Introduction
Hyaluronan (HA) is a high molecular weight
glycosaminoglycan composed of repeating disaccharide units of
D-N-acetylglucosamine (GlcNAc) and D-glucuronic acid (GlcUA)
in the structure GlcNAc-β-(1→4)GlcUA-β-(1→3) (1). Due to its physicochemical composition,
HA is able to trap large amounts of water, and its solutions are
viscous and elastic (1). HA is
synthesized from the precursors uridine 5′-diphosphoglucoronic acid
(UDP-GlcUA) and uridine diphosphate N-acetylglucosamine
(UDP-GlcNAc) (2). The synthesis
occurs at the inner leaflet of the plasma membrane by a
membrane-associated enzyme termed hyaluronan synthase (HAS)
(2). In numerous tissues, HA is a
major component of the extracellular matrix (ECM) and is
non-covalently bound to other components (2). In normal tissues, the presence of HA
creates an environment that facilitates cellular proliferation and
migration (3). Additionally, HA is
required for certain physiological processes such as embryonic
development and wound healing (3). In
cancer, however, the properties of HA enhance tumor invasion,
growth, angiogenesis and metastasis (3–5). Previous
studies have revealed that HA participates in the cell adhesion
(6) and migration (7) phases of tumor metastasis. In several
human tumors of epithelial origin, including breast (4), ovarian (8), prostate (9), gastric (10) and colon cancer (11), accumulation of HA in the tumor tissue
was associated with invasive and metastatic properties, and led to
a worse prognosis.
Previous studies have used 4-methylumbelliferone
(MU) as a fluorescent indicator to measure various types of
enzymatic activity (12). The present
authors have reported that MU inhibits HA synthesis and ECM
formation in cultured human skin fibroblasts (13). Using rat 3Y1 fibroblasts that stably
expressed HAS2, a novel mechanism of MU-mediated inhibition of HA
synthesis was demonstrated, in which glucuronidation of MU by
endogenous uridine diphosphate (UDP)-glucuronyl transferase results
in depletion of UDP-GlcUA (14).
Subsequently, the present authors reported that HA synthesis in
B16F-10 melanoma cells is suppressed by MU, and that knockdown of
extracellular HA decreases cellular adherence and locomotion in
vitro (15). In addition, the
present authors have demonstrated that suppression of HA in the
liver of C57BL/6 mice reduces inhabitation of metastatic nodules
in vivo when melanoma cells were injected into the lateral
tail vein of the mice (16). MU has
been widely investigated as an inhibitor of HA synthesis, and has
been proposed as an anticancer agent. Piccioni et al
(17) reported that MU induced
apoptosis in mouse hepatocellular carcinoma models, and led to a
decrease in the amount of hepatic stellate cells. Lokeshwar et
al (18) reported that MU
inhibited the proliferation and invasion of prostate cancer cells,
while Pályi-Krekk et al (19)
demonstrated that MU reduced the amount of HA in breast cancer
cells, which led to enhanced binding of trastuzumab.
Pancreatic cancer is one of the most malignant
neoplasms, and 80–85% of patients present with advanced
unresectable tumors (20). The annual
number of mortalities caused by pancreatic cancer has been
gradually increasing, while the prevalence and mortality of other
common types of cancer have been declining (21). Despite continuous improvements in the
detection and management of pancreatic cancer, only 4% of patients
live for 5 years subsequent to diagnosis (20–22). One
of the major reasons for this dismal prognosis is the poor response
of pancreatic cancer cells to the majority of chemotherapeutic
agents currently available (20).
Therefore, a novel therapeutic agent for the treatment of
progressive pancreatic cancer is urgently required.
Regarding the role of HA in human pancreatic tissue,
a previous immunohistochemical study using human pancreatic cancer
tissues revealed a stronger expression of HA and HAS2 in these
tissues compared with healthy pancreas tissues, and that increased
expression of HA and HAS2 was associated with a significantly
poorer prognosis (23). Accordingly,
the present authors considered the possibility that MU exerts an
anticancer effect on pancreatic cancer. This hypothesis is
supported by the results of Nakazawa et al (24), who reported that MU inhibited HA
synthesis and ECM formation in primary and metastatic tumors of
human pancreatic cancer cells. However, the distribution of HA in
pancreatic cancer tissue remains unknown, and the structural
changes caused by MU in the ECM have not been sufficiently
investigated to date.
In the present study, the in vitro
antiproliferative effect and cytotoxicity of MU were examined in
MIA PaCa-2, a human pancreatic cancer cell line. HA synthesis and
localization in cancer tissues were analyzed quantitatively and
immunohistochemically. Furthermore, MU-mediated structural changes
of cancer cells and ECM in the tumor tissue were investigated using
an electron microscope. The suitability of MU as an anticancer
agent for pancreatic cancer is also discussed.
Materials and methods
Materials
The 4-methylumbelliferone (MU) and hyaluronidase
from Streptomyces hyalurolyticus were purchased from Wako
Pure Chemicals Industries, Ltd. (Osaka, Japan). Actinase E was
purchased from Kaken Pharmaceutical Co., Ltd. (Tokyo, Japan).
Dulbecco's modified Eagle's medium (DMEM) was purchased from
Nacalai Tesque, Inc. (Kyoto, Japan). All other reagents were of
analytical grade and were obtained from commercial sources.
Tumor cells
The human pancreatic cancer cell line MIA PaCa-2 was
kindly provided by the Department of Pharmacy of Hirosaki
University Hospital (Hirosaki, Japan). The cells were routinely
maintained as monolayer cultures in DMEM supplemented with 10%
heat-inactivated fetal bovine serum (Nichirei Biosciences, Inc.,
Tokyo, Japan), L-glutamine (Nacalai Tesque, Inc.), sodium pyruvate
(Nacalai Tesque, Inc.) and antibiotic antimycotic solution
(Sigma-Aldrich Japan Co., LLC., Tokyo, Japan) at 37°C in a mixture
of 5% CO2 and 95% humidified air.
Mice
A total of 30 male C.B-17/lcr-scid mice were
purchased from Japan Clea (Tokyo, Japan). The mice were housed
under controlled light-dark cycles, temperature and humidity, with
water and food ad libitum. The mice were used when they were
6-week-old and weighted ~25 g. All animal experiments were
performed according to the Guidelines for Animal Experimentation of
Hirosaki University (Hirosaki, Japan).
Particle exclusion assay
Cell surface HA was visualized using a particle
exclusion assay. Fixed horse erythrocytes (Nippon Biotest
Laboratories Inc., Tokyo, Japan) were reconstituted in 9.6 mM
phosphate-buffered saline (PBS; pH 7.4; Nacalai Tesque, Inc.) at a
density of 5×108 cells/ml. MIA PaCa-2 cells were
cultured in 100-mm dishes. Extracellular HA was visualized under a
light microscope (IX71N-22/PH; Olympus Corporation, Tokyo, Japan)
by adding the erythrocyte suspension (3 ml) to the growth medium,
with or without MU. To determine the composition of the
extracellular halo, MU-free dishes were pre-incubated with 3.0 U/ml
Streptomyces hyalurolyticus hyaluronidase (Wako Pure
Chemicals Industries, Ltd.) for 1 h prior to the assay (25).
Immunohistochemical staining of
pancreatic cancer cells
Chamber slides were used for staining cells (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Medium (1 ml)
containing 1×104 MIA PaCa-2 cells was added to the
chamber slides. Following 24-h incubation, 0.1–1.0 mM MU dissolved
in dimethyl sulfoxide (DMSO; Wako Pure Chemicals Industries, Ltd.)
was added, ensuring that the concentration of DMSO in the medium
did not exceed 0.1%. Upon 48-h incubation, cells were washed twice
with PBS (2 ml/wash), fixed with 4% paraformaldehyde buffered with
PBS at room temperature for 2 h and rinsed twice with PBS.
Following antigen retrieval, tissue samples were incubated for 32
min at 37°C with 2.5 µg/ml biotinylated HA binding protein (HABP;
catalog no., BC41; Hokudo Co., Ltd., Osaka, Japan). Streptavidin
horseradish peroxidase conjugate and 3,3-diaminobenzidine (DAB)
from the iVIEW DAB universal kit (Ventana Medical Systems, Inc.,
Tucson, AZ, USA) was used to visualize the results. Negative
controls were stained with HABP following digestion of the tissue
sections with 3.0 U/ml Streptomyces hyalurolyticus
hyaluronidase (26).
Analysis of HA synthesis in cells
Medium (9 ml) containing 6×105 MIA PaCa-2
cells was seeded in a 100-mm culture dish and incubated for 24 h.
Cells were incubated with 0.1–1.0 mM MU dissolved in DMSO.
Following 48-h incubation, the medium was removed. Cells were
washed twice with PBS (5 ml/wash) and suspended in PBS (0.5 ml).
The suspension was mixed with 5 ml lysis buffer [1% NP-40, 150 mM
NaCl, 50 mM Tris-HCl buffer (pH 8.0), 0.5% sodium deoxycholate and
0.1% sodium dodecyl sulfate; Wako Pure Chemicals Industries, Ltd.],
vortexed for 1 min and placed on ice for 30 min. Upon
centrifugation (MRX-152; Tomy Seiko Co., Ltd., Tokyo, Japan) of the
suspension at 10,000 × g at 4°C for 10 min, the supernatant
containing HA was transferred into another tube and kept at −80°C
until analysis. HA was measured using an enzyme-linked binding
protein assay kit in a sandwich format, according to the
manufacturer's protocol (Biotech Trading Partners, Encinitas, CA,
USA) (27).
Proteins were quantified using the dye-binding
Bradford method (Bio-Rad Laboratories, Inc., Hercules, CA, USA),
according to the manufacturer's protocol (28). Bovine serum albumin (Sigma-Aldrich
Japan Co., LLC.) in 5 mM Tris-HCl buffer (pH 7.0; Wako Pure
Chemicals Industries, Ltd.) was used as a standard.
Cell growth assay
Medium (2 ml) containing 1×105 cells was
seeded into 6-well plates (Iwaki®; Analytical
Technologies Group, LLC, Groton, CT, USA). Following 12-h
incubation, 0.1–1.0 mM MU dissolved in DMSO was added. Upon 24, 48
or 72-h incubation with MU, each well was washed twice with PBS (5
ml/wash), detached from the plates by addition of 0.5 ml
trypsin/ethylenediaminetetraacetic acid (EDTA; Thermo Fisher
Scientific, Inc.) and suspended in 2 ml PBS. The cell count of the
suspensions was assessed using an automated cell counter (TC20™;
Bio-Rad Laboratories, Inc.) (29).
Lactate dehydrogenase (LDH) assay
The cytotoxicity of MU was analyzed using
LDH-Cytotoxic Test (Wako Pure Chemicals Industries, Ltd.) (30). Medium (50 µl) containing
5×104 cells was added to 96-well plates
(Iwaki®; Analytical Technologies Group, LLC). Following
12-h incubation, 0.1–1.0 mM MU dissolved in DMSO was added, and
cells were further incubated for 2 h. Next, the plates were
centrifuged (MRX-152; Tomy Seiko Co., Ltd.) at 158 × g for 3 min,
and the supernatants were collected. The concentration of LDH in
the supernatants was measured at an absorbance (Abs) wavelength of
560 nm using a microplate spectrophotometer (xMark™; Bio-Rad
Laboratories, Inc.). Cytotoxicity was calculated using the
following formula, where Abssample, Absneg
and Abspos are the Abs values of the sample, negative
control and positive control, respectively:
Cytotoxicity (%) =
(Abssample-Absneg)/(Abspos-Absneg)
× 100
Analysis of tumor growth
A suspension of 4×106 MIA PaCa-2 cells
was subcutaneously injected into the dorsal tissue of the mice, and
2 weeks later, MU was orally administered at a dose of 2 g/kg/day,
mixed with bait in order to reduce stress in the animals. Once a
week, tumor size was compared between the MU-treated (n=14) and
control groups (n=15) using a caliper, and the tumor volume was
calculated as length × width2 x 0.52 (31).
Analysis of HA synthesis in pancreatic
cancer tissue
Mice were euthanized, and the tumors were removed.
The dry weight of each tumor sample was measured upon desiccation
at 60°C for 4 h. Each sample was minced and digested overnight at
55°C with 0.25% actinase E/10 mM Tris-HCl (1 ml; pH 8.0), followed
by incubation at 100°C for 10 min to terminate the reaction. Upon
centrifugation of the suspension at 10,000 × g at 4°C for 10 min,
the supernatant containing HA was collected, and the concentration
of HA was measured using an enzyme-linked binding protein assay kit
in a sandwich format, as previously described (27,32).
Immunohistochemical staining of
tumor
The removed tumor was fixed in 10% formalin (Wako
Pure Chemicals Industries, Ltd.) dissolved in 70% ethanol (Wako
Pure Chemicals Industries, Ltd.) and 5% glacial acetic acid (Wako
Pure Chemicals Industries, Ltd.). The samples were embedded in
paraffin (Wako Pure Chemicals Industries, Ltd.) using routine
procedures (33), sliced at a
thickness of 4 µm (CRM-440; Sakura Finetek Japan Co., Ltd.) and
analyzed using the iVIEW DAB Universal kit. Antigen retrieval was
conducted using cell conditioning solution [CC1-Tris based EDTA
buffer (pH 8.0); Ventana Medical Systems, Inc.], and tissue samples
were then incubated for 32 min at 37°C with 2 µg/ml biotinylated
HABP. The aforementioned iVIEW DAB Universal kit was used to
visualize the results. Negative controls were stained with HABP
following digestion of the tissue sections with 100 U/ml
Streptomyces hyalurolyticus hyaluronidase (34).
Transmission electron microscopy
Several small pieces of tissue established by the
transplantation of MIA PaCa-2 cells were immersed in Karnovsky's
fixative (25% glutaralldehyde, 2% paraformaldehyde and 0.1 M
cacodylate buffer) for >24 h at 4°C. Upon rinsing in 0.1 M
cacodylate buffer (pH 7.2; Wako Pure Chemicals Industries, Ltd.),
the tissues were post-fixed in 0.1% ruthenium red (Wako Pure
Chemicals Industries, Ltd.) and 2% osmium tetroxide solution (TAAB
Laboratories Equipment, Bershire, England) overnight at 4°C. Next,
the tissues were dehydrated in a graded ethanol series and embedded
in Epon 812 (Okenshoji Co., Ltd., Tokyo, Japan). Ultrathin sections
were prepared, stained with uranyl acetate (TAAB Laboratories
Equipment) and lead citrate (TAAB Laboratories Equipment), and
examined under a JEM-1230EX transmission electron microscope (JEOL,
Ltd., Tokyo, Japan).
Statistical analysis
Statistical analysis was performed using the
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Inhibition of HA synthesis in MIA
PaCa-2 cells by MU
The effect of MU on HA synthesis in the surrounding
ECM of MIA PaCa-2 cells was examined using a particle exclusion
assay. When the cells were incubated without MU, the ECM halo could
be observed (Fig. 1A), whereas no
halo could be observed when the cells were incubated with 0.1 or
0.5 mM MU (Fig. 1B and C,
respectively). Cells treated with Streptomyces
hyalurolyticus hyaluronidase prior to the assay also exhibited
no halo (Fig. 1D).
Immunohistochemical staining for HA indicated that MIA PaCa-2 cells
retained HA on their cell surface (Fig.
1E). However, HA staining by HABP could not be observed when
the cells were incubated with 0.5 mM MU (Fig. 1F), and cells treated with
Streptomyces hyalurolyticus hyaluronidase prior to HABP
treatment were not stained (Fig. 1G).
Quantitative analysis of cellular HA was also performed (Fig. 1H). MU caused a decline in HA retention
in a dose-dependent manner, and incubation with 1.0 mM MU caused a
29% decrease of HA, compared with the control (P<0.05).
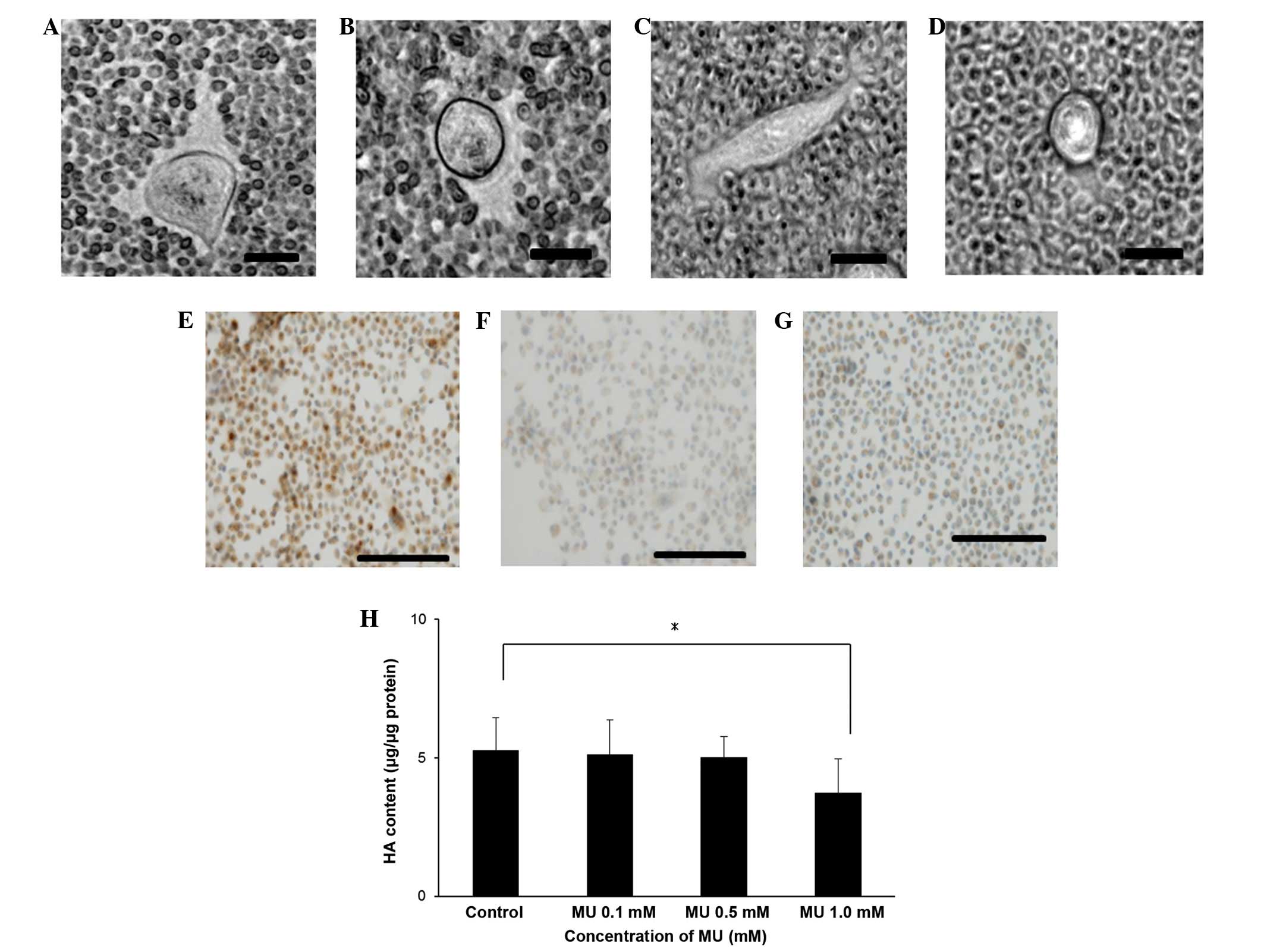 | Figure 1.(A-D) Visualization of the ECM. The
ECM surrounding MIA PaCa-2 cells was visualized using a particle
exclusion assay in (A) control cells, (B) cells incubated with 0.1
mM MU, (C) cells incubated with 0.5 mM MU and (D) cells incubated
with 3.0 U/ml Streptomyces hyalurolyticus hyaluronidase. Bar
size, 20 µm; magnification, ×400; staining, biotinylated HA binding
protein. Immunohistochemical staining of (E) control pancreatic
cancer cells, (F) cells incubated with 0.5 mM MU and (G) cells
incubated with 3.0 U/ml Streptomyces hyalurolyticus
hyaluronidase. Bar size, 100 µm; magnification, ×200; staining,
biotinylated HA binding protein. (H) Effect of MU on the synthesis
of HA by MIA PaCa-2 cells. Cells were incubated with 0.1, 0.5 or
1.0 mM MU for 48 h, prior to the extraction and quantification of
HA. Data are represented as the mean ± standard error of the mean
of eight replicates. *P<0.05. ECM, extracellular matrix; MU,
4-methylumbelliferone; HA, hyaluronan. |
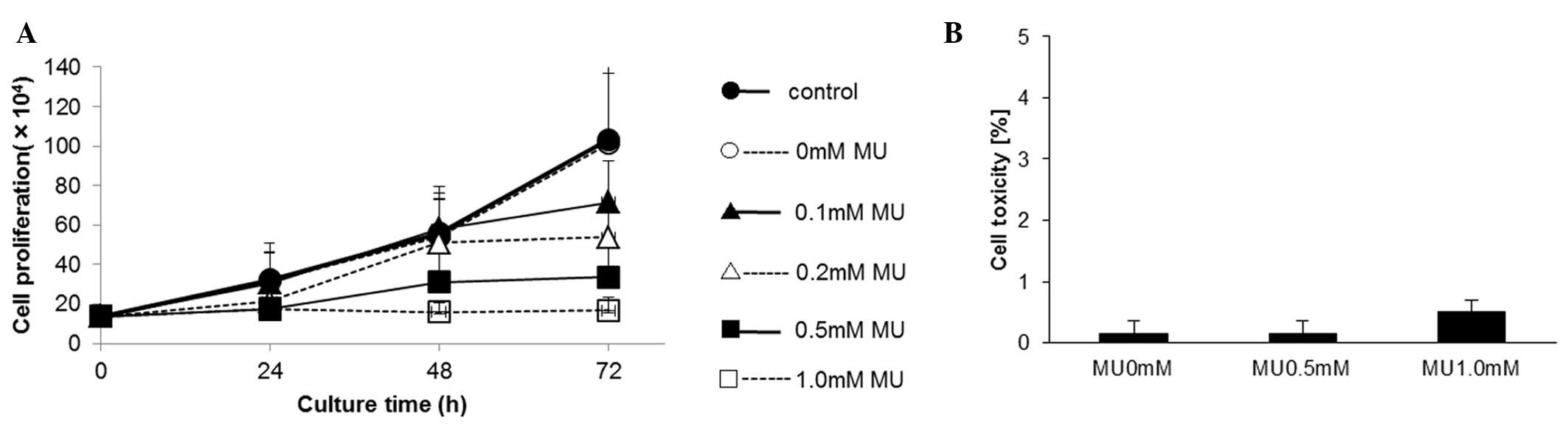
Effect of MU on the proliferation of
MIA PaCa-2 cells
The proliferation of MIA PaCa-2 cells was evaluated
using a cell counter, which revealed that MU inhibited cell
proliferation in a dose-dependent manner. Upon 72-h incubation,
cell proliferation was inhibited by 30.6% by 0.1 mM MU, 48.0% by
0.2 mM MU and 67.5% by 0.5 mM MU, compared with the control
(Fig. 2A). The cytotoxicity of MU was
determined using an LDH assay. Following MU administration, the LDH
activity did not increase in the culture medium (Fig. 2B).
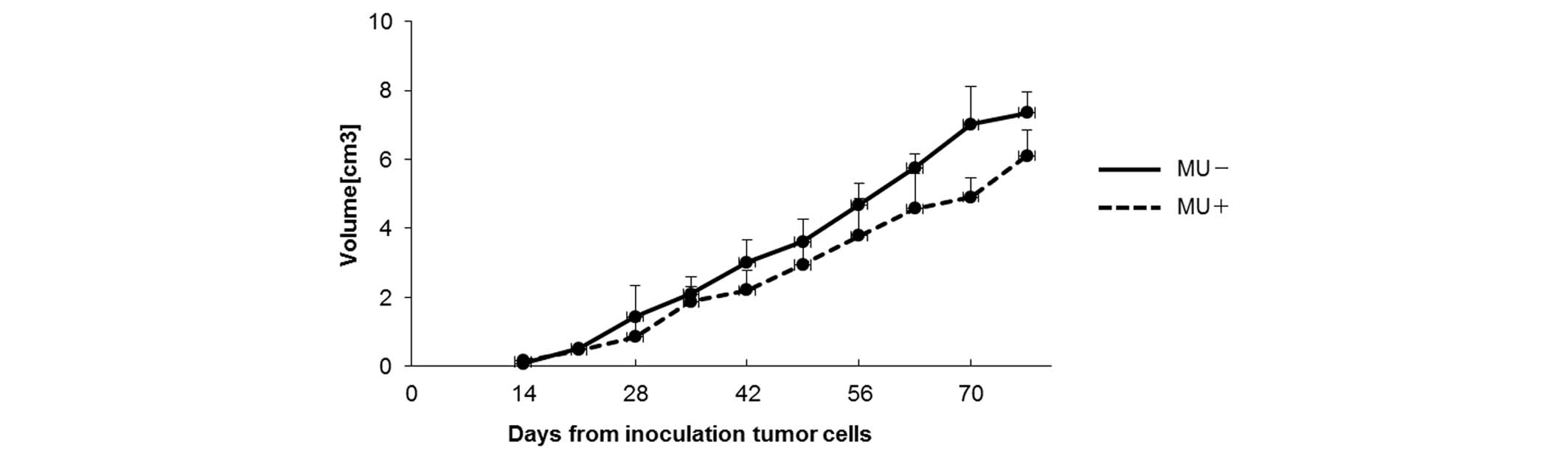
Effect of MU on human pancreatic
cancer in vivo
To examine the effect of MU in vivo,
suspensions containing 4×106 MIA PaCa-2 cells were
subcutaneously injected into mice, and MU was administered orally
every day. On day 70 post-inoculation, the tumor volume was 4.91
cm3 in the MU-treated group vs. 7.02 cm3 in
the control group (Fig. 3). The
majority of mice succumbed to cachexia, and no differences in body
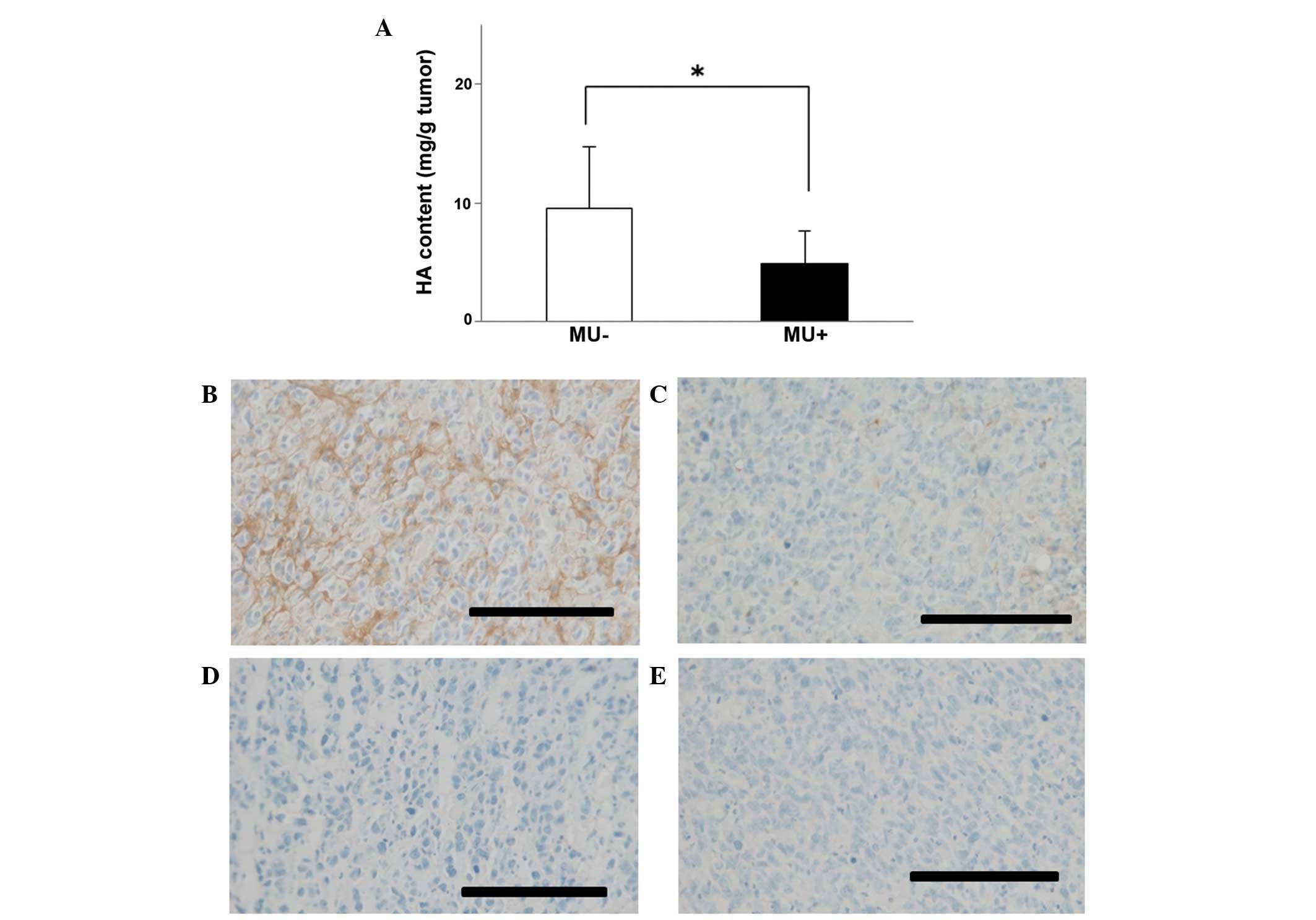
weight were observed between the treatment and control groups. HA
content in tumors was 4.89 mg/g in the MU-treated group vs. 9.59
mg/g in the control group, indicating that the quantity of HA in
the tumor was significantly (P<0.05) decreased by MU treatment
(Fig. 4A). Immunohistochemical
staining of the tumors demonstrated that HA was present around the
cancer cells, and in comparison with the untreated controls, the
levels of HA were reduced in mice that had been treated with MU
(Fig. 4B and C). Tissue sections that
had been treated with 100 U/ml Streptomyces hyalurolyticus
hyaluronidase prior to staining with HABP exhibited no HA (Fig. 4D and E).
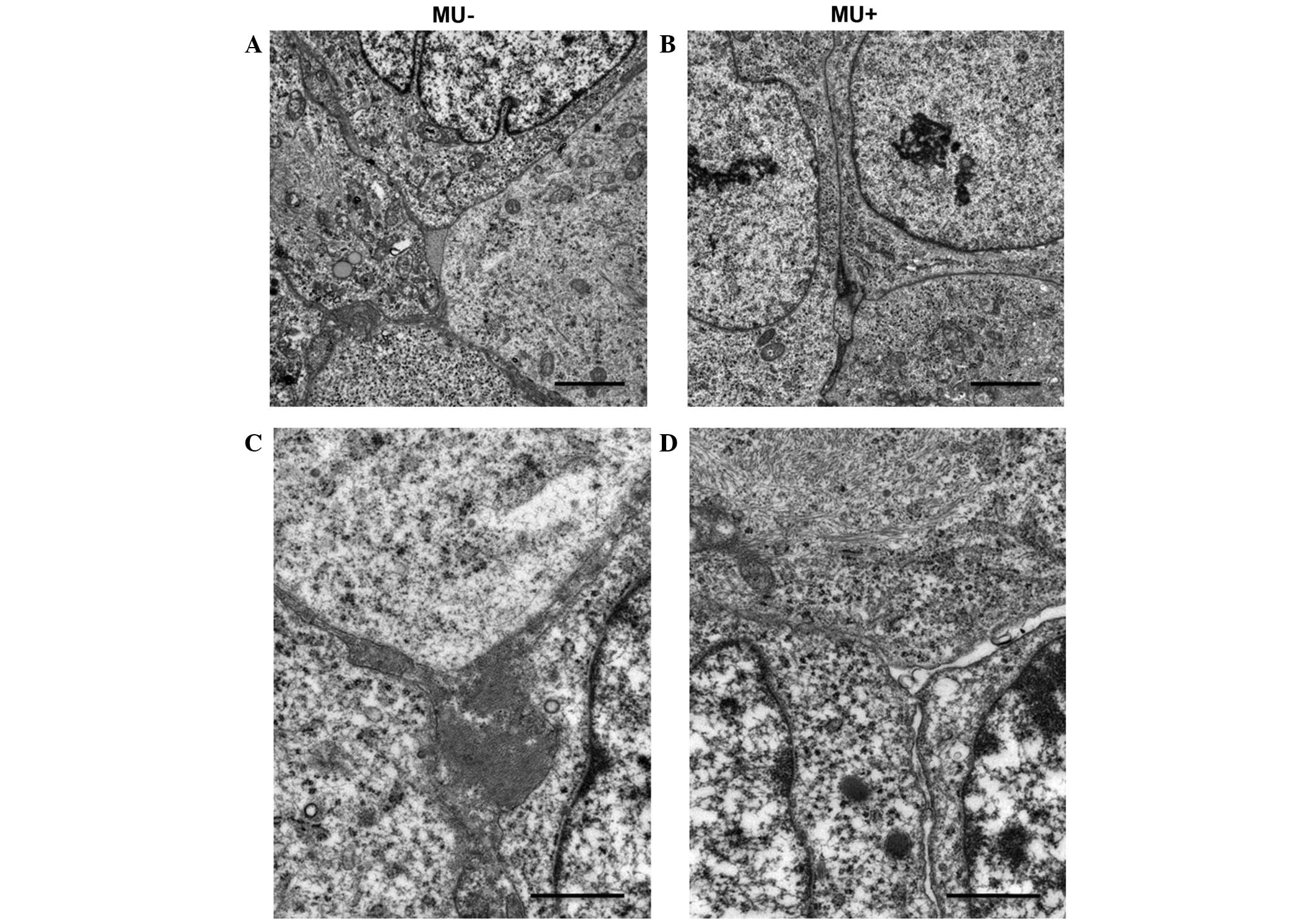
Effect of MU on the microenvironment
of the ECM of cancer cells
Electron microscopy was used to detect changes in
the microenvironment surrounding the inoculated MIA PaCa-2 cells.
Cell nuclei exhibited numerous mitotic features. MIA PaCa-2 cells
had increased tumor cellularity, displayed a strong, proliferative
capability and grew without stroma. In the MU-treated group,
necrosis was suppressed, and the number of apoptotic bodies was not
increased, compared with the control group. No disruption of the
nuclei or differences in the morphology of the nuclei and
cytoplasmic organelles were observed between the MU-treated and the
control groups (Fig. 5A and B). By
contrast, the intercellular space was less cohesive in the
MU-treated group, compared with the control group. In addition, the
MU-treated group exhibited interstitial oligochromia (Fig. 5C and D).
Discussion
HA is intimately involved in the biology of cancer
(7). HA accumulates into the stroma
of various human tumors and modulates the intracellular signaling
pathways involved in cell proliferation and migration (6,7).
Metastatic tumors are enriched in HA, and the healthy fibroblasts
that surround malignant cells are stimulated to synthesize HA
(35). HA-rich environments enhance
tumor invasion and metastasis, and are associated with poorly
differentiated phenotypes and poor prognosis in human
adenocarcinoma (36). Delivery of
macromolecules to cancer cells is inhibited by the ECM, since HA
supplies a high intratumoral fluidic pressure, which prevents the
diffusion and penetration of anticancer agents into the tumor
tissue (37). This suggests that
cancer cells are dependent on HA-rich environments for survival,
self-replication, migration to other tissues and drug resistance.
Therefore, regulation of HA metabolism is important for the
treatment of cancer. Several studies have reported that MU inhibits
the synthesis of HA and the preservation of HA in the ECM (13–15). As a
result, MU has been widely investigated as an inhibitor of HA
synthesis, and has been proposed as an anticancer agent (15–19). The
anticancer effect of MU has been reported in various types of
cancer, including malignant melanoma (15,16),
hepatocellular carcinoma (17),
prostate cancer (18) and breast
cancer (19). However, the effect of
MU on pancreatic cancer remains unknown. Pancreatic cancer has the
worst prognosis of all solid tumors (20). Conventional treatments such as
chemotherapy, molecular-targeted therapy, radiotherapy and
combinations of all three hardly improve prognosis (20,21).
Accordingly, the present authors hypothesized that MU may be useful
for the treatment of pancreatic cancer.
In the present study, MIA PaCa-2 cells were observed
to retain HA on the cellular surface when they were not treated
with MU; however, when MU was administered into the culture medium,
this was suppressed, as evidenced by biochemical and
immunohistochemical assays. Furthermore, MU inhibited cell
proliferation in a dose-dependent manner without causing any damage
to cell membranes. These results suggest that MU is able to inhibit
the growth of pancreatic tumors by suppressing HA synthesis.
Correspondingly, the volume of tumors formed in mice upon
inoculation of MIA PaCa-2 cells was reduced following oral
administration of MU, and a reduction on the levels of HA present
in tumor tissue was confirmed by quantitative analysis.
Immunohistochemical staining revealed that HA was abundant in the
ECM surrounding the pancreatic cancer cells of the inoculated mice
when the animals were not exposed to MU. By contrast, when treated
with MU, a decrease in HA retention in the ECM was observed.
Electron microscopy was used to examine the structural changes in
the ECM, and the gap between the cells was observed to be weaker in
the MU-treated group than in the untreated group. Structural
changes in the ECM caused by MU may aid anticancer agents and
immunocompetent lymphocytes to diffuse through the intercellular
space and successfully reach the cancer cells. The remodeling of
the ECM is therefore expected to act as an adjuvant
chemosensitizer, thus enabling the treatment of otherwise
intractable tumors. As a result, MU is a potentially useful
anticancer therapeutic agent for pancreatic cancer.
An alternative method for controlling the cellular
levels of HA is through genetic manipulation. However, HA
deficiency leads to developmental problems. Camenisch et al
(38) generated HAS2 knockout mice,
and reported that targeted depletion of the HAS2 gene resulted in
embryonic mortality due to extensive abnormalities in the
development of the cardiovascular system. It has also been reported
that HA is involved in various physiological processes that are
essential for maintenance of life (3), and therefore, precise control over the
extent of HA reduction is required for the successful clinical
application of HA modulation as an anticancer treatment.
In the present study, MU treatment mediated an
decrease in tumor volume, reduced the quantity of intratumoral HA
and did not show any harmful effect on the animals. This is
supported by the fact that MU has been used as a choleretic and
antispasmodic agent for patients with motor disorders of the
duodenal papilla (39,40), and it is one of the active ingredients
in coumarin that has been habitually consumed for a long time,
particularly in Europe (41).
Suitability for oral administration, low toxicity and low cost are
additional advantages of MU (42).
The use of hyaluronidase for the treatment of cancer
has been demonstrated in several studies (43,44).
However, hyaluronidase is produced from bovine testis, and its
clinical use has been discontinued due to allergic reactions, since
the enhancement of hyaluronidase activity in healthy tissues has
been observed to cause inflammation or pain in joints (43,44).
Targeting cluster of differentiation (CD)44 has been suggested as
an alternative candidate to targeting HA (45). CD44 has attracted attention as a stem
cell marker of pancreatic cancer, along with CD24 and
epithelial-specific antigen (46).
CD44 is able to bind to HA in the ECM and enhance cancer cell
proliferation, invasion and metastasis (45). Pancreatic cancer cells expressing CD44
demonstrated the characteristic features of cancer stem cells,
including upregulation of signaling pathways, self-renewal and
enhanced tumorigenesis (46). These
properties enable cancer cells to develop tolerance against
chemotherapeutic agents and radiation (46). In a previous study, an anti-CD44
antibody demonstrated anticancer efficacy by controlling HA
bioactivity. However, this antibody is expensive, and similar to
other molecular targeting agents, it may not be used in a clinical
setting for a long time.
In conclusion, in the present study, MU demonstrated
an antitumor effect without exerting any toxic effects in cells. MU
was able to inhibit HA synthesis in cancer cells, thus making the
ECM more appropriate for drug delivery. Further studies are
required on the side effects of MU in vivo and the
synergistic effect of combining MU with other anticancer agents.
The use of MU may contribute to more effective pancreatic cancer
treatments due to its chemosensitizer properties.
Acknowledgements
The authors would like to thank Professor Hiroshi
Kijima and Dr Satoko Morohashi (Department of Pathology and
Bioscience, Hirosaki University, Graduate School of Medicine,
Hirosaki, Japan) for their excellent technical help, as well as
Professor Makoto Hayakari (Department of Pharmacy, Hirosaki
University Hospital, Hirosaki, Japan) for kindly providing the
cells used in the present study.
Glossary
Abbreviations
Abbreviations:
|
HA
|
hyaluronan
|
|
HAS
|
hyaluronan synthase
|
|
MU
|
4-methylumbelliferone
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DMSO
|
dimethyl sulfoxide
|
|
PBS
|
phosphate-buffered saline
|
|
ECM
|
extracellular matrix
|
References
|
1
|
Weissmann B and Meyer K: The structure of
hyalobiuronic acid and of hyaluronic acid from umbilical cord. J Am
Chem Soc. 76:1753–1757. 1954. View Article : Google Scholar
|
|
2
|
Weigel PH, Hascall VC and Tammi M:
Hyaluronan synthases. J Biol Chem. 272:13997–14000. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Menzel EJ and Farr C: Hyaluronidase and
its substrate hyaluronan: Biochemistry, biological activities and
therapeutic uses. Cancer Lett. 131:3–11. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Auvinen P, Tammi R, Parkkinen J, Tammi M,
Agren U, Johansson R, Hirvikoski P, Eskelinen M and Kosma VM:
Hyaluronan in peritumoral stroma and malignant cells associates
with breast cancer spreading and predicts survival. Am J Pathol.
156:529–536. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang L, Underhill CB and Chen L:
Hyaluronan on the surface of tumor cells is correlated with
metastatic behavior. Cancer Res. 55:428–433. 1995.PubMed/NCBI
|
|
6
|
Simpson MA, Reiland J, Burger SR, Furcht
LT, Spicer AP, Oegema TR Jr and McCarthy JB: Hyaluronan synthase
elevation in metastatic prostate carcinoma cells correlates with
hyaluronan surface retention, a prerequisite for rapid adhesion to
bone marrow endothelial cells. J Biol Chem. 276:17949–17957. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Itano N, Atsumi F, Sawai T, Yamada Y,
Miyaishi O, Senga T, Hamaguchi M and Kimata K: Abnormal
accumulation of hyaluronan matrix diminishes contact inhibition of
cell growth and promotes cell migration. Proc Natl Acad Sci USA.
99:3609–3614. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Anttila MA, Tammi RH, Tammi MI, Syrjänen
KJ, Saarikoski SV and Kosma VM: High levels of stromal hyaluronan
predict poor disease outcome in epithelial ovarian cancer. Cancer
Res. 60:150–155. 2000.PubMed/NCBI
|
|
9
|
Lokeshwar VB, Rubinowicz D, Schroeder GL,
Forgacs E, Minna JD, Block NL, Nadji M and Lokeshwar BL: Stromal
and epithelial expression of tumor markers hyaluronic acid and
HYAL1 hyaluronidase in prostate cancer. J Biol Chem.
276:11922–11932. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Setälä LP, Tammi MI, Tammi RH, Eskelinen
MJ, Lipponen PK, Agren UM, Parkkinen J, Alhava EM and Kosma VM:
Hyaluronan expression in gastric cancer cells is associated with
local and nodal spread and reduced survival rate. Br J Cancer.
79:1133–1138. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ropponen K, Tammi M, Parkkinen J,
Eskelinen M, Tammi R, Lipponen P, Agren U, Alhava E and Kosma VM:
Tumor cell-associated hyaluronan as an unfavorable prognostic
factor in colorectal cancer. Cancer Res. 58:342–347.
1998.PubMed/NCBI
|
|
12
|
Collier AC, Tingle MD, Keelan JA, Paxton
JW and Mitchell MD: A highly sensitive fluorescent microplate
method for the determination of UDP-glucuronosyl transferase
activity in tissues and placental cell lines. Drug Metab Dispos.
28:1184–1186. 2000.PubMed/NCBI
|
|
13
|
Nakamura T, Takagaki K, Shibata S, Tanaka
K, Higuchi T and Endo M: Hyaluronic-acid-deficient extracellular
matrix induced by addition of 4-methylumbelliferone to the medium
of cultured human skin fibroblasts. Biochem Biophys Res Commun.
208:470–475. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kakizaki I, Kojima K, Takagaki K, Endo M,
Kannagi R, Ito M, Maruo Y, Sato H, Yasuda T, Mita S, et al: A novel
mechanism for the inhibition of hyaluronan biosynthesis by
4-methylumbelliferone. J Biol Chem. 279:33281–33289. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kudo D, Kon A, Yoshihara S, Kakizaki I,
Sasaki M, Endo M and Takagaki K: Effect of a hyaluronan synthase
suppressor, 4-methylumbelliferone, on B16F-10 melanoma cell
adhesion and locomotion. Biochem Biophys Res Commun. 321:783–787.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoshihara S, Kon A, Kudo D, Nakazawa H,
Kakizaki I, Sasaki M, Endo M and Takagaki K: A hyaluronan synthase
suppressor, 4-methylumbelliferone, inhibits liver metastasis of
melanoma cells. FEBS Lett. 579:2722–2726. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Piccioni F, Malvicini M, Garcia MG,
Rodriguez A, Atorrasagasti C, Kippes N, Piedra Buena IT, Rizzo MM,
Bayo J, Aquino J, et al: Antitumor effects of hyaluronic acid
inhibitor 4-methylumbelliferone in an orthotopic hepatocellular
carcinoma model in mice. Glycobiology. 22:400–410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lokeshwar VB, Lopez LE, Munoz D, Chi A,
Shirodkar SP, Lokeshwar SD, Escudero DO, Dhir N and Altman N:
Antitumor activity of hyaluronic acid synthesis inhibitor
4-methylumbelliferone in prostate cancer cells. Cancer Res.
70:2613–2623. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pályi-Krekk Z, Barok M, Isola J, Tammi M,
Szöllosi J and Nagy P: Hyaluronan-induced masking of ErbB2 and
CD44-enhanced trastuzumab internalisation in trastuzumab resistant
breast cancer. Eur J Cancer. 43:2423–2433. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Raimondi S, Maisonneuve P and Lowenfels
AB: Epidemiology of pancreatic cancer: An overview. Nat Rev
Gastroenterol Hepatol. 6:699–708. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Berrino F, De Angelis R, Sant M, Rosso S,
Bielska-Lasota M, Coebergh JW and Santaquilani M: EUROCARE Working
group: Survival for eight major cancers and all cancers combined
for European adults diagnosed in 1995-99: Results of the EUROCARE-4
study. Lancet Oncol. 8:8682007. View Article : Google Scholar
|
|
23
|
Cheng XB, Sato N, Kohi S and Yamaguchi K:
Prognostic impact of hyaluronan and its regulators in pancreatic
ductal adenocarcinoma. PLoS One. 8:e807652013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakazawa H, Yoshihara S, Kudo D, Morohashi
H, Kakizaki I, Kon A, Takagaki K and Sasaki M:
4-methylumbelliferone, a hyaluronan synthase suppressor, enhances
the anticancer activity of gemcitabine in human pancreatic cancer
cells. Cancer Chemother Pharmacol. 57:165–170. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Knudson CB and Toole BP: Changes in the
pericellular matrix during differentiation of limb bud mesoderm.
Dev Biol. 112:308–318. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin W, Shuster S, Maibach HI and Stern R:
Patterns of hyaluronan staining are modified by fixation
techniques. J Histochem Cytochem. 45:1157–1163. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chichibu K, Matsuura T, Shichijo S and
Yokoyama MM: Assay of serum hyaluronic acid in clinical
application. Clin Chim Acta. 181:317–323. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barbosa H, Slater NK and Marcos JC:
Protein quantification in the presence of poly(ethylene glycol) and
dextran using the Bradford method. Anal Biochem. 395:108–110. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nojiri S and Joh T: Albumin suppresses
human hepatocellular carcinoma proliferation and the cell cycle.
Int J Mol Sci. 15:5163–5174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Decker T and Lohmann-Matthes ML: A quick
and simple method for the quantitation of lactate dehydrogenase
release in measurements of cellular cytotoxicity and tumor necrosis
factor (TNF) activity. J Immunol Methods. 115:61–69. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Andarini S, Kikuchi T, Nukiwa M, Pradono
P, Suzuki T, Ohkouchi S, Inoue A, Maemondo M, Ishii N, Saijo Y, et
al: Adenovirus vector-mediated in vivo gene transfer of OX40 ligand
to tumor cells enhances antitumor immunity of tumor-bearing hosts.
Cancer Res. 64:3281–3287. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stern M and Stern R: An ELISA-like assay
for hyaluronidase and hyaluronidase inhibitors. Matrix. 12:397–403.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Luna LG: Manual of Histologic Staining
Methods of the Armed Forces Institute of Pathology. 3rd edition.
Blakiston Division, McGraw-Hill; New York: 1968
|
|
34
|
Meyer LJ and Stern R: Age-dependent
changes of hyaluronan in human skin. J Invest Dermatol.
102:385–389. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Knudson W, Biswas C and Toole BP:
Interactions between human tumor cells and fibroblasts stimulate
hyaluronate synthesis. Proc Natl Acad Sci USA. 81:6767–6771. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sironen RK, Tammi M, Tammi R, Auvinen PK,
Anttila M and Kosma VM: Hyaluronan in human malignancies. Exp Cell
Res. 317:383–391. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Buckway B, Wang Y, Ray A and Ghandehari H:
Overcoming the stromal barrier for targeted delivery of HPMA
copolymers to pancreatic tumors. Int J Pharm. 456:202–211. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Camenisch TD, Spicer AP, BrehmGibson T,
Biesterfeldt J, Augustine ML, Calabro A Jr, Kubalak S, Klewer SE
and McDonald JA: Disruption of hyaluronan synthase-2 abrogates
normal cardiac morphogenesis and hyaluronan-mediated transformation
of epithelium to mesenchyme. J Clin Invest. 106:349–360. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garrett ER, Venitz J, Eberst K and Cerda
JJ: Pharmacokinetics and bioavailabilities of hymecromone in human
volunteers. Biopharm Drug Dispos. 14:13–39. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fontaine L, Grand M, Molho D, Chabert MJ
and Boschetti E: Choleretic, spasmolytic and general pharmacologic
activities of 4-methylumbelliferone. Therapie. 23:51–62. 1968.(In
French). PubMed/NCBI
|
|
41
|
Hoult JR and Payá M: Pharmacological and
biochemical actions of simple coumarins: Natural products with
therapeutic potential. Gen Pharmacol. 27:713–722. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Karbownik MS and Nowak JZ: Hyaluronan:
Towards novel anti-cancer therapeutics. Pharmacol Rep.
65:1056–1074. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baumgartner G, Gomar-Höss C, Sakr L,
Ulsperger E and Wogritsch C: The impact of extracellular matrix on
the chemoresistance of solid tumors - experimental and clinical
results of hyaluronidase as additive to cytostatic chemotherapy.
Cancer Lett. 131:85–99. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Whatcott CJ, Han H, Posner RG, Hostetter G
and Von Hoff DD: Targeting the tumor microenvironment in cancer:
Why hyaluronidase deserves a second look. Cancer Discov. 1:291–296.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Takada M, Yamamoto M and Saitoh Y: The
significance of CD44 in human pancreatic cancer: II. The role of
CD44 in human pancreatic adenocarcinoma invasion. Pancreas.
9:753–757. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tang SC and Chen YC: Novel therapeutic
targets for pancreatic cancer. World J Gastroenterol.
20:10825–10844. 2014. View Article : Google Scholar : PubMed/NCBI
|