Introduction
The pentaspan membrane glycoprotein prominin-1, also
known as cluster of differentiation (CD)133, was initially
described as a cell surface antigen specific for hematopoietic stem
cells and progenitor cells (1,2). The
biological function of CD133 remains unclear. However, it is
currently used in the identification and isolation of
tumor-initiating cells from certain malignant tumors, whereby it
correlates with poor prognosis (3–5).
Tumor-initiating cells, also called cancer stem cells (CSCs), are
characterized by their self-renewal capacity and the ability to
generate cell subpopulations during tumor growth (6–8). Although
CD133 is a reasonable marker of various CSCs, several types of
cancer arise from cells with different markers (9,10).
Pancreatic cancer is the fifth most lethal cancer in
developed countries, with a 5-year survival rate of <6%
(11). Early metastasis, late
diagnosis and the low effectiveness of the currently available
therapies contribute to its high mortality rate (12). A previous study on pancreatic cancer
identified that cells expressing CD44, CD24 and epithelial-specific
antigen surface markers were associated with an increase in the
tumorigenic and self-renewal capacity of tumor cells isolated from
primary tumors or low-passage tumor xenografts (8). However, these markers do not identify
CSCs within all pancreatic tumors, and other studies revealed that
the use of CD133 to isolate tumor-initiating cells yielded
populations of cells with enhanced tumorigenic potential, high
resistance to standard chemotherapy and a close association with
metastatic phenotype (7,13). In addition, a recent study confirmed
that enforced expression of CD133 enhanced the aggressive behavior
of pancreatic cancer cells (14).
MUC1 is a heavily glycosylated transmembrane
glycoprotein expressed at low levels in the apical surfaces of
epithelial cells (15). This
glycoprotein possesses oncogenic properties, and is overexpressed
in >80% of pancreatic tumors, contributing to tumor progression,
metastasis and mortality in patients with pancreatic cancer
(16–20). The MUC1 gene encodes a protein
comprised of a large extracellular domain with a tandem repeat
region, a transmembrane domain and a highly conserved cytoplasmic
domain (MUC1-CD), which participates in several oncogenic signaling
pathways (21). MUC1-CD is highly
conserved, and contains seven tyrosine residues and several serine
and threonine residues that represent potential docking sites for
proteins with Src homology 2 domains and recognition sites for
receptor tyrosine kinases and other kinases, including protein
kinase C delta (PKCδ), glycogen synthase kinase 3 beta (GSK3β) and
ErbB receptors such as epidermal growth factor receptor (EGFR)
(22). Furthermore, MUC1-CD contains
a serine-rich motif that functions as a β-catenin binding site, and
the phosphorylation of MUC1-CD modulates this affinity (23). MUC1-CD/β-catenin interactions enhance
the malignant phenotype of tumor cells by regulating the activity
of the T-cell factor/lymphoid enhancer factor (TCF/LEF) family of
transcription factors, thus modulating the expression of several
genes involved in the tumorigenic phenotype, including target genes
in the Wnt signaling pathway (24).
Recently, a transmembrane cleaved form of MUC1 has been reported to
exert an important role in chemoresistance to standard chemotherapy
agents (25), and to potentially
serve as an accurate marker of pluripotency in human embryonic stem
cells (26). The expression of MUC1
in CSCs has been documented by a novel antibody against
tumor-associated MUC1 that recognizes a sequence in the tandem
repeat region of MUC1, which is different from the sequences
recognized by the majority of commercially available antibodies
against MUC1 (27).
Based on the reported associations of MUC1 with
CSCs, the present study aimed to investigate the potential
contribution of MUC1 to the oncogenic signaling pathways of
CD133+ pancreatic cancer cells. The results revealed
that MUC1/β-catenin interactions are associated with enhanced
tumorigenic properties of CD133+ pancreatic cancer
cells.
Materials and methods
Cell culture
The human pancreatic cell line HPAF-II was obtained
from the American Type Culture Collection (Manassas, VA, USA), and
was cultured in RPMI 1640 medium Gibco; Thermo Fisher Scientific
Inc.,Waltham, MA USA containing GlutaMAXTMI (Gibco; Thermo Fisher
Scientific, Inc.) and 25 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (Gibco; Thermo
Fisher Scientific, Inc.), supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) and 50 mg/ml gentamicin
(Invitrogen; Thermo Fisher Scientific, Inc.). Cells were grown at
37°C with 5% CO2 in a humidified atmosphere.
CD133 cell-surface expression analysis
by flow cytometry
The expression levels of CD133 in the HPAF-II cell
line were assessed by flow cytometry with an
anti-CD133/2-phycoerythrin (PE) monoclonal antibody (MAb)
[#130-080-901; mouse immunoglobulin (IgG)1; Miltenyi Biotec GmbH,
Bergisch Gladbach, Germany]. A mouse IgG1 MAb served as a control
(#130-092-212; Miltenyi Biotec GmbH).
To perform flow cytometry analysis, cells were
trypsinized when 80% of confluence was reached. For each analysis,
5×105 cells were used. Cells were incubated with a mouse
IgG1 MAb solution (1:80) for 10 min at 4°C, and next resuspended in
an anti-CD133/2-PE antibody solution (1:10) for 10 min at 4°C in
the dark. Upon incubation, the cells were washed with 0.1% PBS two
times, and resuspended in 500 µl magnetic-activated cell sorting
(MACS) buffer [phosphate-buffered saline (PBS) supplemented with
0.5% bovine serum albumin (BSA) and 2 mM ethylenediaminetetraacetic
acid (EDTA)], prior to be analyzed in a FACSCalibur™ flow cytometer
(BD Biosciences, Franklin Lakes, NJ, USA). A cell suspension that
was only incubated with mouse IgG1 MAb was used as a control.
Analysis of the results was performed using FlowJo version 7.2.5
software (FlowJo, LLC, Ashland, OR, USA).
MACS
Cell subpopulations (CD133− and CD133+)
were isolated using a MACS system and microbeads coupled to
anti-CD133/1 MAb (Miltenyi Biotec GmbH).
Magnetic separation was performed using the
MidiMACSTM magnetic separation kit (Miltenyi Biotec GmbH),
according to the manufacturer's protocol with minor alterations.
Briefly, 1×108 cells were washed twice with 0.1% PBS and
passed through a pre-separation filter (30 µm) in order to remove
cell clumps. Subsequently, the cell suspension was incubated with
human IgG FcR Blocking Reagent (1:3 in MACS buffer; Miltenyi
Biotec, GmbH) and CD133 microbeads (1:5; Miltenyi Biotec, GmbH) for
30 min at 4°C. Following the incubation step, cells were washed
twice with 0.1% PBS, and the pellet was resuspended in 500 µl MACS
buffer (PBS supplemented with 0.5% BSA and 2 mM EDTA). The cell
suspension was then transferred to an LS column (Milteny Biotec
GmbH) previously hydrated with 3 ml buffer, and placed in a
magnetic support. The total effluent was collected as the
CD133− fraction, and the column was then washed three
times with 3 ml buffer. Next, the column was removed from the
magnet and, with the aid of a plunger, 5 ml MACS buffer were used
to flush the microbeads-labeled cells out of the column. The
effluent was collected as the CD133+ fraction.
The CD133− cell subpopulation was
subsequently passed through the LS column, and washed three times
with 1 ml MACS buffer to further deplete the remaining
CD133+ cells. Both CD133+ and
CD133− fractions were centrifuged (300 × g; Centrifuge
5810R; Eppendorf, Hamburg, Germany), and the pellets were
resuspended in culture medium and maintained at 37°C in a
humidified atmosphere containing 5% CO2.
In vivo tumorigenic assay
The present in vivo tumorigenic assay was
approved by the Institutional Animal Care and Use Committee of the
University of Nebraska Medical Center (Omaha, USA; protocol
98-088-03FC). Three groups of non-obese diabetic/severe combined
immunodeficiency (NOD/SCID) mice (n=5/group) were subcutaneously
injected in the right dorsal flanks with 3,500 HPAF-II cells
[wild-type (wt), CD133low or CD133+]. Mice were bred and maintained
under pathogen-free conditions, which included: A 12 h light/12 h
dark cycle, 6 AM/6 PM; water bag accessible at all times; Nestlets
(Animal Specialties and Provisions, LLC, Quakertown, PA, USA) or
NestPaks (WF Fisher and Son, Inc., Somerville, NJ, USA) for
enrichment; 18–23°C with 40–60% humidity; and Standard Chow food,
similar to LabDiet 5010 (protein 23%; fat content not less than
4.5%). Animals were observed twice a day by trained veterinary
staff and once a day by laboratory staff from the Eppley Institute
for Research in Cancer and Allied Disease (Omaha, NE, USA). Mice
were euthanized 4 weeks following cell injection, which was the
time point when it was necessary to euthanize the first mouse due
to the initial signs of suffering. The maximum tumor size achieved
was 263.8 mm3. Animals were sacrificed with the aid of
CO2. Following 5 min without signs of heartbeat or
respiration, the animals were subjected to cervical dislocation to
ensure mortality. Tumors were collected, fixed in 10% formalin
(Thermo Fisher Scientific Inc.) and embedded in paraffin (Thermo
Fisher Scientific Inc.) prior to sectioning (Shandon™ Finesse™ 325
microtome; Thermo Fisher Scientific Inc.). Growth of internal
tumors was evaluated by direct examination (palpation), or by
careful observation of animal behavior and estimation of
post-procedure pain, discomfort, distress or morbidity. Anesthesia,
when required, was induced by intraperitoneal administration of
ketamine hydrochloride (100 mg/ml; injectable-RL 3760;
NDC-0409-2051-05; Hospira, Inc., Lake Forest, IL, USA) and xylazine
hydrochloride (20 mg/ml; injectable-AnaSed NADA; 139–236; Lloyd,
Inc., Shenandoah, IA, USA).
Immunohistochemistry (IHC)
Tumor xenografts were paraffin-embedded and
sectioned at 4-µm thickness. IHC staining to detect CD133 protein
expression in tumor xenografts was performed using the Dako
EnVision System (Dako, Glostrup, Denmark). Antigen retrieval was
performed in an IHC-Tek™ Epitope Retrieval Steamer Set (IHC World,
LLC, Woodstock, MD, USA) for 40 min with 10 mM citrate buffer pH
6.0 (Thermo Fisher Scientific Inc.), following deparaffinization in
xylene (Thermo Fisher Scientific Inc.) and rehydration. The slides
were cooled for 20 min at room temperature, and endogenous
peroxidase was blocked with 3% H2O2 (Merck
Millipore, Darmstadt, Germany) for 5 min. Primary antibody
incubation was performed for 1 h at room temperature with a mouse
anti-human CD133/1 MAb (1:25; clone AC133; Miltenyi Biotech GmbH).
Slides were next washed in Tris-buffered saline with Tween 20
(Grisp, Porto, Portugal), and incubated with Dako REAL
EnVision-horseradish peroxidase (HRP) secondary antibody (Dako) for
30 min at room temperature. For signal detection, the slides were
incubated for 5 min with 3,3′-diaminobenzidine chromogen (Dako).
Next, tissues were counterstained with hematoxylin (Richard-Allan
Scientific™; Thermo Fisher Scientific Inc.) for 3 min, dehydrated,
cleared, mounted with Histomount medium (Richard-Allan Scientific™;
Thermo Fisher Scientific Inc.) and cover slipped. Hematoxylin and
eosin (Richard-Allan Scientific™; Thermo Fisher Scientific Inc.)
staining was performed upon antigen retrieval following a standard
protocol (28).
Protein extraction and western blot
analysis
The expression levels of MUC1-CD and oncogenic
signaling proteins were evaluated by western blotting. Unsorted
HPAF-II cells and sorted CD133low and CD133+ cell subpopulations
were cultured to 80–90% confluence. Upon washing twice with PBS,
lysis buffer [10 mM Tris pH 7.4, 150 mM NaCl, 0.1% (v/v) sodium
dodecyl sulfate (SDS; Bio-Rad Laboratories, Inc., Hercules, CA,
USA), 1 mM phenylmethylsulfonyl fluoride and 1% (v/v) Triton X-100]
was added, and cells were scraped. Cell lysates were incubated on
ice for 1 h and centrifuged (14,000 × g; Centrifuge 5417R;
Eppendorf) for 30 min at 4°C to collect the supernatants. Protein
content was assessed wit a bicinchoninic acid protein assay kit
(Bio-Rad Laboratories, Inc.), as described in the manufacturer's
protocol.
Protein extracts were analyzed by 10%
SDS-polyacrylamide gel electrophoresis (Invitrogen; Thermo Fisher
Scientific, Inc.), transferred to a polyvinylidene fluoride
membrane (GE Healthcare Life Sciences, Chalfont, UK) and incubated
overnight at 4°C with anti-MUC-1 Armenian hamster MAb (1:300;
catalogue no. Ab-5; Thermo Fisher Scientific, Inc.), anti-EGFR
mouse MAb (1:200; catalogue no. sc-81449; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), anti-PKCδ rabbit MAb (1:200; catalogue no.
sc-213; Santa Cruz Biotechnology, Inc.), anti-GSK3β mouse MAb
(1:200; catalogue no. sc-53931; Santa Cruz Biotechnology),
anti-growth factor receptor-bound protein 2 (GRB2) mouse MAb
(1:200; catalogue no. sc-8034; Santa Cruz Biotechnology, Inc.),
anti-β-catenin MAb (1:1,000; catalogue no., 610153; BD Biosciences)
and anti-β-actin MAb (1:2,000; catalogue no. sc-69879; Santa Cruz
Biotechnology, Inc.) in 5% non-fat milk diluted in PBS containing
0.1% Tween 20 (Sigma-Aldrich, St. Louis, MO, USA). Next, membranes
were washed three times with PBS containing 0.1% Tween 20, and
incubated with the corresponding goat anti-Armenian hamster
(catalogue no. sc-2443), anti-mouse (catalogue no. sc-2005) or
anti-rabbit (catalogue no. sc-2004) peroxidase conjugated antibody
(1:2,000; Santa Cruz Biotechnology, Inc.) in 5% non-fat milk
diluted in PBS containing 0.1% Tween 20. Proteins were visualized
using an enhanced chemiluminescence detection kit (Bio-Rad
Laboratories, Inc.).
Immunoprecipitation assay
The interaction between MUC1-CD and β-catenin in the
HPAF-II cell line was evaluated by immunoprecipitation. Proteins
from cell lysates (750 µg) were incubated overnight at 4°C with
protein G-agarose beads (Sigma-Aldrich) previously linked to
anti-MUC1 Ab-5 MAb and normal Armenian hamster IgG (eBioscience,
Inc., San Diego, CA, USA). Following three washes, the immune
complexes were dissociated from the beads with reducing
NuPAGE® buffer (Invitrogen; Thermo Fisher Scientific,
Inc.). The immunoprecipitates and cell lysates were separated in
12% Tris-glycine gels (Invitrogen; Thermo Fisher Scientific, Inc.),
and immunoblotted following the aforementioned procedure.
In situ proximity ligation assay
(PLA)
PLA was used to assess the close proximity (and
putative interaction) between MUC1-CD and β-catenin in tumor
xenografts. PLAs were performed using Duolink® In
Situ Detection Reagents Brightfield (Olink Bioscience, Uppsala,
Sweden), according to the manufacturer's protocol. Antigen
retrieval was performed in an IHC-Tek™ Epitope Retrieval Steamer
Set for 40 min with 10 mM citrate buffer pH 6.0, following
deparaffinization and rehydration. Subsequently, the slides were
incubated at 37°C for 30 min with a blocking solution (Olink
Bioscience) in a humidity chamber.
For β-catenin staining, the mouse primary antibody
was used under the same conditions as the ones above described for
IHC, and a secondary anti-mouse antibody conjugated with
Duolink® In Situ PLA® Probe Anti-Mouse
MINUS (Olink Bioscience) was added, followed by 1-h incubation at
37°C in a humidity chamber.
For MUC1 staining, anti-MUC1 Ab-5 primary antibody
directly conjugated with DuolinkII Probemarker Plus (Olink
Bioscience) was used. The conjugation of the antibody with the
probe was performed following the manufacturer's protocol, and
hybridization was conducted for 1 h at 37°C in a humidity chamber.
Following the ligation of the probes for 30 min at 37°C,
amplification of the signal was performed for 120 min at 37°C; both
steps occurred in a humidity chamber. To detect the signal, the
slides were incubated with HRP-labeled probes and chromogen
(catalogue no., DUO92012-30RXN; Olink Bioscience). Subsequently,
tissues were counterstained with hematoxylin, dehydrated, cleared
and mounted with Histomount medium.
Results
Isolation of a CD133+ cell
subpopulation from the HPAF-II cell line
Low-passage/highly tumorigenic samples of the
HPAF-II cell line (104 cells produced tumors in 100% of animals;
data not shown) were evaluated for CD133 expression levels by flow
cytometry. The results indicated that low-passage HPAF-II cells
contained ~4% CD133+ cells (Fig. 1A).
These cells were isolated using MACS, and both CD133+ and
CD133− subpopulations were cultured. To evaluate the
enrichment obtained with the sorting methodology used, CD133 was
again measured by flow cytometry in the above two cell
subpopulations prior to injection into immunodeficient mice
(Fig. 1B). The results revealed that
the CD133+ subpopulation was highly enriched in CD133+ cells.
However, the putative CD133− subpopulation retained a
very low percentage of cells expressing CD133, and was therefore
called CD133low. Repeated selection did not improve the performance
of this procedure (data not shown).
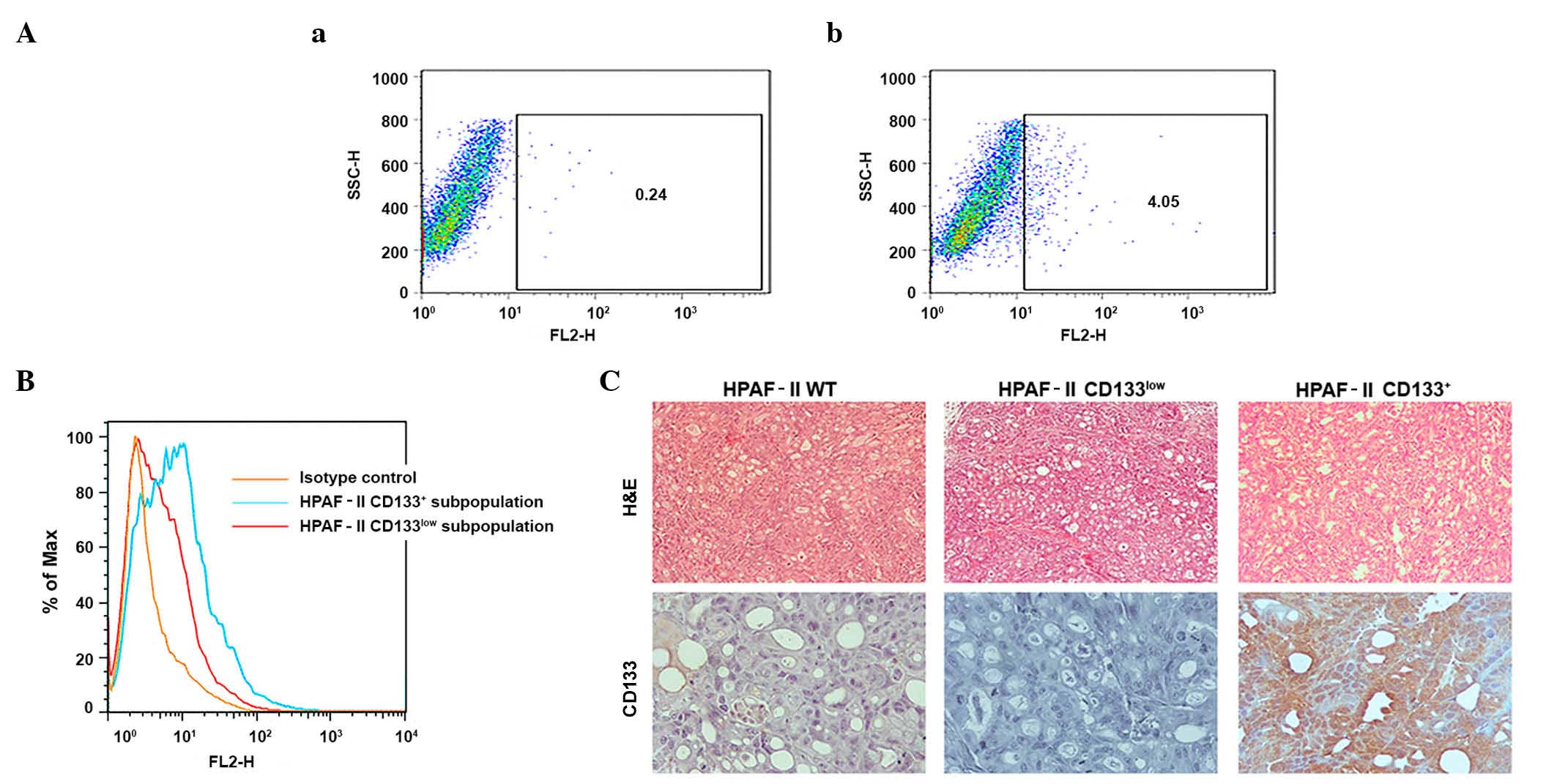 | Figure 1.Validation of the CSC model. (A)
Identification of a CSC subpopulation (CD133+ cells) in
the HPAF-II pancreatic cancer cell line and evaluation of CD133
expression in cell subpopulations sorted by flow cytometry. (a)
Isotype stained cells were used as controls. (b) HPAF-II cells
stained with CD133/2-phycoerythrin monoclonal antibody. (B)
Enrichment of HPAF-II CD133+ subpopulation isolated by
magnetic-activated cell sorting represented on a frequency
distribution histogram. The HPAF-II CD133+ subpopulation
exhibits 8.89% of CD133+ cells, while the HPAF-II
CD133low subpopulation exhibited 3.07% of
CD133+ cells, representing an enriched and a depleted
population, respectively. (C) CD133 expression in HPAF-II tumor
xenografts determined by immunohistochemistry (magnification,
×400). Hemotoxylin and eosin staining was used to reveal the
morphology of the tumors (magnification, ×100). CSC, cancer stem
cell; CD, cluster of differentiation; H&E, hematoxylin and
eosin; SSC-H, measures cell granularity or internal complexity;
FH2, phycoerythrin detection; % of Max, % of maximum (normalization
method). |
The sorted cells were evaluated for tumorigenicity
and tumor phenotype. The CD133+ enriched subpopulation
exhibited increased tumorigenic potential when injected
subcutaneously into NOD/SCID mice, since higher number of tumors
were obtained from these cells (Table
I), and tumor growth was initiated at earlier time points (3
weeks), compared with the CD133low population (Table I). IHC analysis demonstrated that the
xenografts recapitulated the HPAF-II CD133 subpopulation expression
levels. Tumors derived from the HPAF-II CD133+
subpopulation retained high CD133 expression levels, with limited
negative cells, while the HPAF-II CD133low xenografts
were largely negative for expression of CD133. The parental HPAF-II
wt-derived xenografts displayed a small percentage of
CD133+ cells, similar to the original cell line
(Fig. 1C).
 | Table I.In vivo tumorigenic assay. |
Table I.
In vivo tumorigenic assay.
|
| Time
(weeks)a |
|---|
|
|
|
|---|
| Cell
subpopulation | 1 | 2 | 3 | 4 |
|---|
| HPAF-II wt | 0/5 | 0/5 | 0/5 | 1/5 |
| HPAF-II
CD133low | 0/5 | 0/5 | 0/5 | 1/5 |
| HPAF-II
CD133+ | 0/5 | 0/5 | 2/5 | 4/5 |
Expression of MUC1 in the
CD133+ cell subpopulation
In order to evaluate the relevance of MUC1
glycoprotein in CD133+ cell biology, the expression levels of MUC1
were analyzed in CD133low and CD133+ subpopulations by
immunoblotting (Fig. 2A). HPAF-II
CD133+ cells were highly enriched in MUC1 expression, compared with
HPAF-II wt and HPAF-II CD133low cells. In addition, MUC1 expression
levels in the HPAF-II CD133low cell subpopulation were lower than
in HPAF-II wt cells (Fig. 2A).
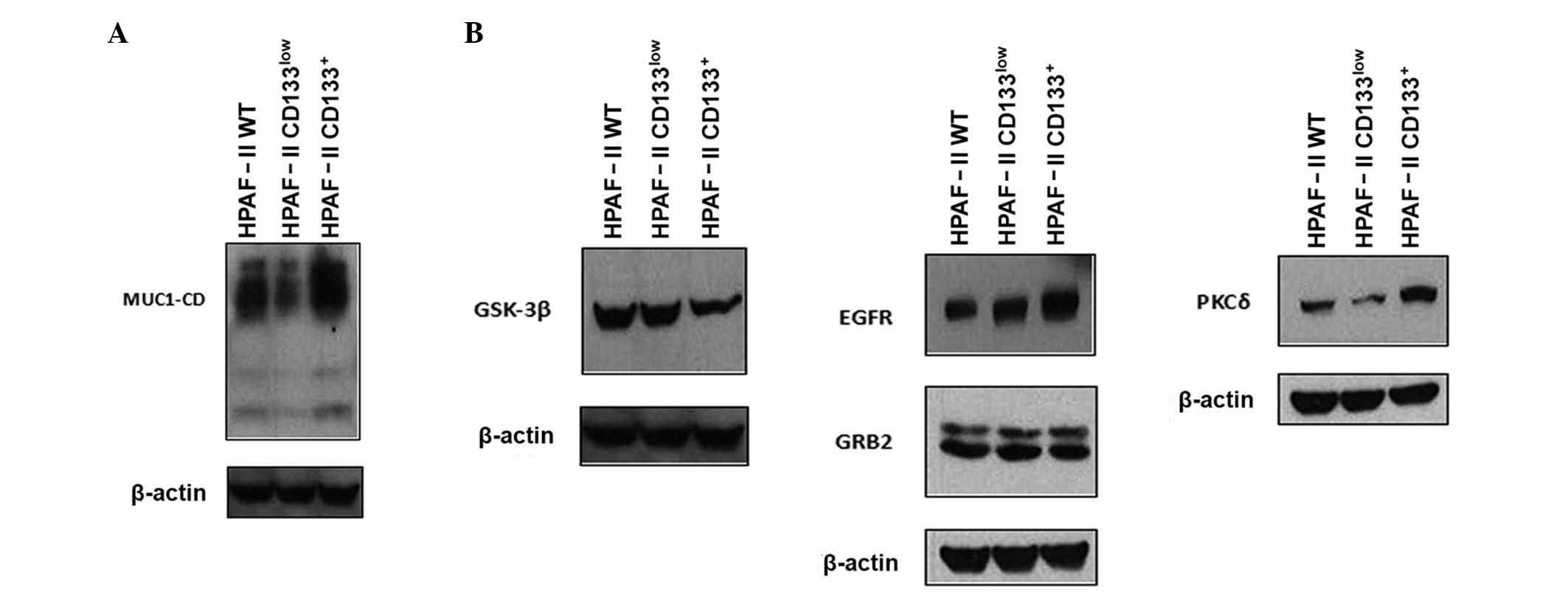 | Figure 2.Expression of MUC1 and signaling
partners. (A) Expression of MUC1 in HPAF-II wt, HPAF-II
CD133low and HPAF-II CD133+ cells was
evaluated by western blotting. β-actin was used as a loading
control. (B) Expression of MUC1 signaling partners (epidermal
growth factor receptor, growth factor receptor-bound protein 2,
protein kinase C delta and glycogen synthase kinase 3 beta) in
HPAF-II wt, HPAF-II CD133low and HPAF-II
CD133+ cells was evaluated by western blotting. β-actin
was used as a loading control. MUC1, mucin 1; MUC1-CD, mucin 1
cytoplasmic domain; EGFR, epidermal growth factor receptor; PKCδ,
protein kinase C delta; GSK3β, glycogen synthase kinase 3 beta;
GRB2, growth factor receptor-bound protein 2; CD, cluster of
differentiation; wt, wild-type. |
Expression of MUC1 signaling partners
in CD133+ cells
MUC1 function in oncogenic pathways depends on the
phosphorylation of its CD by several kinases, including EGFR, PKCδ
and GSK3β (21). Since it is not
possible to assess the phosphorylation status of MUC1-CD due to the
unavailability of antibodies sensitive to phosphorylation, the
expression of selected kinases in CD133+ cells was evaluated by
immunoblot analysis. The results indicated that CD133+ cells were
enriched in EGFR and PKCδ expression, whereas CD133low cells were
enriched in GSK3β expression. The protein expression levels of GRB2
were equivalent in HPAF-II wt, HPAF-II CD133+ and HPAF-II CD133low
cells (Fig. 2B).
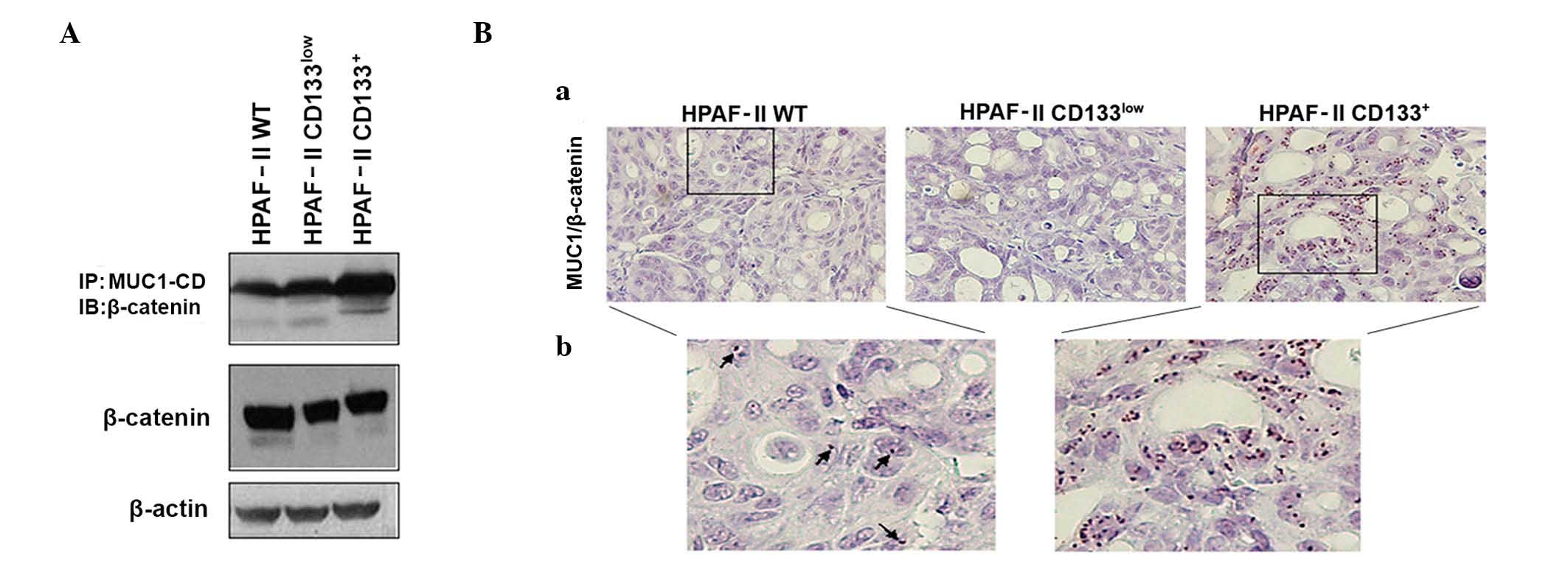
MUC1 and β-catenin interaction in the
HPAF-II cell line and tumor xenografts
MUC1-CD contains docking sites for β-catenin, and
the interactions between MUC1 and β-catenin are known to contribute
to the malignant phenotype of tumor cells by modifying the
expression of target genes in the Wnt signaling pathway (24,29). To
assess if MUC1/β-catenin interaction was potentiated in CD133+
cells, MUC1 was immunoprecipitated from cell lysates of HPAF-II wt,
HPAF-II CD133+ and HPAF-II CD133low cells, followed by β-catenin
immunoblotting. An enrichment in MUC1/β-catenin interaction was
observed in HPAF-II CD133+ cells. β-catenin expression levels were
similar in all conditions (Fig.
3A).
MUC1/β-catenin interactions in tumor xenografts were
confirmed by in situ PLA. Significant interactions between
MUC1 and β-catenin were observed in HPAF-II wt and HPAF-II
CD133+-derived xenografts, but almost no interactions
were observed in HPAF-II CD133low-derived tumors. The
most abundant interactions were observed in the CD133+
tumors (Fig. 3B). In all cases, the
interaction signals were predominantly observed in the nuclei of
the cells.
Discussion
In the present study, the involvement of CD133 and
MUC1 in the highly tumorigenic low-passage pancreatic cancer cell
line HPAF-II, which was derived from the ascites of a pancreatic
cancer patient, was investigated (30). Using a well-established CD133
selection method, an isolated CD133+ cell subpopulation
was demonstrated to exhibit features associated with CSCs (enhanced
tumorigenicity) and concomitant enriched expression of MUC1.
CSCs are known to aberrantly activate canonical
signaling pathways (31–33). Recently, a MUC1 spliced form was
reported to be associated with the differentiation status of stem
cells (34). In the present study,
the expression of MUC1 and oncogenic signaling transducers (EGFR,
PKCδ, GSK3β and GRB2), as well as the MUC1/β-catenin association,
were characterized in pancreatic cancer cells that expressed CD133.
MUC1-CD, EGFR and PKCδ expression levels were increased in the
HPAF-II CD133+ subpopulation, while GSK3β expression was
decreased, and no significant differences were observed regarding
GRB2 and β-catenin expression levels. These results clearly
demonstrate that pancreatic HPAF-II CD133+ cells have a
distinct expression profile, which includes MUC1 and its associated
signaling partners, compared with the subpopulation of cells that
do not express the stem cell surface marker CD133.
MUC1-CD contains docking sites for molecules such as
β-catenin, and the association of these proteins is modulated by
motifs that may be phosphorylated by several kinases, namely EGFR,
PKCδ, GSK3β and GRB2 (35). The
phosphorylation of MUC1-CD influences its interaction with
β-catenin, which directly binds at the MUC1-CD motif SAGNGGSSLS
(22,23). In the present study, increased
interactions between MUC1-CD and β-catenin were observed in the
HPAF-II CD133+ subpopulation, which was correlated with
enhanced expression of EGFR and PKCδ, and decreased expression of
GSK3β (24,36–42). It is
known that MUC1-CD phosphorylation by EGFR and PKCδ promotes
interactions between β-catenin and MUC1, while phosphorylation by
GSK3β leads to a decrease in this association (39–41). It
was observed in the present study that EGFR and PKCδ expression
were upregulated, while GSK3β expression was downregulated, in the
HPAF-II CD133+ subpopulation, compared with the HPAF-II
CD133low subpopulation, which likely explains the
observed increase in MUC1-CD/β-catenin interactions in the
CD133+ subpopulation, despite the fact that the
steady-state levels of β-catenin remained unchanged in these
cells.
The interactions between MUC1-CD/β-catenin influence
several tumorigenic processes. Binding of MUC1-CD to β-catenin
suppresses the capacity of β-catenin to interact with E-cadherin at
adherens junctions, resulting in the loss of cell-cell adhesion,
thus playing a relevant role in tumor invasion (43). The MUC1-CD/β-catenin complex
contributes to β-catenin stabilization by blocking its
GSK3β-mediated phosphorylation and consequently its degradation in
the proteasome (24). Furthermore,
MUC1-CD/β-catenin is translocated to the nucleus, where it may
enhance the activity of β-catenin in association with TCF/LEF
transcription factors, thus promoting cell proliferation and
survival through upregulation of the transcription of Wnt target
genes (36–38,42).
Recently, this process has been associated with a metastatic gene
expression signature and an epithelial-to-mesenchymal transition
phenotype of tumor cells (44). In
the present study, the results of in situ PLA for tumor
xenografts revealed that CD133+ tumors exhibited
frequent MUC1-CD/β-catenin interactions, with the MUC1-CD/β-catenin
complex being mainly present in the cellular nuclei, where it
presumably binds to transcription factors and activates the
transcription of genes involved in cell proliferation and
survival.
In summary, the present study has demonstrated for
the first time that pancreatic CD133+ cells display
enhanced expression of MUC1 and its associated signaling partners.
CD133 and MUC1 expression are associated with an aggressive tumor
phenotype, partly through the production of enhanced
MUC1-CD/β-catenin interactions, and this may partly explain the
behavior of pancreatic CSCs.
References
|
1
|
Yin AH, Miraglia S, Zanjani ED,
AlmeidaPorada G, Ogawa M, Leary AG, Olweus J, Kearney J and Buck
DW: AC133, a novel marker for human hematopoietic stem and
progenitor cells. Blood. 90:5002–5012. 1997.PubMed/NCBI
|
|
2
|
Miraglia S, Godfrey W, Yin AH, Atkins K,
Warnke R, Holden JT, Bray RA, Waller EK and Buck DW: A novel
five-transmembrane hematopoietic stem cell antigen: Isolation,
characterization, and molecular cloning. Blood. 90:5013–5021.
1997.PubMed/NCBI
|
|
3
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
4
|
Todaro M, Francipane MG, Medema JP and
Stassi G: Colon cancer stem cells: Promise of targeted therapy.
Gastroenterology. 138:2151–2162. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen S, Song X, Chen Z, Li X, Li M, Liu H
and Li J: CD133 expression and the prognosis of colorectal cancer:
A systematic review and meta-analysis. PLoS One. 8:e563802013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dalerba P, Dylla SJ, Park IK, Liu R, Wang
X, Cho RW, Hoey T, Gurney A, Huang EH, Simeone DM, et al:
Phenotypic characterization of human colorectal cancer stem cells.
Proc Natl Acad Sci USA. 104:10158–10163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hermann PC, Huber SL, Herrler T, Aicher A,
Ellwart JW, Guba M, Bruns CJ and Heeschen C: Distinct populations
of cancer stem cells determine tumor growth and metastatic activity
in human pancreatic cancer. Cell Stem Cell. 1:313–323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li C, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Florek M, Haase M, Marzesco AM, Freund D,
Ehninger G, Huttner WB and Corbeil D: Prominin-1/CD133, a neural
and hematopoietic stem cell marker, is expressed in adult human
differentiated cells and certain types of kidney cancer. Cell
Tissue Res. 319:15–26. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shmelkov SV, Butler JM, Hooper AT, Hormigo
A, Kushner J, Milde T, St Clair R, Baljevic M, White I, Jin DK, et
al: CD133 expression is not restricted to stem cells, and both
CD133+ and CD133, metastatic colon cancer cells initiate
tumors. J Clin Invest. 118:2111–2120. 2008.PubMed/NCBI
|
|
11
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Collins A and Bloomston M: Diagnosis and
management of pancreatic cancer. Minerva Gastroenterol Dietol.
55:445–454. 2009.PubMed/NCBI
|
|
13
|
Moriyama T, Ohuchida K, Mizumoto K, Cui L,
Ikenaga N, Sato N and Tanaka M: Enhanced cell migration and
invasion of CD133+ pancreatic cancer cells cocultured
with pancreatic stromal cells. Cancer. 116:3357–3368. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nomura A, Banerjee S, Chugh R, Dudeja V,
Yamamoto M, Vickers SM and Saluja AK: CD133 initiates tumors,
induces epithelial-mesenchymal transition and increases metastasis
in pancreatic cancer. Oncotarget. 6:8313–8322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gendler SJ: MUC1, the renaissance
molecule. J Mammary Gland Biol Neoplasia. 6:339–353. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Costa NR, Paulo P, Caffrey T,
Hollingsworth MA and Santos-Silva F: Impact of MUC1 mucin
downregulation in the phenotypic characteristics of MKN45 gastric
carcinoma cell line. PLoS One. 6:e269702011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jonckheere N and Van Seuningen I: The
membrane-bound mucins: From cell signalling to transcriptional
regulation and expression in epithelial cancers. Biochimie.
92:1–11. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Roy LD, Sahraei M, Subramani DB, Besmer D,
Nath S, Tinder TL, Bajaj E, Shanmugam K, Lee YY, Hwang SI, et al:
MUC1 enhances invasiveness of pancreatic cancer cells by inducing
epithelial to mesenchymal transition. Oncogene. 30:1449–1459. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carraway KL III, Funes M, Workman HC and
Sweeney C: Contribution of membrane mucins to tumor progression
through modulation of cellular growth signaling pathways. Curr Top
Dev Biol. 78:1–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bafna S, Kaur S and Batra SK:
Membrane-bound mucins: The mechanistic basis for alterations in the
growth and survival of cancer cells. Oncogene. 29:2893–2904. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Singh AP, Chauhan SC, Bafna S, Johansson
SL, Smith LM, Moniaux N, Lin MF and Batra SK: Aberrant expression
of transmembrane mucins, MUC1 and MUC4, in human prostate
carcinomas. Prostate. 66:421–429. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carson DD: The cytoplasmic tail of MUC1: A
very busy place. Sci Signal. 1:pe352008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamamoto M, Bharti A, Li Y and Kufe D:
Interaction of the DF3/MUC1 breast carcinoma-associated antigen and
beta-catenin in cell adhesion. J Biol Chem. 272:12492–12494. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang L, Chen D, Liu D, Yin L, Kharbanda S
and Kufe D: MUC1 oncoprotein blocks glycogen synthase kinase
3beta-mediated phosphorylation and degradation of beta-catenin.
Cancer Res. 65:10413–10422. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fessler SP, Wotkowicz MT, Mahanta SK and
Bamdad C: MUC1* is a determinant of trastuzumab (Herceptin)
resistance in breast cancer cells. Breast Cancer Res Treat.
118:113–124. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hikita ST, Kosik KS, Clegg DO and Bamdad
C: MUC1* mediates the growth of human pluripotent stem cells. PLoS
One. 3:e33122008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Curry JM, Thompson KJ, Rao SG, Besmer DM,
Murphy AM, Grdzelishvili VZ, Ahrens WA, McKillop IH, Sindram D,
Iannitti DA, et al: The use of a novel MUC1 antibody to identify
cancer stem cells and circulating MUC1 in mice and patients with
pancreatic cancer. J Surg Oncol. 107:713–722. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Suvarna SK, Layton C and Bancroft JD: The
hematoxylins and eosinBancroft's Theory and Practice of
Histological Techniques. 7th. Churchill Livingstone; London: pp.
178–179. 2013
|
|
29
|
Moon RT, Kohn AD, De Ferrari GV and Kaykas
A: WNT and beta-catenin signalling: Diseases and therapies. Nat Rev
Genet. 5:691–701. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Metzgar RS, Gaillard MT, Levine SJ, Tuck
FL, Bossen EH and Borowitz MJ: Antigens of human pancreatic
adenocarcinoma cells defined by murine monoclonal antibodies.
Cancer Res. 42:601–608. 1982.PubMed/NCBI
|
|
31
|
Vathipadiekal V, Saxena D, Mok SC,
Hauschka PV, Ozbun L and Birrer MJ: Identification of a potential
ovarian cancer stem cell gene expression profile from advanced
stage papillary serous ovarian cancer. PLoS One. 7:e290792012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoon CH, Kim MJ, Kim RK, Lim EJ, Choi KS,
An S, Hwang SG, Kang SG, Suh Y, Park MJ and Lee SJ: c-Jun
N-terminal kinase has a pivotal role in the maintenance of
self-renewal and tumorigenicity in glioma stem-like cells.
Oncogene. 31:4655–4666. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rohner A, Spilker ME, Lam JL, Pascual B,
Bartkowski D, Li QJ, Yang AH, Stevens G, Xu M, Wells PA, et al:
Effective targeting of Hedgehog signaling in a medulloblastoma
model with PF-5274857, a potent and selective Smoothened antagonist
that penetrates the blood-brain barrier. Mol Cancer Ther. 11:57–65.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Horn G, Gaziel A, Wreschner DH,
Smorodinsky NI and Ehrlich M: ERK and PI3K regulate different
aspects of the epithelial to mesenchymal transition of mammary
tumor cells induced by truncated MUC1. Exp Cell Res. 315:1490–1504.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vos HL, de Vries Y and Hilkens J: The
mouse episialin (Muc1) gene and its promoter: Rapid evolution of
the repetitive domain in the protein. Biochem Biophys Res Commun.
181:121–130. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baldus SE, Mönig SP, Huxel S, Landsberg S,
Hanisch FG, Engelmann K, Schneider PM, Thiele J, Hölscher AH and
Dienes HP: MUC1 and nuclear beta-catenin are coexpressed at the
invasion front of colorectal carcinomas and are both correlated
with tumor prognosis. Clin Cancer Res. 10:2790–2796. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Behrens J, von Kries JP, Kühl M, Bruhn L,
Wedlich D, Grosschedl R and Birchmeier W: Functional interaction of
beta-catenin with the transcription factor LEF-1. Nature.
382:638–642. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang L, Ren J, Chen D, Li Y, Kharbanda S
and Kufe D: MUC1 cytoplasmic domain coactivates Wnt target gene
transcription and confers transformation. Cancer Biol Ther.
2:702–706. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li Y and Kufe D: The human DF3/MUC1
carcinoma-associated antigen signals nuclear localization of the
catenin p120(ctn). Biochem Biophys Res Commun. 281:440–443. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ren J, Li Y and Kufe D: Protein kinase C
delta regulates function of the DF3/MUC1 carcinoma antigen in
beta-catenin signaling. J Biol Chem. 277:17616–17622. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schroeder JA, Thompson MC, Gardner MM and
Gendler SJ: Transgenic MUC1 interacts with epidermal growth factor
receptor and correlates with mitogen-activated protein kinase
activation in the mouse mammary gland. J Biol Chem.
276:13057–13064. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wen Y, Caffrey TC, Wheelock MJ, Johnson KR
and Hollingsworth MA: Nuclear association of the cytoplasmic tail
of MUC1 and beta-catenin. J Biol Chem. 278:38029–38039. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yuan Z, Wong S, Borrelli A and Chung MA:
Down-regulation of MUC1 in cancer cells inhibits cell migration by
promoting E-cadherin/catenin complex formation. Biochem Biophys Res
Commun. 362:740–746. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gnemmi V, Bouillez A, Gaudelot K, Hémon B,
Ringot B, Pottier N, Glowacki F, Villers A, Vindrieux D, Cauffiez
C, et al: MUC1 drives epithelial-mesenchymal transition in renal
carcinoma through Wnt/β-catenin pathway and interaction with SNAIL
promoter. Cancer Lett. 346:225–236. 2014. View Article : Google Scholar : PubMed/NCBI
|