Introduction
The main therapeutic achievements in treating cancer
have been historically obtained by limiting cancer cell
proliferation or division using non-specific, thereafter, targeted
therapies against these cells. More recently, an increasing
interest has emerged on the role of the tumor microenvironment in
the development of cancer lesions (1). Within this microenvironment, immune
cells which can mediate the destruction of cancer cells and the
immune system as a whole offer a number of novel potential targets
for additional therapies (2), a
proportion of which have provided practical applications for
treating patients. The first efficient treatments using
immunotherapies targeting PD-1/PD-L1 were beneficial for patients
with melanoma (3) and such treatments
were recently approved for treating lung cancer as well (4). Blood vessels, which are part of the
tumor microenvironment and already the targets of anti-angiogenic
drugs in clinical practice for treating colon or lung cancers, also
serve a crucial role as a natural physical barrier which regulates
the immune response. Indeed, endothelial cells which line the inner
side of blood vessels, regulate the extravasation of circulating
immune cells into tissues. Under non-activated, normal conditions,
endothelial cells form a non-adhesive surface which does not allow
for strong interactions with circulating immune cells (5). Upon stimulation by pro-inflammatory
cytokines, endothelial cells become activated and express high
levels of cell surface leukocyte adhesion molecules such as E- and
P-selectin, intercellular adhesion molecule-1 (ICAM-1) and vascular
cell adhesion molecule-1 (VCAM-1). These molecules participate in
the capture of circulating immune cells through the rolling,
arrest, firm adhesion, and extravasation of leukocytes from the
circulation toward the tissues (5).
This process is particularly important in the context of cancer
where activation of endothelial cells directly controls the
penetration of cytotoxic immune cells in the tumor foci (6,7). The gene
coding for EGF-like domain-containing protein 7 (egfl7) is mainly
expressed in blood vessel endothelial cells (8–10) and its
expression is deregulated in human cancer. High expression levels
of egfl7 were correlated with a more advanced stage of human colon
cancer (11), with poor survival in
human hepatocarcinoma (12), and with
a higher tumor grade in human gliomas (13). Egfl7 expression was also associated
with poor overall survival in human pancreatic cancer (14). Furthermore, a previous study
demonstrated in breast cancer tissue samples that a high egfl7
protein score in cancer cells corresponded with low VCAM-1, ICAM-1,
and interferon-γ in the tumors (15).
Using experimental approaches, it has been demonstrated that, when
expressed by tumor cells, egfl7 promotes tumor escape from immunity
by downregulating the activation of tumor blood vessels (15). Egfl7 expressed constitutively by blood
vessel endothelial cells continuously inhibits the expression of
leukocyte-adhesion cell surface receptors and thus limits
endothelium activation. In the context of cancer, egfl7 expressed
by tumor blood vessels similarly prevents the cytotoxic immune
cells from entering the tumor mass, thus protecting tumor from
immune destruction (15).
To the best of our knowledge, no previous study has
addressed the role and expression of egfl7 in blood vessels of the
tumor microenvironment. In the present study, a large cohort of
human breast cancer lesions were included and the expression levels
of egfl7 and that of ICAM-1 as a marker of endothelium activation,
were evaluated in peritumoral vessels of human breast cancer
samples.
Materials and methods
Human samples
The patient population is described in details in
the main section. Human tissue samples were processed and stored at
the pathology laboratory of the Centre Oscar Lambret cancer clinic
(Lille, France). Following surgery, tissues were fixed in 4%
paraformaldehyde and embedded in paraffin. For each tumor, analyses
were performed on sections from the paraffin block which had been
used for estrogen receptor, progesterone and HER2 expression
analyses and for diagnosis, considered by the pathologists to be
the most representative of the lesion.
Antibodies
For staining human tissues, a primary monoclonal
rabbit anti-human ICAM-1 antibody (Abcam, Cambridge, UK; ab53013),
a monoclonal mouse anti-human CD31 (DAKO #M823, clone JC/70A), a
monoclonal mouse anti-CD34 (DAKO #M7165, clone QBend-10), and a
goat biotinylated anti-rabbit IgG (BA-1000, Vector laboratories)
were used. For staining mouse tissues, an anti-mouse ICAM-1
antibody (Abcam ab25375), an anti-rat-Alexa 488 antibody (Life
Technologies A21208), and a biotinylated rat anti-mouse-CD31
(Pharmingen 553371) were used. Antibodies were reconstituted and
stored according to the manufacturers' recommendations.
Immunostaining of human tumor
samples
Four successive paraffin sections (4-µm) were
prepared from each sample paraffin block and laid on superfrost +
slides, dried overnight at 56°C, and rehydrated before staining.
For ICAM-1 staining, slides were placed in methanol, 0.3%
H2O2 for 20 min at room temperature, rinsed
in TRIS-HCl 10 mM, pH 7.5, NaCl 0.15 M (TRIS buffer saline, TBS)
for 5 min, incubated in Antigen retrieval, solution citrate (Vector
Laboratories, Peterborough, UK) for 20 min at 90°C, then for 20 min
at room temperature, and rinsed twice in TBS. Slides were incubated
in TRIS-HCl 0.1 M, pH 7.5, NaCl 0.15 M, 20% goat serum, 0.5%
blocking buffer from TSA Biosystem (Perkin Elmer, Beaconsfield, UK)
for 2 h and incubated in rabbit anti-human ICAM-1 (1:200) overnight
at 4°C in a humidified atmosphere. Slides were then washed in TBS
for 5 min twice and incubated in goat biotinylated anti-rabbit IgG
(1:250) for 45 min. Slides were then washed three times in TBS for
5 min, incubated in Streptavidin-A-HRP (TSA Biosystem kit, 1:100)
in blocking buffer for 30 min, washed 3 times in TRIS-HCl 0.1 M, pH
7.5, NaCl 0.15 M, Tween-20 0.25% (TNT) for 5 min, then incubated in
biotinylated tyramine (1:50) in amplification buffer (TSA
Biosystem, Perkin Elmer), washed 3 times in TNT for 5 min,
incubated in streptavidin-A-HRP (TSA Biosystem kit, Perkin Elmer,
1:100) in blocking buffer for 30 min, washed twice in TNT for 5 min
and the immune complexes revealed using DAB kit (Vector
Laboratories) under microscope observation. Staining was stopped
with tap water and slides counterstained with hemalun, dehydrated
and mounted.
CD31 and CD34 immunostainings were performed using a
BenchMark ULTRA automat and dedicated reagents (Ventana Medical
Systems, Roche Diagnostics, Basel, Switzerland). For CD31 staining,
slides were deparaffinized, incubated at 95°C for 36 min in cell
conditioning buffer 1, washed in Reaction Buffer. Endogenous
peroxydases were inactivated using UV INHIBITOR for 4 min and
slides further washed in Reaction Buffer. Slides were then
incubated in anti-CD31 (Dako, Agilent Technologies, Inc., Santa
Clara, CA, USA, #M0823, 1:20) for 28 min, rinsed in Reaction Buffer
and incubated in HRP UNIV MULT for 8 min, washed in Reaction
Buffer, developed using UV DAB and counterstained in Hematoxylin
for 4 min, then in Bluing Reagent, washed in Reaction Buffer before
dehydration and mounted. For CD34 staining, slides were processed
similarly except that the cell conditioning buffer 1 treatment was
performed for 8 min and the slides were incubated with mouse
anti-CD34 (DAKO, clone QBend-10, 1:50) for 20 min.
Stained slides were analyzed by two independent
oncologists, including a breast cancer pathologist, using an
Axioplan 2 microscope (Zeiss) and compared to the corresponding
hematoxylin/phloxin safran-stained slides used for identification
of the tumor sub-regions for diagnosis. Image acquisition was done
using the ZEN2012 blue edition software (Carl Zeiss AG, Oberkochen,
Germany).
In situ hybridization
Paraffin sections (4-µm) were laid on superfrost +
slides and dried overnight at 56°C. Digoxigenin-labelled sens and
anti-sens egfl7 probes were prepared from a pCMV-sport 6 plasmid
containing the full length human egfl7 cDNA using the Riboprobe
Gemini system II kit (Promega Corporation, Inc., Madison, WI, USA).
Slides were processed using a Ventana Discovery automat as
described previously (8).
Cell transfection, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Primary human umbilical vein endothelial cells
(Lonza Biologics PLC, Slough, UK) were cultured in EGM-2 medium and
passaged according to the manufacturers' recommendations. Cells
(4×104/well) between passage 2 and 5 were plated in 0.33
cm2 wells and transfected with 50 nM non-targeting small
interfering RNA (siRNA, siCtrl, D-001810-01, Dharmacon, Inc.,
Lafayette, CO, USA), or with siRNA targeting egfl7 [siEgfl7,
J-015668-10, Dharmacon, Inc., (15,16)] or
ICAM-1 (siICAM-1, L-003502-00 Dharmacon, Inc.) using Lipofectamine
RNAiMax (Life Technologies, ThermoFisher Scientific, Inc., Waltham,
MA, USA). Three days later, the cells were lysed and total RNA
isolated using the Nucleospin RNA kit and reagents (Macherey-Nagel,
Düren, Germany). RNA were quantified using a Nanodrop (ThermoFisher
Scientific, Inc.) and RT was performed using the High Capacity
Reverse Transcription kit (Life Technologies, ThermoFisher
Scientific, Inc.). qPCR was performed in duplex reactions, mixing
TaqMan FAM-labelled probes for human egfl7 (Hs00211952_m1) or for
ICAM-1 (Hs00164932.m1) with the normalizing β2-microglobulin (B2M)
VIC-labelled probe in the same tube and processed in a StepOne
machine. Data were expressed as 2−ΔΔCq where ΔCq=Cq of
gene-Cq of B2M, and ΔΔCq=ΔCq sample-ΔCq control.
Ethics
Sample storage, handling and analysis were performed
according to the European regulations and the Helsinki Declaration.
Patient consent and legal authorizations were obtained for all the
analyses performed and for the processing of patients personal
data. The protocol was approved by the ‘Comité de Protection des
Personnes Nord-Ouest IV’ on January 12, 2010.
Statistical analysis
All statistical analysis was performed using STATA
11.2 statistical software (StataCorp, College Station, TX, USA).
Analyses used Chi2 and Fisher's exact tests for categorical data
and Kruskall-Wallis test for continuous variables and Cuzick's test
for trend across ordered groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Patient population
Patients included in the present study underwent
surgery for breast cancer at the Oscar Lambret Center cancer clinic
between January 1st, 2005 and July 31st, 2005, for
histologically-proven invasive ductal carcinoma (IDC), invasive
lobular carcinoma (ILC), or ductal carcinoma in situ (DCIS).
Patients were at least 18 years-old, had not been previously
treated with chemotherapy, and had not received neo-adjuvant
chemotherapy. Patients whose tumors exhibited a histology-proven
lobular carcinoma in situ were excluded from the study. The
cohort therefore included: 30 DCIS and 174 invasive carcinomas,
among which 131 IDC (64.2%, Table I),
27 ILC (13.2%), and 16 others types (tubular carcinoma, apocrine,
or neuroendocrine carcinoma, 7.8%). The population was composed of
39 stage 1 (22.4%), 98 stage 2 (56.3%), and 37 stage 3 carcinomas
(21.2%). Among invasive cancers, 20 (11.5%) exhibited HER2 gene
amplification as defined by a 3+ score using immunohistochemistry
or >=2+ score when using chromogenic in situ
hybridization. Hormonal receptors were positive as defined by
estrogen receptor >=10% and/or progesterone receptor >=10% in
85.8% of cases. A total of 13 triple negative breast cancer (7.5%),
as defined by estrogen and progesterone receptor equal to zero
associated with Her2 negative expression, were identified within
this cohort.
 | Table I.Patient population. |
Table I.
Patient population.
| Tumors | N | % |
|---|
| Histological
type | 204 | 100 |
|
IDC | 131 | 64.2 |
|
ILC | 27 | 13.2 |
|
DCIS | 30 | 14.7 |
|
Other | 16 | 7.8 |
| SBR grade | 174 | 100 |
| 1 | 39 | 22.4 |
| 2 | 98 | 56.3 |
| 3 | 37 | 21.3 |
| TNM
classification | 166 | 100 |
| T | 13 | 7.5 |
|
T0-T1a | 42 | 24.1 |
|
T1b | 65 | 37.4 |
|
T1c | 43 | 24.7 |
| T2 | 3 | 1.7 |
| T3 |
|
|
| TNM
classification | 204 | 100 |
| N | 112 | 54.9 |
| NO | 59 | 28.9 |
| N1 | 4 | 1.9 |
| N2 | 8 | 3.9 |
| N3 | 21 | 10.3 |
| Nx |
|
|
| Hormonal
receptor | 204 | 100 |
|
HR+ | 175 | 85.8 |
|
HR− | 26 | 12.7 |
| UK | 3 | 1.5 |
| HER2 status | 174 | 100 |
|
Negative | 151 | 86.8 |
|
Positive | 20 | 11.5 |
|
Unknown | 3 | 1.7 |
| Triple
negative | 13/174 | 7.5 |
Parallel tissue sections were prepared for the
positive identification of blood vessels, for the quantitation of
activated vessels among the identified blood vessels, and for the
quantitation of egfl7 expression levels in the same tumor areas.
The analysis was concentrated on peritumoral vessels and the tumor
sample block which had been used for the initial diagnosis of the
patient's lesion was selected, thus being the most representative
and the closest tissue to the tumor.
Identification of blood vessels
In order to quantify the activated blood vessels in
peritumoral areas, the initial step was to establish a staining
procedure for positively identifying blood vessels in the human
tissue sections. For this purpose, immunostaining using anti-CD34
and anti-CD31 antibodies were compared, as both are widely used
markers of endothelial cells (17).
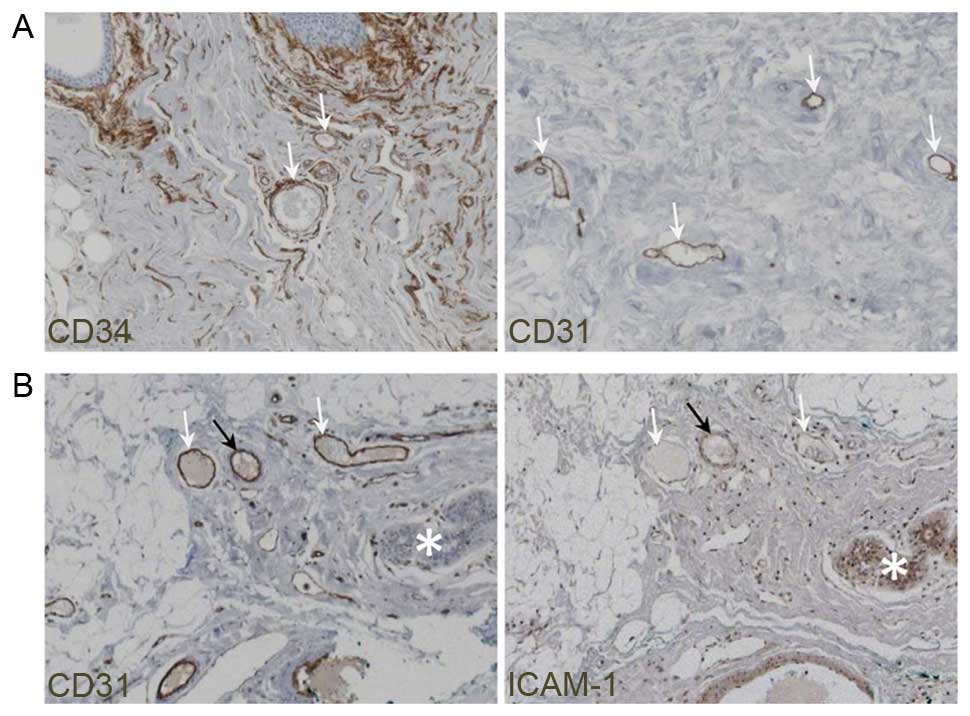
Staining human breast cancer samples with either antibody clearly
revealed the blood vessels (Fig. 1A)
but staining for CD31 provided a more specific signal and gave rise
to a much lower background than staining for CD34, which also
stained non-vascular structures. Therefore all breast cancer tissue
sections were subsequently identified with CD31 staining and blood
vessels were positively identified as CD31+
structures.
Identification of activated blood
vessels in peritumoral areas
Activated blood vessel endothelial cells in breast
cancer samples were identified following ICAM-1 immunostaining.
ICAM-1 staining was membranous in endothelial cells. Its expression
in vascular structures was confirmed by checking the CD31 staining
in parallel sections of the same area (Fig. 1B). In case of an absence of
ICAM-1+ vessels and in order to dismiss any technical
artifact on a specific sample, ICAM-1+ staining was
verified on lobular islets or galactophoric channels of the same
sample, as these structures also stain for this marker. The
percentage of ICAM-1+ activated vessels compared with
ICAM-1− non-activated vessels, was evaluated and scored.
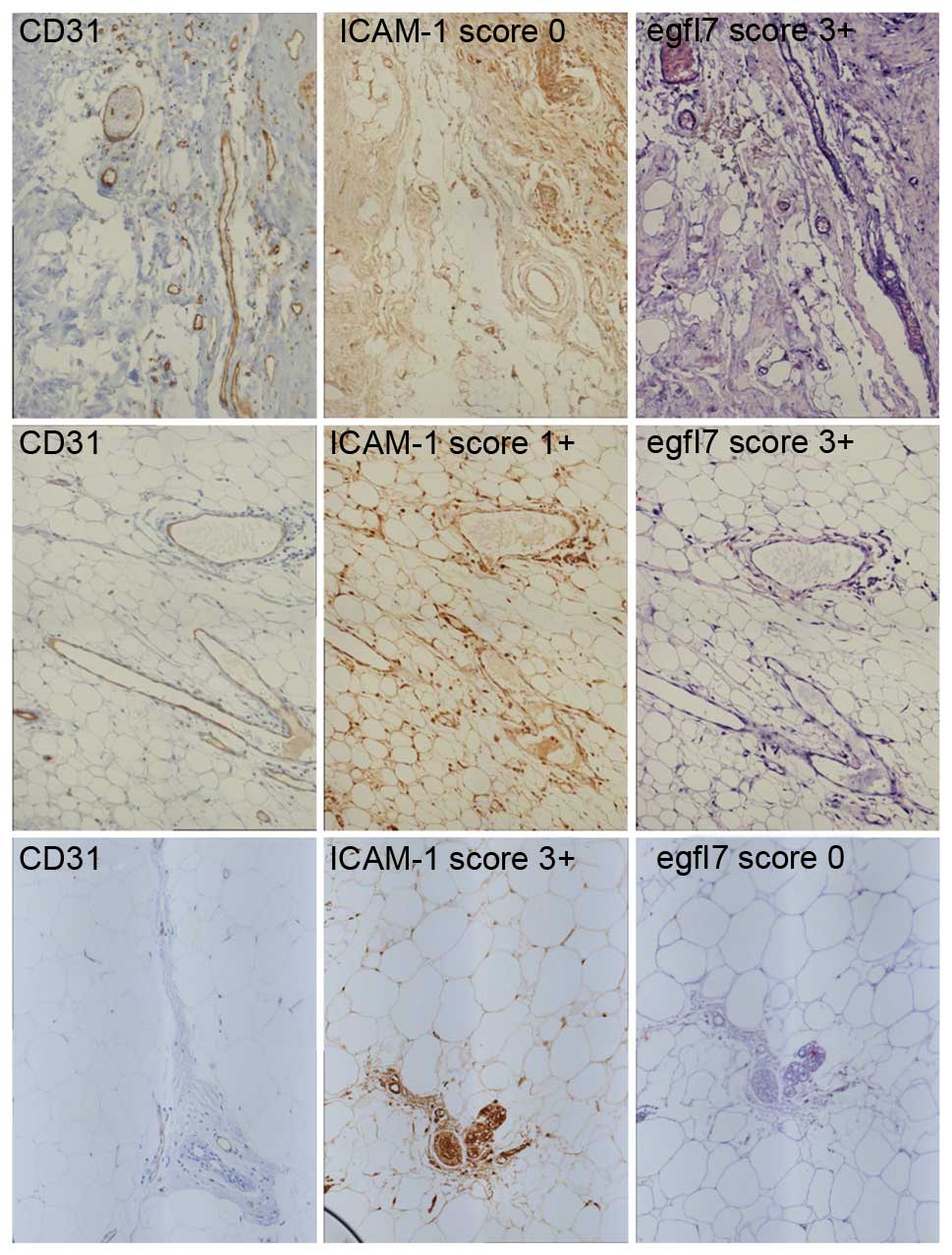
Score 0 was defined as no ICAM-1+ vessels, scores 1+,
2+, and 3+ were defined by 1–29%, 30–60% and >60% of
ICAM-1+ vessels, respectively. Activation of blood
vessels was heterogeneous within any given tumor, as vessels
displayed different states of activation in the same sample
depending on their location and on their size (Figs. 1B and 2). Among the 175 cases analyzed for ICAM-1,
68 lesions (38.9%) were scored 0 for ICAM-1, 62 (35.4%) were scored
1+, 28 (16.0%) were scored 2+ and 17 (9.7%) were scored 3+. A
significant correlation was not identified between ICAM-1
expression in peritumoral blood vessels and clinical data such as
cancer type, grade, metastases, hormone receptors, and HER2 status,
or with the triple-negative type, using the Chi2 test if
theoretical size ≥5 and Fisher exact test otherwise.
Blood vessels are less activated in
areas where egfl7 is highly expressed
Regarding egfl7 transcript detection using in
situ hybridization, positive cases were defined based on the
nuclear staining of endothelial cells and scoring was made using
the same criteria as for ICAM-1 staining. Among the 148 cases
analyzed for egfl7 expression, 63 (42.6%) were scored 0, 30 (20.3%)
were scored 1+, 27 (18.2%) were scored 2+ and 28 (18.9%) were
scored 3+. A significant correlation was not identified between
egfl7 expression in peritumoral area and clinical data including
cancer type, grade, metastases, hormone receptors, nor HER2 status
using the Chi2 test if theoretical size ≥5 and Fisher exact test
otherwise.
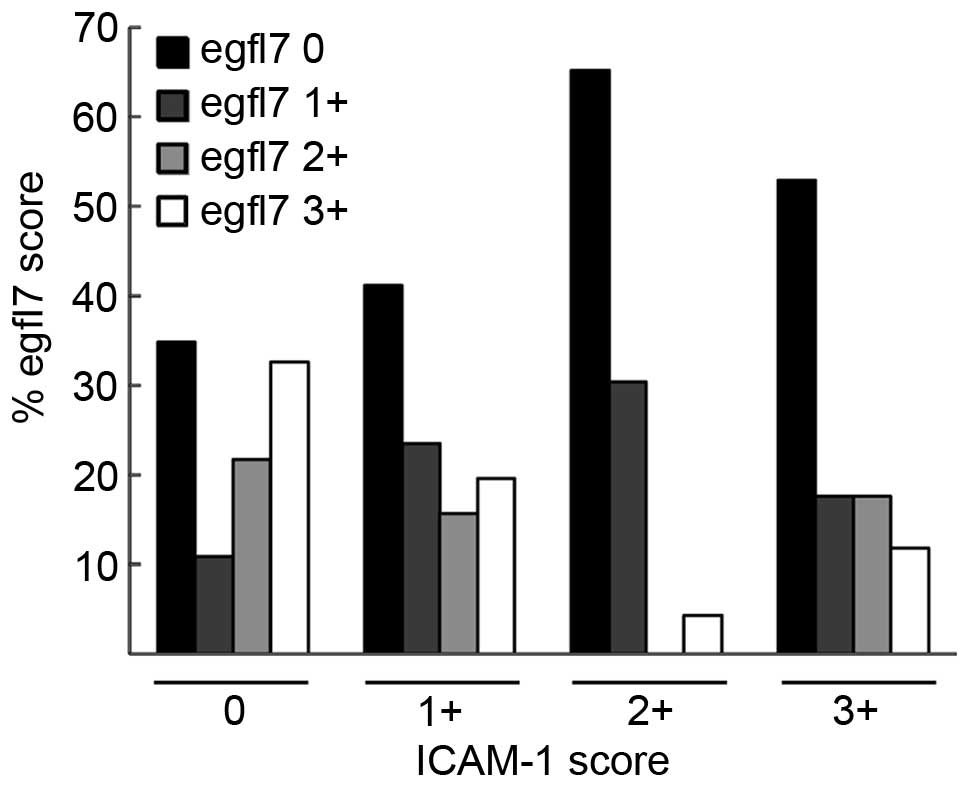
However, a strong and significant correlation was
apparent between the ICAM-1 scores and the egfl7 scores in the
analyzed lesions (P=0.015, Fisher exact test). Cuzick's tests for
trend demonstrated that when the ICAM-1 score increased, the egfl7
score decreased (Fig. 3, P=0.004,
Table II), and vice-versa (P=0.001,
Table II). These observations
directly implied that, regardless of the type and grade of breast
cancer lesions, the expression levels of ICAM-1 and those of egfl7
were somehow linked.
| Table II.Correlations between ICAM-1 scores
and egfl7 scores. |
Table II.
Correlations between ICAM-1 scores
and egfl7 scores.
|
| ICAM-1 score |
|
|
|
|---|
|
|
|
|
|
|
|---|
|
| 0 (0%) | 1+ (0–29%) | 2+ (30–60%) | 3+ (>60%) | Total |
|
|---|
|
|
|
|
|
|
|
|
|---|
|
| n | % | n | % | n | % | n | % | n | % | P |
|---|
| egfl7 score |
|
|
|
|
|
|
|
|
|
| Fisher exact |
| 0 (0%) | 16 | 26.2 | 21 | 34.4 | 15 | 24.6 | 9 | 14.8 | 61 | 100.0 | P=0.015 |
| 1+ (0–29%) | 5 | 18.5 | 12 | 44.4 | 7 | 25.9 | 3 | 11.1 | 27 | 100.0 |
|
| 2+ (30–60%) | 10 | 47.6 | 8 | 38.1 | 0 |
0.0 | 3 | 14.3 | 21 | 100.0 | Test for trend |
| 3+ (>60%) | 15 | 53.6 | 10 | 35.7 | 1 |
3.6 | 2 |
7.1 | 28 | 100.0 |
P=0.004a |
| Total | 46 | 33.6 | 51 | 37.2 | 23 | 16.8 | 17 | 12.4 | 137 | 100.0 |
P=0.001b |
Egfl7 regulates ICAM-1 expression but
ICAM-1 does not affect egfl7 expression in endothelial cells
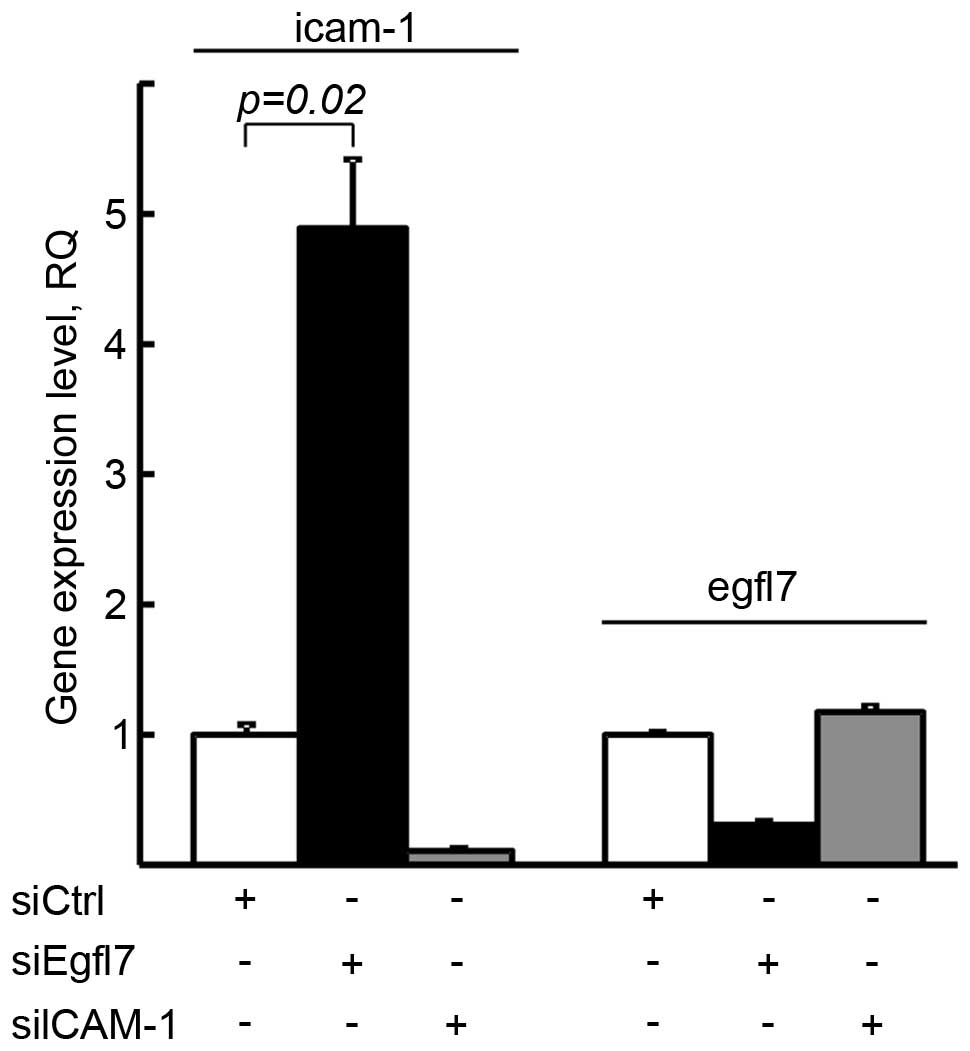
In order to assess which gene between ICAM-1 and
egfl7 regulated the expression of the other one, we deregulated
either egfl7 or ICAM-1 in human primary endothelial cells by RNA
interference and checked for the expression of the other gene.
Targeting egfl7, which lowered its expression by 69.4%, induced a
4.9-fold increase in ICAM-1 expression levels in the same cells. On
the other hand, the downregulation of ICAM-1 expression levels by
89.3% did not significantly affect the expression levels of egfl7
in the same cells (Fig. 4). Thus,
these results show that egfl7 constitutively represses the
expression of ICAM-1 in endothelial cells but that ICAM-1 does not
regulate egfl7 expression.
Discussion
We have previously shown that egfl7 promotes the
escape of tumors from immunity by repressing the activation of
endothelial cells in mice (15). In
this previous study, an inverse correlation between egfl7 protein
levels and those of ICAM-1 were demonstrated in a limited number of
human breast cancer lesions. The present study represents the first
study of the correlation between the expression of egfl7 and that
of ICAM-1 in a large population of patients which allowed for
statistical analysis. This population formed a representative
sample of breast cancer subtypes close to what is typically
described by pathologists in the global population, where 64% are
IDC, 10% are ILC and 7% are of other types (mucinous, tubular) and
where among the IDC, 19% show a Scarf-Bloom-Richardson grade equal
to 3 and are positive for hormonal receptors in 85% of cases, while
HER2 overexpression is detected in 10–15% of the cases (18). Since egfl7 is a secreted protein which
can accumulate in distant tissues from the producing cells
(16), egfl7 transcripts were
selected to analyze using in situ hybridization rather than
following the distribution of the egfl7 protein as before. Indeed,
transcript analysis provides a direct visualization of
egfl7-expressing cells in tumor samples. Furthermore, the study was
focused on the activation of endothelial cells in the tumor
microenvironment, i.e. in peri-tumoral areas of human breast cancer
lesions and on a possible correlation between endothelium
activation and expression levels of egfl7. For that matter, the
analyzed area selected was in the periphery of the main
infiltrating lesion, not close to an in situ carcinoma focus
or a glandular islet, not within a necrotic zone, nor within any
healing zone consecutive to biopsy, so as to avoid any possible
inflammation and activation of blood vessels other than those due
to the presence of the main tumor. In order to identify vascular
vessels without any ambiguity, the choice of a good histological
marker was important. CD34 is a transmembrane glycoprotein
expressed in hematopoietic stem cells, endothelial cells, in
fibroblasts, and in stromal cells. It is commonly used as a marker
for endothelia in pathology labs, though it is not specific to this
tissue (19). CD31 (or
platelet-endothelial cell adhesion molecule-1) is a cell membrane
protein expressed by endothelial cells, platelets, and
hematopoietic cells. It is more particularly expressed in
endothelial cells (17) and is widely
used for visualizing these cells in animal models. It should
however be noted that although both markers reveal endothelial
cells, neither one is strictly specific for blood vessels. The
positive identification of blood vessels was therefore achieved by
crossing the CD31 staining information with the identification of
vessels by the breast cancer pathologist and the oncologist on the
base of morphological criteria, so as to exclude any potential
non-vascular CD31+ structure.
Previous studies have addressed the expression of
ICAM-1 in breast cancer lesions but with different aims (20–23).
Higher levels were correlated with a more aggressive tumor
phenotype (20) and an increased
ICAM-1 staining has been observed in blood vessels of breast cancer
tissues compared to normal tissues (21). Furthermore, fewer numbers of
leukocytes infiltrated in ductal breast carcinoma were also
associated with lower levels of ICAM-1 expression on tumor
endothelial cells (22). More
recently, ICAM-1 has been identified as a marker for triple
negative breast cancers and shown to be a promising target for
treating these tumors (23). All
these studies were focused on the expression of ICAM-1 in the tumor
but did not specifically address the expression of ICAM-1 in the
tumor microenvironment excluding the tumor area in itself, thus
leading to different results. We have previously reported that
egfl7 protein expression was associated with improved prognosis
factors in human breast cancer lesions (24) but this previous study also focused on
egfl7 expression in cancer areas, not peritumoral areas focused on
in the present study. A correlation between egfl7 protein
expression in tumoral foci with lower grade and hormonal receptor
expression was identified (24), but
there was no evidence that expression of egfl7 transcripts
in peritumoral vessels could be correlated with any of these
prognosis factors, thus confirming the fact that the localization
of egfl7 protein secreted within tumor areas and the expression of
its transcripts in peri-tumoral blood vessels are apparently not
linked. Notably, in the present study, egfl7 transcripts were
identified in cancer cells, demonstrating that these cells also
produce egfl7 although it is predominantly an endothelial gene in
normal tissues (8–10). This suggests that the Egfl7 protein
detected in the earlier study (24)
was at least in part produced by cancer cells themselves.
The most interesting observation made in the present
study was the inverse correlation between the expression levels of
egfl7 and those of ICAM-1 in the tumor microenvironment. This
correlation was due to the fact that egfl7 regulates the expression
of ICAM-1 in endothelial cells and not the reverse. This confirmed
previous observations that egfl7 negatively regulates the
expression of ICAM-1 (15) while, on
the other hand, the potential effects of ICAM-1 on egfl7 expression
have never previously been reported. ICAM-1 expression is mainly
regulated by the NFkB and the MAPK/Erk pathways (25–28). It is
most probable that Egfl7 regulates the expression of ICAM-1 through
the NFkB pathway in the breast cancer microenvironment similarly to
in coronary endothelial cells where Egfl7 treatment reduced a
cyclosporin A-induced increase in NFκB activity and ICAM-1
expression (29), but such regulation
is not yet demonstrated.
The results of the present study underline the
important role served by egfl7 in the regulation of the activation
of blood vessels in cancer lesions. They suggest that egfl7 may be
a valuable therapeutic target to consider in order to enhance the
efficiency of immune therapies against cancer. As a matter of fact,
targeting egfl7 has been shown to increase tumor blood vessel
damage and to enhance the tumor response to anti-VEGF treatment in
various murine models (30) but the
effects on the infiltration of immune cells into the tumors has not
been assessed. Whether an anti-egfl7 therapy would increase the
efficiency of immunotherapies in treating cancer remains to be
assessed. Understanding the mechanisms of endothelium activation in
cancer may allow for an improved selection of patients which could
benefit from immunotherapies. Regarding breast cancer, this would
be particularly pertinent for the highly heterogeneous triple
negative population, which would appear to be a good candidate for
such refinement in diagnosis.
Acknowledgements
The present study was supported by Ligue Régionale
contre le Cancer, Institut National du Cancer (grant no.
INCA_6598), and Fondation ARC (grant no. SFI20111203644). Dr
Fabrice Soncin is Directeur de Recherche of the Institut National
de la Santé et de la Recherche Médicale.
Glossary
Abbreviations
Abbreviations:
|
B2M
|
β2-microglobulin
|
|
DCIS
|
ductal carcinoma in situ
|
|
egfl7
|
EGF-like domain-containing protein
7
|
|
ICAM-1
|
intercellular adhesion molecule-1
|
|
IDC
|
invasive ductal carcinoma
|
|
ILC
|
invasive lobular carcinoma
|
|
siRNA
|
small interfering RNA
|
|
TBS
|
TRIS buffer saline
|
|
VCAM-1
|
vascular cell adhesion molecule-1
|
References
|
1
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen DS and Mellman I: Oncology meets
immunology: The cancer-immunity cycle. Immunity. 39:1–10. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Topalian SL, Hodi FS, Brahmer JR,
Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD,
Sosman JA, Atkins MB, et al: Safety, activity, and immune
correlates of anti-PD-1 antibody in cancer. N Engl J Med.
366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brahmer J, Reckamp KL, Baas P, Crinò L,
Eberhardt WE, Poddubskaya E, Antonia S, Pluzanski A, Vokes EE,
Holgado E, et al: Nivolumab versus docetaxel in advanced
squamous-cell non-small-cell lung cancer. N Engl J Med.
373:123–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Muller WA: Mechanisms of transendothelial
migration of leukocytes. Circ Res. 105:223–230. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Herberman RB, Nunn ME, Holden HT and
Lavrin DH: Natural cytotoxic reactivity of mouse lymphoid cells
against syngeneic and allogeneic tumors. II. Characterization of
effector cells. Int J Cancer. 16:230–239. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shrikant P and Mescher MF: Control of
syngeneic tumor growth by activation of CD8+ T cells: Efficacy is
limited by migration away from the site and induction of
nonresponsiveness. J Immunol. 162:2858–2866. 1999.PubMed/NCBI
|
|
8
|
Soncin F, Mattot V, Lionneton F, Spruyt N,
Lepretre F, Begue A and Stehelin D: VE-statin, an endothelial
repressor of smooth muscle cell migration. Embo J. 22:5700–5711.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Parker LH, Schmidt M, Jin SW, Gray AM,
Beis D, Pham T, Frantz G, Palmieri S, Hillan K, Stainier DY, et al:
The endothelial-cell-derived secreted factor Egfl7 regulates
vascular tube formation. Nature. 428:754–758. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fitch MJ, Campagnolo L, Kuhnert F and
Stuhlmann H: Egfl7, a novel epidermal growth factor-domain gene
expressed in endothelial cells. Dev Dyn. 230:316–324. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Diaz R, Silva J, Garcia JM, Lorenzo Y,
García V, Peña C, Rodríguez R, Muñoz C, García F, Bonilla F and
Domínguez G: Deregulated expression of miR-106a predicts survival
in human colon cancer patients. Genes Chromosomes Cancer.
47:794–802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu F, Yang LY, Li YF, Ou DP, Chen DP and
Fan C: Novel role for epidermal growth factor-like domain 7 in
metastasis of human hepatocellular carcinoma. Hepatology.
50:1839–1850. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang CH, Li XJ, Zhou YZ, Luo Y, Li C and
Yuan XR: Expression and clinical significance of Egfl7 in malignant
glioma. J Cancer Res Clin Oncol. 136:1737–1743. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou L, Li J, Zhao YP, Guo JC, Cui QC,
Zhou WX, Zhang TP, Wu WM, You L and Shu H: Prognostic significance
of epidermal growth factor-like domain 7 in pancreatic cancer.
Hepatobiliary Pancreat Dis. 13:523–528. 2014. View Article : Google Scholar
|
|
15
|
Delfortrie S, Pinte S, Mattot V, Samson C,
Villain G, Caetano B, Lauridant-Philippin G, Baranzelli MC,
Bonneterre J, Trottein F, et al: Egfl7 promotes tumor escape from
immunity by repressing endothelial cell activation. Cancer Res.
71:7176–7186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lelièvre E, Hinek A, Lupu F, Buquet C,
Soncin F and Mattot V: VE-statin/egfl7 regulates vascular
elastogenesis by interacting with lysyl oxidases. Embo J.
27:1658–1670. 2008.Can't. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miettinen M: Immunohistochemistry of soft
tissue tumours - review with emphasis on 10 markers.
Histopathology. 64:101–118. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Penault-Llorca F and Arnould L: Adjuvant
breast cancer: Which clinical and pathological characteristics in
2007? Bull Cancer. 97:1421–1426. 2010.(In French). PubMed/NCBI
|
|
19
|
Fina L, Molgaard HV, Robertson D, Bradley
NJ, Monaghan P, Delia D, Sutherland DR, Baker MA and Greaves MF:
Expression of the CD34 gene in vascular endothelial cells. Blood.
75:2417–2426. 1990.PubMed/NCBI
|
|
20
|
Schröder C, Witzel I, Müller V, Krenkel S,
Wirtz RM, Jänicke F, Schumacher U and Milde-Langosch K: Prognostic
value of intercellular adhesion molecule (ICAM)-1 expression in
breast cancer. J Cancer Res Clin Oncol. 137:1193–1201. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fox SB, Turner GD, Leek RD, Whitehouse RM,
Gatter KC and Harris AL: The prognostic value of quantitative
angiogenesis in breast cancer and role of adhesion molecule
expression in tumor endothelium. Breast Cancer Res Treat.
36:219–226. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bouma-ter Steege JC, Baeten CIM, Thijssen
VL, Satijn SA, Verhoeven IC, Hillen HF, Wagstaff J and Griffioen
AW: Angiogenic profile of breast carcinoma determines leukocyte
infiltration. Clin Cancer Res. 10:7171–7178. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo P, Huang J, Wang L, Jia D, Yang J,
Dillon DA, Zurakowski D, Mao H, Moses MA and Auguste DT: ICAM-1 as
a molecular target for triple negative breast cancer. Proc Natl
Acad Sci USA. 111:14710–14715. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Philippin-Lauridant G, Baranzelli M-C,
Samson C, Fournier C, Pinte S, Mattot V, Bonneterre J and Soncin F:
Expression of Egfl7 correlates with low-grade invasive lesions in
human breast cancer. Int J Oncol. 42:1367–1375. 2013.PubMed/NCBI
|
|
25
|
Rao VN, Huebner K, Isobe M, Ar RA, Croce
CM and Reddy ES: elk, tissue-specific ets-related genes on
chromosomes X and 14 near translocation breakpoints. Science.
244:66–70. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kuldo JM, Westra J, Asgeirsdóttir SA, Kok
RJ, Oosterhuis K, Rots MG, Schouten JP, Limburg PC and Molema G:
Differential effects of NF-{kappa}B and p38 MAPK inhibitors and
combinations thereof on TNF-{alpha}- and IL-1{beta}-induced
proinflammatory status of endothelial cells in vitro. Am J Physiol
Cell Physiol. 289:C1229–C1239. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Maeng YS, Min JK, Kim JH, Yamagishi A,
Mochizuki N, Kwon JY, Park YW, Kim YM and Kwon YG: ERK is an
anti-inflammatory signal that suppresses expression of
NF-kappaB-dependent inflammatory genes by inhibiting IKK activity
in endothelial cells. Cell Signal. 18:994–1005. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou Z, Connell MC and MacEwan DJ:
TNFR1-induced NF-kappaB, but not ERK, p38MAPK or JNK activation,
mediates TNF-induced ICAM-1 and VCAM-1 expression on endothelial
cells. Cell Signal. 19:1238–1248. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Badiwala MV, Guha D, Tumiati L, Joseph J,
Ghashghai A, Ross HJ, Delgado DH and Rao V: Epidermal growth
factor-like domain 7 is a novel inhibitor of neutrophil adhesion to
coronary artery endothelial cells injured by calcineurin
inhibition. Circulation. 124(11 Suppl): S197–S203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Johnson L, Huseni M, Smyczek T, Lima A,
Yeung S, Cheng JH, Molina R, Kan D, De Mazière A, Klumperman J, et
al: Anti-Egfl7 antibodies enhance stress-induced endothelial cell
death and anti-VEGF efficacy. J Clin Invest. 123:3997–4009. 2013.
View Article : Google Scholar : PubMed/NCBI
|