Introduction
The existing approaches for glioblastoma multiforme
(GBM) treatment are not very effective. Despite the efforts of
oncologists, only 10% of patients survive >18 month after
diagnosis, and 5-year survival is almost nonexistent. The median
survival time of patients with GBM is 12–15 months (1). One of the reasons that traditional
treatment procedures are ineffective is tumor invasive growth
caused by the high activity rate of cancer stem cells (CSCs). This
invasive process is combined with the high infiltration rate of
tumor cells into brain parenchyma, meaning that radical resection
of the tumor cannot be performed. This is primary reason for
relapse and prolonged tumor growth (1–3).
Traditional treatment methods, which include maximal surgical
resection followed by a combination of radiotherapy and
chemotherapy (usually temozolomide), have almost no effect on tumor
cells infiltrating the brain parenchyma, and there are currently
almost no medication or technologies that are able to eliminate
CSCs effectively. One of the possible ways to improve the treatment
of glioblastoma is to develop biomedical drugs that are able to
control the proliferative and reproductive functions of CSCs. The
development of such medication is more successful when using
patients' own stem cells (SCs) as a basic material (4–6).
SCs have been used in the complex treatment of
malignant tumors for >50 years. The leading clinics of Russia,
Europe and the USA have perfected a method of SC extraction from
the peripheral blood of patients (1).
The cell concentrate is based on separated mononuclear cells with a
high rate of hematopoietic SCs (CD34+/CD45+). The sphere of their
application is not limited to blood system recovery following
high-dose chemotherapy; they are widely used for treating acute
diseases and traumas of the nervous system, and may be of great
interest in neuro-oncology (7,8).
After being introduced into the vasculature, SCs are
able to migrate in a certain direction until they reach the tumor
nidus in the brain. The main source of chemo-attractants for SCs is
damaged nervous tissue and tumor cells themselves. Previous studies
have identified >80 cytokines, growth factors and other ligands
controlling SC migration processes via their corresponding type of
sensory receptors (4,5,9). In the
nidus, SCs can exhibit both an antagonistic and stimulating
influence on the tumor. However, no objective criteria have been
suggested that allow evaluation of the impact of cell
transplantation on the neoplastic process. The most useful
criterion of evaluating this medication effect on CSCs may be
qualitative and quantitative changes that microglia undergo.
First, the transplantation of autologous SCs
extracted from patients' peripheral blood enables the
transformation of monocytes into microglia elements of the tumor
that have an ambiguous role in invasive growth processes. Second,
microglial cells release a large amount of
H2O2, NO and other cytokines that damage
nervous tissue (6). Microglial cell
cultivation together with glioblastoma CSCs intensifies these
abilities, supporting the hypothesis of a controversial association
between microglia and CSCs (10).
Finally, a type of microglia that counteracts invasive tumor
development has been identified (11). It is obvious that histophysiological
evaluation of the influence of microglial cells on glioblastoma
growth will become a crucial factor in understanding the way that
cell culture modification occurs and allow criteria to be proposed
for defining the efficiency of biomedical cell-based treatment.
The present study aimed to understand the nature of
interaction between microglial cells and CSCs in an experimental
glioblastoma model in vivo.
Materials and methods
Ethical approval
The study was approved by the Ethics Committee of
the School of Biomedicine, Far Eastern Federal University
(Vladivostok, Russia; Minutes of Meeting No. 1 of February 2,
2015).
Equipment
Slides were analyzed using an Axio Scope A1
microscope (Carl Zeiss AG, Jena, Germany), a FluoView FV 1200MPE
(Olympus Corporation, Tokyo, Japan) for deeper imaging of
immunohistochemically stained biomaterials, and a Zeiss LSM 710
META confocal (laser scanning) microscope (Carl Zeiss AG) for
histology.
Animals
Mature (3–4 months old) female Wistar rats (n=60)
weighing 200–220 g were obtained from Charles River Laboratories
(Erkrath, Germany) and maintained at our facility at 24°C with
standard atmospheric conditions, and fed with standard pellet rat
feed and tap water as drinking water. On every 10th day of the
experiment, 15 rats were sacrificed following C6 injection.
Sacrifice was conducted as follows: Rats were anesthetized by
intraperitoneal injection with a mixture of Zoletil (Virbac,
Carros, France) and Rometar (Bioveta, Ivanovice na Hané, Czech
Republic) at a ratio of 1:4 (10 mg/kg). Once they reached a
surgical level of anesthesia, their hearts were surgically exposed.
Following incision to the right atrium, the left ventricle was
injected with 10 ml cold 0.9% NaCl solution followed by 20 ml cold
fixative (4% paraformaldehyde in 0.1 M phosphate buffer, pH
7.2).
C6 glioma cell line
The C6 glioma cell line (CCL-107; American Type
Culture Collection, Manassas, VA, USA) was used in the present
study. In comparison with other experimental glioma tumors (CNS1,
RT-2, BT4C, F98, RG2, T9), С6 has the largest proportion of CSCs
(12). The cells were cultivated at
37°C with 5% CO2 in 25-cm2 vials filled with
Dulbecco's modified Eagle medium (DMEM) medium containing 10% fetal
bovine serum (FBS) and Antibiotic-Antimycotic (100X), containing
10,000 IU/ml penicillin, 10,000 µg/ml streptomycin and 25 µg/ml
fungizone (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The culture medium was renewed every 3 days. After reaching
95% confluence, the cells were treated with TrypLE Express Enzyme
solution (Gibco; Thermo Fisher Scientific, Inc.; 12604-021) for 7
min at 37°C, placed into a 15-ml vial and centrifuged for 3 min at
120 × g. The supernatant was removed, DMEM + 10% FBS + 100X
Antibiotic-Antimiotic fresh medium was added to the pellet and the
culture was transferred into a new 25-cm2 culture
vial.
Following suspension in fresh medium, the cells were
counted using a hemocytometer and their viability was determined by
staining with 0.4% trypan blue (Gibco; Thermo Fisher Scientific,
Inc.; 15250-061). Prior to implantation into rat brains, C6 cells
were assigned immunocytochemical characteristics, as follows.
Immunocytochemical characteristics of
C6 glioma cells
Cells were fixed with 4% paraformaldehyde
(Sigma-Aldrich, St. Louis, MO, USA; P6148) for 20 min at 4°C then
washed 3 times for 10 min each with PBS pH 7.4 (0.1 М; cat no.
10010031; Gibco; Thermo Fisher Scientific, Inc.). Cells were then
treated with 0.2% Tween-20 (cat no. P9416; Sigma-Aldrich),
permeabilized with 0.2% Triton X-100 (cat no. T8787; Sigma-Aldrich)
and blocked with 0.3% bovine serum albumin (cat no. A2058;
Sigma-Aldrich) (15 min incubation for each reagent; room
temperature), followed by washing with PBS working solution.
Staining was performed using the following primary
antibodies: p53 mouse (dilution, 1:100; cat no. AHO0152; Novex;
Thermo Fisher Scientific, Inc.), nestin rabbit (dilution, 1:100;
cat no. 5413; Sigma-Aldrich), glial fibrillary acidic protein mouse
(dilution, 1:50; cat no. 7260; Abcam, Cambridge, UK), β III tubulin
mouse (dilution, 1:100; cat no. 7751; Abcam), S100 rabbit (cat no.
868; Abcam; dilution, 1:100) and C-X-C chemokine receptor type 4
(CXCR4) rabbit (cat no. 2074; Abcam; dilution, 1:100) antibodies.
All primary antibodies were incubated for 18 h at 4°C. Cells were
then washed with PBS working solution 3 times for 10 min each and
incubated with anti-rabbit secondary antibodies conjugated with
Alexa 633 (dilution, 1:500; cat no. A21071) or Alexa 488 (Novex;
cat no. A11029; Thermo Fisher Scientific, Inc.; dilution, 1:500)
for 2 h at 37°C. Subsequently, the cells were washed with PBS
working solution 3 times for 10 min each. Nuclei were contrasted
with DAPI (cat no. D1306; Molecular Probes; Thermo Fisher
Scientific, Inc.) for 7 min at 22°C, washed with PBS working
solution 2 times for 10 min and placed in Mowiol 40–88 (cat no.
324590; Sigma-Aldrich). Secondary antibodies Alexa 488 and Alexa
633 were also used alone without primary antibodies as controls.
Primary and secondary antibodies were used according to the
manufacturer's protocol.
Glioblastoma modeling
Animals were anesthetized by injection with a 1:4
ratio mixture of Zoletil 100 and Rometar at a dose of 10 mg/kg.
Soft tissues of the head were dissected in sterile conditions,
according to previously determined coordinates, and a burr hole was
made with a micro drill (1.0 mm diameter; Harvard Bioscience, Inc.,
Holliston, MA, USA).
Using stereotaxic instruments for rats (Nirishige
Ltd., Tokyo, Japan) and rat brain atlas coordinates (anterior,
interaural 2.4 mm; lateral, 3.0 mm; ventral, 4.5 mm) (13), 2×105 C6 glioma cells were
introduced into the caudoputamen in 20 µl DMEM with a Hamilton
syringe (Thermo Fisher Scientific, Inc.) at a rate of 2 µl/min.
Tumor development was monitored once per week by MRI on a BioSpec
scanner (Bruker, Billerica, MA, USA). On the 10th, 20th and 30th
days after implantation, 15 animals were sacrificed, while the
remaining animals were left to assess survivability.
Histological staining
Rats were anesthetized by intraperitoneal injection
with a mixture of Zoletil 100 (Virbac) and Rometar (Bioveta) at a
ratio of 1:4 (10 mg/kg) and transcardially perfused with 10 ml cold
0.9% NaCl saline followed by 20 ml cold fixative (4%
paraformaldehyde in 0.1 M phosphate buffer, pH 7.2). The brains
were immediately removed and post-fixed for 12 h at 4°C in fresh
buffered 4% paraformaldehyde. After rinsing, the brains were
processed and embedded in paraffin, according to standard embedding
techniques. Coronal cuts of the paraffin-embedded brain were made
until the tumor was exposed, after which 7-µm slices were obtained
using a microtome. All three areas of interest (tumor, near-tumor
area and symmetric area of the opposite hemisphere) were present on
each slide. The near-tumor area was defined as the 200-µm of tissue
around the tumor edge. Paraffin-embedded brain sections (7 µm) were
stained with hematoxylin-eosin (Bio Optica, Milano, Italy),
according to the standard procedure for morphological tissue
analysis.
Immunohistochemistry
Paraffinized sections of the brain (7 µm) were
deparaffinized and incubated in 3% hydrogen peroxide to block
endogenous peroxidase prior to immunohistochemical staining. After
three washes in 0.1 M phosphate buffer (pH 7.2), sections were
treated for 60 min in a 2% bovine serum albumin solution (cat no.
SC-2323; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and 0.25%
Triton X-100 (GERBU Biotechnik GmbH, Heidelberg, Germany). Brain
sections were then incubated with primary antibodies on glass
slides in a humidified chamber at 4°C for 24 h. After three washes
with 0.1 M phosphate buffer (pH 7.2), the sections were incubated
in a secondary antibody solution for 60 min. Primary mouse
monoclonal antibodies against proliferating cell nuclear antigen
(PCNA; dilution, 1:2,000; cat no. ab29; Abcam) and transforming
growth factor-β2 (TGF-β2; dilution, 1:100; cat no. ab36495; Abcam);
and rabbit polyclonal antibodies against ionized calcium-binding
adapter molecule-1 (IBA-1; dilution, 1:500; cat no. ab108539;
Abcam), nestin (dilution, 1:100; cat no. N5413; Sigma-Aldrich),
CXCR4 (dilution, 1:500; cat no. ab2074; Abcam) and interleukin-1β
(IL-1β; dilution, 1:1,000; cat no. ab9722; Abcam) were used.
Appropriate anti-rabbit (cat no. PI-1000) and anti-mouse (cat no.
PI-2000) secondary antibodies conjugated to horseradish peroxidase
were used, according to the manufacturer's instructions (dilution,
1:100; Vector Laboratories, Inc., Burlingame, CA, USA). Negative
controls were treated with PBS instead of primary antibody. After
washing with 0.1 M phosphate buffer (pH 7.2), sections were treated
for 5–10 min with two chromogens, DAB Plus (Thermo Fisher
Scientific, Inc.) and NovaRED (Vector Laboratories, Inc.), to
elicit the immunoperoxidase reaction. The sections were washed with
0.1 M phosphate buffer (pH 7.2), dehydrated and embedded in
Toluene-Free Mounting Medium (cat no. CS705; Dako, Glostrup,
Denmark).
Image analysis
The number of PCNA-immunopositive cells was
calculated as a proportion of the total GBM area using the ImageJ
software package (version 1.41; National Institutes of Health,
Bethesda, MD, USA). The Scion Image software package (version
4.0.3; Scion Corporation, Frederick, MD, USA) was used to determine
the immunohistochemically stained area of microglia/macrophage
elements in sections of rat brain using staining of IBA-1.
Different areas of the brain preparations were captured for image
analysis, with at least 80 images captured in each group of
animals. Following image acquisition, all images were saved in the
TIF format (1,080×800 pixels). Micrograph processing included the
following steps: Converting the image to black and white,
background subtraction, contrast enhancement, binarization, noise
reduction and measurement. Image preprocessing included calibration
(pixels translated to mm2) and the first three steps of
micrograph processing. Per-pixel image background subtraction was
used to adjust image brightness. The standard morphological filters
Erode and Dilate were used to reduce background noise. Erode
removes background irregularities as well as pieces of the cell
soma and processes that extend beyond the slice thickness. Dilate
returns the cell area to a value comparable to the area before
applying the Erode filter. Image binarization was necessary to
correct filter operations and the area of objects (positive
staining) present in the area of interest was used as the unit of
measurement.
Statistical analysis
The data obtained by immunohistochemistry were
subjected to statistical analysis using one-way analysis of
variance followed by Tukey's post-hoc test. Data are expressed as
the mean ± standard error of the mean, with each experiment
repeated three times. P<0.05 was considered to indicate a
statistically significant difference. All statistical tests were
performed using GraphPad Prism software (version 4.00; GraphPad
Software, Inc., La Jolla, CA, USA).
Results
Phenotypic characteristics of
implanted cells
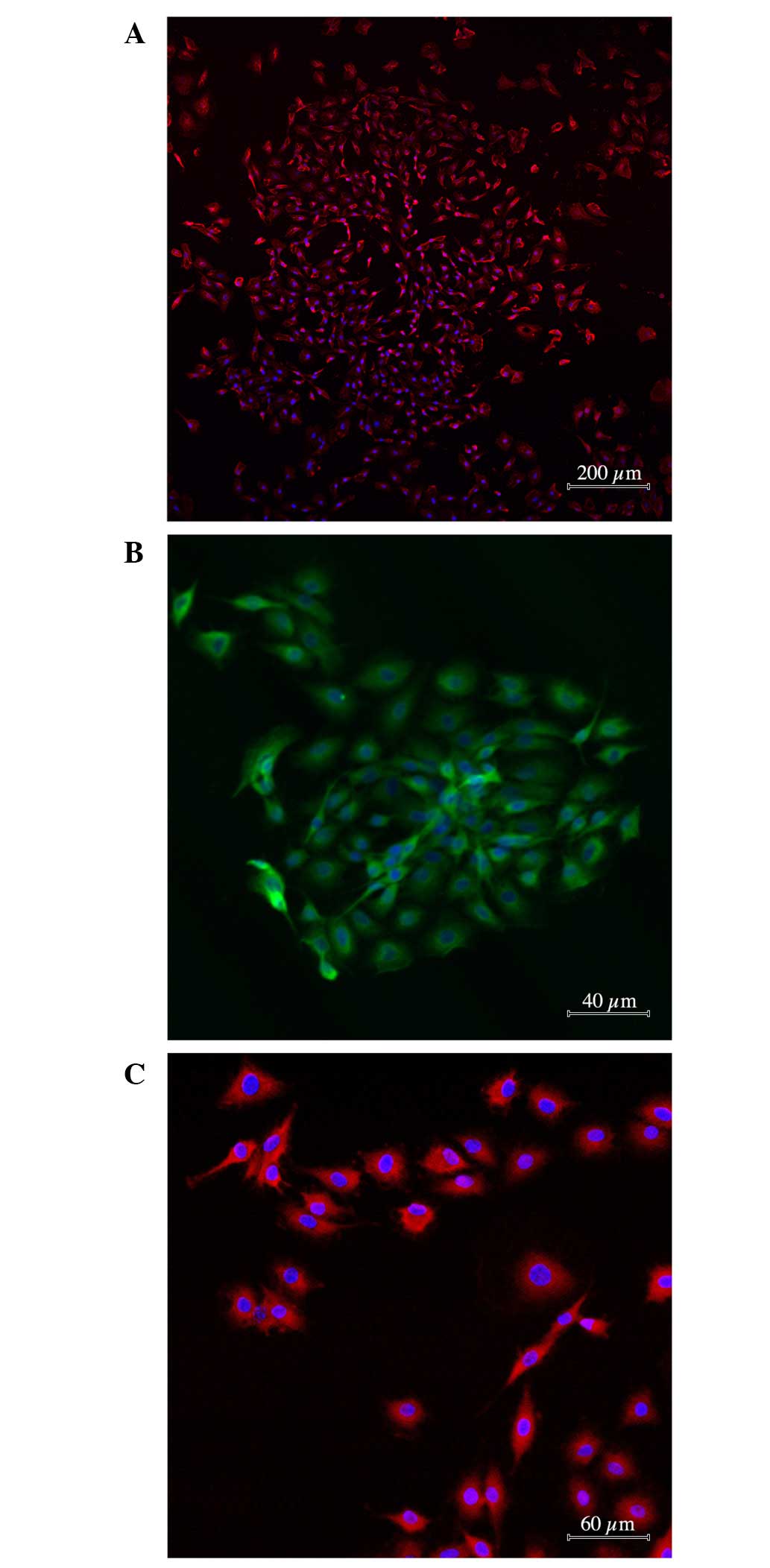
As indicated in Fig.
1, the majority of the C6 glioma specimens (96.0±6.8%) stained
positive for antibodies against nestin and mutant protein p53
(88.4±3.8%). A marked proportion of tumor cells expressed CXCR4
receptor (62.4±8.3%). However, the proportion of cells with antigen
properties of glial cells (nestin-negative; S100- and
GFAP-positive) or neural differentiation was not substantial
(1.0±0.1%; Fig. 1А-C).
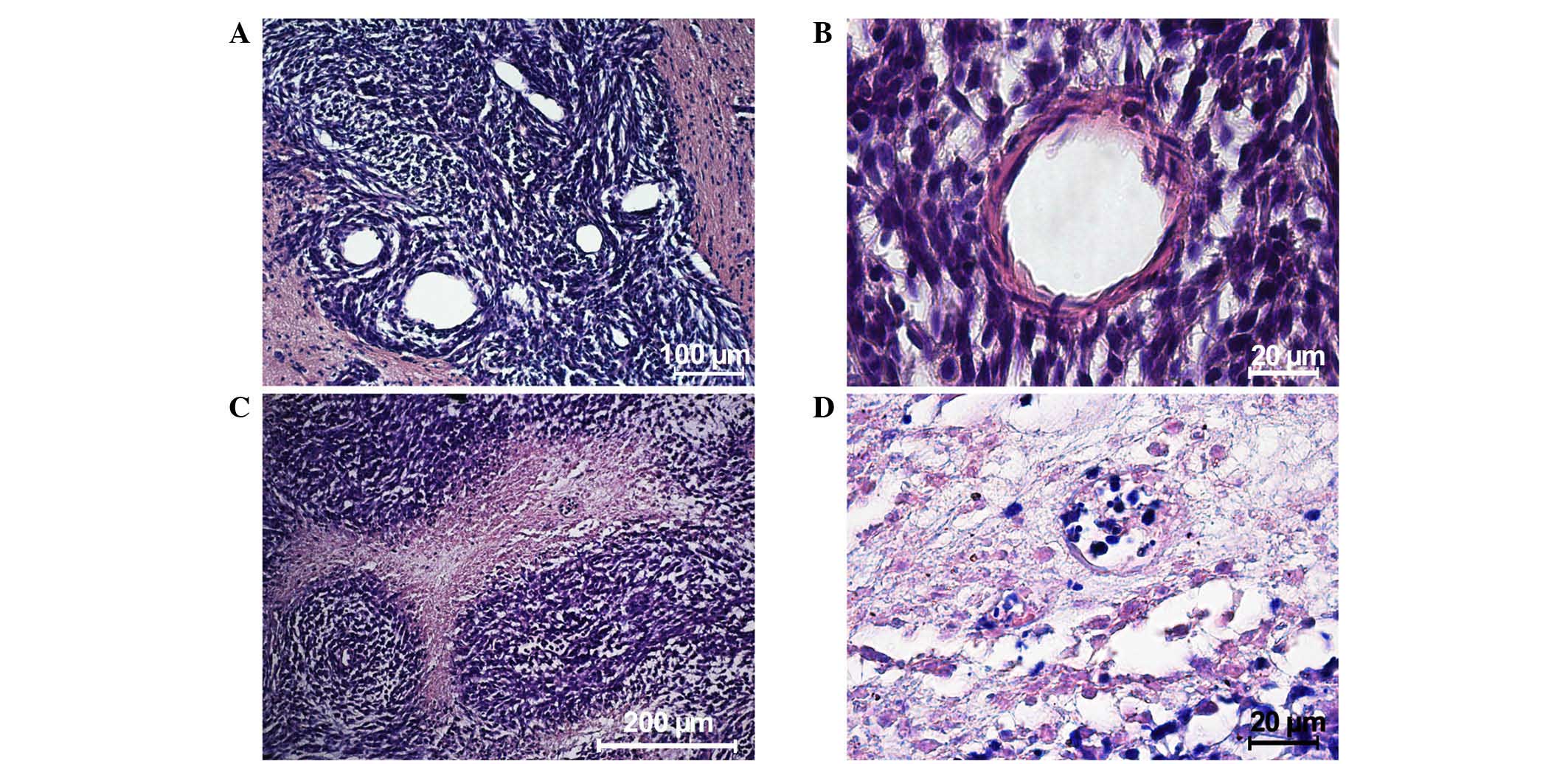
Tumor morphology
An MRI scan performed 10 days after the implantation
of tumor cells showed massive tumors (mean ± SEM, 5.6±1.4mm3) in
the brains of all the animals. Morphological analysis revealed that
the tumor was comprised of variously shaped cells with different
numbers of nuclei of different sizes. By day 20, the neoplastic
tissue contained a large number blood microvessels, indicating a
high metabolism. At the periphery of the tumor nidus, the glioma
cells invaded the brain parenchyma with dystrophic changes and the
creation of chords surrounded by a large number of cells (Fig. 2А). A little farther from the primary
nodule, the glioma cells were clustered into conglomerates creating
secondary satellite tumors. Rapid growth of a feeding blood vessel
in the center of the secondary nidus intensified neoplastic
development and invasive processes (Fig.
2B).
On days 20–30, microscopic observation identified
the start of tumor cell death. Necrosis observed in the center of
the tumor nodule close to the feeding blood vessels indicated that
the blood system was unable to satisfy the needs of the
fast-multiplying cells. By day 30, the tumor cells created thick
nodule-like clusters around the feeding blood vessel showing a
typical ‘roseola’ pattern (Fig. 2C)
with necrotic areas occupying the main space of the morphological
images (Fig. 2D). The mean life
expectancy of the experimental animals was 34.0±8.1 days.
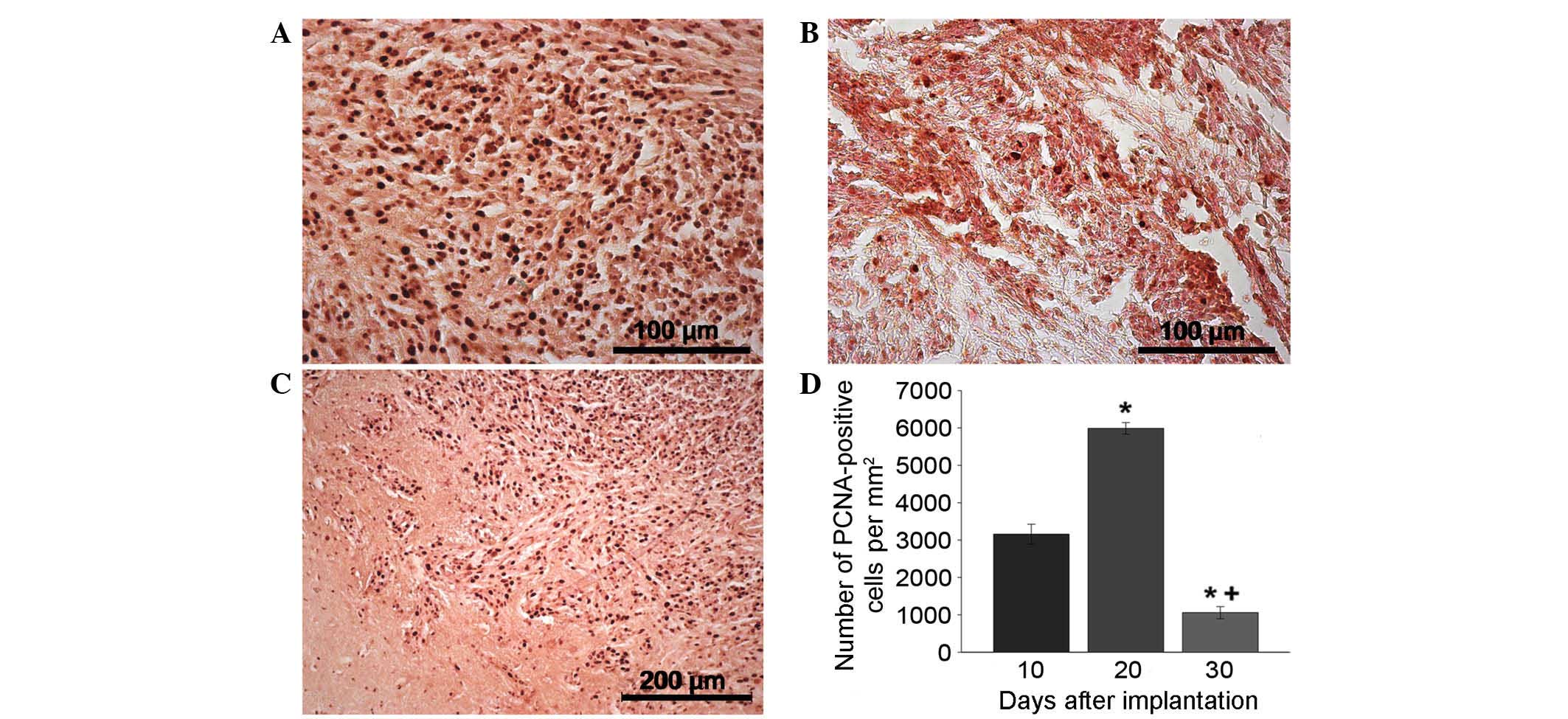
Tumor immunocytochemistry
PCNA antibody staining was used to indicate the
extent of proliferation (Fig. 3A-D).
PCNA staining revealed a sharp increase in the proliferation speed
of the tumor nodule on day 20 compared with day 10 that may be
associated with the intensification of tumor cell division due to
the development of the neoplastic blood supply network (Fig. 3A and D). Outside the tumor nidus, the
maximum proportion of PCNA-positive cells was concentrated in the
adjacent tissues, satellite nodules and blood vessels (data not
shown). By day 30, the number of proliferating cells in the tumor
tissue significantly decreased (Fig.
3В and D). Furthermore, by day 20, PCNA-positive cells had been
located along the tumor borders in areas of the brain with the
maximum infiltration of glioma cells due to their invasive growth
(Fig. 3С). Stand-alone proliferating
cells were observed in the brain tissue a significant distance from
the tumor nidus (data not shown).
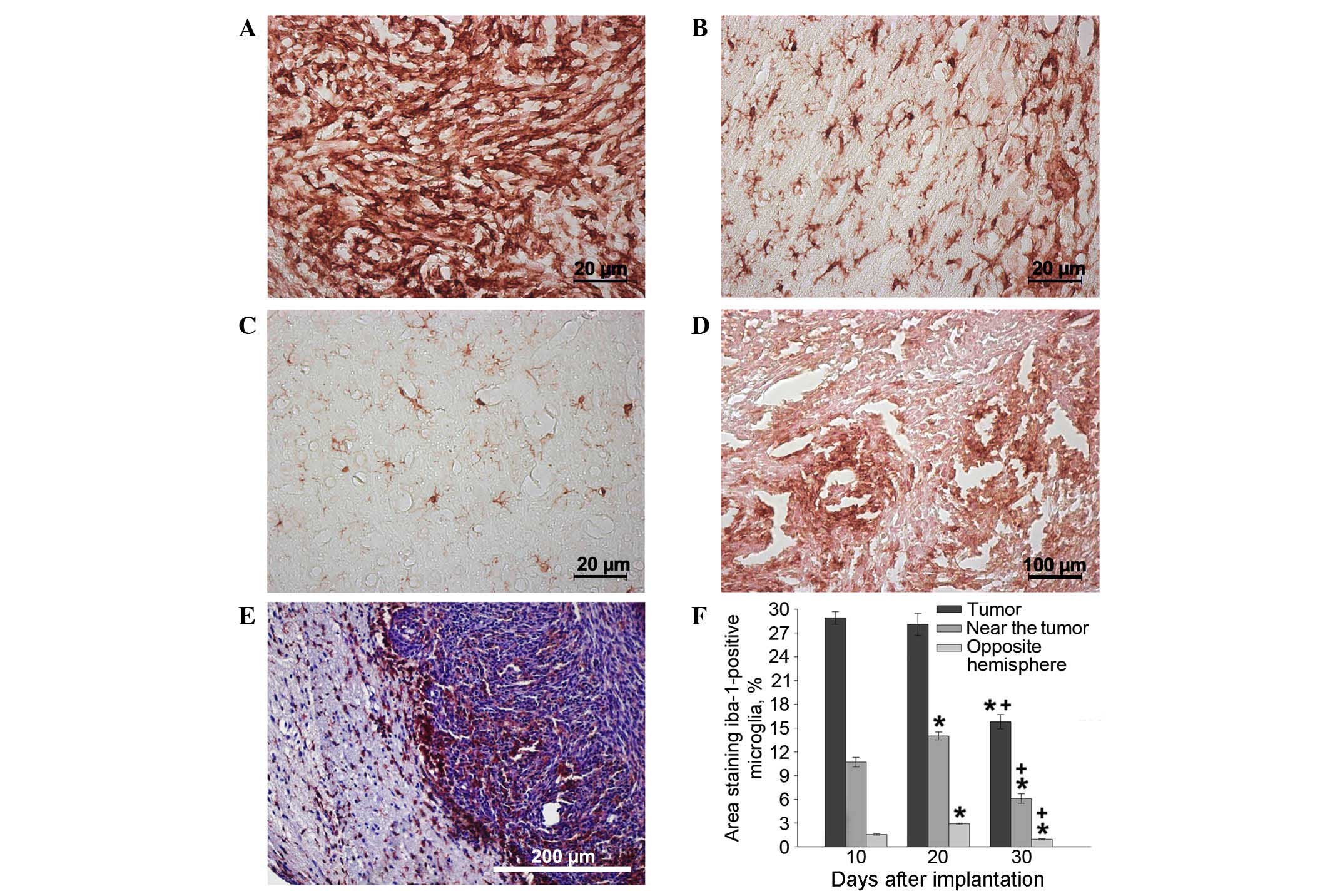
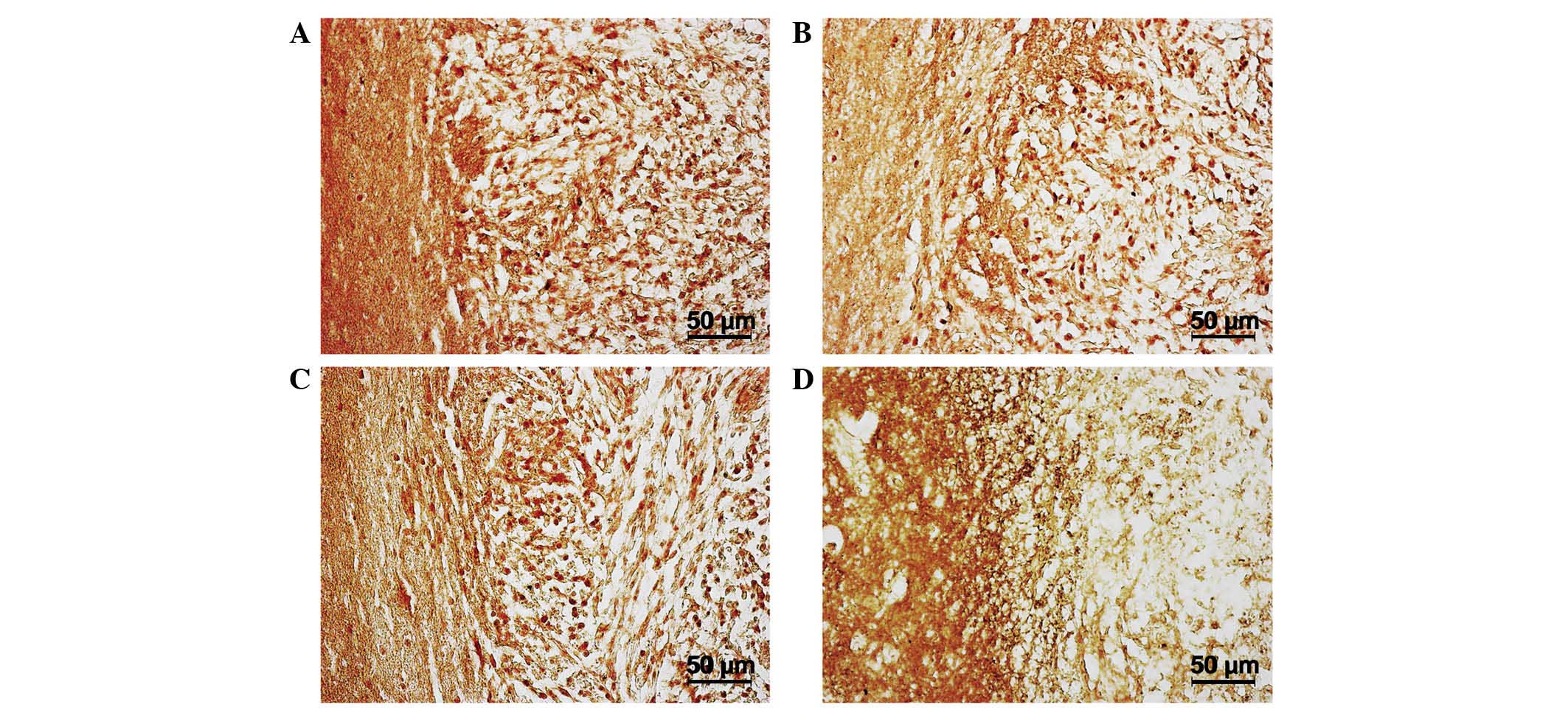
The maximum concentration of microglial cells was
observed in the tumor tissue itself and adjacent glioma invasion
areas on days 10–20, as opposed to in tissues from the opposite
hemisphere of the brain (Fig. 4A-F).
Significant clusters of small IBA-1-positive cells with an
elongated shape and short extensions were concentrated in the brain
parenchyma a small distance from the tumor nodule and in the
endothelium of hypertrophied blood vessels (Fig. 4B). The changes in microglial cell
concentration in the neoplastic nodule between days 10 and 20 may
be caused by the transformation of monocytes migrating from the
newly formed capillaries into resident macrophages, as well as by
the local migration of microglia from the adjacent brain tissues.
The number of IBA-1-positive cells in the brain parenchyma of the
hemisphere containing the tumor was significantly larger than the
number in the intact (opposite) hemisphere 20 days after
implantation (Fig. 4С and F).
After day 20, the number of microglial cells in the
tumor nodus began decreasing. Cells of round and amoeboid shape
were visible in parabasal concentrations of glioma cells, creating
thick lines around necrotic areas (Fig.
4D). In addition, by day 30, a marked number of microglial
cells were localized along the borders of the tumor nidus,
particularly in areas of glioma invasion into the brain parenchyma.
By day 30, the microglial cells were almost absent from the tumor
nodule tissue but were instead forming a rim of tumor cells
(Fig. 4E and F).
The aforementioned results are due to the complex
and specific interaction of microglia with tumor cells and CSCs,
and are associated with the secretion of certain cytokines. On day
30, the areas of perifocal invasion with the maximum concentration
of microglial cells were stained with antibodies against IL-1β,
nestin, CXCR4 and TGF-2β. The tumor and near-tumor areas were
heavily infiltrated with nestin; this indicated the presence of
poorly differentiated cells, either CSCs or neural SCs, which, as
demonstrated by the high CXCR4 expression in this location, may
have migrated here. Increased expression of IL-1β was present
within the tumor, and TGF-β expression was demonstrated in the
near-tumor areas (Fig. 5A-D).
Discussion
CSCs are the driving force of glioblastoma growth
and one of the primary causes of therapeutic resistance in cancer.
The basic properties of CSCs allow quick proliferation and the
establishment of angiogenesis, allowing fast accumulation of the
necessary resources. This notion is confirmed by the present PCNA
staining results. In addition, CSCs occupy the leading position in
the tissue hierarchy, over other somatic cells and SCs, and extend
neoplastic processes into adjacent tissues (14).
The glioma cells used in the current study are
immunopositive for nestin, one of the key markers of neural SCs.
Nestin is a selective marker of highly malignant tumors, including
breast cancer of the basal epithelium, melanoma and glioblastoma.
In addition, CXCR4 protein is one of the major markers for
different types of SCs, including CSCs (15). C6 glioma cells have a mutation in the
most important tumor suppressor in humans, the p53 gene. This
mutation means that C6 glioma cells are categorized as a mutant
cell line with immunophenotypic properties of SCs. In the present
study, single-step implantation of 0.2×106 C6 glioma
cells into a rat brain stimulated the rapid development of invasive
tumors with high proliferation rates.
High proliferation speed is a major contributing
factor to the neoplastic process of glioma cells. By day 20 of the
current study, the tumor cell proliferation rate was faster than
the angiogenesis rate (data not shown), resulting in the generation
of a hypoxic microenvironment. Massive hypoxic death of tumor cells
stimulates the production of hypoxia-inducible factor molecules,
whose major targets are angiogenesis-associated genes. Thus, by
stimulating the production of angiogenic genes, such as vascular
endothelial growth factor, hypoxia completes the circle and
contributes to the selection of the most resistant clones of tumor
cells (16).
Hypoxia has a multifunctional effect. The low
partial pressure of oxygen prevents the creation of active
radicals, significantly reducing the effects of radiation and
creating optimum conditions for CSC survival. Hypoxia is an
important factor involved in the expression of genes responsible
for the biosynthesis of cytokines, which initiate the directed
migration of somatic cells and SCs into a tumor (17).
Microglia consist of 30% of cells recruited to the
tumor environment (18,19). The main factor in this process is
stromal cell-derived factor 1α (also known as C-X-C motif chemokine
12). The ligand is released as a response to hypoxic damage of
tissues and actively attracts SCs of all types, including
monocytes, into the neoplastic area where they can be transformed
into microphages and microgliocytes (20). Another important inducer of SC and
monocyte migration to the tumor nidus is monocyte chemoattractant
protein-1; this ligand was initially identified in malignant glioma
(21).
Microglial cells form complex secretory units by
interacting with CSCs in the neoplastic nidus. For example, highly
malignant glioma cells produce colony stimulating factor-1,
attracting microglia and microphages that secrete epidermal growth
factor (EGF). EGF accelerates the rate of proliferation, and
induces the production of matrix metalloproteinase (MMP)2, 9 and 14
from tumor cells, causing the destruction of tissue barriers and
the extension of neoplastic processes into adjacent tissues
(22).
The tumor modeled in the current study was
characterized by excessive production of cytokines, as indicated by
the significant number of microglial cells concentrated in the
tumor nidus itself and the adjacent tissues. However, as the tumor
cells began to die due to hypoxia, the number of microglia and
macrophages in the tumor tissue decreased, as evidenced by the
continuous IBA-1-positive cell number reduction. Subsequently,
microglial cells were concentrated along the border of the tumor
nidus in the area of glioma cell invasion into the brain
parenchyma. It is possible that, in this case, hypoxia was an
important inductor of recruited cell traffic, but not the main
controller.
The concentration of microglia at the tumor
periphery may be explained by the influence of the specific local
microenvironment created by the CSCs. The presence of CSCs in the
neoplastic invasion area was demonstrated by local positive
staining for nestin and CXCR4 antibody. CXCR4 and nestin expression
were noted in a significant proportion of glioma cells, indicating
that these cells have CSC functions. Furthermore, CXCR4 is a
membrane component of microglial cells and monocytes that are
attracted by the tumor. Thus, there is a high probability that the
microenvironment created by CSCs selectively activates microglia
with an M2 (anti-inflammatory) phenotype, facilitating tumor growth
and invasive processes (23).
On days 20–30 of the current study, areas of
concentrated microglial cells were observed along the tumor border
and in adjacent parts of the nervous tissue by staining with
antibodies against IL-1β. This cytokine is able to induce NO
synthetase, leading to excessive NO formation that can damage
neurons and increase hypoxia (24).
Notably, TGF-β-immunopositive cells were
concentrated in the area of tumor invasion in the present study.
TGF-β is one of the main ligands responsible for the invasive
properties of glioblastoma; it triggers epithelial-mesenchymal
transition in glioma cells, causing cells to a obtain locomotor
phenotype and induce MMP synthesis. A previous study revealed that
TGF-β is able to cause metastasis and contributes to tumor
development via an autocrine mechanism. TGF-β produced by microglia
intensifies glioma cell proliferation and invasive processes
(25). In addition, the TGF-β
signaling pathway, as well as Notch and Sonic Hedgehog cascades, is
a key controllers of CSC proliferation functions (26).
Thus, after being implanted into the brain, CSCs are
able to trigger proliferation that creates a hypoxic
microenvironment, inducing directed migration of different somatic
cells and SCs that are necessary for angiogenesis and proteolysis.
A substantial proportion of cells recruited by the tumor are
microglial cells that have a strategic role in invasive growth.
TGF-β and other ligands produced by the microglia trigger
epithelial-mesenchymal transition in glioma cells, intensifying CSC
proliferative functions and creating an immunosuppressive medium
that facilitates the invasion process.
SCs migrating into neoplastic tissue are able to
have a direct anti-tumor effect. The mechanism of this effect is SC
stimulation of the Akt and Wnt signaling pathways (27,28), tumor
cell cycle arrest in the G1 phase or direct induction of apoptosis
(29). This property also means that
SCs could be utilized to deliver medication directly to the tumor
nidus or to release specific ligands that interact with major
pathways of signal transduction in CSCs, suppressing the production
of cytokines stimulating invasive growth by microglial cells.
The production of cytokines by microglial cells to
intensify invasive processes is suppressed by activating the signal
transducer and activator of transcription 3 signaling pathway
(11) and p38 mitogen-activated
protein kinase (30). Currently,
research is aimed at identifying novel substances that can activate
these signaling pathways. Using minocycline and doxycycline
suppresses tumor microglia activity by blocking MMP synthesis
(31). However, there is a more
promising method of regulating microglia and CSC function: By using
cell adhesion receptors to stimulate integrins.
Integrins have a key role in CSC remodeling of the
local microenvironment that occurs during invasive growth.
Suppressing integrin expression reduces the size of a tumor in
vivo and reduces microglial cell density at the border of a
glioma (32).
In 2014, our previous study reported that the focal
adhesion pathway in CSCs of the U87 human glioblastoma cell line is
safe and that it is possible to stimulate this pathway with
SC-based medication with a modified transcriptomic profile
(6). Microglia have a strategically
important role in promoting invasive growth, as they are
concentrated in the areas with the highest activity of CSCs that
are the primary target of biomedical cell-based treatment. We
propose that changes in microglia and subsequent tumor growth
suppression may be important criteria in evaluating the
effectiveness of cell-based treatment for glioblastoma in
vivo.
In summary, CSCs are among the leading reasons
behind therapeutic resistance in glioblastoma. Currently there are
no effective ways to eliminate CSCs within the patient's body.
Development of biomedical approaches based on autologous SCs is a
possible way to resolve this problem. Regional human SCs hold high
antitumor potential. These migrate towards cancer and accumulate in
the neoplastic tissue. We hypothesized that the most likely
direction of SC differentiation within cancer foci is into elements
of cancer microglia.
The present work describes the in vivo
dynamics of microgliacyte distribution during invasive glial tumor
growth. The findings demonstrated that during glioblastoma
development, microglia localization corresponds to nestin-positive,
CSC-rich areas of active tumor cell proliferation and invasion.
Active production of TGF-β and IL-1 within this area indicates of
the presence of complex regulatory relationships between
interacting cells which, in theory, could be controlled by
transplanting modified SCs. Changes in microglia localization and
concomitant inhibition of tumor growth can be used for assessment
of antitumor therapy effectiveness.
Acknowledgements
The present study was funded by the Ministry of
Education and Science of the Russian Federation (project no.,
14.575.21.0038; project ID, RFMEF157514X0038).
References
|
1
|
Omuro A and DeAngelis LM: Glioblastoma and
other malignant gliomas: A clinical review. JAMA. 310:1842–1850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Louis DN, Perry A, Burger P, Ellison DW,
Reifenberger G, von Deimling A, Aldape K, Brat D, Collins VP,
Eberhart C, et al: International society of neuropathology-haarlem
consensus guidelines for nervous system tumor classification and
grading. Brain Pathol. 24:429–435. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cruceru ML, Neagu M, Demoulin JB and
Constantinescu SN: Therapy targets in glioblastoma and cancer stem
cells: Lessons from hematopoietic neoplasms. J Cell Mol Med.
17:1218–1235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bryukhovetskiy I, Bryukhovetsky A,
Khotimchenko Y, Mischenko P, Tolok E and Khotimchenko R:
Combination of the multipotent mesenchymal stromal cell
transplantation with administration of temozolomide increases
survival of rats with experimental glioblastoma. Mol Med Rep.
12:2828–2834. 2015.PubMed/NCBI
|
|
6
|
Bryukhovetskiy A, Shevchenko V, Kovalev S,
Chekhonin V, Baklaushev V, Bryukhovetskiy I and Zhukova M: To the
novel paradigm of proteome-based cell therapy of tumors: Through
comparative proteome mapping of tumor stem cells and
tissue-specific stem cells of humans. Cell Transplant. 23(Suppl 1):
S151–S170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saraceni F, Shem-Tov N, Olivieri A and
Nagler A: Mobilized peripheral blood grafts include more than
hematopoietic stem cells: The immunological perspective. Bone
Marrow Transplant. 50:886–891. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Syme R, Bajwa R, Robertson L, Stewart D
and Glück S: Comparison of CD34 and monocyte-derived dendritic
cells from mobilized peripheral blood from cancer patients. Stem
Cells. 23:74–81. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moore XL, Lu J, Sun L, Zhu CJ, Tan P and
Wong MC: Endothelial progenitor cells ‘homing’ specificity to brain
tumors. Gene Ther. 11:811–818. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ye XZ, Xu SL, Xin YH, Yu SC, Ping YF, Chen
L, Xiao HL, Wang B, Yi L, Wang QL, et al: Tumor-associated
microglia/macrophages enhance the invasion of glioma stem-like
cells via TGF-β1 signaling pathway. J Immunol. 189:444–453. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
da Fonseca AC and Badie B: Microglia and
macrophages in malignant gliomas: Recent discoveries and
implications for promising therapies. Clin Dev Immunol.
2013:2641242013.PubMed/NCBI
|
|
12
|
Barth RF and Kaur B: Rat brain tumor
models in experimental neuro-oncology: The C6, 9L, T9, RG2, F98,
BT4C, RT-2 and CNS-1 gliomas. J Neurooncol. 94:299–312. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Paxinos G and Watson C: The Rat Brain in
Stereotaxic Coordinates (6th). Elsevier Academic Press. San Diego:
2007.
|
|
14
|
Lathia JD, Mack SC, Mulkearns-Hubert EE,
Valentim CL and Rich JN: Cancer stem cells in glioblastoma. Genes
Dev. 29:1203–1217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Trautmann F, Cojoc M, Kurth I, Melin N,
Bouchez LC, Dubrovska A and Peitzsch C: CXCR4 as biomarker for
radioresistant cancer stem cells. Int J Radiat Biol. 90:687–699.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao D, Najbauer J, Garcia E, Metz MZ,
Gutova M, Glackin CA, Kim SU and Aboody KS: Neural stem cell
tropism to glioma: Critical role of tumor hypoxia. Mol Cancer Res.
6:1819–1829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wierenga AT, Vellenga E and Schuringa JJ:
Convergence of hypoxia and TGFβ pathways on cell cycle regulation
in human hematopoietic stem/progenitor cells. PLoS One.
9:e934942014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Held-Feindt J, Hattermann K, Müerköster
SS, Wedderkopp H, Knerlich-Lukoschus F, Ungefroren H, Mehdorn HM
and Mentlein R: CX3CR1 promotes recruitment of human
glioma-infiltrating microglia/macrophages (GIMs). Exp Cell Res.
316:1553–1566. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Coniglio SJ, Eugenin E, Dobrenis K,
Stanley ER, West BL, Symons MH and Segall JE: Microglial
stimulation of glioblastoma invasion involves epidermal growth
factor receptor (EGFR) and colony stimulating factor 1 receptor
(CSF-1R) signaling. Mol Med. 18:519–527. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Greenbaum A, Hsu YM, Day RB, Schuettpelz
LG, Christopher MJ, Borgerding JN, Nagasawa T and Link DC: CXCL12
in early mesenchymal progenitors is required for haematopoietic
stem-cell maintenance. Nature. 495:227–230. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Desbaillets I, Tada M, de Tribolet N,
Diserens AC, Hamou MF and Van Meir EG: Human astrocytomas and
glioblastomas express monocyte chemoattractant protein-1 (MCP-1) in
vivo and in vitro. Int J Cancer. 58:240–247. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Markovic DS, Vinnakota K, Chirasani S,
Synowitz M, Raguet H, Stock K, Sliwa M, Lehmann S, Kälin R, van
Rooijen N, et al: Gliomas induce and exploit microglial MT1-MMP
expression for tumor expansion. Proc Natl Acad Sci USA.
106:12530–12535. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu A, Wei J, Kong LY, Wang Y, Priebe W,
Qiao W, Sawaya R and Heimberger AB: Glioma cancer stem cells induce
immunosuppressive macrophages/microglia. Neuro Oncol. 12:1113–1125.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu Y, Xue J, Yang Y, Zhou X, Qin C, Zheng
M, Zhu H, Liu Y, Liu W, Lou G, et al: Lipocalin 2 upregulation
protects hepatocytes from IL1-β-induced stress. Cell Physiol
Biochem. 36:753–762. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ikushima H, Todo T, Ino Y, Takahashi M,
Miyazawa K and Miyazono K: Autocrine TGF-beta signaling maintains
tumorigenicity of glioma-initiating cells through Sry-related
HMG-box factors. Cell Stem Cell. 5:504–514. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Caja L, Bellomo C and Moustakas A:
Transforming growth factor β and bone morphogenetic protein actions
in brain tumors. FEBS Lett. 589:1588–1597. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qiao L, Xu Z, Zhao T, Zhao Z, Shi M, Zhao
RC, Ye L and Zhang X: Suppression of tumorigenesis by human
mesenchymal stem cells in a hepatoma model. Cell Res. 18:500–507.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moreno E: Cancer: Darwinian tumour
suppression. Nature. 509:435–436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cousin B, Ravet E, Poglio S, De Toni F,
Bertuzzi M, Lulka H, Touil I, André M, Grolleau JL, Péron JM, et
al: Adult stromal cells derived from human adipose tissue provoke
pancreatic cancer cell death both in vitro and in vivo. PLoS One.
4:e62782009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Ren X, Shi M, Jiang Z, Wang H, Su
Q, Liu Q, Li G and Jiang G: Downregulation of STAT3 and activation
of MAPK are involved in the induction of apoptosis by HNK in
glioblastoma cell line U87. Oncol Rep. 32:2038–2046.
2014.PubMed/NCBI
|
|
31
|
Sun JS, Yang YJ, Zhang YZ, Huang W, Li ZS
and Zhang Y: Minocycline attenuates pain by inhibiting spinal
microglia activation in diabetic rats. Mol Med Rep. 12:2677–2682.
2015.PubMed/NCBI
|
|
32
|
Färber K, Synowitz M, Zahn G, Vossmeyer D,
Stragies R, van Rooijen N and Kettenmann H: An alpha5beta1 integrin
inhibitor attenuates glioma growth. Mol Cell Neurosci. 39:579–585.
2008. View Article : Google Scholar : PubMed/NCBI
|