Introduction
Diffuse large B-cell lymphoma (DLBCL), the most
common type of non-Hodgkin's lymphoma (NHL) in adults, accounts for
approximately 30–40% of newly diagnosed lymphomas worldwide
(1). Using gene expression profiling,
DLBCL can be divided into three distinguishable subtypes; the
germinal center B-cell like (GBC), activated B-cell like (ABC) and
primary mediastinal B-cell lymphoma subtypes (1–3). These
subtypes differ in the expression of thousands of genes and arise
at separate stages of B-cell differentiation. The ABC subgroup has
a poor prognosis with a short survival time compared with the GCB
subgroup (2–4). In addition, the frequency of the ABC
subtype in Asian countries, such as Japan and China, is increased
compared with the GCB subtype. The ABC subtype is also more common
in certain Asian countries compared with in the western world
(5,6).
Environmental factors, including viruses and
bacteria, may contribute to the development of cancer, and have
been previously indicated in cervical cancer, hepatocellular
carcinoma, gastric cancer and lymphoproliferative disorders
(7–10). Viruses may be involved in the
development of cancer by activating the innate immune system, as
well as activating intracellular signaling cascades that control
viral infection and tumor cell growth (11). Bacteria, viruses and parasites may
also directly contribute to carcinogenesis through the transfection
and integration of oncogenes, which results in malignant cell
transformation (12,13). Viruses involved in carcinogenesis may
be targets for diagnosis, prevention and therapies (11).
Although the incidence of NHL has increased during
the last three decades, the etiology of the most common types
remains unclear (American Cancer Society, 2001). Chromosomal
translocations are common in numerous NHL subtypes, which could
result in the inactivation of tumor suppressor genes or activation
of oncogenes (14). Viruses may
induce genetic damage to the genome of lymphoid cells and
dysregulate normal cell growth (15).
Certain viruses have been specifically linked to particular types
of lymphomas, including Epstein-Barr virus (EBV), human
T-lymphotropic virus type 1, herpes virus-8 and hepatitis (16–18).
Prevention of viral infections should reduce the number of
individuals at risk of lymphoma development; thus recognizing the
viruses involved in the tumorigenesis of NHL is important.
The virus genome has been shaped to interact with
host cell regulatory and signaling networks. Viruses may alter host
cell proteins in all steps of the viral cycle and utilize common
host cell response pathways for pathogenic invasion (19,20). The
Janus kinase (JAK)/signal transducers and activators of
transcription (STAT) pathway may be activated by viral infections
that result in maintained cell growth (21–23).
Replicating hepatitis B virus (HBV) modulates STAT signaling by the
upregulation of STAT3, which contributes to immune suppression,
viral persistence and malignant transformation (21). As a part of the JAK/STAT pathway,
STAT3 has been shown to be upregulated in several types of
malignancies, including B-cell lymphomas (24). STAT3 has been previously suggested as
a therapeutic target of ABC DLBCL, since STAT3 and nuclear factor κ
B activation may be associated with a poor survival in this
subgroup of patients (24).
A previous study analyzed the gene and micro RNA
(miRNA) expression patterns in the lymphoma tissues of patients
from two geographically varied regions with differing loads of
microbial infections, and found a difference in the expression of
STAT3 (25). The patient populations
were from Sweden (a low infectious disease region) and Egypt (a
high endemic infectious disease region) (25). STAT3 was overexpressed in Swedish
patients compared with Egyptian patients (26) and miR-1234 was shown to be a possible
regulator of STAT3 expression (27).
The main aim of the present study was to analyze the
presence of microbial agents in the lymphoma tissues of patients
with ABC DLBCL, and to associate this with STAT3 and miR-1234
expression.
Materials and methods
Patients and controls
In total, 47 ABC DLBCL patient biopsy samples were
obtained from the National Cancer Institute, Cairo, Egypt, between
July 2003 and January 2008, and 14 ABC DLBCL patient biopsy samples
were obtained from the Karolinska University Hospital, Stockholm,
Sweden, between October 2005 and September 2011. The biopsy
material was obtained at the time of diagnosis and embedded in
paraffin. The median ages of the Swedish and Egyptian patients were
57 and 66 years, respectively. The gender distribution (%
male/female) was 53/47 and 43/57 in Swedish and Egyptian patients,
respectively. Reactive lymph nodes from teh same patients were
collected as controls, from the National Cancer Institute, Cairo,
Egypt (n=10) and from the Karolinska University Hospital,
Stockholm, Sweden (n=10). The study was approved by the Regional
Ethics Committee (www.epn.se) and written informed
consent was obtained from all patients.
Histopathological diagnosis
Primary selection of DLBCL patients was performed
using formalin-fixed (FF) paraffin-embedded (PE) material stained
with eosin and hematoxylin and automated immunohistochemistry (IHC)
for cluster of differentiation (CD)20, CD3, Ki67, B-cell lymphoma
(Bcl)-2, Bcl-6, CD10 and melanoma associated antigen (mutated) 1,
according to the algorithm described by Hans et al (28). This algorithm has recently been
confirmed as one of two IHC algorithms that can predict the
non-germinal center cell origin with a high accuracy, and has 86%
concordance with gene expression profiling results (29). FFPE blocks classified as the non-GCB
DLBCL subtype, as described previously (30), were selected for the study and were
analyzed by the Department of Pathology.
Handling of patient and control
material
The tissue blocks were cut into 20-µm thick
sections, using sterile blades. All benches, instruments and
pipettes were cleaned with RNaseZap solution (Ambion; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). A previous study reported that
FFPE materials may be used for gene expression analysis, and are
comparable with fresh frozen material (30).
RNA extraction
Extraction of RNA from PE material with the Ambion
Recover All Total Nucleic Acid Isolation optimized for FFPE samples
(Thermo Fisher Scientific, Inc.) was successfully performed
following the manufacturer's instructions. RNA samples were stored
at −80°C until required for amplification. In brief, 20-µm thick
sections were treated with xylene, centrifuged at full speed
(10,000 × g) and washed with 100% ethanol twice. The pellets were
air dried for 15 min. Digestion buffer and protease were added,
followed by 3 h incubation at 50°C. Isolation additive was added
followed by 100% ethanol, and the samples were transferred to a
filter cartridge. Two rounds of centrifugation with Wash 1 and Wash
2/3 was performed. DNase combined with 10X DNase buffer was added
followed by 30 min incubation at room temperature. The membranes
were then washed with Wash 1 and Wash 2/3. The RNA was eluted in 30
µl RNase free water preheated at 95°C for 30 min. Yield and quality
was measured using the Nanodrop Technologies ND 1000 (Thermo Fisher
Scientific, Inc.). Absorbance ratios (260/280 for DNA and 260/230
for RNA) of >1.8 were considered to be of high purity.
Complementary DNA (cDNA) preparation
and amplification from PE DLBCL material
RNA (100 ng) was used for amplification according to
the protocols of the WT-Ovation FFPE RNA Amplification System V2
(Nugen Technologies, Inc., San Carlos, CA, USA), with a Spike
control. All incubations were performed using the 1294 Techne
Progene Peltier Thermal cycler thermal cycler (Scientific support,
Inc., Hayward, CA, USA). The purification step was performed using
Agencourt RNA Clean purification beads (Beckman Coulter, Inc.,
Fullerton, CA, USA). The amplified cDNA was purified using DNA
Clean & Concentrator kit (Zymo Research Corporation, Irvine,
CA, USA). The yield and quality of the cDNA was measured using the
Nanodrop Technologies ND 1000. cDNA to be used in the microarray
analysis were pooled using the same concentration of each
sample.
Microbial detection array (MDA)
Nucleic acid concentrations were determined using
the Invitrogen Qubit Fluorometer (Thermo Fisher Scientific, Inc.).
DNA and RNA were randomly amplified using a protocol described
previously (31). Following
amplification, the amplified samples were purified using the Qiagen
QIAquick PCR Purification kit (Qiagen GmbH, Hilden, Germany).
Amplified nucleic acids were fluorescently labeled using the Roche
NimbleGen One-Color DNA Labeling kit (Roche Applied Science,
Madison, WI, USA), according to the recommended protocols. DNA was
purified subsequent to labeling, and hybridized using the NimbleGen
Hybridization kit (Roche Applied Science) and the Lawrence
Livermore Microbial Detection Array (LLMDA), according to
manufacturers' instructions. Microarrays were allowed to hybridize
for 17 h and washed using the NimbleGen Wash Buffer kit (Roche
Applied Science), according to manufacturer's instructions. The
samples were then scanned on an Axon GenePix 4000B 5 µM scanner
from Molecular Devices, LLC (Sunnyvale, CA, USA). Signals on the
LLMDA were analyzed using the maximum likelihood analysis method
(32). In brief, probes were
identified when binding intensity to the probes exceeded a
threshold equal to the 99th percentile of intensities for the
negative control probes. Targets in an internal database of viral
sequences were screened against stringency criteria and a log-odd
score was computed for each target. A selection algorithm was
applied to find the collection of targets most likely to be present
in the sample. At every forward selection step, a conditional
log-odd score was computed for each remaining target. The target
with the largest conditional log-odd score was selected and added
to the collection. The conditional log-odd score is a value
representing the probability of being the detected target. Higher
log-odd scores have an increased probability of being the detected
target compared with lower log-odd scores.
HBV antigen and antibody analyses of
patient serum
DNA extracted from the serum of Swedish and Egyptian
patients was used to analyze the presence of the HBVsAg by
polymerase chain reaction (PCR), by detecting HBV core genes as
previously described (33). Briefly,
100 µl of reaction mixture containing 10 µl of extracted DNA, 50 mM
potassium chloride, 10 mM Tris-hydrochloric acid (pH 8.3), 2 mM
magnesium chloride, 200 µM deoxyribonucleosides, 2.5 units Taq
polymerase (PerkinElmer, Inc., Waltham, MA, USA), and 20 pmol of
each of the oligonucleotide primers [C1 sense, CTGGGAGGAGTTGGGGGA
(1730–1747) and C2 antisense, GTAGAAGAATAAAGCCC (2503–2487)] for
the core genes. Amplification was performed for one cycle of 95°C
for 5 min, followed by 35 cycles that consisted of denaturating for
1 min at 94°C, annealing for 1 min at 55°C and extension for 1.5
min at 72°C. Antibodies against HBV were analyzed in Swedish and
Egyptian patient serum using a chemiluminescent microparticle
immunoassay (Abbott Diagnostics, Lake Forest, IL, USA). In brief,
serum sample was mixed with paramagnetic microparticles coated with
recombinant HBsAg (rHBsAg; Abbott Diagnostics). Subsequent to
washing, acridium-marked rHBsAg-conjugate (Abbott Diagnostics) was
added. Following additional washing, pre-trigger and trigger
solution were added. The chemiluminescence reaction was then
measured and the concentration of anti-HBs was set by a
pre-designed calibration curve (Abbott Diagnostics).
Relative expression of STAT3 in
HBVsAg-positive (HBV+) and -negative (HBV-) Egyptian patients
To evaluate the relative expression of STAT3 and
micro RNA (miR)-1234 in association with HBV, HBV+ and HBV−
Egyptian and Swedish patients were analyzed individually using
Applied Biosystems HT7900 quantitative PCR (Thermo Fisher
Scientific, Inc.). For the STAT3 relative expression analysis, the
High Capacity RNA to cDNA kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was used for generating cDNA, according to
manufacturer's recommendations. cDNA was further amplified (250 ng)
by applying the TaqMan PreAmp Master Mix kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.), according to manufacturer's
recommendations. The TaqMan assays designed for STAT3 and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were used (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The relative
expression of individual patient samples was calculated using the
reference gene GAPDH. For the miR-1234 relative expression
analysis, total RNA was poly-adenylated using yeast polyA
polymerase (Affymetrix, Inc., Santa Clara, CA, USA) for 1 h at
37°C, followed by 10 min at 65°C, according to manufacturer's
recommendations. The RNA was precipitated over night using sodium
acetate-isopropanol at −20°C, washed in ethanol and diluted in
H2O. The reverse transcription reaction was performed
using the TaqMan miRNA reverse transcription kit (Thermo Fisher
Scientific, Inc.) using 1.17 µM universal RT primer. The reaction
was incubated at 72°C for 5 min and put on ice prior to adding the
RT enzyme. The reaction was then incubated at 42°C for 45 min,
followed by 5 min at 85°C. qPCR was performed using an Applied
Biosystems HT7900 qPCR machine (Thermo Fisher Scientific, Inc.)
with miR-1234 primers and TaqMan universal PCR master mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The relative
expression of miR-1234 was calculated relative to the expression of
5S rRNA. The mean Cq values and the relative expression for STAT3
and miR-1234 expression were calculated, as previously described
(34).
Statistics
Statistical analysis was performed using GraphPad
Prism version 5 (GraphPad Software, Inc., La Jolla, CA, USA).
Statistical calculation was performed by a one-way analysis of
variance, followed by post-hoc Tukey's honest significant
difference test or Student's t-test. P<0.05 was used to indicate
a statistically significant difference.
Results
Microbial detection in Swedish and
Egyptian patients using MDA analysis
Viruses were detected by the MDA, a high-density
oligonucleotide array for the detection and discovery of viruses
and bacteria. Viruses that were found in all groups (controls and
patients) were excluded, leaving exclusively those found in each
patient group. Among excluded viruses were influenza virus A and
human endogenous retroviruses found in the patient and control
groups. Human immunodeficiency virus (HIV) was excluded as it was
found in Swedish patients and controls. Clinically irrelevant
viruses, including plant and animal viruses that have not been
reported to infect human cells (for example faba bean necrotic
yellow virus, porcine circovirus and melon necrotic spot virus)
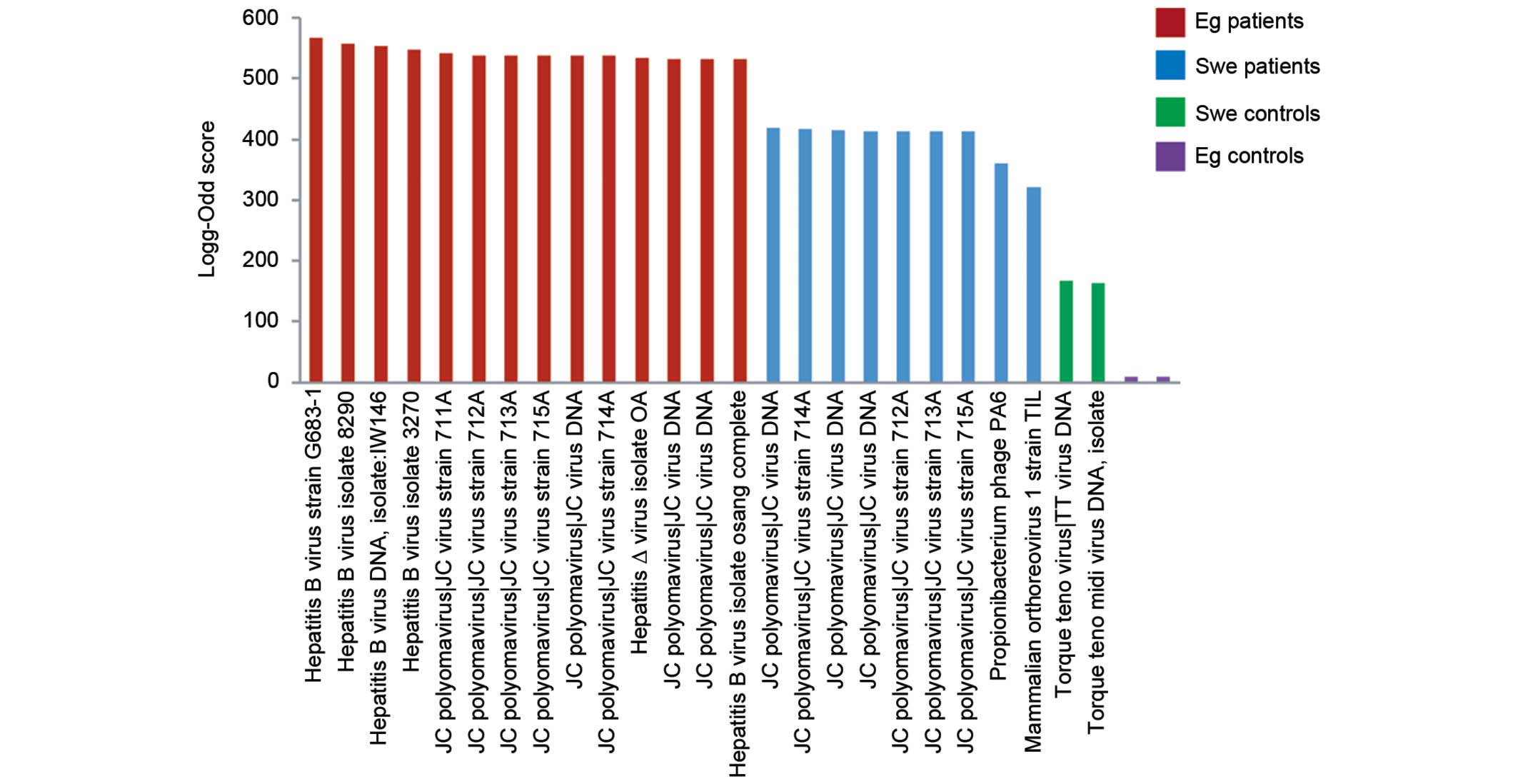
were also excluded. The log-odd scores were plotted in a bar graph
(Fig. 1). The HBV had the highest
log-odd scores, including the complete genome, and was only found
in Egyptian patients. JC polyomavirus (JCV) was found in Egyptian
and Swedish patients, with a higher log-odd score in the Egyptian
patients, but not in Swedish and Egyptian controls. Generally, the
highest log-odd scores were found in Egyptian patients rather than
in Swedish patients. Few viruses were found in the control groups.
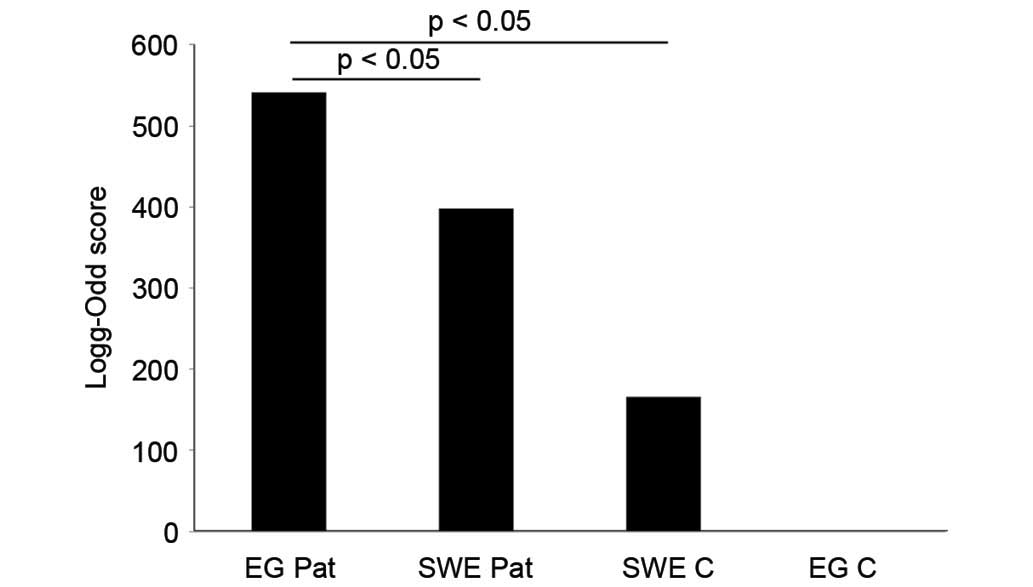
To visualize the differences in log-odd scores between the groups,
a mean log-odd score was calculated for each group. A statistically
significant difference was found between the mean log-odd scores
for each patient and control group (P=0.032 for Swedish controls
versus Egyptian patients; P=0.041 for Swedish patients versus
Egyptian patients; Fig. 2).
Serum HBVsAgs and antibody
responses
HBV showed the highest log-odd score and the
presence of the complete genome, but only in Egyptian patients
using lymphoma tissues. HBVsAgs could be detected in the serum of
38% of Egyptian patients, but not in any of the Swedish patients.
According to previous literature, the prevalence in the general
population of Egypt for the HBsAg is 4% (35), and in Swedish patients <1%
(36). The difference, with regard to
the presence of HBVsAg, between the Egyptian patients and the
general Egyptian population was statistically significant
(P<0.05) (35). HBV antibodies
could be detected in 100% of the Egyptian patients, but in none of
the Swedish patients.
STAT3 and miR-1234 in association with
serum HBVsAg positivity
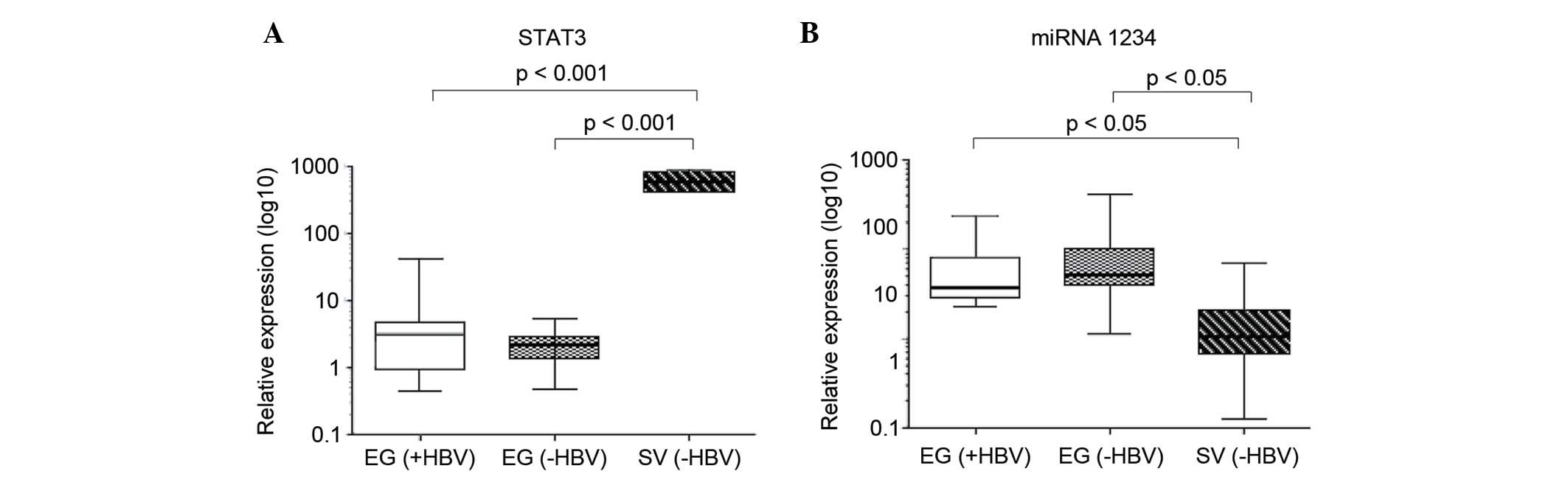
Based on the presence of the HBVsAgs in serum,
Egyptian patients were divided into a HBV+ and HBV− groups. All
Swedish patients were HBV-. The relative expression of STAT3 did
not differ between HBV+ and HBV-Egyptian patients, but was
significantly increased in the Swedish patients compared with the
total Egyptian patient population (P=0.00098 versus HB+ and
P=0.00093 versus HB− Egyptian patients; Fig. 3A). A study has recently shown that
miR-1234 may regulate STAT3, and that the relative expression of
miR-1234 is associated with HBV status (data to be published). In
the present study, there was no difference in the relative
expression of miR-1234 when comparing HBV+ and HBV− Egyptian
patients; however, HBV− Swedish patients showed a significantly
lower relative expression of miR-1234 compared with the total
Egyptian patient population (P=0.044 versus HB+ and P=0.042 versus
HB− Egyptian patients; Fig. 3B).
miR-1234 expression was inversely associated with the expression
patterns of STAT3.
Discussion
Virus-linked human cancers may be responsible for
15% of all cancers, which makes viruses the second most important
risk factor for cancer development after tobacco (37,38). The
present study compared the presence of microbial agents in the
lymphoma tissues of two groups of ABC DLBCL patients from a high
(Egypt) and low (Sweden) infectious disease region (39). The LLMDA system used in the present
study was developed at Lawrence Livermore National Laboratory,
Livermore, CA, USA, and contains probes to detect any RNA and DNA
of sequenced viruses and bacteria (32,40).
Several viruses were found and most the prevalent were HBV and JCV.
HBV was not detected in the Swedish patients or in either control
group. JCV was also present in Egyptian and Swedish patients, but
not in the controls. Overall, the Egyptian group showed higher
virus log-odd scores compared with Swedish patients. The HBVsAg
could be found in 38% of the Egyptian patients. There was no
difference in STAT3 expression compared between the HBV+ and HBV−
groups of Egyptian patients. STAT3 has previously been reported to
be dysregulated by HBV, resulting in viral persistence and
malignant transformation (21,41).
Viral infection of host cells may induce cell
proliferation and immune suppression, which are mechanisms that may
contribute to the viral-induced development of human tumors
(21). HIV infected patients have an
increased risk of developing, for example, Bcl and Kaposi sarcoma
(38). Adenovirus and poxvirus have
cell-transforming properties, but none of these have been found in
human tumor cells (38). EBV has been
recognized as a viral agent involved in the development of certain
Bcls. Barzon et al (42)
reported that patients with primary adrenal Bcl harbored EBV and
JCV together, and suggested that these viruses may act as cofactors
in the development of lymphomas (42).
Hepatitis B and C have been associated with
hepatocellular carcinoma and dermatological, hematological,
endocrinological and autoimmune disorders (43–45). HBV
reactivation may be observed in patients during treatment with
rituximab (46); however, no
conclusive evidence of an association between NHL and hepatitis
virus infection has been reported (47). Two studies showed that HBV infection
was associated with an increased risk of DLBCL in Korean
populations (48,49). An association between HBV infection
and NHL was suggested in patients with persistent HBsAg (50). A study of a Korean population reported
that 12% of NHL patients were HBV+ compared with 6% of the control
population (51). According to the
Centers for Disease Control and Prevention, 2–7% of the Egyptian
population are HBV+ (52). These
numbers were confirmed by a study reporting that 4% of Egyptian
blood donors were HBV+ (35). The
results of the present study show that lymphoma tissues of the
Egyptian patients were HBV+ and that 38% of the patients were HBV+.
None of the Swedish patients or any of the controls were HBV+. The
results indicate that Egyptian patients have an increased viral
load and that HBV was the most common virus. Since none of the
Swedish patients were HBV+, the virus is unlikely to be associated
with the development of DLBCL originating in Sweden. However, these
results may be of importance for treatment strategies in Egyptian
patients, since a previous study has demonstrated the reactivation
of HBV in patients from high endemic infectious regions during
treatment with rituximab (46).
The first human polyomaviruses (PyV), BK
polyomavirus (BKV) and JCV were isolated in 1971 (53,54). Human
PyVs are common in the general population but rarely cause
clinically apparent disease (55).
Sources of infection that have been suggested include virus-bearing
fluids, contaminated food and water (56,57). The
association of JCV with human malignancies is controversial;
however, several studies have shown JCV to be associated with human
cancers as colorectal-, gastric-, lung− and brain cancers (58–62). In a
study by Flægstad et al (63),
BKV was reported to be expressed in the tissue samples of
neuroblastomas, but not found in control tissues. This suggests
that BKV may be important in the development of neuroblastomas
through the inactivation of cell cycle regulation and apoptotic
effects, which contribute to malignant transformation (63).
BKV and JCV were recently classified as ‘possibly
carcinogenic to humans’ by the WHO international cancer research
working committee (64). The
contribution of PyVs to cancer progression can be challenged in
three ways First, the PyV may establish chromosomal instability at
an early stage of infection that contributes to cancer progression,
but PyV may not be detectable until full progression to malignancy.
Secondly, PyVs may find favorable conditions in an already
transformed cell, but be neither necessary for nor contribute to
the oncogenic characteristics. Thirdly, PyV may be detected in an
anatomically connected compartment, but be unrelated to the
malignancy (55). JCV may establish a
latent infection and e reactivated upon immunosuppression; one a
site of latency has been shown to be lymphoid cells (65). The results of the present study showed
that JCV may be detected in Swedish and Egyptian patients, but not
in Swedish and Egyptian control groups. These data are notable as
it may suggest a role for JCV in DLBCL.
Previous studies on Swedish and Egyptian patients
showed that STAT3 was overexpressed in Swedish patients compared
with Egyptian patients (26), and
that miR-1234 may be a possible regulator of STAT3 (data to be
published). No association between STAT3 and HBV expression was
detected in the present study. However, the regulation of STAT3 by
miR-1234 may also be associated with other environmental factors
and to the increased viral load as noted in the Egyptian
patients.
The current study showed that Egyptian and Swedish
patients had detectable JCV, which was not detected in Swedish and
Egyptian controls, and HBV was only detected in Egyptian patients.
Extended studies on the viral involvement in DLBCL are warranted in
order to analyze whether specific infections are associated with
the development of the disease. Such studies may be of value for
the development of novel prevention and treatment strategies.
References
|
1
|
A clinical evaluation of the International
Lymphoma Study Group classification of non-Hodgkin's lymphoma, .
The Non-Hodgkin's Lymphoma Classification Project. Blood.
89:3909–3918. 1997.PubMed/NCBI
|
|
2
|
Alizadeh AA, Eisen MB, Davis RE, Ma C,
Lossos IS, Rosenwald A, Boldrick JC, Sabet H, Tran T, Yu X, et al:
Distinct types of diffuse large B-cell lymphoma identified by gene
expression profiling. Nature. 403:503–511. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rosenwald A, Wright G, Chan WC, Connors
JM, Campo E, Fisher RI, Gascoyne RD, Muller-Hermelink HK, Smeland
EB, Giltnane JM, et al: The use of molecular profiling to predict
survival after chemotherapy for diffuse large-B-cell lymphoma. N
Engl J Med. 346:1937–1947. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wright G, Tan B, Rosenwald A, Hurt EH,
Wiestner A and Staudt LM: A gene expression-based method to
diagnose clinically distinct subgroups of diffuse large B cell
lymphoma. Proc Natl Acad Sci USA. 100:9991–9996. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu YH, Xu FP, Zhuang HG, Lai KC, Xie D,
Luo DL, Li L, Luo XL, Xu J, Zhang MH, et al: Clinicopathologic
significance of immunophenotypic profiles related to germinal
center and activation B-cell differentiation in diffuse large
B-cell lymphoma from Chinese patients. Hum Pathol. 39:875–884.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yamauchi A, Fujita S, Ikeda J, Nakamichi
I, Fukuhara S, Hino M, Kanakura Y, Ogawa H, Sugiyama H, Kanamaru A
and Aozasa K: Diffuse large B-cell lymphoma in the young in Japan:
A study by the Osaka Lymphoma Study Group. Am J Hematol.
82:893–897. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jarrett RF: Viruses and
lymphoma/leukaemia. J Pathol. 208:176–186. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Walboomers JM, Jacobs MV, Manos MM, Bosch
FX, Kummer JA, Shah KV, Snijders PJ, Peto J, Meijer CJ and Muñoz N:
Human papillomavirus is a necessary cause of invasive cervical
cancer worldwide. J Pathol. 189:12–19. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wong BC, Lam SK, Wong WM, Chen JS, Zheng
TT, Feng RE, Lai KC, Hu WH, Yuen ST, Leung SY, et al: Helicobacter
pylori eradication to prevent gastric cancer in a high-risk region
of China: A randomized controlled trial. JAMA. 291:187–194. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang JY, Dai M, Wang X, Lu WQ, Li DS,
Zhang MX, Wang KJ, Dai LP, Han SG, Zhou YF and Zhuang H: A
case-control study of hepatitis B and C virus infection as risk
factors for hepatocellular carcinoma in Henan, China. Int J
Epidemiol. 27:574–578. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Moore PS and Chang Y: Why do viruses cause
cancer? Highlights of the first century of human tumour virology.
Nat Rev Cancer. 10:878–889. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
zurHausen H: Oncogenic DNA viruses.
Oncogene. 20:7820–7823. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Parsonnet J: Microbes and malignancy:
Infection as a cause of human cancers. Oxford University Press; New
York, NY: 1999
|
|
14
|
Nogai H, Dörken B and Lenz G: Pathogenesis
of non-Hodgkin's lymphoma. J Clin Oncol. 29:1803–1811. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Capello D, Scandurra M, Poretti G,
Rancoita PM, Mian M, Gloghini A, Deambrogi C, Martini M, Rossi D,
Greiner TC, et al: Genome wide DNA-profiling of HIV-related B-cell
lymphomas. Br J Haematol. 148:245–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Engels EA, Chatterjee N, Cerhan JR, Davis
S, Cozen W, Severson RK, Whitby D, Colt JS and Hartge P: Hepatitis
C virus infection and non-Hodgkin lymphoma: Results of the NCI-SEER
multi-center case-control study. Int J Cancer. 111:76–80. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fisher SG and Fisher RI: The epidemiology
of non-Hodgkin's lymphoma. Oncogene. 23:6524–6534. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mele A, Pulsoni A, Bianco E, Musto P,
Szklo A, Sanpaolo MG, Iannitto E, De Renzo A, Martino B, Liso V, et
al: Hepatitis C virus and B-cell non-Hodgkin lymphomas: An Italian
multicenter case-control study. Blood. 102:996–999. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Davey NE, Travé G and Gibson TJ: How
viruses hijack cell regulation. Trends Biochem Sci. 36:159–169.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Smith SB, Dampier W, Tozeren A, Brown JR
and Magid-Slav M: Identification of common biological pathways and
drug targets across multiple respiratory viruses based on human
host gene expression analysis. PLoS One. 7:e331742012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Koeberlein B, zur Hausen A, Bektas N,
Zentgraf H, Chin R, Nguyen LT, Kandolf R, Torresi J and Bock CT:
Hepatitis B virus overexpresses suppressor of cytokine signaling-3
(SOCS3) thereby contributing to severity of inflammation in the
liver. Virus Res. 148:51–59. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weber-Nordt RM, Egen C, Wehinger J, Ludwig
W, Gouilleux-Gruart V, Mertelsmann R and Finke J: Constitutive
activation of STAT proteins in primary lymphoid and myeloid
leukemia cells and in Epstein-Barr virus (EBV)-related lymphoma
cell lines. Blood. 88:809–816. 1996.PubMed/NCBI
|
|
23
|
Heim MH, Moradpour D and Blum HE:
Expression of hepatitis C virus proteins inhibits signal
transduction through the Jak-STAT pathway. J Virol. 73:8469–8475.
1999.PubMed/NCBI
|
|
24
|
Lam LT, Wright G, Davis RE, Lenz G,
Farinha P, Dang L, Chan JW, Rosenwald A, Gascoyne RD and Staudt LM:
Cooperative signaling through the signal transducer and activator
of transcription 3 and nuclear factor-{kappa}B pathways in subtypes
of diffuse large B-cell lymphoma. Blood. 111:3701–3713. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Parkin DM: The global health burden of
infection-associated cancers in the year 2002. Int J Cancer.
118:3030–3044. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hogfeldt T, Bahnassy AA, Kwiecinska A,
Osterborg A, Tamm KP, Porwit A, Zekri AR, Lundahl J, Khaled HM,
Mellstedt H and Moshfegh A: Patients with activated B-cell like
diffuse large B-cell lymphoma in high and low infectious disease
areas have different inflammatory gene signatures. Leuk Lymphoma.
54:996–1003. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Högfeldt T, Johnsson P, Grandér D,
Bahnassy AA, Porwit A, Eid S, Österborg A, Zekri AR, Lundahl J,
Khaled MH, et al: Expression of microRNA-1234 related signal
transducer and activator of transcription 3 in patients with
diffuse large B-cell lymphoma of activated B-cell like type from
high and low infectious disease areas. Leuk Lymphoma. 55:1158–1165.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hans CP, Weisenburger DD, Greiner TC,
Gascoyne RD, Delabie J, Ott G, Muller-Hermelink HK, Campo E,
Braziel RM, Jaffe ES, et al: Confirmation of the molecular
classification of diffuse large B-cell lymphoma by
immunohistochemistry using a tissue microarray. Blood. 103:275–282.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meyer PN, Fu K, Greiner TC, Smith LM,
Delabie J, Gascoyne RD, Ott G, Rosenwald A, Braziel RM, Campo E, et
al: Immunohistochemical methods for predicting cell of origin and
survival in patients with diffuse large B-cell lymphoma treated
with rituximab. J Clin Oncol. 29:200–207. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jacobson TA, Lundahl J, Mellstedt H and
Moshfegh A: Gene expression analysis using long-term preserved
formalin-fixed and paraffin-embedded tissue of non-small cell lung
cancer. Int J Oncol. 38:1075–1081. 2011.PubMed/NCBI
|
|
31
|
Victoria JG, Wang C, Jones MS, Jaing C,
McLoughlin K, Gardner S and Delwart EL: Viral nucleic acids in
live-attenuated vaccines: Detection of minority variants and an
adventitious virus. J Virol. 84:6033–6040. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gardner SN, Jaing CJ, McLoughlin KS and
Slezak TR: A microbial detection array (MDA) for viral and
bacterial detection. BMC Genomics. 11:6682010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Okamoto H, Tsuda F, Sakugawa H,
Sastrosoewignjo RI, Imai M, Miyakawa Y and Mayumi M: Typing
hepatitis B virus by homology in nucleotide sequence: Comparison of
surface antigen subtypes. J Gen Virol. 69:2575–2583. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
El-Gilany AH and El-Fedawy S: Bloodborne
infections among student voluntary blood donors in Mansoura
University, Egypt. East Mediterr Health J. 12:742–748.
2006.PubMed/NCBI
|
|
36
|
European Centre for Disease Prevention and
Control, . Hepatitis B and C in the EU neighborhood, ECDC.
Technical Report. 2010, http://ecdc.europa.eu/en/publications/_layouts/forms/Publication_DispForm.aspx?ID=300&List=4f55ad51-4aed-4d32-b960-af70113dbb90Accessed
June 15, 2016.
|
|
37
|
zurHausen H: Intracellular surveillance of
persisting viral infections. Human genital cancer results from
deficient cellular control of papillomavirus gene expression.
Lancet. 2:489–491. 1986.PubMed/NCBI
|
|
38
|
zurHausen H: Viruses in human cancers.
Science. 254:1167–1173. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
World Health Organisation, . Media Centre,
Hepatitis B. http://www.who.int/mediacentre/factsheets/fs204/en/Accessed.
June 15–2016
|
|
40
|
Erlandsson L, Rosenstierne MW, McLoughlin
K, Jaing C and Fomsgaard A: The microbial detection array combined
with random Phi29-amplification used as a diagnostic tool for virus
detection in clinical samples. PLoS One. 6:e226312011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim K, Kim KH and Cheong J: Hepatitis B
virus X protein impairs hepatic insulin signaling through
degradation of IRS1 and induction of SOCS3. PLoS One. 5:e86492010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Barzon L, Trevisan M, Marino F, Guzzardo V
and Palù G: Primary bilateral adrenal B-cell lymphoma associated
with EBV and JCV infection. Infect Agent Cancer. 4:12009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Anzola M: Hepatocellular carcinoma: Role
of hepatitis B and hepatitis C viruses proteins in
hepatocarcinogenesis. J Viral Hepat. 11:383–393. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dammacco F, Sansonno D, Piccoli C,
Racanelli V, D'Amore FP and Lauletta G: The lymphoid system in
hepatitis C virus infection: Autoimmunity, mixed cryoglobulinemia,
and Overt B-cell malignancy. Semin Liver Dis. 20:143–157. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gumber SC and Chopra S: Hepatitis C: A
multifaceted disease. Review of extrahepatic manifestations. Ann
Intern Med. 123:615–620. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Niitsu N, Hagiwara Y, Tanae K, Kohri M and
Takahashi N: Prospective analysis of hepatitis B virus reactivation
in patients with diffuse large B-cell lymphoma after rituximab
combination chemotherapy. J Clin Oncol. 28:5097–5100. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Okan V, Yilmaz M, Bayram A, Kis C, Cifci
S, Buyukhatipoglu H and Pehlivan M: Prevalence of hepatitis B and C
viruses in patients with lymphoproliferative disorders. Int J
Hematol. 88:403–408. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Engels EA, Cho ER and Jee SH: Hepatitis B
virus infection and risk of non-Hodgkin lymphoma in South Korea: A
cohort study. Lancet Oncol. 11:827–834. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kang J, Cho JH, Suh CW, Lee DH, Oh HB,
Sohn YH, Chi HS, Park CJ, Jang SS, Lee KH, et al: High prevalence
of hepatitis B and hepatitis C virus infections in Korean patients
with hematopoietic malignancies. Ann Hematol. 90:159–164. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Marcucci F, Spada E, Mele A, Caserta CA
and Pulsoni A: The association of hepatitis B virus infection with
B-cell non-Hodgkin lymphoma-a review. Am J Blood Res. 2:18–28.
2012.PubMed/NCBI
|
|
51
|
Kim JH, Bang YJ, Park BJ, Yoo T, Kim CW,
Kim TY, Heo DS, Lee HS and Kim NK: Hepatitis B virus infection and
B-cell non-Hodgkin's lymphoma in a hepatitis B endemic area: A
case-control study. Jpn J Cancer Res. 93:471–477. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Centers for Disease Control and
Prevention, . Viral Hepatitis - Hepatitis B Information. http://www.cdc.gov/hepatitis/hbv/Accessed. June
29–2016
|
|
53
|
Gardner SD, Field AM, Coleman DV and Hulme
B: New human papovavirus (B.K.) isolated from urine after renal
transplantation. Lancet. 1:1253–1257. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Padgett BL, Walker DL, ZuRhein GM,
Eckroade RJ and Dessel BH: Cultivation of papova-like virus from
human brain with progressive multifocal leucoencephalopathy.
Lancet. 1:1257–1260. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dalianis T and Hirsch HH: Human
polyomaviruses in disease and cancer. Virology. 437:63–72. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bofill-Mas S, Formiga-Cruz M,
Clemente-Casares P, Calafell F and Girones R: Potential
transmission of human polyomaviruses through the gastrointestinal
tract after exposure to virions or viral DNA. J Virol.
75:10290–10299. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bofill-Mas S, Pina S and Girones R:
Documenting the epidemiologic patterns of polyomaviruses in human
populations by studying their presence in urban sewage. Appl
Environ Microbiol. 66:238–245. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Boldorini R, Pagani E, Car PG,
Omodeo-Zorini E, Borghi E, Tarantini L, Bellotti C, Ferrante P and
Monga G: Molecular characterisation of JC virus strains detected in
human brain tumours. Pathology. 35:248–253. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Delbue S, Pagani E, Guerini FR, Agliardi
C, Mancuso R, Borghi E, Rossi F, Boldorini R, Veggiani C, Car PG
and Ferrante P: Distribution, characterization and significance of
polyomavirus genomic sequences in tumors of the brain and its
covering. J Med Virol. 77:447–454. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Laghi L, Randolph AE, Chauhan DP, Marra G,
Major EO, Neel JV and Boland CR: JC virus DNA is present in the
mucosa of the human colon and in colorectal cancers. Proc Natl Acad
Sci USA. 96:7484–7489. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lin PY, Fung CY, Chang FP, Huang WS, Chen
WC, Wang JY and Chang D: Prevalence and genotype identification of
human JC virus in colon cancer in Taiwan. J Med Virol.
80:1828–1834. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zheng H, Aziz HO Abdel, Nakanishi Y,
Masuda S, Saito H, Tsuneyama K and Takano Y: Oncogenic role of JC
virus in lung cancer. J Pathol. 212:306–315. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Flaegstad T, Andresen PA, Johnsen JI,
Asomani SK, Jorgensen GE, Vignarajan S, Kjuul A, Kogner P and
Traavik T: A possible contributory role of BK virus infection in
neuroblastoma development. Cancer Res. 59:1160–1163.
1999.PubMed/NCBI
|
|
64
|
Bouvard V, Baan RA, Grosse Y,
Lauby-Secretan B, El Ghissassi F, Benbrahim-Tallaa L, Guha N and
Straif K: WHO International Agency for Research on Cancer Monograph
Working Group: Carcinogenicity of malaria and of some
polyomaviruses. Lancet Oncol. 13:339–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lafon ME, Dutronc H, Dubois V, Pellegrin
I, Barbeau P, Ragnaud JM, Pellegrin JL and Fleury HJ: JC virus
remains latent in peripheral blood B lymphocytes but replicates
actively in urine from AIDS patients. J Infect Dis. 177:1502–1505.
1998. View
Article : Google Scholar : PubMed/NCBI
|