Introduction
Macrophage migration inhibitory factor (MIF) is a
pro-inflammatory cytokine that was first identified in 1966 in a
study by Bloom and Bennett (1), which
reported that T lymphocytes released a factor able to inhibit the
random movement of macrophages. The MIF gene is localized on
chromosome 22q11.2 and codes for a transcript 800 bp in length. The
MIF protein is composed of 115 amino acids with a molecular weight
of 12.5 kDa in the monomeric form. The active form of MIF is a
homotrimer: Each monomer exhibits two anti-parallel α-helices and
six β-strands (2). This cytokine
shares homology with the bacterial enzyme 4-oxalocrotonate
tautomerase (3). Additionally, in a
study on melanin biosynthesis, MIF catalyzed the conversion of
D-dopachrome to 5,6-dihydroxyindole-2-carboxylic acid and
this was dependent on its tautomerase activity (4).
In addition to T lymphocytes, MIF is secreted by a
variety of other cells, including epithelial cells, endothelial
cells and macrophages (5). MIF also
exhibits the properties of a stress hormone, as it is expressed at
high concentrations in the anterior pituitary gland, from which its
release is triggered by corticotrophin-releasing hormone (6). During inflammatory responses, MIF
counterbalances the immunosuppressive activity of glucocorticoids.
This effect may be explained by the fact that MIF inhibits
mitogen-activated protein kinase (MAPK) phosphatase-1, which is
induced by glucocorticoids (7). In
addition, MIF has been implicated in several types of inflammatory
diseases, including atherosclerosis, rheumatoid arthritis, systemic
lupus erythematosus, inflammatory bowel disease, psoriasis and
diabetes (8,9). Therefore, at present, studies are
focusing on the identification and development of pharmacological
agents capable of interfering with MIF activity.
MIF and cancer
In addition to inflammatory diseases, MIF has also
been demonstrated to be overexpressed in solid tumors, such as
lung, colorectal, breast, cervical, prostate, and head and neck
cancer, where it may exhibit a crucial function in tumor
progression (cell proliferation and invasiveness) and tumor-induced
angiogenesis (10–15). It is hypothesized that the latter
processes are modulated by MIF binding to its cognate receptor,
cluster of differentiation (CD)74 [also known as the invariant
chain of the major histocompatibility complex class II] in
association with CD44. While CD74 provides the binding site,
downstream signal transduction pathways [MAPK and AKT pathways] are
activated via CD44.
Globally, the MIF effects in cancer may be mainly
explained by signaling through the CD74 receptor, since we recently
showed that i) CD74 is upregulated in oral cavity carcinomas
compared with benign lesions, ii) knockdown of CD74 in the murine
squamous cell carcinoma SCCVII cell line decreases in vitro
proliferation, migration, MMP9 secretion and VEGF production, and
iii) SCCVII CD74-knockdown cells orthotopically inoculated in mice
have a weaker growth capacity than scramble cells (16).
However, additional receptors may be involved in the
effects of MIF in cancer insofar as its interaction with the
chemokine receptor CXCR4 may induce metastasis. Indeed, Dessein
et al (17) showed that MIF
binding to CXCR4 was associated with invasion and metastasis in
human colon carcinoma cells.
MIF and carcinogenesis
Carcinogenesis refers to the processes by which
normal cells are transformed into cancer cells. Several clinical
studies have revealed that MIF expression is increased in cancer
tissues compared with corresponding normal tissues. For example, a
previous study demonstrated that in gastric cancer, positive MIF
expression rates were 12, 52 and 96% in normal mucosal, gastritis
and gastric cancer tissues, respectively (18). Similar observations have also been
reported in pancreatic cancer, melanoma, hepatocellular carcinoma,
malignant glioma and cervical adenocarcinoma (13,19–22).
Furthermore, Zhao et al (23) demonstrated that serum MIF levels may
aid to differentiate cancer patients with hepatocellular carcinoma
from individuals with other liver diseases, such as cirrhosis, when
using a reference threshold of 35.3 ng/ml. Similarly, De Souza
et al (24) reported that in
oral squamous cell carcinoma patients, MIF serum levels decreased
following tumor resection and thus, serum MIF was proposed as a
biomarker.
In our previous studies, a significant increase in
MIF immunostaining was observed in hypopharyngeal carcinoma, oral
cavity carcinoma and laryngeal carcinoma when compared with normal
epithelium, and low and high-grade dysplasia and carcinoma,
respectively (15,25,26). In
addition, our previous study also revealed that in breast cancer
patients, MIF expression was increased in cancer tissues when
compared with tumor-free breast tissues in glandular and stromal
compartments (12). Therefore, these
results provide compelling evidence that MIF is involved in tumor
biology.
MIF and disease prognosis
Kamimura et al (27) reported that low nuclear MIF expression
was correlated with a worse prognosis in lung adenocarcinoma and
thus, it was postulated that the intracellular distribution of MIF
has prognostic significance. By contrast, subsequent studies
indicated that high MIF expression in cancer was correlated with
poor patient survival. For example, Tomiyasu et al (28) demonstrated that high MIF expression in
lung cancer tissues was correlated with heavy smoking status and a
poorer prognosis. Furthermore, overexpression of MIF correlates
with a worse prognosis in hepatocellular carcinoma, which is
characterized by a high frequency of recurrence, large tumor size,
high tumor-node-metastasis stage and prominent vascular invasion
(23,29). In oral squamous cell carcinoma,
increased MIF expression correlates with a higher pathological (p)T
and pN status, positive perineural invasion and tumor depth
(30). In addition, in metastatic
melanoma, high MIF expression is associated with faster disease
progression (31). In addition, high
MIF mRNA expression in pancreatic ductal carcinoma correlates with
a poor survival when compared with tumors exhibiting low MIF mRNA
expression (32).
With regard to circulating MIF, Zhao et al
(23) reported that an increase in
MIF serum level to 90 ng/ml (normal value, 15 ng/ml) corresponded
to a poor prognosis for patients with hepatocellular carcinoma. A
similar observation was reported in colorectal cancer, where serum
MIF levels were elevated in patients with hepatic or lymphoid
metastasis when compared with those without metastasis (11). Furthermore, gastric cancer patients
with a serum MIF level of ≥6,600 pg/ml exhibited a poorer prognosis
than those with lower serum MIF levels (33).
In our previous study, it was demonstrated that
serum MIF levels were higher in head and neck squamous cell
carcinoma patients compared with healthy volunteers, and high MIF
immunostaining in tumor tissues was found to correlate with a poor
prognosis in terms of local tumor recurrence, nodal metastasis
involvement and overall survival (15). Recently, similar results have been
reported in oral squamous cell carcinoma and gastric cancer
(30,34). Overall, these clinical studies
indicate that MIF expression levels in serum and tumor tissue may
be of prognostic value in numerous cancer types.
MIF and cell proliferation
Shi et al (35)
demonstrated that MIF interacts with the CD74/CD44 receptor complex
in stably transfected mammalian kidney COS-7/M6 cells, leading to
MIF signal transduction via the activation of Src, Ras, MAPK kinase
and extracellular-signal-regulated kinase (ERK). It is hypothesized
that this pathway may account for enhanced proliferative activity
(31). Indeed, the interaction of MIF
with its receptor CD74 results in the stimulation of ERK1/2 leading
to cyclin D1 expression (36) and
enhancement of proliferative activity (35). In addition, MIF may enhance cell
proliferation by activating not only the ERK1/2 pathway but also
the phosphoinositide 3-kinase/AKT pathway. Notably, in a previous
study, immunocytochemistry and western blotting revealed that the
addition of recombinant human MIF to the cell medium of the gastric
cancer MGC-803 cell line resulted in increased AKT phosphorylation
(37).
MIF has been also reported to act as a potent
inhibitor of the transcriptional activity of p53 by direct
interaction between MIF and p53, and stabilization of the p53-MDM2
complex (38). This association
prevents p53 translocation from the cytoplasm to the nucleus, and
consequently represses the p53-induced cell cycle arrest and
apoptosis, further supporting cell survival.
Several studies have confirmed that MIF contributes
to cell proliferation in cancer. One previous study demonstrated
that MIF expression was downregulated by short hairpin (sh)RNA in
the murine ovarian cancer ID8 cell line, and the proliferation of
inoculated tumor cells was reduced when compared with
mock-transfected cells, as revealed by a decrease in the percentage
of Ki-67-immunopositive cells (39).
MIF-silencing studies have revealed comparable reductions in the
tumor cell proliferation of human melanoma and hepatocellular
carcinoma cell lines, whereby a decrease in cyclin D1 expression
was reported in vitro and in vivo, respectively
(31,20). Another study reported that
cyclin-dependent kinase 4, cyclin D2 and cyclin E2 expression was
downregulated following MIF-knockdown in HeLa 229 cells (22). In hepatocellular carcinoma, increased
MIF expression, induced by cell transfection, potentiated the
promoter activity of the hepatopoietin gene (a mediator involved in
liver regeneration), leading to enhanced cell proliferation
(40). Hu et al (41) also demonstrated that MIF potentiates
cell proliferation, as the release of MIF from human hepatic
sinusoidal endothelial cells (HHSECs) increased colorectal
carcinoma cell proliferation. Notably, nude mice injected with
colorectal cell lines mixed with mock-transfected HHSECs developed
larger tumors than mice injected with cancer cells mixed with
anti-MIF short hairpin (sh)RNA-transfected HHSECs (41).
Pharmacological inhibition of MIF by small molecule
antagonists reduces cell proliferation. For example, in a previous
study, a BrdU assay revealed that in glioblastoma cell lines
treated with isoxazoline (ISO)-1, cell proliferation was inhibited
in a concentration-dependent manner (42). Similarly, the use of MIF or CD74
neutralizing antibodies has been demonstrated to inhibit human
prostate cancer cell (DU-145) proliferation (14).
In our previous study (15), shRNA-mediated MIF-knockdown in a
murine squamous cell carcinoma cell line (SCCVII) decreased cell
proliferation when compared with a control scramble shRNA cell
line. These findings were confirmed in vivo by the observed
reduction in tumor growth following injection of the SCCVII
MIF-knockdown cells into C3H/Hen mice compared with mice inoculated
with scramble RNA-transfected cells (15). Finally, treatment of SCCVII cells with
the MIF inhibitor 4-IPP (4-iodo-6-phenylpyrimidine) resulted in a
dose-dependent decrease in cell proliferation (43). Thus, these findings indicate that MIF
exhibits a critical function in tumor cell proliferation via its
CD74 receptor, which activates the MAPK and AKT pathways.
MIF and tumor cell invasion
Tumor invasion and metastasis are highly critical in
cancer progression. Several studies have indicated that MIF
contributes to cell invasiveness in cancer. Notably, Rendon et
al (10) revealed that in lung
adenocarcinoma, MIF promotes Ras-related C3 botulinum toxin
substrate 1 activity and thus, tumor cell motility via lipid raft
activation. Another study reported that colorectal cancer cells
cultured in medium conditioned with mock-transfected HHSECs exhibit
increased intracellular F-actin expression compared with cells
cultured in medium conditioned by anti-MIF shRNA-transfected
HHSECs, indicating that cytoskeletal remodeling promotes colorectal
cancer cell migration (41). Ren
et al (44) revealed that
decreased MIF expression in a neuroblastoma cell line reduces cell
invasiveness in vitro and metastasis formation in
vivo: 70% of athymic nude mice injected intravenously with
MIF-expressing cells developed lung metastasis, whereas only 10–20%
of mice developed lung metastasis when MIF was underexpressed.
Furthermore, exposure to antibodies raised against MIF or CD74 has
been shown to block the in vitro migration of prostate
cancer DU-145 cells through Matrigel-coated membranes (14). In pancreatic cancer cell lines, the
overexpression of MIF was associated with an increase in cell
invasiveness in vitro and in vivo, with distant
metastasis to the liver, spleen, lymph nodes and intestine
(32). Furthermore, the
MIF-associated promotion of cell invasiveness in vitro has
also been demonstrated in colorectal carcinoma, nasopharyngeal
carcinoma, adenoid cystic carcinoma and oral squamous cell
carcinoma (11,30,45,46).
Notably, matrix metalloproteinase-9 (MMP-9) expression appears to
be correlated with MIF expression, as positive immunostaining of
MIF increases with that of MMP-9, and MMP-9 is associated with cell
invasion in adenoid cystic carcinoma tissue (46).
Consistent with these studies, in our previous
study, SCCVII cells treated with 4-IPP exhibited a significantly
lower capacity for migration through Matrigel-coated membranes when
compared with untreated cells, which demonstrated the function of
MIF in cell invasion (43). Taken
together, these observations indicate the involvement of MIF in
tumor invasion and metastasis.
MIF and angiogenesis
Angiogenesis is essential for tumor progression, as
neovascularization sustains tumor cell activity, survival and
dissemination (47). Hypoxia promotes
vessel growth via activation of proliferation and migration of
endothelial cells. This involves stabilization of hypoxia-induced
factor-1α (HIF-1α) through two methods that link MIF and HIF1α: i)
In the extracellular environment, MIF binding to CD74 induces
direct activation of HIF1α, and ii) in the intracellular
compartment, MIF can bind Jab/CSN5, which regulates the stability
of HIF1α by preventing its hydroxylation (48), leading to the expression of
pro-angiogenic factors such as IL-8 and VEGF. In this context, MIF
is involved in angiogenesis, demonstrated most notably by a study
in breast cancer tissue, whereby a correlation was identified
between MIF and IL-8 expression (49). The same study also demonstrated that
increased levels of serum MIF were correlated with an increase in
IL-8 serum level (49). The
association between MIF and IL-8 was also confirmed in
vitro: The addition of recombinant human MIF to breast cancer
MDA-MB-231 and MCF7 cell line culture medium stimulated the
secretion of VEGF and IL-8 (50). The
increase in VEGF mRNA expression following exposure to recombinant
human MIF was also reported in bladder cancer (51). Similarly, in non-small cell lung
cancer, two studies reported that the level of CXC chemokine
increased with MIF expression (51)
and that MIF is required to bind its CD74 receptor to subsequently
activate the transcription of CXCL8 and VEGF genes
(52), which confirms that MIF is
implicated in angiogenesis. In addition, studies have demonstrated
that MIF immunostaining correlates with tumor microvessel density
in hepatocellular carcinoma, and that recombinant human MIF
stimulates endothelial tube formation in vitro, as
demonstrated by human umbilical vein endothelial cell assays
(53). Similarly, addition of
exogenous MIF resulted in an increase of VEGF mRNA in the
colorectal LoVo cell line (11). In
addition, a previous study revealed that the MIF−/−
mouse model of melanoma exhibits reduced vessel density compared
with the wild-type model, as demonstrated by decreased
immunostaining of the endothelial marker CD31 in B16-F10 tumors
(54). Fu et al (55) reported that hypoxia in a 3%
O2 atmosphere increases MIF expression in human vascular
smooth muscle cells, further supporting the hypothesis that MIF and
HIF-1α mediate the response to hypoxia in such a model. Therefore,
these results demonstrate the function of MIF in angiogenesis.
MIF and cancer therapy
MIF may present a novel therapeutic target in the
field of oncology. The inhibition of the effect of MIF on tumor
cells may be achieved by the use small molecule inhibitors, such as
ISO-1 and 4-IPP, with high bioavailability and low toxicity, by the
inhibition of MIF activity by neutralizing antibodies and by
targeting its receptor, CD74.
The small inhibitory agents ISO-1 and 4-IPP have
been demonstrated to inhibit the effects of MIF in cancer. ISO-1
exerts an inhibitory action on cell migration and invasion in
vitro in adenoid cystic carcinoma, as demonstrated by a
decrease in MMP-9 expression (46).
Additionally, an analog of ISO-1, ISO-66, has recently been
reported to be active in melanoma and colon cancer models, where it
decreases tumor growth by stimulating an antitumor immune response
in vivo (56). Regarding head
and neck carcinoma, our previous study investigated the effect of
4-IPP on SCCVII cells and reported that the pharmacological
inhibition of MIF resulted in impaired proliferation and
invasiveness in vitro (43).
Recently, Mawhinney et al (57) developed a novel inhibitor of MIF
enzymatic activity (SCD-19), which was demonstrated to markedly
inhibit the growth of nascent and established murine Lewis lung
carcinoma.
The efficacy of human anti-MIF antibodies in human
carcinoma has been reported by Hussain et al (58). The study revealed that the anti-MIF
antibodies, BaxG03 and BaxB0, inhibit cell proliferation via the
reduction of the phosphorylation of ERK1/2 and AKT in prostate
cancer cell lines. It was also revealed that these antibodies
decreased the migration of the PC3 cell line, with a half-maximal
inhibition in the range of 2–10 nm/l. In addition, the study
evaluated the in vivo efficacy of several anti-MIF
antibodies (BaxG03, BaxB01 and BaxM159) in a xenograft model of
prostate cancer and observed that the tumor size was reduced in
mice that were administered antibodies for 30 days when compared
with the control. Furthermore, the number of Ki-67-positive cells
diminished after treatment compared with the control. In this
study, the median effective dose was 8–14 mg/kg (58).
Regarding the mechanism of action of MIF, its
receptor, CD74, may also present a promising therapeutic target to
inhibit MIF signaling. In this context, a CD74 specific monoclonal
antibody, milatuzumab, has been developed, which has been reported
to enhance the action of doxorubicin in multiple myeloma cell lines
(59). At present, a multiple myeloma
phase I clinical trial combining milatuzumab and doxorubicin is
ongoing (clinicaltrials.gov identifier,
NCT01101594).
In our previous study, the effect of MIF
underexpression on mouse survival following inoculation with SCCVII
cells and subsequent treatment with cisplatin or 5-fluorouracil was
evaluated (15). The results revealed
that the tumors growing after the injection of MIF-knockdown cells
were more sensitive to cisplatin (4 mg/kg twice a week for 2 weeks)
and 5-fluorouracil (20 mg/kg daily for 4 days) treatment than
tumors developing from control cells, which further supports the
hypothesis that MIF downregulation may potentiate the effect of
chemotherapy agents.
Furthermore, a recent study reported that
pharmacological inhibition of the ATPase activity of heat shock
protein 90 (HSP90) resulted in the degradation of MIF in numerous
types of cancer cell (60). Thus,
targeting HSP90 may present a novel strategy for the inhibition of
MIF in cancer.
Conclusion
In this review, the effect of MIF on tumor cell
proliferation, migration and tumor-induced angiogenesis in clinical
and experimental studies was discussed (Table I). Overexpression of MIF in tumor
tissue, as well as high levels of MIF in patient serum, have
demonstrated prognostic value associated with a short survival
time. The poor clinical outcome of cancer patients exhibiting high
MIF expression may be explained by its potentiating effects on
proliferation, invasion and angiogenesis (Fig. 1). Indeed, binding of MIF to its
receptor may stimulate the MAPK pathway and inhibit p53, each
positively impacting tumor cell proliferation and survival. In
addition, the overexpression of MIF was correlated with the
overexpression of MMP-9, further supporting its role in
invasiveness. Furthermore, the action of MIF on angiogenesis may
occur through the activation of HIF1α, and subsequently, the
production of pro-angiogenic factors. A number of the effects of
MIF may be driven by interaction with its membrane receptor CD74,
while interaction with the chemokine receptor CXCR4 has also been
proposed. Significantly, MIF also inhibits p53, inhibiting cell
cycle arrest and apoptosis (Fig. 1).
Altogether, such data provides a novel understanding of the
potential role of MIF in the mechanisms of resistance to cancer
therapy.
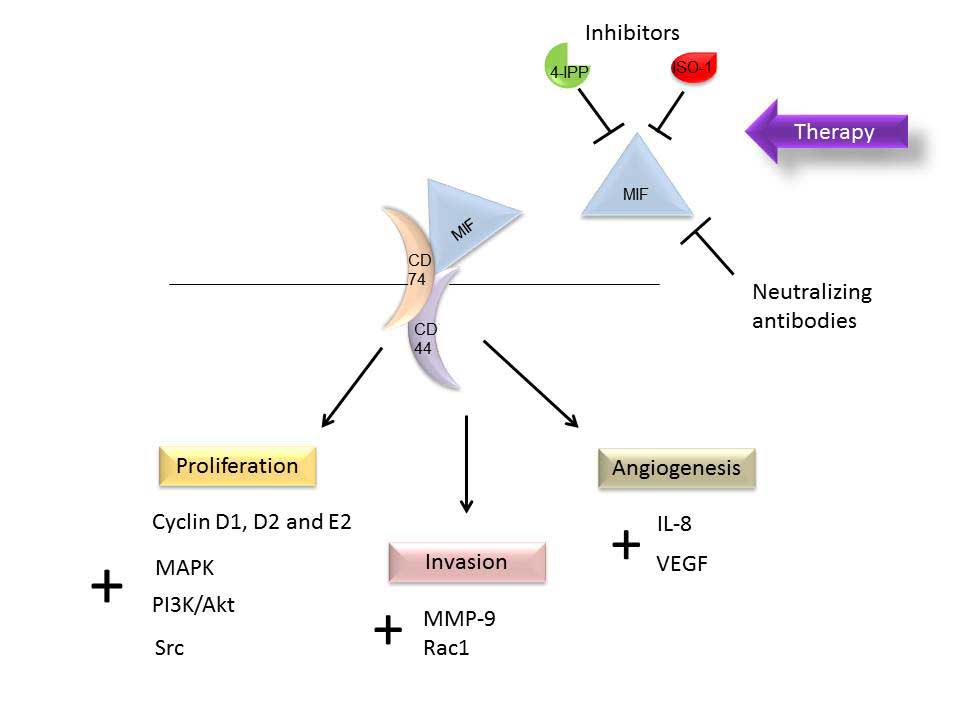 | Figure 1.Schematic representation of the
involvement of MIF in cancer, which affects several pathways in
proliferation, invasion and angiogenesis, via interaction with its
membrane receptor, CD74. Small molecule inhibitors, 4-IPP and
ISO-1, and neutralizing antibodies have demonstrated an inhibitory
effect on MIF. MIF, migration inhibitory factor; CD, cluster of
differentiation; 4-IPP, 4-iodo-6-phenylpyrimidine; ISO-1,
isoxazoline-1; IL, interleukin; MMP-9, matrix metalloproteinase-9;
MAPK, mitogen-activated protein kinase; PI3K, phosphoinositide
3-kinase; VEGF, vascular endothelial growth factor; Rac1,
Ras-related C3 botulinum toxin substrate 1. |
 | Table I.Overview of previous literature
investigating the function of MIF and/or CD74 in carcinogenesis,
prognosis, proliferation, invasion, angiogenesis and reported
responses to targeted therapy. |
Table I.
Overview of previous literature
investigating the function of MIF and/or CD74 in carcinogenesis,
prognosis, proliferation, invasion, angiogenesis and reported
responses to targeted therapy.
| Cancer type |
Carcinogenesisa, refs. |
Prognosisb, refs. |
Proliferationc, refs. |
Invasiond, refs. |
Angiogenesise, refs. |
Therapyf,
refs. |
|---|
| Adenoid cystic
carcinoma |
|
|
| 4 |
| 45 |
| Bladder |
|
|
|
| 50 |
|
| Breast | 12 |
|
|
| 49 |
|
| Cervical | 13 |
|
|
|
|
|
| Colorectal |
| 11 | 41 | 11,41 | 11 | 56 |
| Gastric | 18 | 33,34 | 37 |
|
|
|
| Glioma | 22 |
| 42 |
|
|
|
| Head and neck | 24,25,26,15 | 30,15 |
| 30,43,45 |
| 43 |
| Liver | 21,23 | 23,29 | 21,40 |
| 53 |
|
| Lung |
| 28 |
| 10 | 51,52 | 57 |
| Melanoma | 20 | 31 | 31 |
| 54 | 56 |
| Multiple
myeloma |
|
|
|
|
| 59 |
| Neuroblastoma |
|
|
| 44 |
|
|
| Pancreatic | 19 | 32 |
| 32 |
|
|
| Prostate |
|
| 14 | 14 |
| 58 |
Thus, as MIF is implicated in numerous aspects of
cancer progression, via direct activation of its CD74 receptor and
direct inhibition of the tumor suppressor p53, inhibition of this
factor and/or of its receptor may present a novel treatment
strategy against cancer, alone or in combination with conventional,
as well as targeted therapies.
Acknowledgements
The present review was supported by a grant (no.
7.4656.14) from the National Fund for Scientific Research and the
Honorary Senior Research Associate of the National Fund for
Scientific Research (Belgium).
References
|
1
|
Bloom and Bennett. Mechanism of a reaction
in vitro associated with delayed-type hypersensitivity. Science.
153:80–82. 1966. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun HW, Bernhagen J, Bucala R and Lolis E:
Crystal structure at 2.6-A resolution of human macrophage migration
inhibitory factor. Proc Natl Acad Sci USA. 93:5191–5196. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leng L and Bucala R: Insight into the
biology of macrophage migration inhibitory factor (MIF) revealed by
the cloning of its cell surface receptor. Cell Res. 16:162–168.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rosengren E, Bucala R, Aman P, Jacobsson
L, Odh G, Metz CN and Rorsman H: The immunoregulatory mediator
macrophage migration inhibitory factor (MIF) catalyzes a
tautomerization reaction. Mol Med. 2:143–149. 1996.PubMed/NCBI
|
|
5
|
Conroy H, Mawhinney L and Donnelly SC:
Inflammation and cancer: Macrophage migration inhibitory factor
(MIF)-the potential missing link. QJM. 103:831–836. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nishino T, Bernhagen J, Shiiki H, Calandra
T, Dohi K and Bucala R: Localization of macrophage migration
inhibitory factor (MIF) to secretory granules within the
corticotrophic and thyrotrophic cells of the pituitary gland. Mol
Med. 1:781–788. 1995.PubMed/NCBI
|
|
7
|
Roger T, Chanson AL, KnaupReymond M and
Calandra T: Macrophage migration inhibitory factor promotes innate
immune responses by suppressing glucocorticoid-induced expression
of mitogen-activated protein kinase phosphatase-1. Eur J Immunol.
35:3405–3413. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Santos LL and Morand EF: Macrophage
migration inhibitory factor: A key cytokine in RA, SLE and
atherosclerosis. Clin Chim Acta. 399:1–7. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sánchez-Zamora YI and Rodriguez-Sosa M:
The role of MIF in type 1 and type 2 diabetes mellitus. J Diabetes
Res. 2014:8045192014.PubMed/NCBI
|
|
10
|
Rendon BE, Roger T, Teneng I, Zhao M,
AlAbed Y, Calandra T and Mitchell RA: Regulation of human lung
adenocarcinoma cell migration and invasion by macrophage migration
inhibitory factor. J Biol Chem. 282:29910–29918. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He XX, Chen K, Yang J, Li XY, Gan HY, Liu
CY, Coleman TR and Al-Abed Y: Macrophage migration inhibitory
factor promotes colorectal cancer. Mol Med. 15:1–10. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Richard V, Kindt N, Decaestecker C, Gabius
HJ, Laurent G, Noël JC and Saussez S: Involvement of macrophage
migration inhibitory factor and its receptor (CD74) in human breast
cancer. Oncol Rep. 32:523–529. 2014.PubMed/NCBI
|
|
13
|
Guo P, Wang J, Liu J, Xia M, Li W and He
M: Macrophage immigration inhibitory factor promotes cell
proliferation and inhibits apoptosis of cervical adenocarcinoma.
Tumour Biol. 36:5095–5102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
MeyerSiegler KL, Iczkowski KA, Leng L,
Bucala R and Vera PL: Inhibition of macrophage migration inhibitory
factor or its receptor (CD74) attenuates growth and invasion of
DU-145 prostate cancer cells. J Immunol. 177:8730–8739. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kindt N, Preillon J, Kaltner H, Gabius HJ,
Chevalier D, Rodriguez A, Johnson BD, Megalizzi V, Decaestecker C,
Laurent G and Saussez S: Macrophage migration inhibitory factor in
head and neck squamous cell carcinoma: Clinical and experimental
studies. J Cancer Res Clin Oncol. 139:727–737. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kindt N, Lechien JR, Nonclercq D, Laurent
G and Saussez S: Involvement of CD74 in head and neck squamous cell
carcinomas. J Cancer Res Clin Oncol. 140:937–947. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dessein AF, Stechly L, Jonckheere N,
Dumont P, Monté D, Leteurtre E, Truant S, Pruvot FR, Figeac M,
Hebbar M, et al: Autocrine induction of invasive and metastatic
phenotypes by the MIF-CXCR4 axis in drug-resistant human colon
cancer cells. Cancer Res. 70:4644–4654. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He XX, Yang J, Ding YW, Liu W, Shen QY and
Xia HH: Increased epithelial and serum expression of macrophage
migration inhibitory factor (MIF) in gastric cancer: Potential role
of MIF in gastric carcinogenesis. Gut. 55:797–802. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tan L, Ye X, Zhou Y, Yu M, Fu Z, Chen R,
Zhuang B, Zeng B, Ye H, Gao W, et al: Macrophage migration
inhibitory factor is overexpressed in pancreatic cancer tissues and
impairs insulin secretion function of β-cell. J Transl Med.
12:922014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heise R, VetterKauczok CS, Skazik C, Czaja
K, Marquardt Y, Lue H, Merk HF, Bernhagen J and Baron JM:
Expression and function of macrophage migration inhibitory factor
in the pathogenesis of UV-induced cutaneous nonmelanoma skin
cancer. Photochem Photobiol. 88:1157–1164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang XH, Jian WH, Wu ZF, Zhao J, Wang H,
Li W and Xia JT: Small interfering RNA (siRNA)-mediated knockdown
of macrophage migration inhibitory factor (MIF) suppressed cyclin
D1 expression and hepatocellular carcinoma cell proliferation.
Oncotarget. 5:5570–5580. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mittelbronn M, Platten M, Zeiner P,
Dombrowski Y, Frank B, Zachskorn C, Harter PN, Weller M and
Wischhusen J: Macrophage migration inhibitory factor (MIF)
expression in human malignant gliomas contributes to immune escape
and tumour progression. Acta Neuropathol. 122:353–365. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao YM, Wang L, Dai Z, Wang DD, Hei ZY,
Zhang N, Fu XT, Wang XL, Zhang SC, Qin LX, et al: Validity of
plasma macrophage migration inhibitory factor for diagnosis and
prognosis of hepatocellular carcinoma. Int J Cancer. 129:2463–2472.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
DeSouza MB, Curioni OA, Kanda JL and De
Carvalho MB: Serum and salivary macrophage migration inhibitory
factor in patients with oral squamous cell carcinoma. Oncol Lett.
8:2267–2275. 2014.PubMed/NCBI
|
|
25
|
Cludts S, Decaestecker C, Johnson B,
Lechien J, Leroy X, Kindt N, Kaltner H, André S, Gabius HJ and
Saussez S: Increased expression of macrophage migration inhibitory
factor during progression to hypopharyngeal squamous cell
carcinoma. Anticancer Res. 30:3313–3319. 2010.PubMed/NCBI
|
|
26
|
Kindt N, Lechien J, Decaestecker C,
Rodriguez A, Chantrain G, Remmelink M, Laurent G, Gabius HJ and
Saussez S: Expression of macrophage migration-inhibitory factor is
correlated with progression in oral cavity carcinomas. Anticancer
Res. 32:4499–4505. 2012.PubMed/NCBI
|
|
27
|
Kamimura A, Kamachi M, Nishihira J, Ogura
S, Isobe H, DosakaAkita H, Ogata A, Shindoh M, Ohbuchi T and
Kawakami Y: Intracellular distribution of macrophage migration
inhibitory factor predicts the prognosis of patients with
adenocarcinoma of the lung. Cancer. 89:334–341. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tomiyasu M, Yoshino I, Suemitsu R, Okamoto
T and Sugimachi K: Quantification of macrophage migration
inhibitory factor mRNA expression in non-small cell lung cancer
tissues and its clinical significance. Clin Cancer Res.
8:3755–3760. 2002.PubMed/NCBI
|
|
29
|
Wang D, Luo L, Chen W, Chen LZ, Zeng WT,
Li W and Huang XH: Significance of the vascular endothelial growth
factor and the macrophage migration inhibitory factor in the
progression of hepatocellular carcinoma. Oncol Rep. 31:1199–1204.
2014.PubMed/NCBI
|
|
30
|
Chang KP, Lin SJ, Liu SC, Yi JS, Chien KY,
Chi LM, Kao HK, Liang Y, Lin YT, Chang YS and Yu JS:
Low-molecular-mass secretome profiling identifies HMGA2 and MIF as
prognostic biomarkers for oral cavity squamous cell carcinoma. Sci
Rep. 5:116892015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Oliveira CS, de Bock CE, Molloy TJ,
Sadeqzadeh E, Geng XY, Hersey P, Zhang XD and Thorne RF: Macrophage
migration inhibitory factor engages PI3K/AKT signalling and is a
prognostic factor in metastatic melanoma. BMC Cancer. 14:6302014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Funamizu N, Hu C, Lacy C, Schetter A,
Zhang G, He P, Gaedcke J, Ghadimi MB, Ried T and Yfantis HG:
Macrophage migration inhibitory factor induces epithelial to
mesenchymal transition, enhances tumor aggressiveness and predicts
clinical outcome in resected pancreatic ductal adenocarcinoma. Int
J Cancer. 132:785–794. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xia HH, Yang Y, Chu KM, Gu Q, Zhang YY, He
H, Wong WM, Leung SY, Yuen ST, Yuen MF, et al: Serum macrophage
migration-inhibitory factor as a diagnostic and prognostic
biomarker for gastric cancer. Cancer. 115:5441–5449. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He LJ, Xie D, Hu PJ, Liao YJ, Deng HX,
Kung HF and Zhu SL: Macrophage migration inhibitory factor as a
potential prognostic factor in gastric cancer. World J
Gastroenterol. 21:9916–9926. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi X, Leng L, Wang T, Wang W, Du X, Li J,
McDonald C, Chen Z, Murphy JW, Lolis E, et al: CD44 is the
signaling component of the macrophage migration inhibitory
factor-CD74 receptor complex. Immunity. 25:595–606. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mitchell RA, Liao H, Chesney J,
FingerleRowson G, Baugh J, David J and Bucala R: Macrophage
migration inhibitory factor (MIF) sustains macrophage
proinflammatory function by inhibiting p53: Regulatory role in the
innate immune response. Proc Natl Acad Sci USA. 99:345–350. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li GQ, Xie J, Lei XY and Zhang L:
Macrophage migration inhibitory factor regulates proliferation of
gastric cancer cells via the PI3K/AKT pathway. World J
Gastroenterol. 15:5541–5548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jung H, Seong HA and Ha H: Critical role
of cysteine residue 81 of macrophage migration inhibitory factor
(MIF) in MIF-induced inhibition of p53 activity. J Biol Chem.
283:20383–20396. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hagemann T, Robinson SC, Thompson RG,
Charles K, Kulbe H and Balkwill FR: Ovarian cancer cell-derived
migration inhibitory factor enhances tumor growth, progression, and
angiogenesis. Mol Cancer Ther. 6:1993–2002. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li Y, Lu C, Xing G, Zhu Y and He F:
Macrophage migration inhibitory factor directly interacts with
hepatopoietin and regulates the proliferation of hepatoma cell. Exp
Cell Res. 300:379–387. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu CT, Guo LL, Feng N, Zhang L, Zhou N, Ma
LL, Shen L, Tong GH, Yan QW, Zhu SJ, et al: MIF, secreted by human
hepatic sinusoidal endothelial cells, promotes chemotaxis and
outgrowth of colorectal cancer in liver prometastasis. Oncotarget.
6:22410–24423. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Baron N, Deuster O, Noelker C, Stüer C,
Strik H, Schaller C, Dodel R, Meyer B and Bacher M: Role of
macrophage migration inhibitory factor in primary glioblastoma
multiforme cells. J Neurosci Res. 89:711–717. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kindt N, Laurent G, Nonclercq D, Journé F,
Ghanem G, Duvillier H, Gabius HJ, Lechien J and Saussez S:
Pharmacological inhibition of macrophage migration inhibitory
factor interferes with the proliferation and invasiveness of
squamous carcinoma cells. Int J Oncol. 43:185–193. 2013.PubMed/NCBI
|
|
44
|
Ren Y, Chan HM, Fan J, Xie Y, Chen YX, Li
W, Jiang GP, Liu Q, Meinhardt A and Tam PK: Inhibition of tumor
growth and metastasis in vitro and in vivo by targeting macrophage
migration inhibitory factor in human neuroblastoma. Oncogene.
25:3501–3508. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pei XJ, Wu TT, Li B, Tian XY, Li Z and
Yang QX: Increased expression of macrophage migration inhibitory
factor and DJ-1 contribute to cell invasion and metastasis of
nasopharyngeal carcinoma. Int J Med Sci. 11:106–115. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu H, Chen G, Zhang W, Zhu JY, Lin ZQ,
Gong ZC, Wang FQ, Jia J, Sun ZJ and Zhao YF: Overexpression of
macrophage migration inhibitory factor in adenoid cystic carcinoma:
Correlation with enhanced metastatic potential. J Cancer Res Clin
Oncol. 139:287–295. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Oda S, Oda T, Nishi K, Takabuchi S,
Wakamatsu T, Tanaka T, Adachi T, Fukuda K, Semenza GL and Hirota K:
Macrophage migration inhibitory factor activates hypoxia-inducible
factor in a p53-dependent manner. PLoS One. 3:e22152008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu X, Wang B, Ye C, Yao C, Lin Y, Huang X,
Zhang Y and Wang S: Overexpression of macrophage migration
inhibitory factor induces angiogenesis in human breast cancer.
Cancer Lett. 261:147–157. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Choudhary S, Hegde P, Pruitt JR, Sielecki
TM, Choudhary D, Scarpato K, Degraff DJ, Pilbeam CC and Taylor JA
III: Macrophage migratory inhibitory factor promotes bladder cancer
progression via increasing proliferation and angiogenesis.
Carcinogenesis. 34:2891–2899. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
White ES, Flaherty KR, Carskadon S, Brant
A, Iannettoni MD, Yee J, Orringer MB and Arenberg DA: Macrophage
migration inhibitory factor and CXC chemokine expression in
non-small cell lung cancer: Role in angiogenesis and prognosis.
Clin Cancer Res. 9:853–860. 2003.PubMed/NCBI
|
|
52
|
Coleman AM, Rendon BE, Zhao M, Qian MW,
Bucala R, Xin D and Mitchell RA: Cooperative regulation of
non-small cell lung carcinoma angiogenic potential by macrophage
migration inhibitory factor and its homolog, D-dopachrome
tautomerase. J Immunol. 181:2330–2337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hira E, Ono T, Dhar DK, ElAssal ON,
Hishikawa Y, Yamanoi A and Nagasue N: Overexpression of macrophage
migration inhibitory factor induces angiogenesis and deteriorates
prognosis after radical resection for hepatocellular carcinoma.
Cancer. 103:588–598. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Girard E, Strathdee C, Trueblood E and
Quéva C: Macrophage migration inhibitory factor produced by the
tumour stroma but not by tumour cells regulates angiogenesis in the
B16-F10 melanoma model. Br J Cancer. 107:1498–1505. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Fu H, Luo F, Yang L, Wu W and Liu X:
Hypoxia stimulates the expression of macrophage migration
inhibitory factor in human vascular smooth muscle cells via
HIF-1alpha dependent pathway. BMC Cell Biol. 11:662010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ioannou K, Cheng KF, Crichlow GV,
Birmpilis AI, Lolis EJ, Tsitsilonis OE and Al-Abed Y: ISO-66, a
novel inhibitor of macrophage migration, shows efficacy in melanoma
and colon cancer models. Int J Oncol. 45:1457–1468. 2014.PubMed/NCBI
|
|
57
|
Mawhinney L, Armstrong ME, O'Reilly C,
Bucala R, Leng L, Fingerle-Rowson G, Fayne D, Keane MP, Tynan A,
Maher L, et al: Macrophage migration inhibitory factor (MIF)
enzymatic activity and lung cancer. Mol Med. 20:729–735.
2015.PubMed/NCBI
|
|
58
|
Hussain F, Freissmuth M, Völkel D, Thiele
M, Douillard P, Antoine G, Thurner P, Ehrlich H, Schwarz HP,
Scheiflinger F and Kerschbaumer RJ: Human anti-macrophage migration
inhibitory factor antibodies inhibit growth of human prostate
cancer cells in vitro and in vivo. Mol Cancer Ther. 12:1223–1234.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Stein R, Mattes MJ, Cardillo TM, Hansen
HJ, Chang CH, Burton J, Govindan S and Goldenberg DM: CD74: A new
candidate target for the immunotherapy of B-cell neoplasms. Clin
Cancer Res. 13:5556s–5563s. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Schulz R and Moll UM: Targeting the heat
shock protein 90: A rational way to inhibit macrophage migration
inhibitory factor function in cancer. Curr Opin Oncol. 26:108–113.
2014. View Article : Google Scholar : PubMed/NCBI
|














