Introduction
Advances in contemporary molecular biology have
revealed that a large number of RNAs that have no protein-coding
ability [non-coding (nc)RNAs] exist in various organs and tissues
(1–4).
Many of these ncRNAs are key factors in gene regulation and are
important for normal cellular function as well as disease
pathogenesis (5). Among the different
classes of ncRNAs, long ncRNAs (lncRNAs), which are most commonly
defined as non-protein-coding RNA molecules of >200 nucleotides,
have recently received increasing attention. lncRNAs can be
transcribed from intergenic, intronic and imprinted loci, or from
overlapping or antisense loci adjacent to protein-coding genes.
These diverse transcription patterns of lncRNAs have significant
implications for their various functions, including imprinting, DNA
methylation and X-chromosome dosage compensation, as well as
transcriptional, post-transcriptional and epigenetic regulation
(6–11). The majority of lncRNAs are transcribed
in a developmentally-regulated and cell type-specific manner
(12,13), particularly in the central nervous
system (CNS), where over half of all lncRNAs are expressed
(13–15). Therefore, lncRNAs are now considered
to be highly important for mediating CNS form and function, and
alteration of their expression may cause certain CNS pathologies,
such as Alzheimer's disease, multiple sclerosis, Down's syndrome,
schizophrenia and brain tumors (16).
Gliomas are the most common and aggressive type of
primary adult brain tumor, with an prevalence of 3–6 per 100,000
population in China (17). Their
highly malignant and invasive nature gives rise to a median
survival time of <15 months for patients with glioblastoma (GBM)
undergoing conventional treatment. The pathogenesis of glioma is
complex and involves the aberrant activation of proto-oncogenes and
the inactivation of tumor suppressor genes (18–20).
Growing evidence in the literature has revealed that aberrant
expression of lncRNAs in glioma influence cell proliferation,
apoptosis and invasion through interactions with different
molecules and diverse signaling pathways (21–23),
thereby acting as critical components in the progression of
gliomas. For example, H19, a well-characterized lncRNA in
glioma, drives tumor transformation and contributes to malignant
glioma phenotypes through binding with transcription factor c-Myc
(22,24). CRNDE, the most upregulated
lncRNA in GBM (16), regulates gene
expression through histone methylation/demethylation epigenetic
changes; these histone modifications are induced via interaction
with chromatin modifying complexes CoREST and polycomb repressive
complex 2 (PRC2) (25). In addition,
MEG3, a tumor suppressor lncRNA (23,26),
appears to have an anti-proliferative function in glioma through
the suppression of MDM2 and subsequent activation of the p53
signaling pathway (27).
HOTAIR, which also participates in epigenetic regulation
through PRC2, was demonstrated to be closely associated with tumor
staging, poor prognosis and the molecular subtype of glioma
(17), with knockdown of HOTAIR in
glioma cells exerting a tumor suppressive function accompanied by
significant downregulation of cell-cycle related genes (28,29).
Despite the data obtained from the previous aforementioned studies,
the exact mechanisms by which lncRNAs regulate the development of
gliomas remain largely unclear.
Several studies have suggested that the interaction
between lncRNAs and microRNAs (miRNA/miRs) may have a regulatory
role in cancer (30–33). For example, MEG3 is
methylation-dependent tissue-specific lncRNA that is regulated by
miR-29a and has been reported to contribute to hepatocellular
carcinoma growth (34). Furthermore,
UCA1, an oncogene lncRNA in breast cancer, modulates breast
cancer cell growth and apoptosis by decreasing tumor suppressive
miR-143 (35), and miR-148b-3p, a
member of the miR-148/152 family, was underexpressed (reduced
compared with normal cells) in several cancer cell lines (36–38). In a
study by Zhang et al, miR-148b-3p was found to induce cell
apoptosis by activating caspase-3 and caspase-9, inducing S phase
arrest by regulating cyclin D1 and p21, and inhibiting cell
invasion (39). Although a number of
mRNA targets of miR-148b-3p have been identified, interaction
between miR-148b-3p and an lncRNA has not been reported to date.
Using the online software program starBase v2.0, the present study
identified HOTAIR as a potential lncRNA target of
miR-148b-3p. Subsequently, the current study showed that mutated
HOTAIR promotes the aggressiveness of A172 glioma cells, and
its was determined that miR-148b-3p binds HOTAIR in a
sequence-specific manner. Furthermore, miR-148b-3p reduced
proliferation, cell cycle progression and invasion of A172 cells
through the suppression of HOTAIR expression. Thus, the
current data, at least in part, contributes insight into the
development of glioma.
Materials and methods
Human tissue samples and cell
lines
mRNA and miRNA expression microarray data from 180
samples were downloaded from the Gene Expression Omnibus website
(http://www.ncbi.nlm.nih.gov/geo/;
accession no. GSE4290). The data were compiled from 23 non-tumor,
26 astrocytoma (7 grade II, 19 grade III), 50 oligodendroglioma (38
grade II, 12 grade III) and 81 GBM samples. The tumor sample
expression profile, including HOTAIR expression data, was
also downloaded. HA1800 human astrocytes and A172 glioma cells were
purchased from the Cell Resource Center of Shanghai Institute of
Life Sciences (Shanghai, China). The cell lines were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
calf serum (Gibco; Thermo Fisher Scientific, Inc.), penicillin (100
U/ml) and streptomycin (100 mg/ml) at 37°C in 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells using TRIzol
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's instructions. Briefly, 1 ml TRIzol per
5×105 cells was added to cells, prior to adding 0.2 ml
of chloroform per 1 ml TRIzol. The mixture was mixed vigorously by
hand and allowed to stand for 2–3 min at room temperature. The
mixture was then centrifuged at 10,000 × g for 10 min at 4°C. The
upper clear phase was transferred to a fresh tube and 0.5 ml
isopropanol per 1 ml of the clear phase was added, which was mixed
vigorously by rapid shaking and left to stand for 10 min. The
precipitated RNA was collected by centrifugation at 10,000 × g for
10 min at 4°C and then carefully decanting/pipetting the
supernatant. The RNA precipitate was washed once with 70% ethanol,
dissolved in 25 µl RNase free water, and then stored at −80°C. The
concentration of the Recombinant DNase I RNase-free (Takara
Biotechnology Co., Ltd., Dalian, China) used to treat the RNA
sample was 5 units/µl.
cDNA was synthesized using the HiFi-MMLV cDNA kit
(Beijing ComWin Biotech Co., Ltd., Beijing, China) and qPCR was
conducted using the UltraSYBR Mixture (Beijing ComWin Biotech Co.,
Ltd.). Briefly, 5 µg purified RNA sample was mixed with Primer Mix,
dNTP Mix, DTT, RT-buffer, HiFi-MMLV and RNase-free water using a
pulled pipette (total volume, 20 µl). All qRT-PCR reactions were
run in a StepOnePlus™ Real-Time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The mixture was then incubated at
42°C for 30–50 min and then 85°C for 5 min. The RT products were
quickly centrifuged and stored at −20°C. No cDNA was used as a
negative control. To amplify hsa-miR-148b-3p cDNA, specific RT
primers were used based on its sequence, and the U6 RT primer for
was the same as the U6 reverse PCR primer. The hsa-miR-148b-3p RT
primer was
5′-GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCACCAGAGCCAACACAAAG-3′. PCR
primers were as follows: Forward, 5′-GGCACCACACCTTCTACAAT-3′ and
reverse, 5′-GTGGTGGTGAAGCTGTAGCC-3′ for the β-actin gene;
forward, 5′-CAGTGGGGAACTCTGACTCG-3′ and reverse,
5′-GTGCCTGGTGCTCTCTTACC-3′ for the HOTAIR gene; forward,
5′-CGGTCAGTGCATCACAGAA-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′ for
hsa-miR-148b-3p; and forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′ for U6. All primers were synthesized by
GenScript (Nanjing, China). The relative fold change in mRNA
expression level was calculated using the 2−ΔΔCq method
(40).
Cell transfection
siRNA HOTAIR (siHOTAIR), siRNA
negative control (siNC), miR-148b-3p mimic and miR-148b-3p
inhibitor were synthesized by Biomics Biotechnologies Co., Ltd.
(Nantong, China). The sequences were as follows: Sense,
5′-CCACAUGAACGCCCAGAGAUU-3′ and antisense,
5′-AAUCUCUGGGCGUUCAUGUGG-3′ for si-HOTAIR (41); and sense, 5′-UUCUCCGAACGUGUCACGUTT-3′
and antisense, 5′-ACGUGACACGUUCGGAGAATT-3′ for si-NC (22). Cells were grown tuntil they reached
105 in number in 6-well plates prior to transfection.
One day before transfection, the control group cells were plated at
the same density as the si-HOTAIR and si-NC groups. The
cells were transfected using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
instructions.
Cell viability assay
The effect of HOTAIR downregulation on the
viability of A172 cells was assessed using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. Briefly, cells were trypsinized and seeded at a density of
1×104 cells/well in 96-well plates immediately after
siRNA transfection. Then, 10 µl MTT solution (5 mg/ml) was added
and the plates were incubated for an additional 4 h at 37°C.
Following removal of the medium, formazan crystals were dissolved
in 150 µl dimethyl sulfoxide. The absorbance of the MTT formazan
was measured at 550 nm using a SpectraMax M3 microplate reader
(Molecular Devices, LLC, Sunnyvale, CA, USA). Experiments were
repeated three times using 6 wells for each treatment to ensure the
reproducibility of results.
Flow cytometry analysis (FCM)
Cells were separately harvested 0, 24, 48 and 72 h
after siRNA transfection, fixed in 70% ethanol, and stained with
propidium iodide (Nanjing KeyGen Biotech Co. Ltd., Nanjing, China)
containing RNase A (1 mg/ml; Takara Biotechnology Co., Ltd.) for 30
min at 37°C. Subsequently, 500 µl of cells was filtered through
200-µm mesh sieves, and the cell cycle profiles were assayed using
a Guava easyCyte 8 Flow Cytometer (EMD Millipore, Billerica, MA,
USA).
Matrigel invasion assay
Transwell inserts (diameter, 6.5 mm) with a pore
size of 8 µM (Corning Incorporated, Corning, NY, USA) were coated
with Matrigel (100 µg/well; BD Biosciences, San Jose, CA, USA) and
placed into the wells of 24-well culture plates. Following
transfection with siRNA for 48 h, 1×104 cells were
transferred into the top of the invasion chambers in serum-free
DMEM, and DMEM containing 20% fetal calf serum was added to the
lower chambers. After 24 h of incubation at 37°C, non-invasive
cells were removed with a cotton swab, and the invading cells were
fixed with 4% paraformaldehyde for 15 min, stained with Giemsa for
20 min at room temperature and observed under an ECLIPSE Ti S
microscope (Nikon, Tokyo, Japan). Experiments were independently
repeated three times.
Plasmid construction
To verify whether miR-148b-3p regulates
HOTAIR as a direct target, a predicted binding site (12 bp)
for miR-148b-3p was identified using the online software program
starBase v2.0 (42). The human
HOTAIR fragment containing the putative binding sites for
miR-148b-3p was synthesized, annealed and inserted into the
NotI and XbaI restriction sites of the pmirGLO
luciferase reporter vector (Promega, Madison, WI, USA), downstream
of the luciferase gene, to generate the recombinant vectors
pmirGLO-wild-type (WT) and pmirGLO-mutant (MUT). For the
pmirGLO-MUT construct, 12 mismatches were introduced into the
HOTAIR sequence, producing a change of
GATGCATTTTCTGTGCACTGGtoGATGCACCTCCCACATGTCAG; therefore, a human
HOTAIR fragment containing mutated binding sites was
synthesized. The constructs were validated by Sanger sequencing by
Sangon Biotech Co., Ltd. (Shanghai, China).
Luciferase reporter assay
For the luciferase reporter assay, A172 cells were
co-transfected with miRNA (miR-148b-3p mimics or miR-148-3p mimic
negative control; Biomics Biotechnologies Co., Ltd., Nantong,
China) and reporter vectors (pmirGLO-WT or pmirGLO-MUT) using
Lipofectamine 2000. Luciferase activity was assayed 48 h after
transfection using a Dual-Luciferase Reporter Assay system
(Beyotime Institute of Biotechnology, Haimen, China). The values
were normalized to those obtained for miRNA negative control
transfection. All transfection experiments were performed in
triplicate.
Statistical analysis
All statistical analyses were performed using SPSS
software version 18.0 (SPSS, Inc., Chicago, IL, USA). Data are
expressed as means ± standard deviation of experiments performed in
triplicate. HOTAIR expression in normal tissues and tumor
tissues was compared using analysis of variance (ANOVA) followed by
Student-Newman-Keuls analysis. GraphPad Prism version 5.0 (GraphPad
Software, La Jolla, CA, USA) was used for the graphing. Statistical
analysis on other experiments was performed using one-way ANOVA and
the Student-Newman-Keuls test for multiple comparisons, and
unpaired Student's t-tests for comparisons between groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
HOTAIR expression in glioma tissues
and cells
First, the expression pattern of HOTAIR was
analyzed through whole genome gene expression profiling of 157
glioma and 23 normal tissue samples from NCBI GEO data (accession
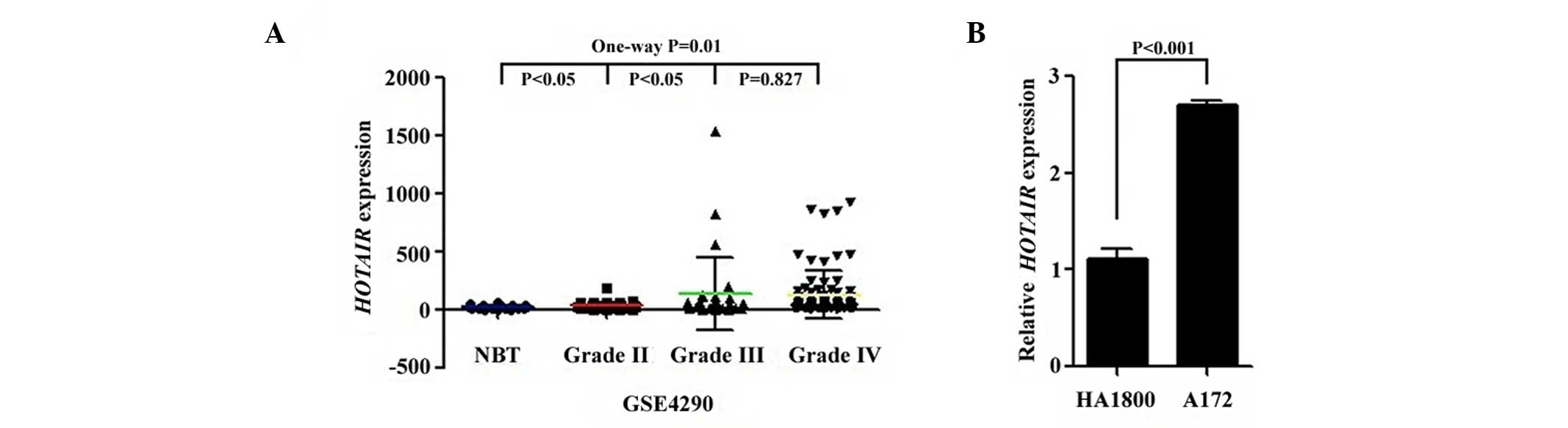
no. GSE4290). As shown in Fig. 1A,
HOTAIR expression was significantly higher in carcinoma
tissues compared with in normal tissues (P=0.010), and grade IV and
III tumor tissues both demonstrated a significant increase in
HOTAIR transcription levels compared with grade II tumor
tissues (P=0.047). However, no significant difference in
HOTAIR expression was observed between grade IV and III
samples. Next, HOTAIR expression was detected in HA1800
human astrocytes and A172 glioma cells by RT-qPCR. The
HOTAIR mRNA level was significantly increased in A172 cells
compared with HA1800 cells (P<0.001; Fig. 1B). These findings indicate that
HOTAIR may have an important role in glioma progression.
Silencing HOTAIR inhibits the
malignant properties of A172 cells
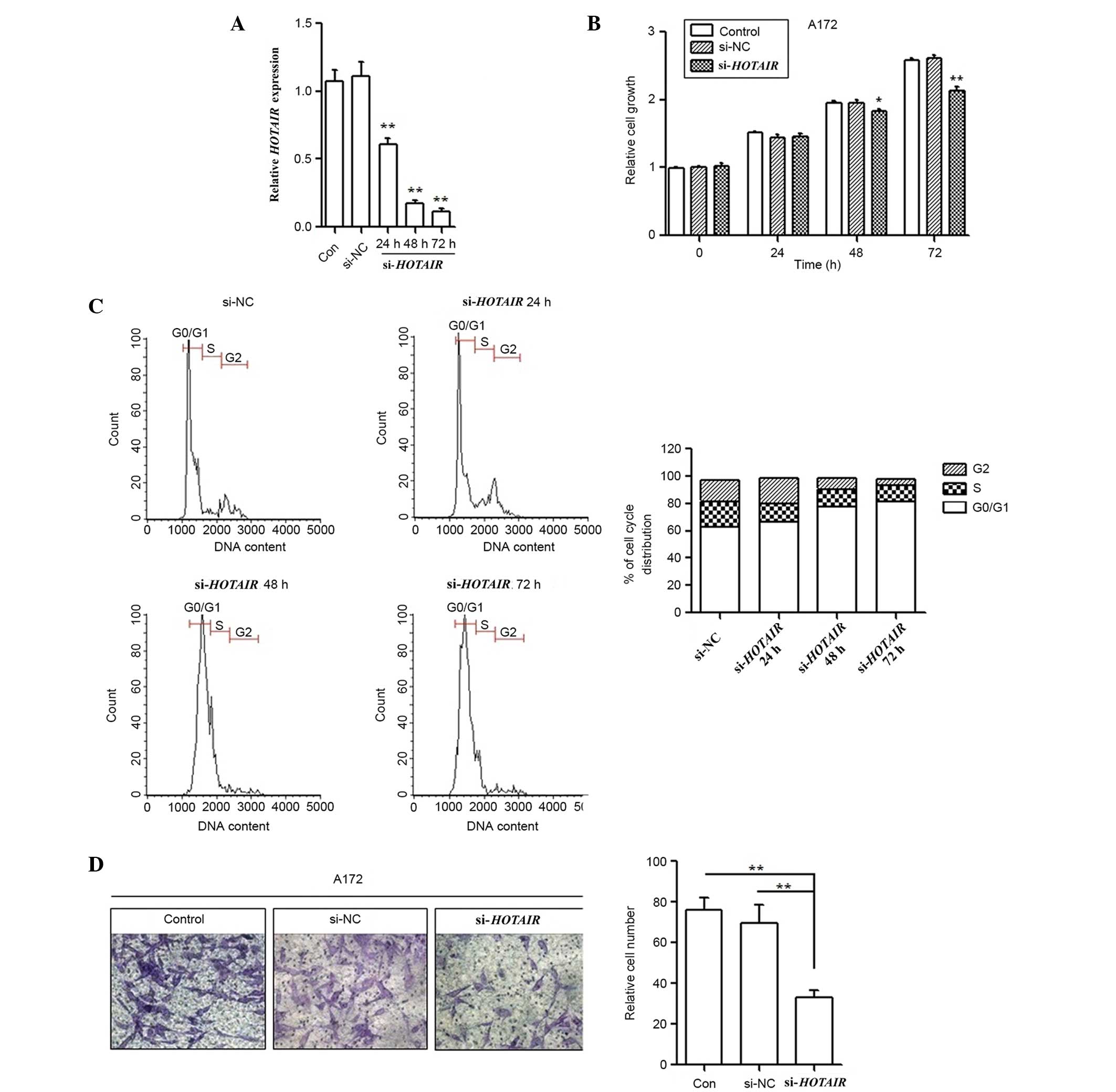
To further investigate the oncogenic role of
HOTAIR in glioma pathogenesis, A172 cell lines were
transfected with si-HOTAIR. Successful transfection was
confirmed by RT-qPCR (Fig. 2A). Cells
were separately collected at 0, 24, 48 and 72 h post-transfection,
and cell growth ability was determined using the MTT assay.
Silencing of HOTAIR expression significantly reduced growth
rates compared with the si-NC and untransfected cell groups at 48
(P=0.012) and 72 h (P<0.001) post-transfection (Fig. 2B). Additionally, FCM was performed
following transfection to assess cell cycle distribution. The
results showed that the cell population in the G1 phase was
increased but the S phase population was decreased after
HOTAIR gene silencing compared with the results observed for
the si-NC cells (Fig. 2C), further
indicating that knockdown of HOTAIR expression may suppress
cancer cell proliferation. In addition, the effect of HOTAIR
knockdown on A172 cell invasion was investigated using a Matrigel
invasion assay. The in vitro Matrigel invasion assay
revealed that the invasiveness of A172 cells transfected with
si-HOTAIR was significantly suppressed compared with the
control and si-NC cells (P=0.007; Fig.
2D).
Reciprocal negative regulation of
miR-148b-3p and HOTAIR
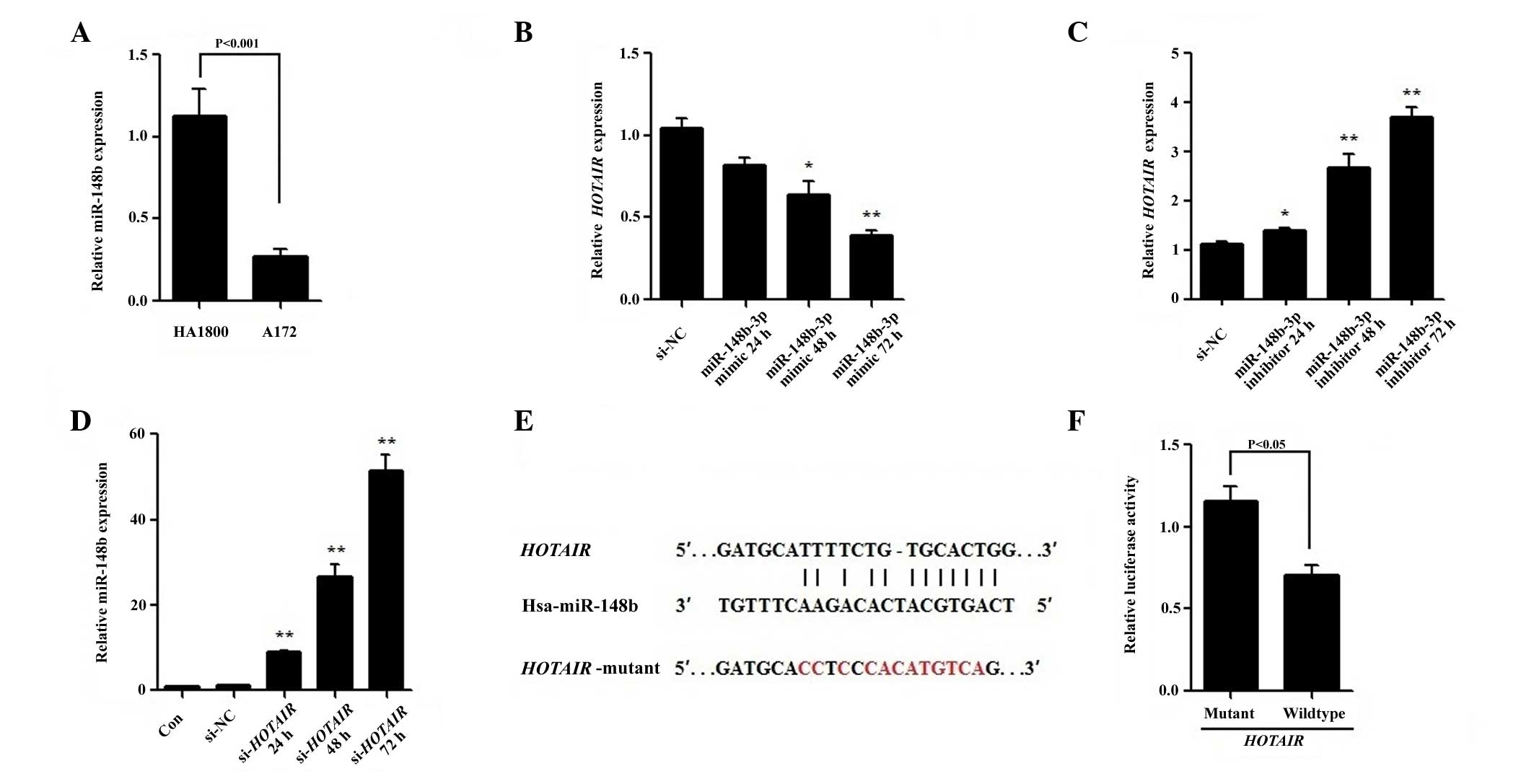
RT-qPCR revealed that the expression level of
miR-148b-3p was significantly lower in A172 cells compared with
HA1800 cells (P<0.001; Fig. 3A).
To determine whether miR-148b-3p is able to suppress HOTAIR
expression, miR-148b-3p mimic or inhibitor were transfected into
A172 cells. As shown in Fig. 3B, the
miR-148b-3p mimic reduced HOTAIR expression in a
dose-dependent manner, decreasing it by ~62% 72 h after
transfection (P<0.001). By contrast, the miR-148b-3p inhibitor
increased the level of HOTAIR in a dose-dependent manner
(Fig. 3C). Furthermore,
si-HOTAIR transfection significantly increased the
expression of miR-148b-3p in A172 cells at all time points
(P<0.001; Fig. 3D), indicating
that there is a strong inverse correlation between HOTAIR
and miR-148b-3p expression levels.
To verify whether miR-148b-3p regulates
HOTAIR as a direct target, a predicted binding site for
miR-148b-3p was identified using the online software program
starBase v2.0 (42) (Fig. 3E) an wild-type or mutant miR-148b-3p
target binding sequences were cloned into the pmirGLO luciferase
vector. Following co-transfection with the miR-148b-3p mimic in
A172 cells, a Dual-Luciferase Assay was performed determine the
luciferase activity. The data shows that cells co-transfected with
the constructs containing the pmirGLO-WT and miR-148b-3p mimic had
significantly lower luciferase activity compared with that of those
transfected with pmirGLO-MUT and miR-148b-3p mimic (P=0.007;
Fig. 3F). All the results indicate
that miR-148b-3p suppresses HOTAIR by binding to
HOTAIR in a sequence-specific manner.
miR-148b-3p inhibits HOTAIR
function
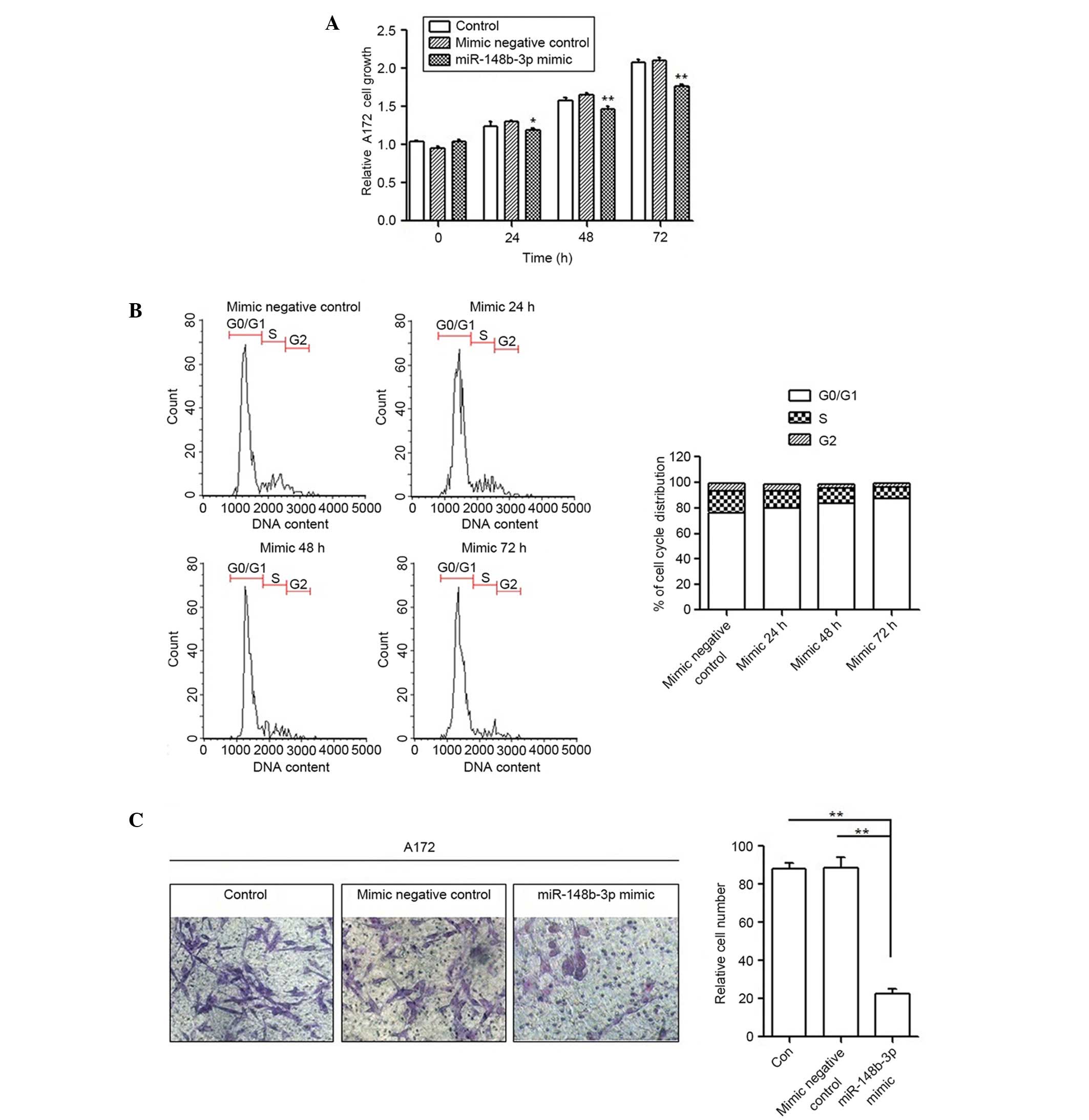
To detect whether the aggressiveness of A172 cells
could be restored by miR-148b-3p, an MTT assay, FCM and a Transwell
invasion assay were performed following transfection with the
miR-148b-3p mimic. Transient transfection with the miR-148b-3p
mimic decreased the proliferation of A172 cells by ~15% at 72 h
compared with the mimic negative control group (P=0.002; Fig. 4A). The miR-148b-3p mimic also
increased the cell population in the G1 phase and decreased the
S-phase population (Fig. 4B),
demonstrating that miR-148b-3p blocks the cell cycle of A172 cells.
Furthermore, the Transwell invasion assay results clearly revealed
that the miR-148b-3p mimic decreased cell invasion to ~70% compared
with the untreated and miR-148b-3p mimic negative controls
(P=0.002; Fig. 4C).
Discussion
Years of research have revealed the essential role
of non-coding lncRNAs and miRNAs in gene regulation, and their
involvement in diverse biological processes, such as development
and disease. Recently, rapid advances in bioinformatic analysis
have expanded understanding of the transcriptome to a genome-wide
level, and the interaction between lncRNA and miRNA has been shown
to provide an additional element of control in gene regulation
(43–45). In support an interaction between miRNA
and lncRNA, the present study demonstrated the reciprocal
repression of miRNA-148b and HOTAIR. The addition of a
miR-148b-3p mimic decreased the expression levels of HOTAIR,
while miR-148b-3p inhibitor increased the levels of HOTAIR.
In addition, knockdown of HOTAIR induced the upregulation of
miR-148b-3p.
HOTAIR is a 2,158-bp lncRNA that is located
within the HOXC cluster of chromosome 12, and is flanked by
HOXC11 and HOXC12. HOTAIR activity is
partially regulated by its interaction with PRC2, which is
comprised of EZH2 (a histone H3K27 methylase), SUZ12
and EED. Since Gupta et al (41) first observed that loss of
HOTAIR can inhibit the invasiveness of cancer cells,
increasing evidence for a significant role of HOTAIR in
carcinogenesis has been documented for several types of cancer,
including glioma. The current study confirmed that HOTAIR is
highly expressed in glioma tissues and the A172 glioma cell line.
Furthermore, knockdown of HOTAIR decreased growth rates,
blocked the cell cycle and suppressed the invasion of glioma cells,
as well as increasing the expression of miR-148b-3p.
Previous studies have shown that miR-148b-3p is
dysregulated in numerous types of cancer (46–48). For
example, in colorectal cancer, it is downregulated and acts as a
tumor suppressor by targeting the CCKBR gene (38). miR-148b-3p is also downregulated in
pancreatic cancer and suppresses cell growth by targeting the
AMPKα1 gene (49). However, it
is upregulated in ovarian cancer and may be involved in the early
stages of ovarian carcinogenesis (50). These results reflect the controversial
roles of miR-148b-3p in different types of cancer. To the best of
our knowledge, the present study is the first to demonstrate that
miR-148b-3p is downregulated in glioma cells. Additionally, an
association between miR-148b-3p and lncRNA expression in cancer has
not been reported until now. In the current study, luciferase
assays indicated that miR-148b-3p reduces HOTAIR expression
through the putative miR-148b-3p binding site in HOTAIR. To
investigate whether miR-148b-3p has a metastasis-suppressing
function in glioma cells, a miR-148b-3p mimic was transfected into
A172 cells, resulting in reduced expression levels of
HOTAIR. The data also showed that miR-148b-3p inhibits cell
proliferation, blocks the cell cycle and reduces cell invasion.
In conclusion, the results of the current study
indicate that miR-148b-3p has a tumor-suppressive role by
downregulating HOTAIR in glioma, providing evidence for the
importance of the interaction between lncRNA and miRNA in
gliomagenesis. These results will aid in providing novel
considerations for the molecule-targeted therapies of glioblastoma.
Additional investigations are underway to further investigate other
molecules involved in the HOTAIR-miR-148b-3p
interaction.
Acknowledgements
The current study was supported in part by research
grants from the Natural Science Foundation of Zhejiang Province,
Youth Fund Project (grant no. LQ12C07001), the Research Fund for
the Doctoral Program of Higher Education of China (grant no.
20133322120002), Zhejiang Medical and Health Science and Technology
Project (grant no. 2014KYA150), the Science and Technology Creative
Activity Plan for University Students in Zhejiang Province (grant
no. 2015R410050) and the National Training Programs of Innovation
and Entrepreneurship for Undergraduates (grant no.
201410344004).
Glossary
Abbreviations
Abbreviations:
|
CNS
|
central nervous system
|
|
DMEM
|
Dulbecco's modified Eagle medium
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
References
|
1
|
Mattick JS: Challenging the dogma: The
hidden layer of non-protein-coding RNAs in complex organisms.
BioEssays. 25:930–939. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mattick JS: RNA regulation: A new
genetics? Nat Rev Genet. 5:316–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Szymanski M, Barciszewska MZ, Erdmann VA
and Barciszewski J: A new frontier for molecular medicine:
Noncoding RNAs. Biochim Biophys Acta. 1756:65–75. 2005.PubMed/NCBI
|
|
4
|
Prasanth KV and Spector DL: Eukaryotic
regulatory RNAs: An answer to the ‘genome complexity’ conundrum.
Genes Dev. 21:11–42. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amaral PP, Dinger ME, Mercer TR and
Mattick JS: The eukaryotic genome as an RNA machine. Science.
319:1787–1789. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Geisler S and Coller J: RNA in unexpected
places: Long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wilusz JE, Sunwoo H and Spector DL: Long
noncoding RNAs: Functional surprises from the RNA world. Genes Dev.
23:1494–1504. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang XQ and Leung GK: Long non-coding
RNAs in glioma: Functional roles and clinical perspectives.
Neurochem Int. 77:78–85. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qureshi IA, Mattick JS and Mehler MF: Long
non-coding RNAs in nervous system function and disease. Brain Res.
1338:20–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mercer TR, Dinger ME, Sunkin SM, Mehler MF
and Mattick JS: Specific expression of long noncoding RNAs in the
mouse brain. Proc Natl Acad Sci USA. 105:716–721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ravasi T, Suzuki H, Pang KC, Katayama S,
Furuno M, Okunishi R, Fukuda S, Ru K, Frith MC, Gongora MM, et al:
Experimental validation of the regulated expression of large
numbers of non-coding RNAs from the mouse genome. Genome Res.
16:11–19. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Carninci P, Kasukawa T, Katayama S, Gough
J, Frith MC, Maeda N, Oyama R, Ravasi T, Lenhard B, Wells C, et al:
RIKEN Genome Exploration Research Group and Genome Science Group
(Genome Network Project Core Group): The transcriptional landscape
of the mammalian genome. Science. 309:1559–1563. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kiang KM, Zhang XQ and Leung GK: Long
Non-Coding RNAs: The Key Players in Glioma Pathogenesis. Cancers
(Basel). 7:1406–1424. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hatanpaa KJ, Burma S, Zhao D and Habib AA:
Epidermal growth factor receptor in glioma: Signal transduction,
neuropathology, imaging, and radioresistance. Neoplasia.
12:675–684. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan H, Parsons DW, Jin G, McLendon R,
Rasheed BA, Yuan W, Kos I, Batinic-Haberle I, Jones S, Riggins GJ,
et al: IDH1 and IDH2 mutations in gliomas. New Eng N Engl J Med.
360:765–773. 2009. View Article : Google Scholar
|
|
20
|
Endersby R and Baker SJ: PTEN signaling in
brain: Neuropathology and tumorigenesis. Oncogene. 27:5416–5430.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gordon FE, Nutt CL, Cheunsuchon P,
Nakayama Y, Provencher KA, Rice KA, Zhou Y, Zhang X and Klibanski
A: Increased expression of angiogenic genes in the brains of mouse
meg3-null embryos. Endocrinology. 151:2443–2452. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi Y, Wang Y, Luan W, Wang P, Tao T,
Zhang J, Qian J, Liu N and You Y: Long non-coding RNA H19 promotes
glioma cell invasion by deriving miR-675. PLoS One. 9:e862952014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang P, Ren Z and Sun P: Overexpression of
the long non-coding RNA MEG3 impairs in vitro glioma cell
proliferation. J Cell Biochem. 113:1868–1874. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Barsyte-Lovejoy D, Lau SK, Boutros PC,
Khosravi F, Jurisica I, Andrulis IL, Tsao MS and Penn LZ: The c-Myc
oncogene directly induces the H19 noncoding RNA by allele-specific
binding to potentiate tumorigenesis. Cancer Res. 66:5330–5337.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ellis BC, Molloy PL and Graham LD: CRNDE:
A long non-coding RNA involved in CanceR, Neurobiology, and
DEvelopment. Front Genet. 3:2702012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Sun S, Pu JK, Tsang AC O, Lee D,
Man VO, Lui WM, Wong ST and Leung GK: Long non-coding RNA
expression profiles predict clinical phenotypes in glioma.
Neurobiol Dis. 48:1–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou Y, Zhong Y, Wang Y, Zhang X, Batista
DL, Gejman R, Ansell PJ, Zhao J, Weng C and Klibanski A: Activation
of p53 by MEG3 non-coding RNA. J Biol Chem. 282:24731–24742. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang K, Sun X, Zhou X, Han L, Chen L, Shi
Z, Zhang A, Ye M, Wang Q, Liu C, et al: Long non-coding RNA HOTAIR
promotes glioblastoma cell cycle progression in an EZH2 dependent
manner. Oncotarget. 6:537–546. 2015.PubMed/NCBI
|
|
29
|
Ma MZ, Li CX, Zhang Y, Weng MZ, Zhang MD,
Qin YY, Gong W and Quan ZW: Long non-coding RNA HOTAIR, a c-Myc
activated driver of malignancy, negatively regulates miRNA-130a in
gallbladder cancer. Mol Cancer. 13:1562014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Q, Huang J, Zhou N, Zhang Z, Zhang A,
Lu Z, Wu F and Mo YY: LncRNAloc285194 is a p53-regulated tumor
suppressor. Nuclei Acids Res. 41:4976–4987. 2013. View Article : Google Scholar
|
|
31
|
Zhang Z, Zhu Z, Watabe K, Zhang X, Bai C,
Xu M, Wu F and Mo YY: Negative regulation of lncRNA GAS5 by miR-21.
Cell Death Differ. 20:1558–1568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jalali S, Bhartiya D, Lalwani MK,
Sivasubbu S and Scaria V: Systematic transcriptome wide analysis of
lncRNA-miRNA interactions. PLoS One. 8:e538232013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Juan L, Wang G, Radovich M, Schneider BP,
Clare SE, Wang Y and Liu Y: Potential roles of microRNAs in
regulating long intergenic noncodingRNAs. BMC Med Genomics. 6(Suppl
1): S72013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Braconi C, Kogure T, Valeri N, Huang N,
Nuovo G, Costinean S, Negrini M, Miotto E, Croce CM and Patel T:
microRNA-29 can regulate expression of the long non-coding RNA gene
MEG3 in hepatocellular cancer. Oncogene. 30:4750–4756. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tuo YL, Li XM and Luo J: Long noncoding
RNA UCA1 modulates breast cancer cell growth and apoptosis through
decreasing tumor suppressive miR-143. Eur Rev Med Pharmacol Sci.
19:3403–3411. 2015.PubMed/NCBI
|
|
36
|
Song YX, Yue ZY, Wang ZN, Xu YY, Luo Y, Xu
HM, Zhang X, Jiang L, Xing CZ and Zhang Y: MicroRNA-148b is
frequently down-regulated in gastric cancer and acts as a tumor
suppressor by inhibiting cell proliferation. Mol Cancer. 10:12011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao Y, Jia HL, Zhou HJ, Dong QZ, Fu LY,
Yan ZW, Sun J, Ren N, Ye QH and Qin LX: Identification of
metastasis-related microRNAs of hepatocellular carcinoma in
hepatocellular carcinoma cell lines by quantitative real time PCR.
Zhonghua Gan Zang Bing Za Zhi. 17:526–530. 2009.(In Chinese).
PubMed/NCBI
|
|
38
|
Song Y, Xu Y, Wang Z, Chen Y, Yue Z, Gao
P, Xing C and Xu H: MicroRNA-148b suppresses cell growth by
targeting cholecystokinin-2 receptor in colorectal cancer. Int J
Cancer. 131:1042–1051. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang JG, Shi Y, Hong DF, Song M, Huang D,
Wang CY and Zhao G: MiR-148b suppresses cell proliferation and
invasion in hepatocellular carcinoma by targeting WNT1/β-catenin
pathway. Sci Rep. 5:80872015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNAceRNA, miRNA-ncRNA and protein-RNA
interaction networks from largescaleCLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Guo L, Zhao Y, Yang S, Zhang H and Chen F:
An integrated analysis of miRNA, lncRNA, and mRNA expression
profiles. Biomed Res Int. 2014:3456052014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu Q, Guo L, Jiang F, Li L, Li Z and Chen
F: Analysis of the miRNA-mRNA-lncRNA networks in ER+ and ER- breast
cancer cell lines. J Cell Mol Med. 19:2874–2887. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yoon JH, Abdelmohsen K and Gorospe M:
Functional interactions among microRNAs and long noncoding RNAs.
Semin Cell Dev Biol. 34:9–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou Z, Su Y and Fa X: Restoration of BRG1
inhibits proliferation and metastasis of lung cancer by regulating
tumor suppressor miR-148b. Onco Targets Ther. 8:3603–3612.
2015.PubMed/NCBI
|
|
47
|
Ghasemkhani N, Shadvar S, Masoudi Y,
Talaei AJ, Yahaghi E, Goudarzi PK and Shakiba E: Down-regulated
microRNA 148b expression as predictive biomarker and its prognostic
significance associated with clinicopathological features in
non-small-cell lung cancer patients. Diagn Pathol. 10:1642015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cimino D, De Pitta C, Bear F, Casara S,
Zampini M, Romualdi C, Damascus C, Pinatel E, Ponzone R, Brisken C,
et al: miR-148b is a major coordinator in a relapse-associated miR
signature in breast tumors. FEBS J. 278:199–200. 2011.
|
|
49
|
Zhao G, Zhang JG, Liu Y, Qin Q, Wang B,
Tian K, Liu L, Li X, Niu Y, Deng SC, et al: miR-148b functions as a
tumor suppressor in pancreatic cancer by targeting AMPKα1. Mol
Cancer Ther. 12:83–93. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chang H, Zhou X, Wang ZN, Song YX, Zhao F,
Gao P, Chiang Y and Xu HM: Increased expression of miR-148b in
ovarian carcinoma and its clinical significance. Mol Med Rep.
5:1277–1280. 2012.PubMed/NCBI
|