Introduction
The mutation of RAS is the most frequent
event during the development of human cancer (1,2). Four
closely related isoforms of the Ras protein exist and consist of
H-Ras, K(A)-Ras, K(B)-Ras and N-Ras (3,4). Mutations
of distinct Ras isoforms are detected in an organ-specific manner
and have different cellular functions (3,4). To be
functionally active, the Ras protein must be associated with
cellular membranes through several post-translational modifications
(5). Farnesyltransferase (FTase) is
an essential enzyme for farnesylation on the cysteine of the CAAX
motif of cytosolic Ras, resulting in the irreversible step of Ras
membrane association (6).
Membrane-anchored Ras protein becomes activated by cycling between
the GDP-bound inactive state and the GTP-bound active state
(5).
Farnesylation by FTase is the first irreversible,
rate-limiting step for Ras membrane association and an obligate
modification for oncogenic Ras biological activity (7). Initial attempts to inhibit FTase have
been made in the development of anti-cancer agents (2,8,9). In vitro and in vivo tumor
models have demonstrated that FTase inhibitors (FTIs) are potent
against various forms of cancer (2,10). FTI
treatment has also been reported to be effective against
H-Ras-transformed cells and H-Ras-driven murine tumors (10,11).
Following treatment with the FTI lonafarnib, transgenic mice with
mammary tumors containing a H-Ras mutation underwent complete tumor
regression (10). However, these
drugs failed to increase survival in clinical trials of patients
with K-Ras mutated pancreatic cancer (12). N-Ras and K-Ras become substrates for
geranylgeranyltransferase I, resulting in alternative prenylation
and membrane targeting (9,13). By contrast, H-Ras has no alternative
prenylation for membrane targeting. Therefore, FTIs are emerging as
an effective therapeutic approach for targeting H-Ras in cancer
(9,13). FTI-277, an FTI, successfully inhibited
Ras membrane association, resulting in prevention of Ras oncogenic
signaling from the cell membrane (14).
Breast cancer has been estimated as the most
commonly diagnosed form of cancer and the second highest cause of
cancer-associated mortality among women in the United States
(15). The metastatic spread of
breast cancer is a major cause of mortality (16). Despite the low frequency (<5%) of
mutated forms of RAS in breast cancer (13), elevated levels of the Ras protein have
been identified in 60–70% of human primary breast cancer cases
(17). It was previously demonstrated
that H-Ras and N-Ras induced a transformed phenotype in human
breast epithelial MCF10A cells (18).
However, H-Ras, but not N-Ras, led to an invasive phenotype in
MCF10A cells (18). The
H-Ras-specific Rac1-MKK3/6-p38 mitogen-activated protein kinase
pathway is required for breast cell motility and invasion (19–21).
The present study investigated the effect of FTI-277
on H-Ras activation and the migratory/invasive phenotypes of breast
cancer cells. The results demonstrated that FTI-277 inhibited
activation of H-Ras, but not that of N-Ras. In addition, FTI-277
effectively inhibited the migratory/invasive abilities of breast
cancer cells through inhibition of H-Ras activation.
Materials and methods
Cell culture conditions and
reagents
H-Ras-MCF10A cells were established and cultured as
previously described (18). Hs578T
and MDA-MB-231 breast cancer cells were purchased from the Korean
Cell Line Bank (Seoul, Korea). Hs578T and MDA-MB-231 cells were
cultured in Dulbecco's modified Eagles's medium (GE Healthcare Life
Sciences, Salt Lake City, UT, USA) supplemented with 10% fetal
bovine serum (Corning Life Sciences, Cambridge, MA, USA) and 100
µg/ml penicillin-streptomycin as previously described (22). FTI-277 (F9803) was purchased from
Sigma-Aldrich (St. Louis, MO, USA) and dissolved in distilled
water.
Immunoblot analysis
Immunoblot analysis was performed as described
previously (23). Protein extracts in
lysis buffer [50 mM Tris, 2% sodium dodecyl sulfate (SDS), 1 mM
ethylenediaminetetraacetic acid (EDTA) and 0.1 M dithiothreitol)
containing protease inhibitor cocktail (Roche Diagnostics GmbH,
Mannheim, Germany) were subjected to 12% SDS-polyacrylamide gel
electrophoresis. The PVDF membrane (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) was probed with rabbit polyclonal anti-human
H-Ras [dilution, 1:1,000 in phosphate-buffered saline with Tween
(PBST); #sc-520] and anti-N-Ras (dilution, 1:1,000 in PBST; sc-519)
antibodies (Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
WesternBright ECL kit was used for detection (#K-12045-D50;
Advansta Inc., Menlo Park, CA, USA). Relative band intensities were
determined by quantification of each band using the Gel Doc™ XR+
system (Bio-Rad Laboratories, Inc.).
Membrane fractions
Membrane fractions were collected as previously
described (24). Cells were treated
with FTI-277 (50 µM) for 24 h and epidermal growth factor (EGF) (10
ng/ml) for 30 min prior to lysis. Cells were rinsed with PBS
buffer, suspended in homogenization buffer (0.25 M sucrose, 25 mM
Tris-HCl and 10 µg/ml leupeptin, pH 7.4), and homogenized in a
ground-glass dounce homogenizer (Heidolph Instruments, Schwabach,
Germany). The lysates were centrifuged for 10 min at 3,000 × g,
resulting in the pellet and supernatant. The supernatant was
centrifuged again at 12,000 × g for 10 min. The resulting pellet
was suspended in chelating buffer (25 mM Tris-HCl, 5 mM EDTA and 10
µg/ml leupeptin, pH 7.4), followed by centrifugation at 12,000 × g
for 10 min. The resulting pellet (membrane fraction) was washed and
resuspended in incubation buffer (25 mM Tris-HCl and 10 µg/ml
leupeptin, pH 7.4).
Ras activity assay
The level of Ras-GTP was measured by Ras assay
reagent (Merck Millipore, Darmstadt, Germany) following the
manufacturer's protocol, as previously described (22). The cells were treated with 10 ng/ml
EGF (Sigma-Aldrich) for 30 min and lysed in Mg2+
lysis/wash buffer. Raf-1 RBD-agarose was added to the lysates and
incubated for 45 min at 4°C. The bead pellet was washed 3 times
with MLB buffer, boiled in 2X Laemmli sample buffer, and subjected
to immunoblot analysis with anti-H-Ras or anti-N-Ras
antibodies.
MTT assay
MTT assay was performed as previously described
(25). Cells (5×103) were
seeded in a 96-well plate and incubated for 24 h at 37°C. Cells
were then treated with various concentrations of FTI-277 (0, 10, 20
and 50 µM) for 24 or 48 h. Following incubation, 25 µl 5 mg/ml MTT
(Sigma-Aldrich) was added and incubated for 3 h, and formazan was
dissolved in 100 µl dimethyl sulfoxide. The optical density was
measured at 540 nm using a Synergy 2 Multi-Mode Reader (BioTek
Instruments, Inc., Winooski, VT, USA). Each sample was assayed in
triplicate.
In vitro invasion/migration
assays
In vitro invasion/migration assays were
performed using a 24-well Transwell unit with polycarbonate filters
(Corning Life Sciences) as previously described (26). The lower side of the filter was coated
with type I collagen (Sigma-Aldrich) and the upper side of the
filter was coated with Matrigel (BD Biosciences, Bedford, MA, USA).
Cells (5×104 cells) suspended in DMEM supplemented with
100 µg/ml penicillin-streptomycin were placed in the upper chamber.
The lower chamber was filled with DMEM supplemented with 10% fetal
bovine serum and 100 µg/ml penicillin-streptomycin. Following 24 h
of incubation, cells that had invaded the lower side of the filter
were solubilized with 0.5% crystal violet solution and measured at
595 nm using a micro-ELISA reader. Each sample was assayed in
triplicate.
Results
FTI-277 inhibits EGF-induced H-Ras
activation in MDA-MB-231 cells
The present study examined the effect of FTI-277 on
the localization and activation of H-Ras and N-Ras in breast cancer
cells. Membrane fractions were extracted from MDA-MB-231 cells in
which both H-Ras and N-Ras may be activated by EGF (22,27,28).
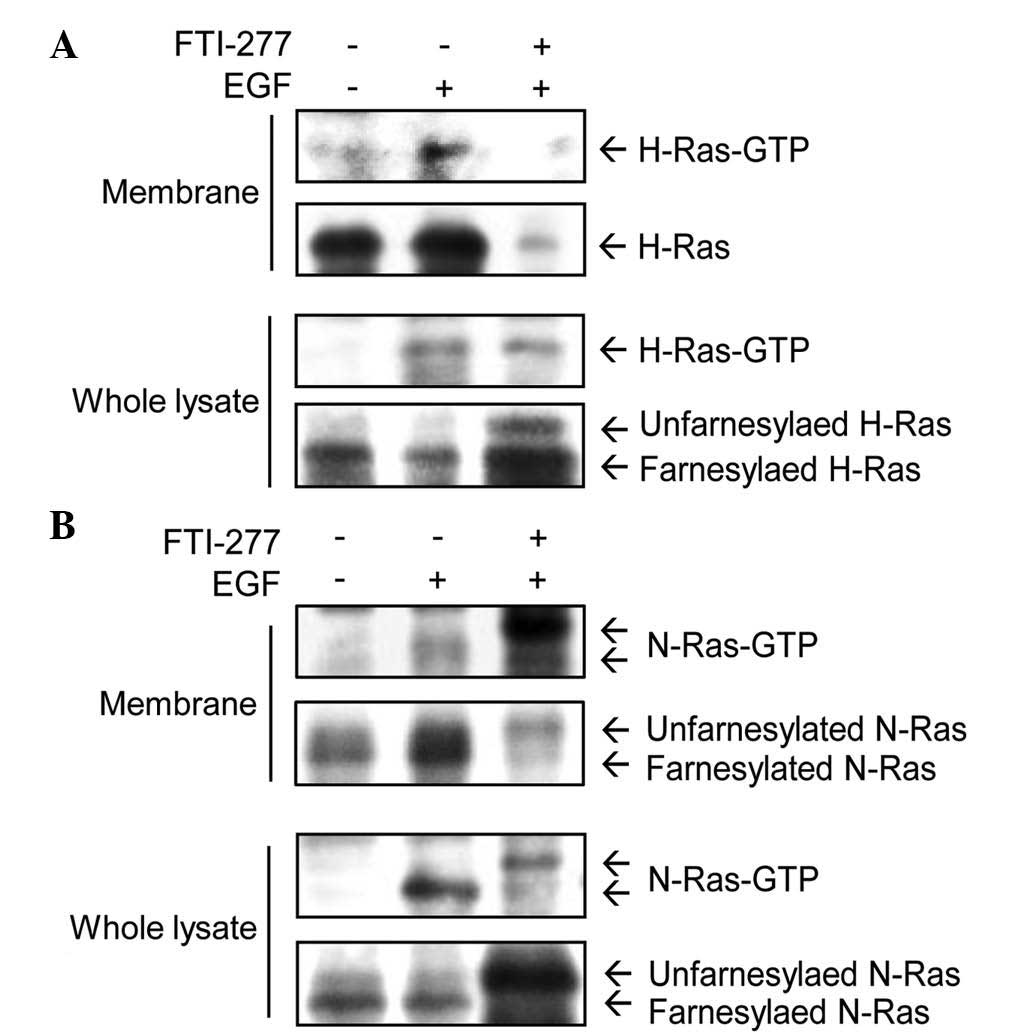
H-Ras-GTP was induced by EGF in the membrane fractions (Fig. 1A). Following treatment with 50 µM
FTI-277 for 24 h, H-Ras-GTP was almost completely abolished in the
membrane fractions. However, in whole cell lysates, FTI-277 did not
inhibit H-Ras-GTP induced by EGF. Unfarnesylated H-Ras (upper band)
was detected following FTI-277 treatment in whole cell lysates, but
not in the membrane fractions (Fig.
1A). These results indicate that FTI-277 inhibited
farnesylation, and therefore also inhibited the membrane
localization of H-Ras in MDA-MB-231 cells.
EGF treatment induced activation of N-Ras in
membrane fractions and whole cell lysates (Fig. 1B). The level of N-Ras-GTP in membrane
fractions was not decreased by treatment with FTI-277. These
results suggest that N-Ras has an alternative prenylation for
membrane targeting, unlike H-Ras. In combination, these data
demonstrate that FTI-277 inhibited the EGF-induced activation of
H-Ras, but not that of N-Ras, in membrane fractions of MDA-MB-231
cells.
FTI-277 exerts an anti-proliferative
effect in breast cells expressing active H-Ras
The current study subsequently examined the effect
of FTI-277 on the proliferation of breast cells. H-Ras-activated
breast H-Ras-MCF10A cell lines, in which a constitutively active
mutant of H-Ras (G12D) was stably introduced (18), and Hs578T cells, which endogenously
express active H-Ras (G12D), were analyzed (29). MDA-MB-231 cells harboring wild-type
H-Ras and N-Ras were also used (30).
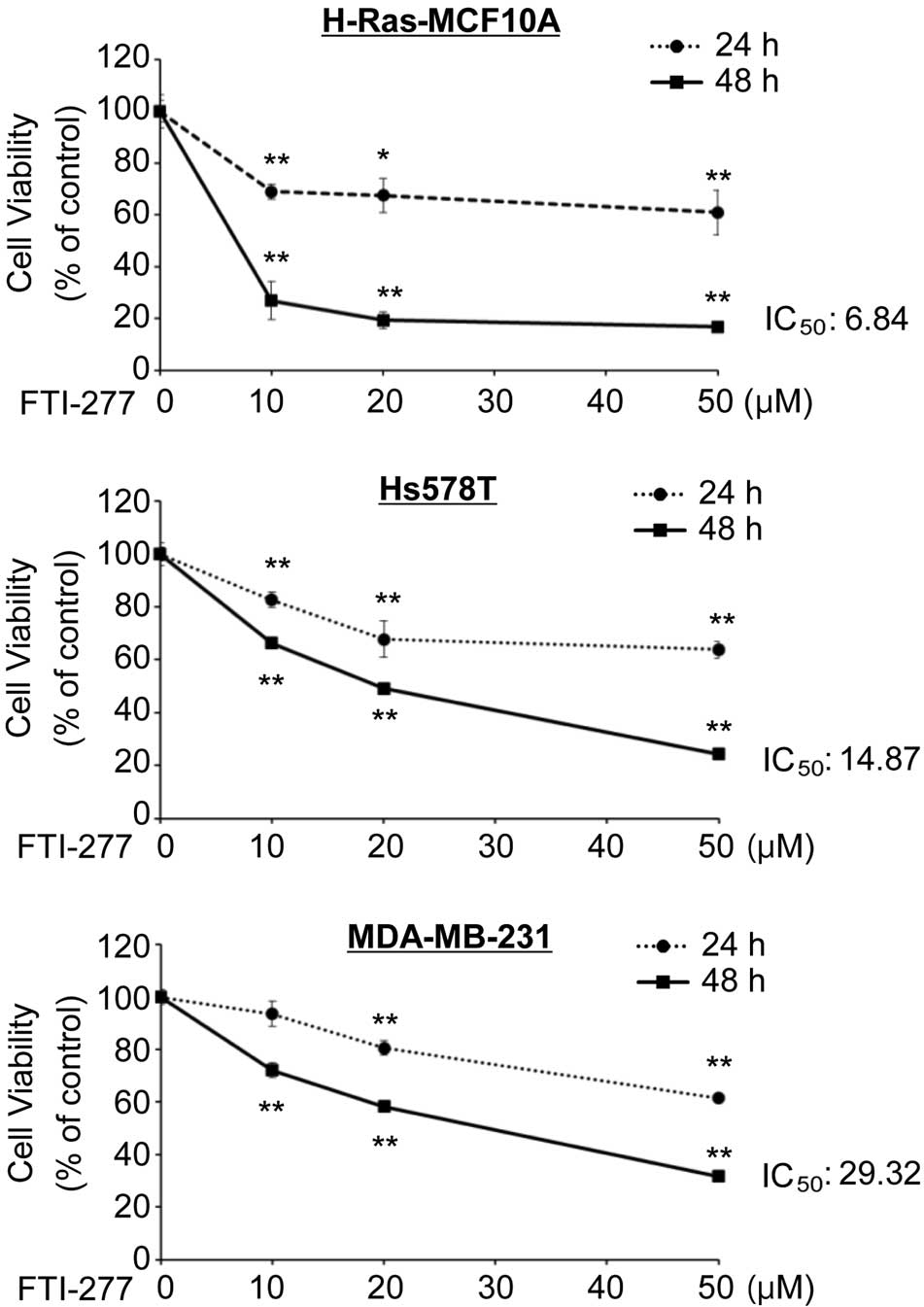
MTT assay demonstrated that FTI-277 inhibited proliferation of the
H-Ras-MCF10A, Hs578T and MDA-MB-231 cells in a dose-dependent
manner (Fig. 2). FTI-277 exerted a
strong anti-proliferative effect on the H-Ras-MCF10A and Hs578T
cells with 50% inhibitory concentration (IC50) values of
6.84 and 14.87 µM for 48 h, respectively. FTI-277 treatment
inhibited proliferation of the MDA-MB-231 cells with an
IC50 value of 29.32 µM for 48 h. The results suggest
that breast cells in which H-Ras is activated may be more
susceptible to FTI-277 compared with the cells with wild-type
H-Ras.
FTI-277 inhibits invasive and
migratory phenotypes of breast cells expressing active H-Ras
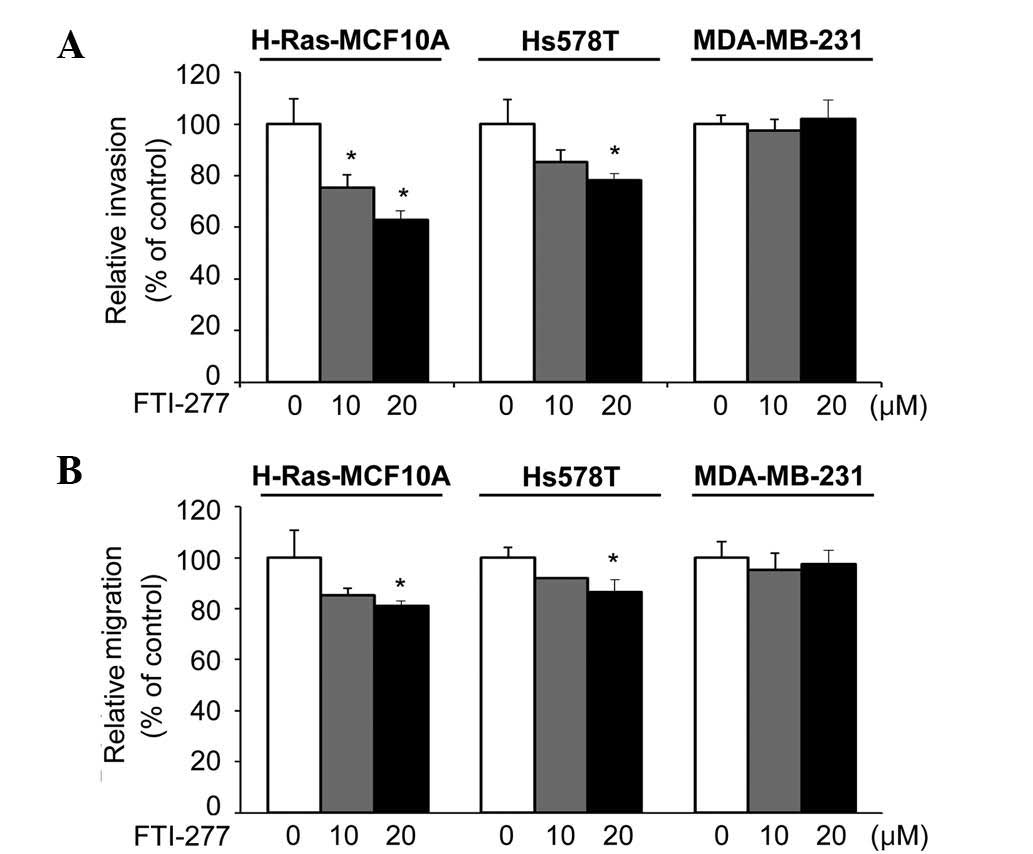
Next, the present study investigated the effect of
FTI-277 on the invasive phenotypes of H-Ras-MCF10A, Hs578T and
MDA-MB-231 cells, which have been previously demonstrated to be
highly invasive (18,19,30). The
invasiveness of the H-Ras-MCF10A and Hs578T cells was significantly
inhibited by FTI-277 (Fig. 3A). By
contrast, FTI-277 did not affect the invasive ability of the
MDA-MB-231 cells.
Similarly, FTI-277 treatment significantly inhibited
the migratory phenotypes of the H-Ras-MCF10A and Hs578T cells, but
not that of the MDA-MB-231 cells (Fig.
3B). Given that the active state of H-Ras is important in
invasive phenotypes of H-Ras-MCF10A and Hs578T cells (18,19,22), the
results suggest that FTI-277 may inhibit the invasive and migratory
abilities of breast cells through inhibition of H-Ras.
FTI-277 inhibits EGF-induced invasion
of MDA-MB-231 cells
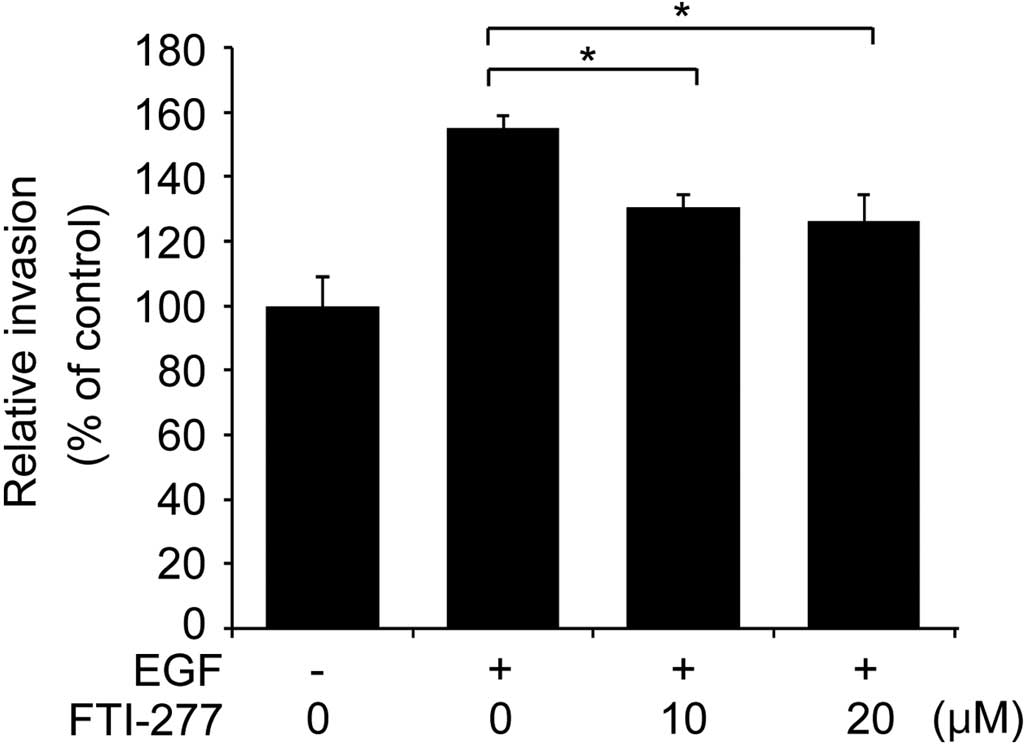
To further examine if the inhibitory effect of
FTI-277 on breast cell invasion involves inhibition of H-Ras
activation, the present study analyzed whether FTI-277 affects the
invasive phenotype of MDA-MB-231 cells treated with EGF. As
presented in Fig. 4, the invasive
phenotype of the MDA-MB-231 cells was enhanced by EGF stimulation.
FTI-277 significantly decreased the EGF-enhanced invasion of the
MDA-MB-231 cells in a concentration-dependent manner (Fig. 4). Given that FTI-277 inhibited the
EGF-induced H-Ras activation in MDA-MB-231 cells (Fig. 1A), the present data indicates that
FTI-277 inhibits breast cell invasion, possibly through inhibition
of H-Ras activation.
Discussion
FTase has been recently suggested as a drug target
in the development of anti-cancer therapy (7,9). In the
present study, it was demonstrated that FTI-277 exerted a more
potent anti-proliferative effect on breast cells expressing an
active mutant of H-Ras (H-Ras-MCF10A and Hs578T cells) than breast
cells expressing wild-type H-Ras (MDA-MB-231 cells). Although H-Ras
mutation is infrequent compared with K-Ras or N-Ras (13), the current data suggests that FTIs may
serve as an effective strategy for targeting H-Ras-mediated
proliferation of breast cancer cells. A promising FTI, tipifarnib,
demonstrated anti-cancer activity against breast cancer in a phase
II study, which warrants further study (9).
In addition, the present study observed that FTI-277
treatment markedly decreased the level of H-Ras-GTP, but not that
of N-Ras-GTP, in membrane fractions of MDA-MB-231 cells stimulated
by EGF. As Ras proteins trigger signaling pathways and activate
downstream effector molecules when they are located in membrane
(31), the results imply that FTI-277
effectively blocked H-Ras activation by inhibiting H-Ras
trafficking to the cell membrane of breast cancer cells.
Furthermore, in whole cell lysates, FTI-277 was not
able to inhibit H-Ras-GTP or N-Ras-GTP in the current study. It was
previously reported that FTI-277 induced the accumulation of
Ras-Raf complexes in the cytoplasm where Ras is GTP-bound, but Raf
kinase was not activated (14).
Hence, it is plausible that the H-Ras-GTP in the cytoplasm of
MDA-MB-231 cells may not be active.
There is increasing evidence that FTIs inhibit the
migration, invasion and metastasis of various forms of cancer,
including breast, head and neck, pancreatic and colon cancer
(32–35). The present study demonstrated that
FTI-277 effectively inhibited the invasive/migratory phenotypes of
H-Ras-MCF10A and Hs578T cells, but not those of MDA-MB-231 cells.
Consistent with these results, a previous study observed that
FTI-277 did not inhibit the Transwell invasion of MDA-MB-231 cells
(36). By contrast, it has been
reported that the transendothelial invasion of MDA-MB-231 cells was
mildly inhibited by FTI-277 (33).
These results suggest that FTI-277 exerts mild or no effect on the
invasion of MDA-MB-231 cells without EGF stimulation, depending on
the different methods of invasion assays.
Notably, EGF-enhanced invasion of MDA-MB-231 cells
was significantly decreased by FTI-277 treatment in the current
study. Given that EGF enhances the invasive phenotypes of
MDA-MB-231 cells by activation of H-Ras and K-Ras (22), these results indicate that FTI-277 may
partially inhibit EGF-increased invasion mediated by H-Ras.
In conclusion, the present study demonstrated that
FTI-277 exerted a potent inhibitory effect on the proliferation and
invasive/migratory abilities of breast cells expressing active
H-Ras. In addition, FTI-277 inhibited EGF-induced H-Ras activation
and invasion of breast cancer cells. This suggests that FTase
inhibition may be an effective therapeutic approach for targeting
H-Ras-mediated proliferation, migration and invasion of breast
cells.
Acknowledgements
The present study was supported by the Priority
Research Centers Program (grant no. 2016R1A6A1A03007648) and the
Bio & Medical Technology Development Program (grant no.
2015M3A9B6074045) of the National Research Foundation of Korea
funded by the Korean government.
References
|
1
|
Barbacid M: Ras genes. Annu Rev Biochem.
56:779–827. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Appels NM, Beijnen JH and Schellens JH:
Development of farnesyl transferase inhibitors: A review.
Oncologist. 10:565–578. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carbone A, Gusella GL, Radzioch D and
Varesio L: Human Harvey-ras is biochemically different from
Kirsten- or N-ras. Oncogene. 6:731–737. 1991.PubMed/NCBI
|
|
4
|
Maher J, Baker DA, Manning M, Dibb NJ and
Roberts IA: Evidence for cell-specific differences in
transformation by N-, H- and K-ras. Oncogene. 11:1639–1647.
1995.PubMed/NCBI
|
|
5
|
Hancock JF and Parton RG: Ras plasma
membrane signalling platforms. Biochem J. 389:1–11. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Reiss Y, Goldstein JL, Seabra MC, Casey PJ
and Brown MS: Inhibition of purified p21ras farnesyl: Protein
transferase by Cys-AAX tetrapeptides. Cell. 62:81–88. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cox AD, Der CJ and Philips MR: Targeting
RAS membrane association: Back to the future for Anti-RAS drug
discovery? Clin Cancer Res. 21:1819–1827. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Brunner TB, Hahn SM, Gupta AK, Muschel RJ,
McKenna WG and Bernhard EJ: Farnesyltransferase inhibitors: An
overview of the results of preclinical and clinical investigations.
Cancer Res. 63:5656–5668. 2003.PubMed/NCBI
|
|
9
|
Vasan N, Boyer JL and Herbst RS: A RAS
renaissance: Emerging targeted therapies for KRAS-mutated non-small
cell lung cancer. Clin Cancer Res. 20:3921–3930. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu M, Bryant MS, Chen J, Lee S, Yaremko
B, Lipari P, Malkowski M, Ferrari E, Nielsen L, Prioli N, et al:
Antitumor activity of SCH 66336, an orally bioavailable tricyclic
inhibitor of farnesyl protein transferase, in human tumor xenograft
models and wap-ras transgenic mice. Cancer Res. 58:4947–4956.
1998.PubMed/NCBI
|
|
11
|
Kohl NE, Omer CA, Conner MW, Anthony NJ,
Davide JP, deSolms SJ, Giuliani EA, Gomez RP, Graham SL, Hamilton
K, et al: Inhibition of farnesyltransferase induces regression of
mammary and salivary carcinomas in ras transgenic mice. Nat Med.
1:792–797. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Van Cutsem E, van de Velde H, Karasek P,
Oettle H, Vervenne WL, Szawlowski A, Schoffski P, Post S, Verslype
C, Neumann H, et al: Phase III trial of gemcitabine plus tipifarnib
compared with gemcitabine plus placebo in advanced pancreatic
cancer. J Clin Oncol. 22:1430–1438. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cox AD, Fesik SW, Kimmelman AC, Luo J and
Der CJ: Drugging the undruggable RAS: Mission possible? Nat Rev
Drug Discov. 13:828–851. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lerner EC, Qian Y, Blaskovich MA, Fossum
RD, Vogt A, Sun J, Cox AD, Der CJ, Hamilton AD and Sebti SM: Ras
CAAX peptidomimetic FTI-277 selectively blocks oncogenic Ras
signaling by inducing cytoplasmic accumulation of inactive Ras-Raf
complexes. J Biol Chem. 270:26802–26806. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hudis CA and Gianni L: Triple-negative
breast cancer: An unmet medical need. Oncologist. 16(Suppl 1):
S1–S11. 2011. View Article : Google Scholar
|
|
17
|
Clair T, Miller WR and Cho-Chung YS:
Prognostic significance of the expression of a ras protein with a
molecular weight of 21,000 by human breast cancer. Cancer Res.
47:5290–5293. 1987.PubMed/NCBI
|
|
18
|
Moon A, Kim MS, Kim TG, Kim SH, Kim HE,
Chen YQ and Kim HR: H-ras, but not N-ras, induces an invasive
phenotype in human breast epithelial cells: A role for MMP-2 in the
H-ras-induced invasive phenotype. Int J Cancer. 85:176–181. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim MS, Lee EJ, Kim HR and Moon A: p38
kinase is a key signaling molecule for H-Ras-induced cell motility
and invasive phenotype in human breast epithelial cells. Cancer
Res. 63:5454–5461. 2003.PubMed/NCBI
|
|
20
|
Shin I, Kim S, Song H, Kim HR and Moon A:
H-Ras-specific activation of Rac-MKK3/6-p38 pathway: Its critical
role in invasion and migration of breast epithelial cells. J Biol
Chem. 280:14675–14683. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song H, Ki SH, Kim SG and Moon A:
Activating transcription factor 2 mediates matrix
metalloproteinase-2 transcriptional activation induced by p38 in
breast epithelial cells. Cancer Res. 66:10487–10496. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Koh MS and Moon A: Activation of H-Ras and
Rac1 correlates with epidermal growth factor-induced invasion in
Hs578T and MDA-MB-231 breast carcinoma cells. Biochem Biophys Res
Commun. 406:25–29. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee HM and Moon A: Amygdalin regulates
apoptosis and adhesion in Hs578T triple-negative breast cancer
cells. Biomol Ther (Seoul). 24:62–66. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kang S, Kim ES and Moon A: Simvastatin and
lovastatin inhibit breast cell invasion induced by H-Ras. Oncol
Rep. 21:1317–1322. 2009.PubMed/NCBI
|
|
25
|
Kim YT, Kim SK, Jeon YJ and Park SJ:
Seahorse-derived peptide suppresses invasive migration of HT1080
fibrosarcoma cells by competing with intracellular α-enolase for
plasminogen binding and inhibiting uPA-mediated activation of
plasminogen. BMB Rep. 47:691–696. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peng Y, Zhong Y and Li G: Tubeimoside-1
suppresses breast cancer metastasis through downregulation of CXCR4
chemokine receptor expression. BMB Rep. May 9–2016.(Epub ahead of
print). PubMed/NCBI
|
|
27
|
Egan SE, Giddings BW, Brooks MW, Buday L,
Sizeland AM and Weinberg RA: Association of Sos Ras exchange
protein with Grb2 is implicated in tyrosine kinase signal
transduction and transformation. Nature. 363:45–51. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ehrhardt A, David MD, Ehrhardt GR and
Schrader JW: Distinct mechanisms determine the patterns of
differential activation of H-Ras, N-Ras, K-Ras 4B, and M-Ras by
receptors for growth factors or antigen. Mol Cell Biol.
24:6311–6323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kraus MH, Yuasa Y and Aaronson SA: A
position 12-activated H-ras oncogene in all HS578T mammary
carcinosarcoma cells but not normal mammary cells of the same
patient. Proc Natl Acad Sci USA. 81:5384–5388. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hollestelle A, Elstrodt F, Nagel JH,
Kallemeijn WW and Schutte M: Phosphatidylinositol-3-OH kinase or
RAS pathway mutations in human breast cancer cell lines. Mol Cancer
Res. 5:195–201. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Prior IA and Hancock JF:
Compartmentalization of Ras proteins. J Cell Sci. 114:1603–1608.
2001.PubMed/NCBI
|
|
32
|
Oh SH, Kang JH, Kyu Woo J, Lee OH, Kim ES
and Lee HY: A multiplicity of anti-invasive effects of farnesyl
transferase inhibitor SCH66336 in human head and neck cancer. Int J
Cancer. 131:537–547. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kusama T, Mukai M, Tatsuta M, Nakamura H
and Inoue M: Inhibition of transendothelial migration and invasion
of human breast cancer cells by preventing geranylgeranylation of
Rho. Int J Oncol. 29:217–223. 2006.PubMed/NCBI
|
|
34
|
Kusama T, Mukai M, Tatsuta M, Matsumoto Y,
Nakamura H and Inoue M: Selective inhibition of cancer cell
invasion by a geranylgeranyltransferase-I inhibitor. Clin Exp
Metastasis. 20:561–567. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nam JS, Ino Y, Sakamoto M and Hirohashi S:
Ras farnesylation inhibitor FTI-277 restores the E-cadherin/catenin
cell adhesion system in human cancer cells and reduces cancer
metastasis. Jpn J Cancer Res. 93:1020–1028. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Denoyelle C, Hong L, Vannier JP, Soria J
and Soria C: New insights into the actions of bisphosphonate
zoledronic acid in breast cancer cells by dual RhoA-dependent and
-independent effects. Br J Cancer. 88:1631–1640. 2003. View Article : Google Scholar : PubMed/NCBI
|