Introduction
Worldwide, hepatocellular carcinoma (HCC) is one of
the most aggressive types of malignancy (1,2). Due to
its poor prognosis, HCC is the third leading cause of
cancer-associated mortality worldwide, a statistic which is even
higher in China (3). There is a
marked geographical variation in the incidence of HCC, and >80%
of cases occur following hepatitis infection. Furthermore, HCC is
typically more severe in China, with a poorer 5-year survival
compared with Western countries (4).
The tumorigenesis of HCC is a multistage process, and it has been
reported that aberrant Toll-like receptor (TLR) signaling may
contribute to the development and progression of HCC. TLRs are
pattern recognition receptors, which have significant functions in
host defense via recognition of pathogen-associated molecular
patterns (5). Studies have suggested
that TLR signaling is involved in the progression of certain
chronic liver diseases. In addition, ongoing clinical trials have
suggested that therapeutic manipulation of TLR pathways may
represent a novel strategy for the reversal of chronic liver
disease (6). Following activation by
their respective ligands, TLRs initiate intracellular
pro-inflammatory/anti-inflammatory signaling cascades through the
recruitment of various adaptor proteins. TLR-associated signaling
pathways are therefore tightly regulated to prevent inappropriate
generation of pro-inflammatory cytokines and interferons, and
thereby prevent aberrant autoimmune and inflammatory processes.
Evidence has indicated that TLR4/5 are involved in
the carcinogenesis of hepatic cancer, indicating that anti-TRLs are
useful and applicable in this disease entity (7,8). In normal
liver tissues, TLRs have crucial roles in the processes of wound
healing and regeneration. In addition, TLRs are involved in the
pathogenesis and progression of various inflammatory liver
diseases, including autoimmune liver disease, non-alcoholic
steatohepatitis, alcoholic liver disease, fibrogenesis and chronic
hepatitis B/C virus (HBV/HCV) infection (9). Further studies have suggested that
TLR-based therapies may represent a promising strategy for the
treatment of viral hepatitis. Since hepatitis is closely associated
with liver cirrhosis and further HCC, and the signaling that was
observed to function in hepatitis also functions in cancer biology
(10,11), it was hypothesized that TLR-based
therapies may be useful therapeutic strategies for the malignancies
associated with hepatic tumors. Our preliminary experiments
revealed that TLR7 is expressed in HCCs, therefore, the effects of
treatment with a TLR7 agonist were investigated in the present
study.
In human tissues, TLR7 is only expressed on human
plasmacytoid dendritic cells, which TLR7 agonist Imiquimod was able
to bind (12). Imiquimod has been
demonstrated to be a strong inducer of interferon-γ and a wide
range of pro-inflammatory mediators (13,14).
Immune system reactions and disorders are regarded as critical
processes in tumor development and initiation; however, the
mechanism underlying the effects of the TLR7 agonist in regulating
cancer initiation and development has remained to be elucidated.
Cancer stem cells (CSCs) were regarded as the roots of tumor
recurrence, and also contribute to chemoradiotherapy resistance
(15,16). TAM tyrosine kinases regulate the
targeted genes involved in the homeostatic regulation of cytokine
receptors and TLR-mediated signal transduction pathways. A loss of
TAM receptors, attributed to exaggerated inflammatory responses by
microglia and characterized by increased mitogen-activated protein
kinase, IκB kinase (IKK) and nuclear factor (NF)-κB activation, as
well as elevated production of pro-inflammatory cytokines, is
detrimental to stem cell proliferation (10). IKK, NF-κB and interleukin-6 (IL6)
signaling have previously been demonstrated to be involved in the
regulation of cancer stem cells; therefore, the present study aimed
to elucidate whether the TLR7 agonist exhibited suppressive effects
on hepatic cancer cells, potentially through IKK and NF-κB
activation.
Materials and methods
Cell culture
The HCCLM3 and MHCC97-H human HCC cell lines were
purchased from the American Type Culture Collection (Manassas, VA,
USA) and cultured in Dulbecco's modified Eagle's medium (DMEM;
Invitrogen, Carlsbad, CA, USA), containing 10% fetal bovine serum
(FBS) and 1% penicillin and streptomycin (Invitrogen). Mammospheres
(CSCs) obtained from the HCCLM3 and MHCC97-H cell lines were
cultured in DMEM/Ham's F-12 medium, supplemented with 10 ng/ml
epidermal growth factor, 10 ng/ml human basic fibroblast growth
factor, 1 µg/ml hydrocortisone, 4 µg/ml insulin and 1% penicillin
and streptomycin (Invitrogen) (17).
All cells were cultured in 5% CO2 at 37°C.
Western blot analysis
Proteins were harvested from the HCC cells after 48
h with radioimmunoprecipitation assay lysis buffer (BioTeke Corp.,
Beijing, China) and 60 µg cellular protein was subjected to 10%
SDS-PAGE separation. Proteins were transferred to polyvinylidene
difluoride microporous membranes (EMD Millipore, Billerica, MA,
USA) and blots were probed with rabbit monoclonal IL6 antibody
(1:2500; ab32530; Abcam, Cambridge, MA, USA), internal control
GAPDH (1:5000; sc-25778), mouse monoclonal immunoglobulin G
(IgG)2a IKK antibody (1:1000; sc-376114) and rabbit
polyclonal IgG NF-κB antibody (1:1000; sc-372) (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). All primary antibodies were
incubated overnight at 4°C.
Cell proliferation assay
Imiquimod
[1-(2-methylprophyl)-1H-imadazo(4,5c)quinoline-4-amine] is a
synthetic low-molecular-weight compound, which is a topical immune
response modifier that upregulates immune responses. Imiquimod
(catalog no. R837; InvivoGen, San Diego, CA, USA) and TLR7 ligand
(cat. no. ODN 20959; Miltenyi Biotec Inc., Auburn, CA, USA) were
used. The cells of each group were treated with 100 nM, 1 µM, 10
µM, 100 µM and 1mM Imiquimod and/or 10 nM, 100 nM, 1 µM and 10 µM
TLR7 ligand for 24, 48 or 72 h prior to MTT detection. The MTT
(Sigma-Aldrich, St Louis, MO, USA) spectrophotometric dye assay was
used to detect cell proliferation ability. HCCLM3 and MHCC97-H
cells were plated in 96-well plates at a density of 2000
cells/well, respectively. At 24, 48 and 72 h post-treatment, cells
were incubated with 10 µl MTT for 4 h. The color was developed by
incubating the cells with 100 µl dimethyl sulfoxide; and the
absorbance was detected at a wavelength of 490 nm using a Bio-Rad
550 Ultramark™ Microplate Reader (Bio-Rad Laboratories, Inc.
Hercules, CA, USA). The data were obtained from three independent
experiments.
Sphere formation assays
To explore the effects of Imiquimod and TRL7 ligand
on the self-renewal of HCC cells, cells of the various groups were
plated in ultra-low attachment (non-adherent condition) dishes
(Corning, Inc., Corning, NY, USA) to test their ability to form
primary mammospheres when incubated at 37°C. On day 7, cell
mammosphere numbers were counted under the low-power field of an
inverted microscope (DMI3000B; Leica Microsystems, Wetzlar,
Germany); briefly, the mammospheres were collected by
centrifugation at 450 × g for 5 min at 4°C, and dissociated
mechanically with a pipette using 0.05% Trypsin-EDTA (Invitrogen).
The obtained spheres were passed through a 40-µm sieve and analyzed
microscopically for single cellularity. The mammosphere forming
efficiency (MFE) was calculated as the percentage ratio between the
obtained spheres and plated cells [mean ± standard deviation (SD)
of three independent experiments] (18). Obtained mammospheres of the various
groups were disaggregated into single cells, and these cells were
re-suspended and plated to test the ability of self-renewal of the
second generation for ≤7 days. The number of secondary mammospheres
was also calculated as the mean ± SD.
Immunofluorescence assay
The HCCLM3 and MHCC97-H cells
(1–3×104/well) were plated in a chamber 20 h prior to
staining, then fixed in 10% formalin (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) for 15 min. The cells were blocked with 5%
goat serum (cat. no. ab7481; Abcam), and incubated with the primary
monoclonal rabbit anti-Snail antibody (1:500; cat. no. 3879; Cell
Signaling Technology, Inc., Danvers, MA, USA) overnight at 4°C. The
membranes were then washed three times with phosphate-buffered
saline (PBS; Calbiotech Inc., Spring Valley, CA, USA) with Tween-20
(Thermo Fisher Scientific, Inc.) (PBST) for 15 min. The membranes
were then incubated with a horseradish peroxidase-conjugated goat
anti-mouse IgG secondary antibody (cat. no. sc-2060; 1:,5000; Santa
Cruz Biotechnology, Inc.) for 1 h at room temperature and washed
with PBST. Next, the membranes were incubated with Alexa
Fluor® 488 secondary antibody (goat IgG; cat. no.
Z25002; 1:1,000; Invitrogen) for 1 h at room temperature. Next, the
membranes were incubated with DAPI (Thermo Fisher Scientific, Inc.)
for 10 min, then washed in PBS. Fluorescence was visualized using a
Leica microscope (DMI3000B; Leica Microsystems).
Flow cytometric analysis
To further confirm the influence of Imiquimod and
the TLR7 ligand on the self-renewal ability of stem cells the
ALDEFLUOR™ fluorescent reagent system (STEMCELL Technologies, Inc.,
Vancouver, Canada) was used to separate aldehyde dehydrogenase
(ALDH)+ cells from the various groups, as previously
described (15). Flow cytometer setup
and data acquisition were followed by ALDH-based Cell Detection kit
analysis (ALDEFLUOR).
Statistical analysis
Data are expressed as the mean ± SD and were
analyzed by Student's t-test and χ2 test, using SPSS for
Windows version 16.0 (SPSS, Inc., Chicago, IL, USA).
Results
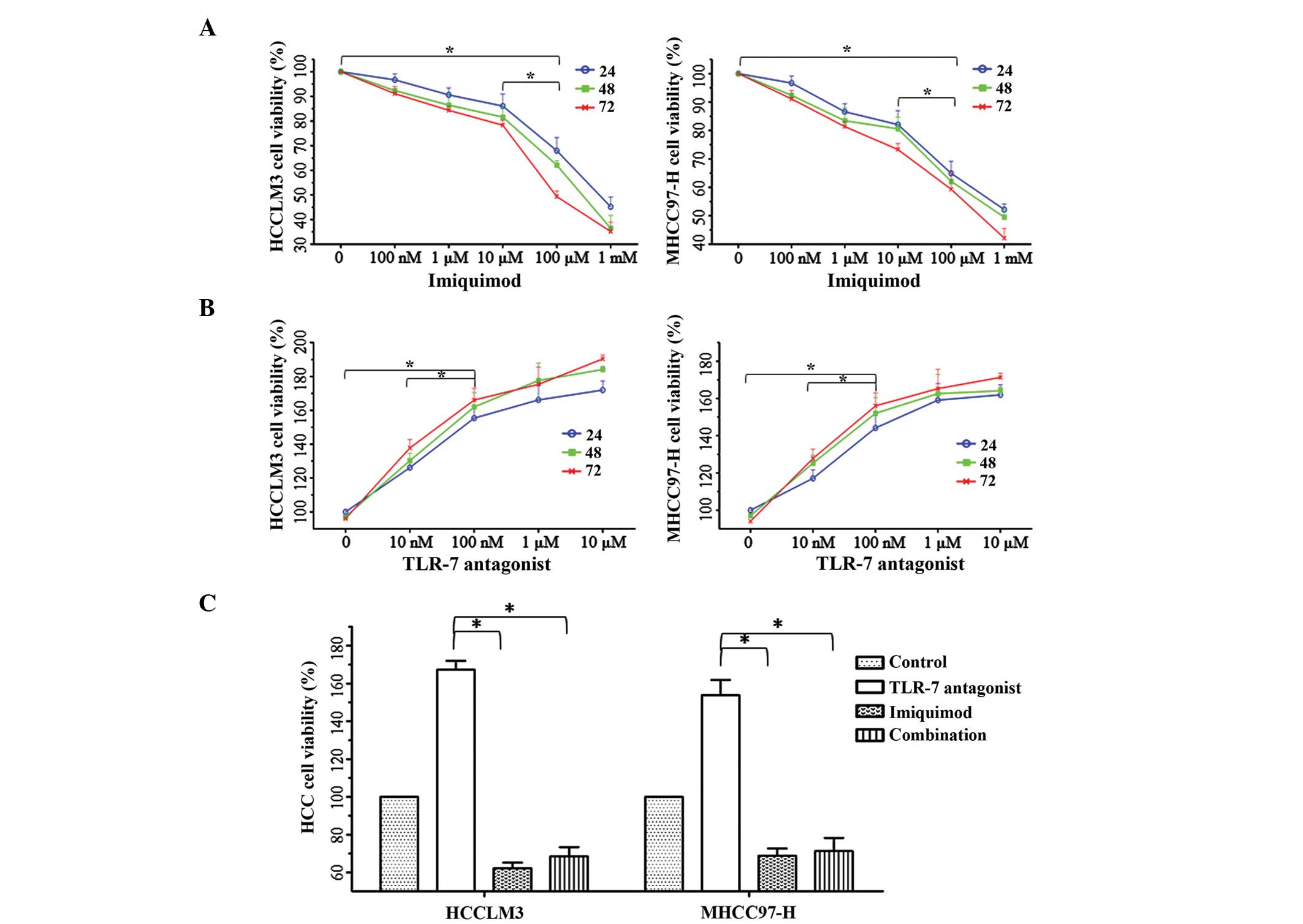
The functions of TLR7 signaling in HCC cell
proliferation. To identify the effects of the TLR7 signaling
pathway on the proliferative potential of HCC cells, the cells were
treated with TLR7 ligand and/or Imiquimod. Cell proliferation was
evaluated using the MTT method at 24, 48 and 72 h, and the
proliferation of cancer cells treated with 100 µM Imiquimod for 48
h was suppressed most effectively, compared with the other groups
(P<0.01; Fig. 1A). Conversely, 100
nM TLR7 ligand significantly induced cell proliferation (Fig. 1B); however, the induction of
proliferation was blocked by the addition of 100 µM Imiquimod,
indicating that 100 µM Imiquimod effectively reversed the oncogenic
function of the TLR7 ligand following 48 h of incubation (Fig. 1C).
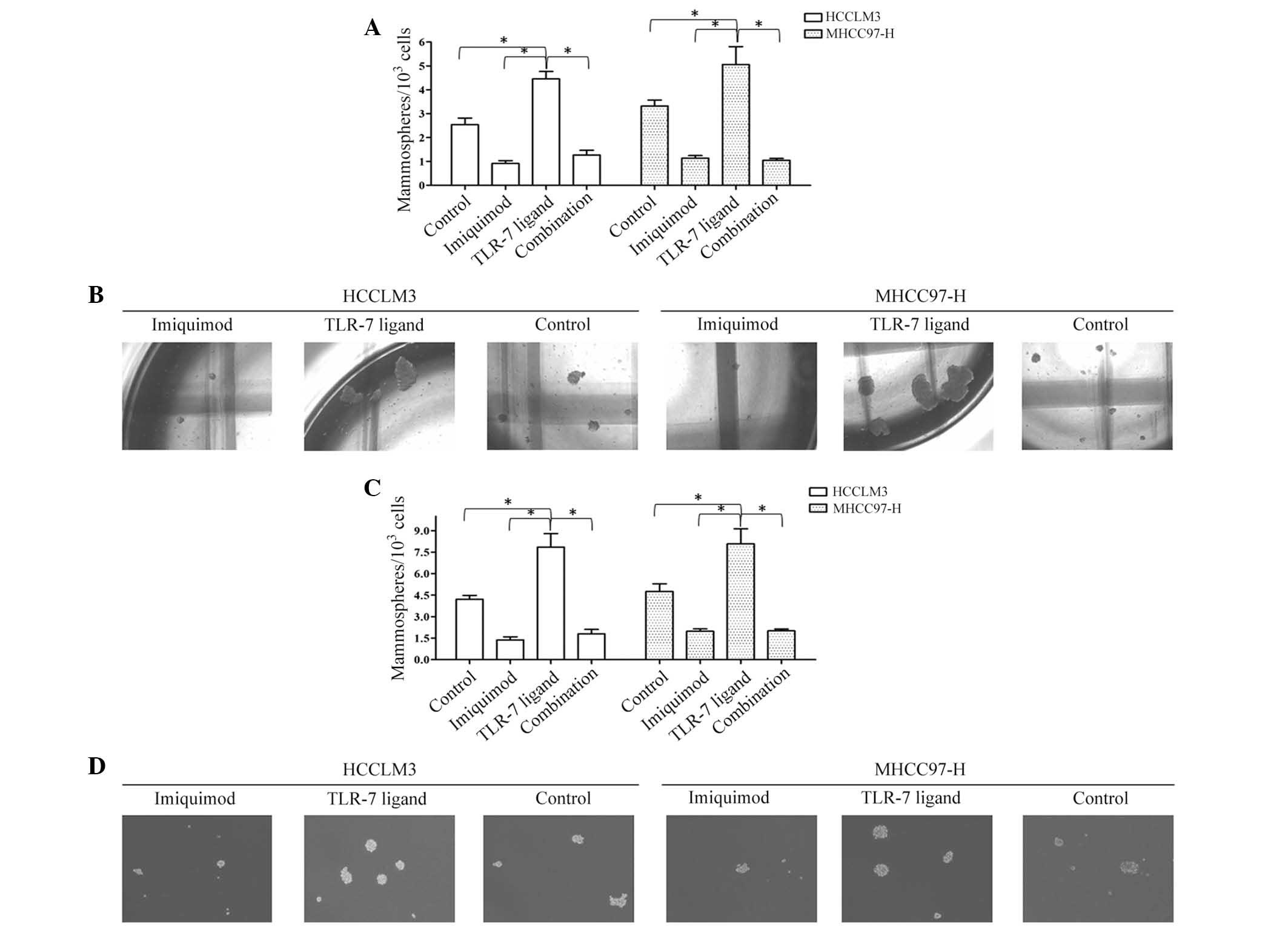
Imiquimod suppresses mammosphere
formation ability of HCC cells
The MFE of Imiquimod-treated HCCLM3 (HCCLM3-I) cells
(0.92±0.11%) was significantly lower than that of the HCCLM3
control group (2.54±0.28%). In addition, the MFE of
Imiqumod-treated MHCC97 (MHCC97-H-I) cells (1.14±0.104%) was
markedly lower than that of the MHCC97-H control group (3.32±0.25%)
(P<0.01; Fig. 2A and B).
Disaggregation of primary mammospheres and secondary plating of
suspended cells resulted in the repeated formation of mammospheres,
which suggested that the mammospheres of each group in the first
round contained cells capable of self-renewal. The MFE of HCCLM3-I
secondary mammospheres (1.37±0.22%) was lower than that of the
HCCLM3 control group (4.21±0.27%); and the MFE of MHCC97-H-I
secondary mammospheres (1.98±0.16%) was lower than that of the
MHCC97-H control group (4.76±0.53%) (P<0.01; Fig. 2C and D). To further confirm the
suppressive effects of Imiquimod on HCC stem cells, the proportion
of ALDH+ cells was detected in the various groups, and
Imiquimod was demonstrated to significantly inhibit CSC numbers
(P<0.01; Fig. 3A).
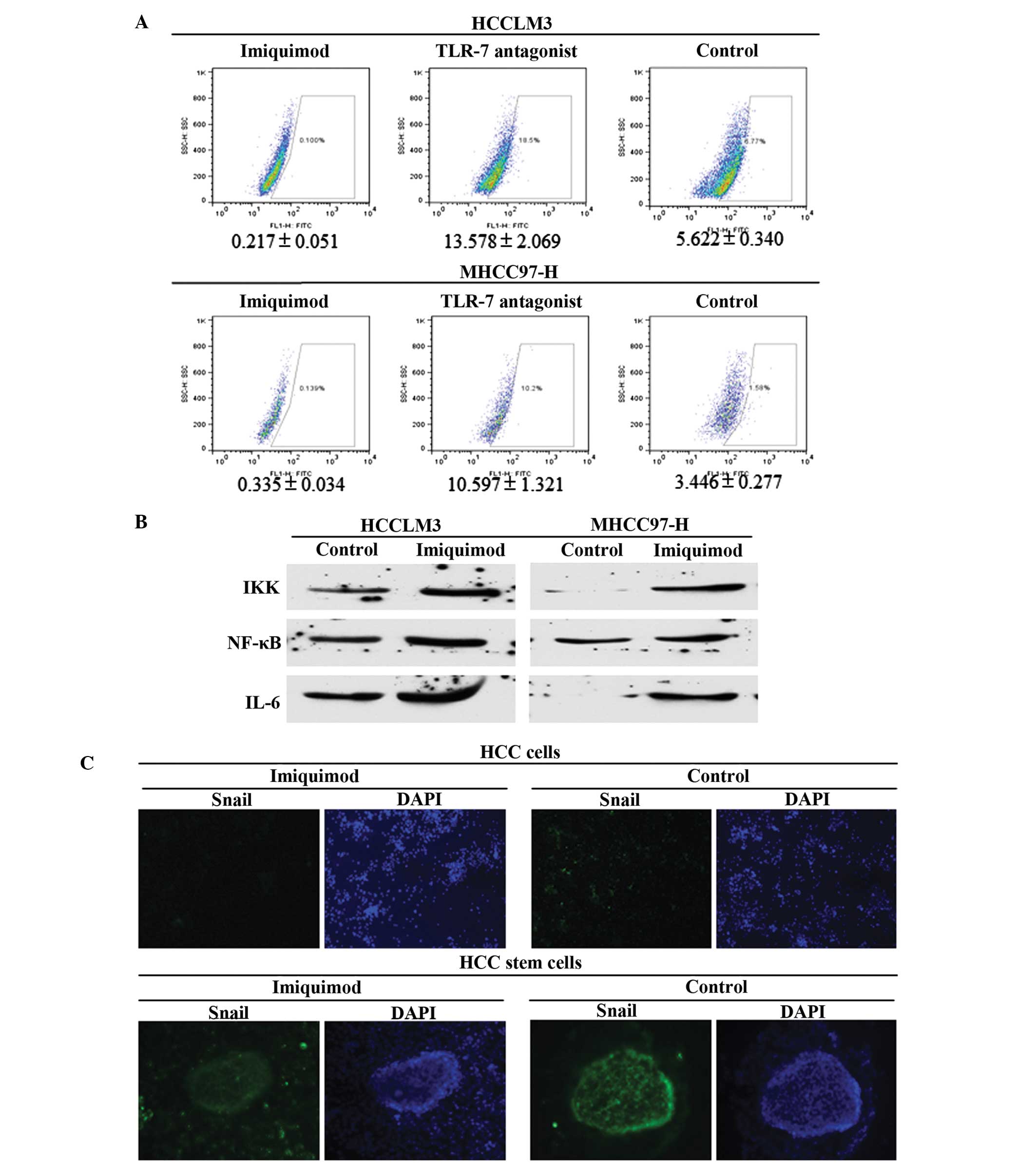 | Figure 3.Suppression and associated mechanisms
of Imiquimod on HCC stem cells. Signaling pathways implicated in
TLR7 agonist induced cell repression were also evaluated. (A)
Imiquimod effectively decreased ALDH+ cells in the two
HCC cell lines, and also decreased TLR7 ligand-induced stem cell
number, P<0.01. (B) Imiquimod treatment decreased IKK, NF-κB and
IL6 expression, which are all downstream genes of TLR7 signaling,
indicating that traditional TLR7-IKK-NF-κB-IL6 signaling likely
functioned in HCC cells. (C) The results of Snail staining in the
various groups of HCC cells and HCC cancer stem cells revealed that
Imiquimod treatment decreased Snail expression. Snail signal was
stronger in mammospheres than that in cancer cells (magnification,
HCC cells, ×20; HCC stem cells, ×40). HCC, hepatocellular
carcinoma; TLR7, Toll-like receptor 7; ALDH, aldehyde
dehydrogenase; IKK, IκB kinase; NF-κB, nuclear factor-κB; IL6,
interleukin-6. |
Imiquimod reverses TLR7 ligand-induced
mammosphere formation ability
The MFE of TLR7 ligand-treated HCCLM3 (HCCLM3-L)
cells (4.46±0.31%) was elevated significantly, compared with that
of the control group (2.54±0.28%) (P<0.01). In addition, the MFE
of TLR7 ligand-treated MHCC97-H (MHCC97-H-L) cells (5.06±0.75%) was
markedly higher than that of the MHCC97-H control group
(3.32±0.25%), P<0.01 (Fig. 2A and
B). The MFE of HCCLM3-L secondary mammospheres was 7.85±0.94%,
compared with 4.21±0.27% of the HCCLM3 control group (P<0.01);
while the MFE of MHCC97-H-L secondary mammospheres was higher
(8.07±1.06%) than that of the MHCC97-H control group (4.76±0.53%)
(P<0.01; Fig. 2C and D).
Furthermore, the TLR7 ligand enhanced the number of
ALDH+ cells of HCC, while Imiquimod abrogated the TLR7
ligand-induced increase in mammosphere numbers (Fig. 2) and inhibited the ALDH+
cells (P<0.01). Combined treatment with Imiquimod and TLR7
ligand therefore neutralized the oncogenic function of the TLR7
ligand in the self-renewal of CSCs (P<0.01; Fig. 2).
Imiquimod inhibits HCC cells and CSCs
via the IKK-NFκB-IL6 signaling pathway
To explore the potential mechanisms by which
Imiquimod suppresses CSC numbers, it was hypothesized that
Imiquimod inhibited malignant cellular behaviors in HCC cells and
HCC stem cells through downregulation of IKK-NFκB-IL6 signaling,
which had previously been demonstrated to function in
TLR7-dominated hepatic diseases (8,9,19). The expression levels of IKK, NFκB and
IL6 were all significantly decreased in the Imiquimod-treated group
(P<0.01; Fig. 3B); and were
identical in the TLR7 ligand and Imiquimod combined group (data not
shown). To further confirm the suppressive effects of Imiquimod on
stem cells, Snail, which is an EMT marker, identifying stem cell
pluripotency, was detected in HCC cells using immunofluorescent
staining. The results indicated that Snail expression was
significantly inhibited in Imiquimod-treated HCC cells (Fig. 3C) and HCC cancer stem cells, compared
with that of the control group (Fig.
3C). The results also indicated that the Snail signal was
markedly stronger in stem cells than that in differentiated cancer
cells.
Discussion
Liver cancer is one of the leading causes of
cancer-associated mortality worldwide, and therefore the
improvement of available therapies is required in order to reduce
this risk. Understanding the mechanisms underlying
hepatocarcinogenesis is vital for the development of more effective
treatments. Chronic liver diseases, including chronic hepatitis and
liver cirrhosis, are predisposing factors for the development of
HCC. TLRs have a crucial role in conferring immunity against
microbial pathogens and evidence has confirmed that TLRs are
implicated in the pathogenesis of chronic liver disease, including
that of HCC. TLR7 was previously confirmed to be upregulated in HCC
tissues (20), and TLR7 stimulation
significantly increased the proliferation of HuH7 cells, while
inhibition of TLR7 using chloroquine resulted in significant
inhibition (21). In vivo
experiments revealed that TLR7 inhibition using chloroquine reduced
tumor growth in xenograft models, and chloroquine also decreased
liver fibrosis and tumor growth in the diethylnitrosamine and
N-nitrosomorpholine model (22).
However, how the TLR7 agonist Imiquimod functions in HCC cells,
particularly in stem cells, has remained elusive. The putative
mechanisms underlying Imiquimod functions require further
investigation, which will aid the development of potential novel
therapeutic strategies using the TLR7 agonists for the prevention
of progression of primary liver cancers in susceptible patients,
together with the results of other groups (23). CSCs were demonstrated to be crucial in
the eradication of the tumor group entirely; therefore, the
suppressive effects of TLR agonists on HCC cells and CSCs will have
significance in the treatment of HCC, in particular, the TLR7
agonist Imiquimod (9,23).
In the present study, the proliferative inhibition
of Imiquimod on HCC cell lines was first evaluated, and inhibitory
effects of Imiquimod were observed in the two HCC cell lines.
Following confirmation of the appropriate and effective
concentration of Imiquimod, it was demonstrated that Imiquimod
treatment attenuated TLR7 ligand-induced cancer cell proliferation.
To explore the functions of Imiquimod on HCC stem cells, the cells
were treated with Imiquimod for 48 h, and the self-renewal ability
was determined using mammosphere formation assay. Imiquimod (100
µl) significantly inhibited the self-renewal of HCC stem cells.
Flow cytometry-based ALDEFLUOR fluorescent sorting, which aims to
separate ALDH+ cells, also demonstrated the influence of
Imiquimod on stem cell numbers. For the first time, to the best of
our knowledge, the present results proved that TLR7 agonist
Imiquimod was able to suppress HCC cell and stem cell proliferation
and self-renewal, an effect which may be mediated by
TLR7-IKK-NFκB-IL6 signaling. The systemic administration of the
selective TLR7 agonist resulted in dose-dependent changes in HCC
cells, indicating the potential for novel therapeutic strategies
for the prevention and progression of primary liver through
targeting CSC groups. Further research is required to analyze the
effects of additional TLR7 agonists in the inhibition of hepatic
carcinoma by suppressing the ratios of CSCs and to elucidate their
associated mechanisms. In addition, further in vivo
experiments are required, to confirm the efficacy and safety of
TLR7 agonist-based anticancer therapies.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thun MJ, DeLancey JO, Center MM, Jemal A
and Ward EM: The global burden of cancer: Priorities for
prevention. Carcinogenesis. 31:100–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Shin H-R, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu HC and Santella R: The role of
aflatoxins in hepatocellular carcinoma. Hepat Mon. 12:e72382012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dennison U, McKernan DP, Scully P, Clarke
G, Cryan J and Dinan T: Menstrual cycle influences toll-like
receptor responses. Neuroimmunomodulation. 19:171–179. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kesar V and Odin JA: Toll-like receptors
and liver disease. Liver Int. 34:184–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mencin A, Kluwe J and Schwabe RF:
Toll-like receptors as targets in chronic liver diseases. Gut.
58:704–720. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mittal D, Saccheri F, Vénéreau E, Pusterla
T, Bianchi ME and Rescigno M: TLR4-mediated skin carcinogenesis is
dependent on immune and radioresistant cells. EMBO J. 29:2242–2252.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dapito DH, Mencin A, Gwak GY, Pradere JP,
Jang MK, Mederacke I, Caviglia JM, Khiabanian H, Adeyemi A,
Bataller R, et al: Promotion of Hepatocellular Carcinoma by the
Intestinal Microbiota and TLR4. Cancer Cell. 21:504–516. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji R, Tian S, Lu HJ and Lu Q, Zheng Y,
Wang X, Ding J, Li Q and Lu Q: TAM receptors affect adult brain
neurogenesis by negative regulation of microglial cell activation.
J Immunol. 191:6165–6177. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Larangé A, Antonios D, Pallardy M and
Kerdine-Römer S: TLR7 and TLR8 agonists trigger different signaling
pathways for human dendritic cell maturation. J Leukoc Biol.
85:673–683. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Y, Abel K, Lantz K, Krieg AM,
McChesney MB and Miller CJ: The Toll-like receptor 7 (TLR7)
agonist, Imiquimod and the TLR9 agonist, CpG ODN, induce antiviral
cytokines and chemokines but do not prevent vaginal transmission of
simian immunodeficiency virus when applied intravaginally to rhesus
macaques. J Virol. 79:14355–14370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oosterhoff D, Heusinkveld M, Lougheed SM,
Kosten I, Lindstedt M, Bruijns SC, van Es T, van Kooyk Y, van der
Burg SH and de Gruijl TD: Intradermal delivery of TLR agonists in a
human explant skin model: Preferential activation of migratory
dendritic cells by polyribosinic-polyribocytidylic acid and
peptidoglycans. J Immunol. 190:3338–3345. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
O'Neill LA, Bryant CE and Doyle SL:
Therapeutic targeting of Toll-like receptors for infectious and
inflammatory diseases and cancer. Pharmacol Rev. 61:177–197. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun X, Jiao X, Pestell TG, Fan C, Qin S,
Mirabelli E, Ren H and Pestell RG: MicroRNAs and cancer stem cells:
The sword and the shield. Oncogene. 33:4967–4977. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kreso A and Dick JE: Evolution of the
cancer stem cell model. Cell Stem Cell. 14:275–291. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun X, Qin S, Fan C, Xu C, Du N and Ren H:
Let-7: A regulator of the ERα signaling pathway in human breast
tumors and breast cancer stem cells. Oncol Rep. 29:2079–2087.
2013.PubMed/NCBI
|
|
18
|
Cicalese A, Bonizzi G, Pasi CE, Faretta M,
Ronzoni S, Giulini B, Brisken C, Minucci S, Di Fiore PP and Pelicci
PG: The tumor suppressor p53 regulates polarity of self-renewing
divisions in mammary stem cells. Cell. 138:1083–1095. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abdel-Raouf TA, Ahmed A, Zaki WK, Abdella
HM and Zid MA: Study of toll-like receptor 7 expression and
interferon α in Egyptian patients with chronic hepatitis C
infection and hepatocellular carcinoma. Egypt J Med Hum Genet.
15:387–392. 2014. View Article : Google Scholar
|
|
20
|
Mohamed FE, Al-Jehani RM, Minogue SS,
Andreola F, Winstanley A, Olde Damink SW, Habtesion A, Malagó M,
Davies N, Luong TV, et al: Effect of toll-like receptor 7 and 9
targeted therapy to prevent the development of hepatocellular
carcinoma. Liver Int. 35:1063–1076. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guiducci C, Tripodo C, Gong M, Sangaletti
S, Colombo MP, Coffman RL and Barrat FJ: Autoimmune skin
inflammation is dependent on plasmacytoid dendritic cell activation
by nucleic acids via TLR7 and TLR9. J Exp Med. 207:2931–2942. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tazzyman S, Barry ST, Ashton S, Wood P,
Blakey D, Lewis CE and Murdoch C: Inhibition of neutrophil
infiltration into A549 lung tumors in vitro and in
vivo using a CXCR2-specific antagonist is associated with
reduced tumor growth. Int J Cancer. 129:847–858. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Adams S, Kozhaya L, Martiniuk F, Meng TC,
Chiriboga L, Liebes L, Hochman T, Shuman N, Axelrod D, Speyer J, et
al: Topical TLR7 agonist imiquimod can induce immune-mediated
rejection of skin metastases in patients with breast cancer. Clin
Cancer Res. 18:6748–6757. 2012. View Article : Google Scholar : PubMed/NCBI
|