U87-GSCs;
however, in C6-GSCs, they just reinforced each other and induced
cell cycle arrest at S phase. Based on the experimental results of
the present study, MET may enhance the cytotoxicity of TMZ by
down-regulation of the PI3K/Akt pathway, these results warrant
further in vivo exploration.
Although the present study presents evidence that
MET acts synergistically with TMZ in inhibiting GSCs proliferation
and generating the highest apoptotic rates when compared with
either drug alone, but the exact mechanism of action remains
unclear. Soritau et al (24)
reported that tumor cells isolated from patients with high-grade
gliomas who were treated with TMZ and MET exhibited a significant
decrease in proliferation rate when compared to those treated with
TMZ alone. In conclusion, future trials are required to explore the
detailed mechanisms.
Acknowledgements
The present study was supported by the Young
Scientists Fund of the National Natural Sciene Foundation of China
(grant no. 21401072).
References
|
1
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hermann PC, Bhaskar S, Cioffi M and
Heeschen C: Cancer stem cells in solid tumors. Semin Cancer Biol.
20:77–84. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hirose Y, Katayama M, Mirzoeva OK, Berger
MS and Pieper RO: Akt activation suppresses Chk2-mediated,
methylating agent-induced G2 arrest and protects from
temozolomide-induced mitotic catastrophe and cellular senescence.
Cancer Res. 65:4861–4869. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Molina JR, Hayashi Y, Stephens C and
Georgescu MM: Invasive glioblastoma cells acquire stemness and
increased Akt activation. Neoplasia. 12:453–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Eyler CE, Foo WC, LaFiura KM, McLendon RE,
Hjelmeland AB and Rich JN: Brain cancer stem cells display
preferential sensitivity to Akt inhibition. Stem Cells.
26:3027–3036. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Caporali S, Levati L, Starace G, Ragone G,
Bonmassar E, Alvino E and D'Atri S: AKT is activated in an
ataxia-telangiectasia and Rad3-related-dependent manner in response
to temozolomide and confers protection against drug-induced cell
growth inhibition. Mol Pharmacol. 74:173–183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
De Salvo M, Maresca G, D'Agnano I,
Marchese R, Stigliano A, Gagliassi R, Brunetti E, Raza GH, De Paula
U and Bucci B: Temozolomide induced c-Myc-mediated apoptosis via
Akt signalling in MGMT expressing glioblastoma cells. Int J Radiat
Biol. 87:518–533. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gallia GL, Tyler BM, Hann CL, Siu IM,
Giranda VL, Vescovi AL, Brem H and Riggins GJ: Inhibition of Akt
inhibits growth of glioblastoma and glioblastoma stem-like cells.
Mol Cancer Ther. 8:386–393. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen L, Han L, Shi Z, Zhang K, Liu Y,
Zheng Y, Jiang T, Pu P, Jiang C and Kang C: LY294002 enhances
cytotoxicity of temozolomide in glioma by down-regulation of the
PI3K/Akt pathway. Mol Med Rep. 5:575–579. 2012.PubMed/NCBI
|
|
12
|
Zhang WB, Wang Z, Shu F, Jin YH, Liu HY,
Wang QJ and Yang Y: Activation of AMP-activated protein kinase by
temozolomide contributes to apoptosis in glioblastoma cells via p53
activation and mTORC1 inhibition. J Biol Chem. 285:40461–40471.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Evans JM, Donnelly LA, Emslie-Smith AM,
Alessi DR and Morris AD: Metformin and reduced risk of cancer in
diabetic patients. BMJ. 330:1304–1305. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bowker SL, Majumdar SR, Veugelers P and
Johnson JA: Increased cancer-related mortality for patients with
type 2 diabetes who use sulfonylureas or insulin. Diabetes Care.
29:254–258. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiralerspong S, Palla SL, Giordano SH,
Meric-Bernstam F, Liedtke C, Barnett CM, Hsu L, Hung MC, Hortobagyi
GN and Gonzalez-Angulo AM: Metformin and pathologic complete
responses to neoadjuvant chemotherapy in diabetic patients with
breast cancer. J Clin Oncol. 27:3297–3302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gritti M, Wurth R, Angelini M, Barbieri F,
Peretti M, Pizzi E, Pattarozzi A, Carra E, Sirito R, Daga A, et al:
Metformin repositioning as antitumoral agent: Selective
antiproliferative effects in human glioblastoma stem cells, via
inhibition of CLIC1-mediated ion current. Oncotarget.
5:11252–11268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kast RE, Karpel-Massler G and Halatsch ME:
Can the therapeutic effects of temozolomide be potentiated by
stimulating AMP-activated protein kinase with olanzepine and
metformin? Br J Pharmacol. 164:1393–1396. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sato A, Sunayama J, Okada M, Watanabe E,
Seino S, Shibuya K, Suzuki K, Narita Y, Shibui S, Kayama T and
Kitanaka C: Glioma-initiating cell elimination by metformin
activation of FOXO3 via AMPK. Stem cells Transl Med. 1:811–824.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wurth R, Pattarozzi A, Gatti M, Bajetto A,
Corsaro A, Parodi A, Sirito R, Massollo M, Marini C, Zona G, et al:
Metformin selectively affects human glioblastoma tumor-initiating
cell viability: A role for metformin-induced inhibition of Akt.
Cell Cycle. 12:145–156. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bolen S, Feldman L, Vassy J, Wilson L, Yeh
HC, Marinopoulos S, Wiley C, Selvin E, Wilson R, Bass EB, et al:
Systematic review: Comparative effectiveness and safety of oral
medications for type 2 diabetes mellitus. Ann Intern Med.
147:386–399. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zakikhani M, Blouin MJ, Piura E and Pollak
MN: Metformin and rapamycin have distinct effects on the AKT
pathway and proliferation in breast cancer cells. Breast Cancer
Res. 123:271–279. 2010. View Article : Google Scholar
|
|
22
|
Rattan R, Ali Fehmi R and Munkarah A:
Metformin: An emerging new therapeutic option for targeting cancer
stem cells and metastasis. J Oncol. 2012:9281272012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Quinn BJ, Kitagawa H, Memmott RM, Gills JJ
and Dennis PA: Repositioning metformin for cancer prevention and
treatment. Trends Endocrinol Metab. 24:469–480. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Soritau O, Tomuleasa C, Aldea M, Petrushev
B, Susman S, Gheban D, Ioani H, Cosis A, Brie I, Irimie A, et al:
Metformin plus temozolomide-based chemotherapy as adjuvant
treatment for WHO grade III and IV malignant gliomas. J BUON.
16:282–289. 2011.PubMed/NCBI
|
|
25
|
Chou TC: Drug combination studies and
their synergy quantification using the Chou-Talalay method. Cancer
Res. 70:440–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
27
|
Sachlos E, Risueño RM, Laronde S,
Shapovalova Z, Lee JH, Russell J, Malig M, McNicol JD, Fiebig-Comyn
A, Graham M, et al: Identification of drugs including a dopamine
receptor antagonist that selectively target cancer stem cells.
Cell. 149:1284–1297. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gupta PB, Onder TT, Jiang G, Tao K,
Kuperwasser C, Weinberg RA and Lander ES: Identification of
selective inhibitors of cancer stem cells by high-throughput
screening. Cell. 138:645–659. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Janjetovic K, Vucicevic L, Misirkic M,
Vilimanovich U, Tovilovic G, Zogovic N, Nikolic Z, Jovanovic S,
Bumbasirevic V, Trajkovic V, et al: Metformin reduces
cisplatin-mediated apoptotic death of cancer cells through
AMPK-independent activation of Akt. Eur J Pharmacol. 651:41–50.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu X, Chhipa RR, Pooya S, Wortman M,
Yachyshin S, Chow LM, Kumar A, Zhou X, Sun Y, Quinn B, et al:
Discrete mechanisms of mTOR and cell cycle regulation by AMPK
agonists independent of AMPK. Proc Natl Acad Sci USA.
111:E435–E444. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Meier FE, Lasithiotakis K, Schittek B,
Sinnberg T and Garbe C: Temozolomide combined with the PI3K
inhibitor LY294002 or the mTOR inhibitor rapamycin inhibits
melanoma cell growth, survival and invasion. J Clin Oncol. 25(Suppl
18): 85592007.
|
|
32
|
Kandel ES, Skeen J, Majewski N, Di
Cristofano A, Pandolfi PP, Feliciano CS, Gartel A and Hay N:
Activation of Akt/protein kinase B overcomes a G(2)/m cell cycle
checkpoint induced by DNA damage. Mol Cellular Biol. 22:7831–7841.
2002. View Article : Google Scholar
|