Introduction
Mitochondria are important in cell and energy
metabolism, and are required for cell apoptosis. Mitochondrial
dysfunction has been associated with a wide range of degenerative
and metabolic diseases, including ageing and cancer (1). According to the Warburg hypothesis
(2), dysfunction of mitochondria has
been postulated to render cancer cells resistant to apoptosis.
Mitochondrial DNA (mtDNA) possesses several unique
characteristics (3–5), such as non-Mendelian maternal
inheritance, a mutation rate that is 10–20 times higher than
nuclear DNA, the threshold effect and heteroplasmy (6). In addition, there are 100–1,000
mitochondria per cell and 2–10 copies of circular double-stranded
mitochondrial genomes per mitochondrion (6). When a mutation arises in mtDNA, it
creates a coexistence of wild-type and mutant mtDNA, resulting in
heteroplasmy. When a heteroplasmic cell divides, the two variants
of mtDNA are randomly distributed into the daughter cells,
resulting in genetic drift; thus, the proportions of mutant mtDNA
in these daughter cells ranges between 1 and 100% (4,7). As the
percentage of mutant mtDNA increases to a threshold value, tissue
dysfunction and clinical symptoms appear in the individual, and
apoptosis or necrosis may be initiated (8,9).
Therefore, the proportion of mutant mtDNA may contribute to the
clinical symptoms of mitochondrial disease, including cancer
(9).
Previously, somatic mtDNA mutations, including point
mutations, insertions and deletions, were identified in various
types of human cancer, including non-small cell lung cancer (NSCLC)
(10). Furthermore, mtDNA 8701 and
10398, which code for ATPase 6 and NADH dehydrogenase 3,
respectively, were identified as mutational hotspots in the
mitochondrial genome of patients with lung cancer (11). African American women carrying
wild-type mtDNA 10398A were observed to have an increased risk of
invasive breast cancer (12). In
cancer, it has been hypothesized that these somatic mtDNA
alterations may lead to a switch in the energy supply between
mitochondrial oxidative phosphorylation and aerobic glycolysis,
leading to cancer progression (13).
However, few studies have investigated the prognostic value of
heteroplasmy of mutant mtDNA 10398G in patients with NSCLC. The
proportion of mutant mtDNA must reach the threshold to cause a
clinical phenotype (14); therefore,
in order to predict the clinical outcome of a patient, the
proportion of mutant DNA in the affected or relevant tissue should
be determined.
In the present study, the heteroplasmy of mtDNA
A10398G was assessed in tumor tissues from 129 patients with NSCLC
using an amplification refractory mutation system-quantitative
polymerase chain reaction (ARMS-qPCR) assay. A previous study
demonstrated that the introduction of one or two mismatched
nucleotides immediately 5′ to the mutation site markedly increases
the binding specificity of the allele-specific modified primers
toward the wild-type or mutant sequence targets (15). In addition, the present study analyzed
the prognostic value of mtDNA A10398G. The present results
demonstrated that the proportion of mutant heteroplasmy provides a
novel biomarker in predicting the outcome of patients with NSCLC.
Thus, intervention of heteroplasmy using mtDNA A10398G may become a
novel approach for cancer treatment.
Materials and methods
Patient eligibility and clinical
data
A total of 129 NSCLC tissue specimens were collected
from patients that underwent surgical resection, including lymph
node biopsy, at Zhongnan Hospital of Wuhan University (Wuhan,
China) between July 2006 and July 2011. None of patients had
received radiotherapy or chemotherapy prior to surgery. Clinical
staging was assessed according to the American Joint Committee on
Cancer (AJCC, seventh edition; https://cancerstaging.org/Pages/default.aspx).
Formalin-fixed and paraffin-embedded surgical tissue samples
collected from the Department of Pathology at the Zhongnan Hospital
of Wuhan University were examined according to the World Health
Organization 2004 classification system (16). The patients consisted of 96 men and 33
women (age range, 20–80 years). All participants provided prior
written informed consent. The study protocol was reviewed and
approved by the Institutional Review Board of Zhongnan Hospital of
Wuhan University. Clinical information, including age, gender,
pathological type, tumor-node-metastasis stage, smoking status,
epidermal growth factor receptor (EGFR) mutation status and
clinical follow-up data, was recorded prospectively (Table I). Patients were followed up at
3-month intervals for ≤2 years, then every 6 months for 5 years,
and annually thereafter until the final follow-up on November 26,
2012. The median follow-up time and overall survival (OS) time were
20.6 and 26.3 months respectively; 69 patients (53.5%) succumbed
during this period, 46 patients (35.7%) survived and 14 patients
(10.9%) were lost to follow-up.
 | Table I.Patient characteristics of 129
patients with non-small cell lung cancer. |
Table I.
Patient characteristics of 129
patients with non-small cell lung cancer.
| Variable | Value |
|---|
| Gender, n (%) |
|
| Male | 96
(74.4) |
|
Female | 33
(25.6) |
| Age, years |
|
| Mean | 61.2 |
|
Range | 20–80 |
| Smoking status, n
(%) |
|
| No | 50
(38.8) |
| Yes | 79
(61.2) |
| Histological status,
n (%)a |
| AC | 54
(41.9) |
| SCC | 52
(40.3) |
| ASC | 15
(11.6) |
|
Other | 8
(6.2) |
| EGFR mutation, n
(%) |
|
| No | 115 (89.1) |
| Yes | 14
(10.9) |
| Stage, n
(%)b |
|
| I | 23
(17.9) |
| II | 27
(20.9) |
| III | 63
(48.8) |
| IV | 16
(12.4) |
| Follow-up status, n
(%) |
|
|
Survival | 46
(35.7) |
|
Mortality | 69
(53.5) |
| Lost to
follow-up | 14
(10.9) |
Tissue genomic DNA isolation
Total DNA was extracted from formalin-fixed,
paraffin-embedded tissues using the E.Z.N.A.® FFPE DNA
kit (Omega Bio-Tek, Inc., Norcross, GA, USA), according to the
manufacturer's protocol. The isolated DNA was eluted in 100 µl
Tris-EDTA buffer included in the E.Z.N.A.® FFPE DNA kit
and stored at −20°C until required. The quality of the isolated DNA
was assessed by NanoDrop spectrophotometer (NanoDrop Technologies,
Thermo Fisher Scientific, Inc., Wilmington, DE, USA). The optical
density values of all samples ranged from 1.8 to 2.0.
Primers for ARMS-qPCR
The forward primers for ARMS-qPCR analysis of the
mtDNA A10398G mutation are listed in Table II. The primers were designed by
Primer Premier 6.0 (Premier Biosoft International, Palo Alto, CA,
USA) and generated by Sangon Biotech Co., Ltd. (Shanghai, China).
The numbers in Table I correspond to
the nucleotide positions in GenBank (accession no. NC012920). Two
mismatches were introduced at the penultimate and antepenultimate
nucleotide positions of the mutation site in the forward primers
(Table II) to increase the
specificity of the ARMS reaction, as described by Newton et
al (15). At the mutation site,
the wild-type primer containing an A would perfectly match the
wild-type target sequence, but would be a weak AC mismatch with the
mutant target sequence. Similarly, the mutant primer containing a G
would be a perfect match with the mutant target sequence, but would
be a weak GT mismatch with the wild-type target sequence.
Therefore, two clear TT and CC mismatches at the two nucleotides
immediately 5′ to the mutation site were introduced, which were
hypothesized to increase the primer specificity (15). The reverse primer was at nucleotide
positions 10753–10776 for the amplification of the mtDNA A10398G
mutation. However, the wild-type primer produced two PCR products
of different sizes, indicating that the specificity of the
wild-type primer was not sufficient. Therefore, two and one
nucleotides were added to the forward and reverse primers,
respectively, at each 5 terminal (Table
II), which improved the specificity, since only one PCR product
was generated upon addition of the above nucleotides. PCR products
amplified by the two pairs of primers were identified by agarose
gel electrophoresis and sequencing, as follows.
| Table II.Primers used in amplification
refractory mutation system-quantitative polymerase chain
reaction. |
Table II.
Primers used in amplification
refractory mutation system-quantitative polymerase chain
reaction.
| Primer | Sequence,
5′-3′ |
|---|
| Forward (nucleotide
position 10373–10398) |
|
|
Wild-type sequence | GT
GACTACAAAAAGGATTAGACT g
a A |
|
Wild-type 10398A primer | GT
GACTACAAAAAGGATTAGACT c
t A |
| Mutated
10398G primer |
GACTACAAAAAGGATTAGACT
c t G |
| Reverse (nucleotide
position 10753–10777) |
|
|
Wild-type sequence |
TACTCCAATGCTAAAACTAATCGT C |
|
Antisense strand sequence | G
ACGATTAGTTTTAGCATTGGAGTA |
|
Wild-type 10398A primer | G
ACGATTAGTTTTAGCATTGGAGTA |
| Mutated
10398G primer |
ACGATTAGTTTTAGCATTGGAGTA |
ARMS-qPCR
qPCR was performed using a 20-µl PCR reaction
mixture, containing 100 ng DNA template (2 µl), 10 µl QuantiTect
SYBR-Green PCR Master Mix (Takara Bio, Inc., Otsu, Japan), 200 nM
of each primer (Table II), 0.4 µl
ROX Reference Dye II (Takara Bio, Inc.) and 7.2 µl
ddH2O, according to the manufacture's protocol. As
reference gene, s a single copy region of the nuclear gene β2
microglobulin (β2M), was used, which was amplified with the
following primers: β2M forward, 5′-GCTGGGTAGCTCTAAACAATGTATTCA-3′
and reverse, 5′-CCATGTACTAACAAATGTCTAAAATGGT-3′ (93 bp product).
The PCR cycling conditions were as follows: 2 min at 50°C and 10
min at 95°C; followed by 40 cycles of denaturation for 15 sec at
95°C and annealing/extension for 60 sec at 60°C. The fluorescent
signal intensities were recorded and analyzed using an ABI Prism
7300 Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Dissociation curves were
generated following each run to confirm that the increased
fluorescence intensities were not associated with primer-dimer
formation. The quantification cycle (Cq) values (17) within the linear exponential increase
phase were used to construct the calibration curve and to measure
the original DNA template content, as well as to calculate the
mutant heteroplasmy of each sample.
Preparation of standard curves
The fragments of wild-type and mutant target
sequences were amplified by the specific primers presented in
Table II. Subsequently, the products
were analyzed by electrophoresis on a 2% agarose gel. Bands of
interest were cut from the gel and purified using a commercial kit
(E.Z.N.A Gel Extraction kit; Omega Bio-Tek, Inc.). Plasmids were
constructed by cloning the purified PCR products into the
pGEM®-T Easy Vector (Promega Corporation, Madison, WI,
USA), according to the manufacturer's protocol, and subsequently
transformed into competent JM109 cells (Beijing Transgen Biotech
Co., Ltd., Beijing, China) by the heat shock method. The clones
were identified by digesting the plasmid DNA with NotI
(Fermentas; Thermo Fisher Scientific, Inc.; Pittsburgh, PA, USA).
The positive plasmids were sequenced and their nucleotide sequences
were analyzed using Lasergene version 7.1 software (DNASTAR, Inc.,
Madison, WI, USA). The copy numbers were calculated based on the
size and molecular weight of the plasmid DNA. Five 10-fold serial
dilutions of plasmid DNA between 105 and 109
molecules were prepared, and ARMS-qPCR reactions were performed to
construct the calibration curves for the wild-type and mutant mtDNA
10398 sequences. The slope of the curves was determined and the
corresponding qPCR efficiency for the specific amplification was
calculated according to the following equation (18): E = 10(−1 / slope) - 1.
Measurement of mutant
heteroplasmy
The ARMS-qPCR assay was performed three times for
each wild-type and mutant target sequence of each sample by
relative quantification to detect the heteroplasmy of mtDNA
A10398G. The proportion of the mutant mtDNA that was used to assess
the degree of mutant heteroplasmy was calculated from
ΔCq (Cqwild-type - Cqmutant),
using the formula: Proportion of mutant (%) = 1 /
(1+1/2ΔCq) × 100% (18).
Detection of mtDNA copy number and
EGFR mutations
Quantification of a unique fragment in the human
mitochondrial genome (accession no. NC012920) relative to a single
copy region of the nuclear gene β2M was performed using a qPCR
assay to assess mtDNA content (19),
using reagents purchased from Sangon Biotech Co., Ltd.
PCR-restriction fragment length polymorphism was
employed for mutation analysis of EGFR exons 18, 19 and 21
(19), using reagents purchased from
Sangon Biotech Co., Ltd. The mutant allele of exons 18 and 21 were
not digested by the restriction enzymes ApaI and MscI
(New England BioLabs, Inc., Ipswich, MA, USA), respectively. Exon
19 deletion was distinguished by the generation of PCR products of
different sizes.
Statistical analysis
Statistical analyses were performed with SPSS
version 17.0 (SPSS, Inc., Chicago, IL, USA). The association
between heteroplasmy of mtDNA A10398G and survival time was
analyzed using the χ2 test. Kaplan-Meier survival curves
and the log-rank test were used to analyze OS time. Univariate and
multivariate Cox proportional hazard models were used to assess the
effects of all clinical variables on patient survival (Table I). The parameters were then tested
using the multivariate Cox proportional hazards model, which was
performed to identify the independent variables for predicting
survival. Two-sided P<0.05 was considered to indicate a
statistically significant difference.
Results
Assay specificity, standard
amplification curves and amplification efficiency
The ARMS-qPCR primer system, for which the primers
were specifically designed, exhibited high specificity and qPCR
efficiency. The wild-type primer only amplified the wild-type
target sequence, but did not amplify the mutant target sequence.
Similarly, the mutant primer only targeted the mutant target
sequence, but did not target the wild-type target sequence
(Fig. 1).
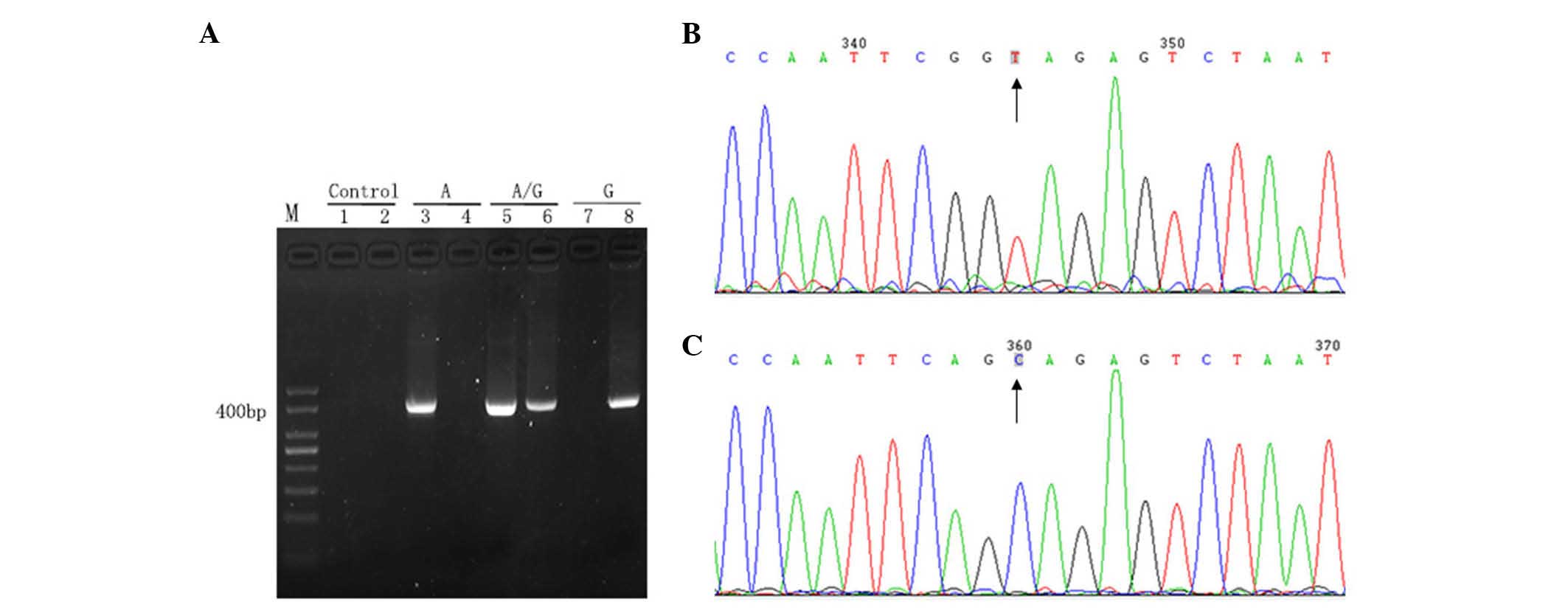 | Figure 1.Analysis of primer specificity. (A)
Gel electrophoresis of the PCR products. Lane M, 50 bp DNA ladder;
lane 1, negative controls of wild-type primers (ddH2O
instead of DNA template was added to the PCR); lane 2, negative
control of mutation patterns (ddH2O instead of DNA
template was added to the PCR); lanes 3 and 4, plasmid with
wild-type sequence amplified using wild-type (lane 3) and mutant
(lane 4) primers; lanes 5 and 6, plasmid with mutant and wild-type
sequences amplified using wild-type (lane 5) and mutant (lane 6)
primers; lanes 7 and 8, plasmid with mutant sequence amplified
using wild-type (lane 7) and mutant (lane 8) primers. (B and C)
Reverse sequence (detecting the sequence of the complementary
strand) results of PCR products extended using two pairs of
primers: (B) Wild-type (TA) and (C) mutant (CG). Results were
unimodal, which indicated high specificity of the primers.
Subsequently, these primers were used to amplify reconstructed
plasmids. PCR, polymerase chain reaction; dd, double distilled. |
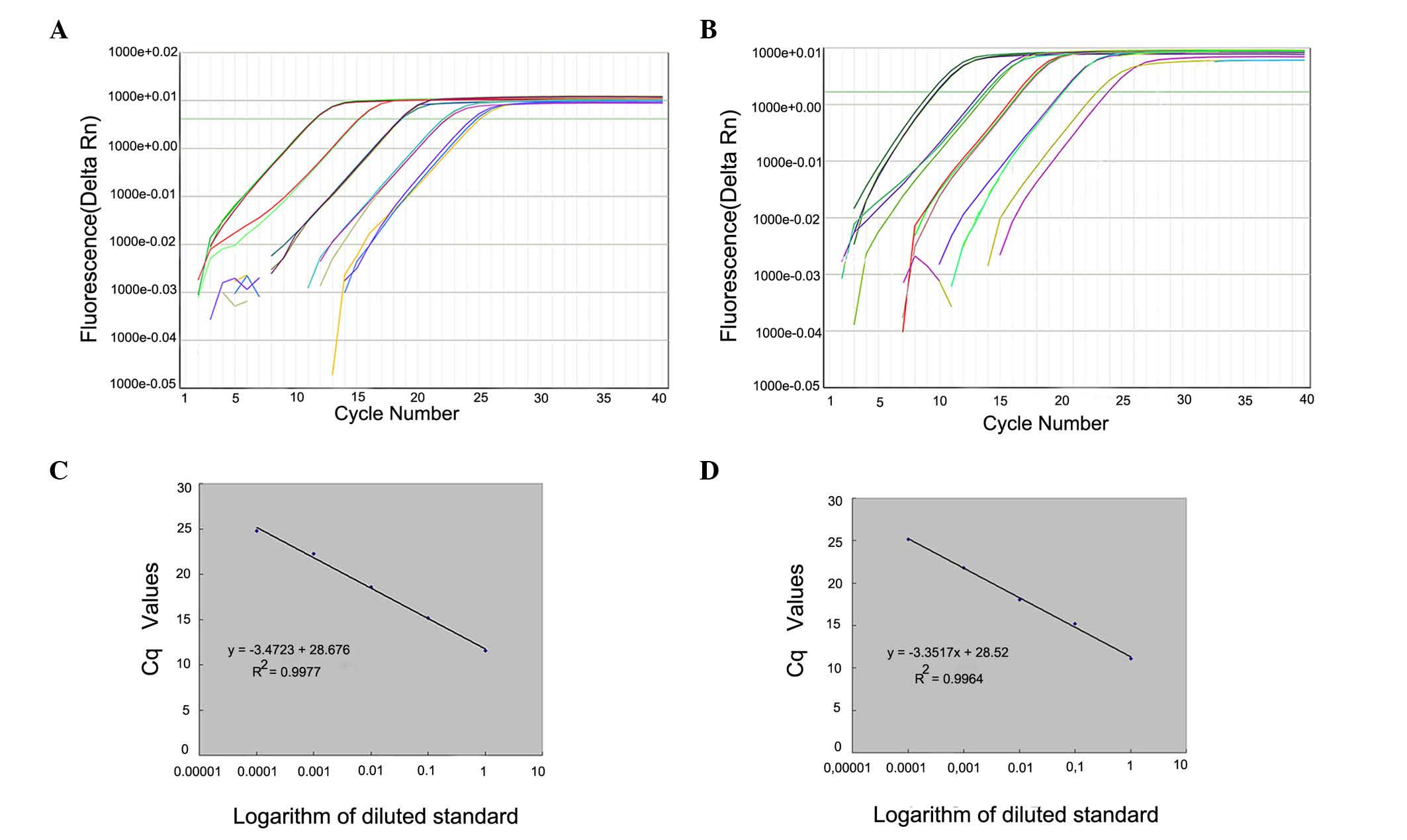
Calibration curves were generated from duplicate
samples with high linearity. Quantitative equations were as
follows: Wild-type, y = −3.3517x + 28.52 (R2=0.9964);
and mutant, y = −3.4723x + 28.676 (R2=0.9977) where ×
and y represent the Cq value of qPCR and the logarithmic value of
the DNA copy numbers, respectively. The corresponding amplification
efficiencies that were calculated from the slopes of two equations,
Ewild-type=98.77% and Emutant=94%, were in
the optimum range, since both efficiency values were >90% and
the difference between them was <5% (Fig. 2).
Prognostic significance of mtDNA
A10398G heteroplasmy
The mean Cq values for wild-type and mutant
sequences in NSCLC tissues ranged between 22.11 and 35.71, and
between 22.10 and 36.06, respectively. The proportion of mutant
heteroplasmy in the NSCLC tissue samples ranged between 0.31 and
97.04%. The mtDNA content of the NSCLC tissue samples ranged
between 23.8 and 1,833.0 copy numbers (data not shown).
There are few studies published at present regarding
heteroplasmic mutations, and none regarding mtDNA 0398
heteroplasmic mutation. In the present study, it was observed that
heteroplasmic mutations were not normally distributed. therefore,
they were divided into low proportion (≤ median) and high
proportion (> median) on the basis of their median mtDNA
content, as previously analyzed (20). A low proportion of mutant mtDNA 10398G
was observed marginally less often in adenocarcinoma compared with
other histological types of NSCLC tissue (32.8 vs. 67.2%,
respectively; χ2=3.567; P=0.050). There was no
association between proportion of mutant mtDNA 10398G, and gender,
age, smoking status, EGFR mutation status or tumor stage
(P>0.05; Table III).
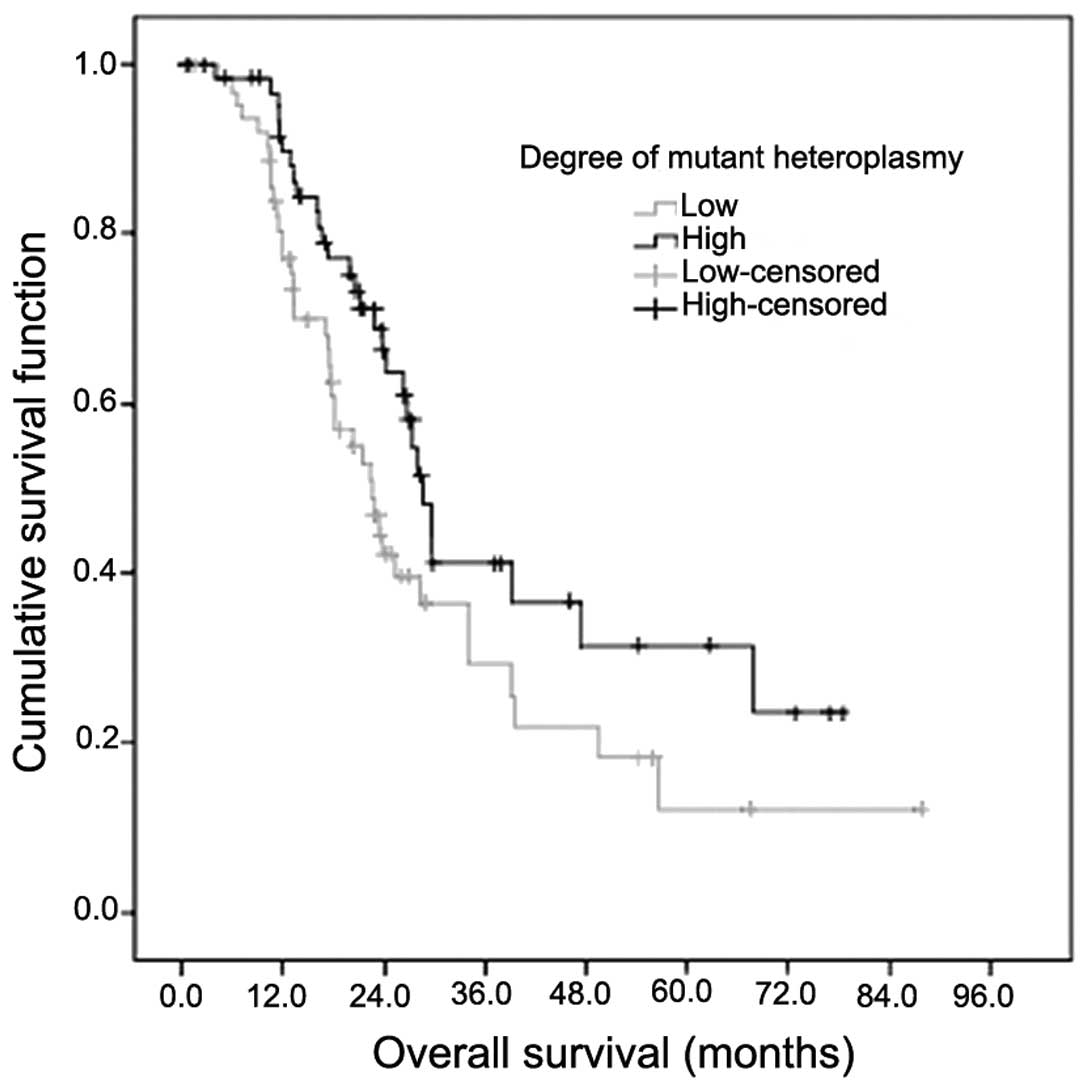
Subsequently, mutant mtDNA 10398G heteroplasmy was subjected to
survival analysis. Patients with a low percentage of mutant mtDNA
10398G heteroplasmy had a significantly shorter survival time
compared with patients with a high proportion of mutant mtDNA
10398G heteroplasmy (median survival time 22.5 vs. 28.7 months,
respectively; χ2=5.656; P=0.017; Fig. 3). All the analyzed clinical parameters
(gender, age, smoking status, histology, EGFR mutation status and
tumor stage) and mtDNA (mutant mtDNA A10398G heteroplasmy and mtDNA
content) were entered into a Cox proportional hazards regression
analysis. The univariate analysis revealed that the EGFR mutation
status (P<0.001), stage of tumor (P=0.044) and mutant mtDNA
A10398G heteroplasmy (P=0.038) were significantly associated with
prognosis. The prognosis exhibited no significant difference for
age (P=0.354), gender (P=0.405), smoking status (P=0.319), tumor
histology (P=0.463) or mtDNA content (P=0.764). In the multivariate
statistical analysis, the Cox regression analysis demonstrated that
EGFR mutation status and possessing a low degree of mutant mtDNA
A10398G were two independent prognostic factors in patients with
NSCLC (χ2=24.595, P<0.001 and χ2= 0.557,
P=0.022, respectively) (Table IV).
Forward: Wald, P=0.05, entry and P=0.10, removal (Wald probability
and statistical method, forward by stepwise selection of
variables).
| Table III.Associations between mutant
heteroplasmy proportions of mtDNA 10398 and clinical parameters in
129 patients with non-small cell lung cancer. |
Table III.
Associations between mutant
heteroplasmy proportions of mtDNA 10398 and clinical parameters in
129 patients with non-small cell lung cancer.
| Variable | Low mutant
proportion, n (%) | High mutant
proportion, n (%) | χ2 | P-value | OR (95% CI) |
|---|
| Total | 64 (49.6) | 65 (50.4) |
|
| Gender |
|
| 0.000 | 1.000 | 1.063
(0.482–2.344) |
|
Male | 48 (50.0) | 48 (50.0) |
|
|
Female | 16 (48.5) | 17 (51.5) |
|
| Age, years |
|
| 0.916 | 0.321 | 0.542
(0.199–1.481) |
|
<50 | 7 (36.8) | 12 (63.2) |
|
|
≥50 | 57 (51.8) | 53 (48.2) |
|
| Smoking status |
|
| 0.063 | 0.720 | 1.169
(0.575–2.375) |
| No | 26 (52.0) | 24 (48.0) |
|
|
Yes | 38 (48.1) | 41 (51.9) |
|
| Histology |
|
| 3.567 | 0.050 | 0.474
(0.232–0.966) |
| AC | 21 (38.9) | 33 (61.1) |
|
|
Other | 43 (57.3) | 32 (42.7) |
|
| EGFR mutation |
|
| 0.064 | 0.778 | 1.357
(0.443–4.158) |
| No | 58 (50.4) | 57 (49.6) |
|
|
Yes | 6
(42.9) | 8
(57.1) |
|
| Stage |
|
| 0.012 | 0.857 | 0.900
(0.443–1.828) |
| I +
II | 24 (48.0) | 26 (52.0) |
|
| III +
IV | 40 (50.6) | 39 (49.4) |
|
| Table IV.Multivariate analysis of overall
survival. |
Table IV.
Multivariate analysis of overall
survival.
| Variables | Wald | P-value | HR | 95% CI |
|---|
| EGFR mutation | 24.595 | <0.001 | 5.662 | (2.854,
11.235) |
| Heteroplasmy |
5.257 |
0.022 | 0.557 | (0.338, 0.919) |
Discussion
Mitochondria are semi-autonomous cytoplasmic
organelles that are involved in the production of energy for cell
metabolism. Each mitochondria contains 2–10 copies of its genome.
Double-stranded circular mtDNA is unique genetic material located
outside of the cell nucleus and has a higher mutation rate compared
with nuclear DNA (6). mtDNA is key in
maintaining organelles that are functionally competent and an
accumulation of somatic mtDNA mutations may affect energy
production; therefore, it is crucial in cancer pathogenesis and
progression (21).
mtDNA mutations have been increasingly observed to
be potentially involved in cancer. mtDNA 8701 and 10398, which code
for ATPase6 and NADH dehydrogenase 3 respectively, were reported to
have the highest mutation frequencies in the mitochondrial genome
in lung cancer. The A-to-G point mutation of mtDNA 10398 (A10398G)
leads to the conversion of Ala to Thr, resulting in an alteration
in the structure of complex I in the mitochondrial electron
transport chain, which is an important site of free radical
production. This polymorphism is associated with several
neurodegenerative disorders. For example, a previous study
demonstrated that African-American female patients with the
wild-type mtDNA 10398A allele had a significantly increased risk of
invasive breast cancer (20). In
addition, the wild-type mtDNA 10398A variant in African American
women with breast cancer provides resistance to cell apoptosis and
promotes metastasis in mice (22). In
NSCLC, patients with a high mtDNA content in combination with the
mutant mtDNA 10398G allele had a significantly increased overall
survival time compared with those that had a low mtDNA content and
the wild-type mtDNA 10398A allele (19).
Presently, few studies have reported the
heteroplasmy of wild-type mtDNA 10398A. However, the potential
involvement of mutant heteroplasmy and tumor biological behavior
has been indicated by studies at other positions; for example, the
proportion of mutant mtDNA 3243 in patients with mitochondrial
disease was detected at between 0.02 and 100% (5). However, the proportion of mutant
mitochondria must reach the threshold to cause a clinical
phenotype. Depending on the proportion of mutant heteroplasmy and
the affected tissue, patients with the mtDNA A3243G mutation may be
completely unaffected, or present with a broad spectrum of
diseases, including diabetes, deafness or a MELAS phenotype
(mitochondrial myopathy, encephalopathy, lactic acidosis and
stroke-like episodes) (23). In
addition, mutation copy number determined using ARMS-qPCR was
associated with the degree of deafness in a small number of
patients with heteroplasmic mutations of mtDNA A1555G (14). Another study reported five family
pedigrees with multiple members who had the mtDNA A1555G mutation,
and exhibited diverse clinical manifestations and various
heteroplasmy levels; the risk of deafness rose with the increase in
heteroplasmy level (24). Warowicka
et al (25) identified that
mtDNA copy numbers with a 4.977 bp deletion were significantly
different at various stages of carcinogenesis in control, low-grade
squamous intraepithelial lesions (L-SIL), high grade-SIL (H-SIL)
and cervical cancer samples. In L-SIL cells, only one third of the
mtDNA copies did not contain a deletion, while in H-SIL and
cervical cancer cells the proportion that contained a deletion
reduced to almost half (24).
However, to the best of our knowledge, the association between the
mtDNA G10398A polymorphism and NSCLC prognosis has not been
reported. In the present study, the overall survival time increased
by 27.6% and the mortality risk decreased in NSCLC patients with a
high proportion of mutant mtDNA 1039 compared with a low proportion
of mutant mtDNA 10398G (median survival time, 28.7 vs. 22.5 months,
respectively; χ2=5.656; P=0.017).
The present study confirmed that the ARMS-qPCR assay
is a rapid, sensitive, reliable and cost-effective one-step qPCR
method for the quantification of mutant mtDNA heteroplasmy. This
method may be used for the quantification of heteroplasmy for any
mtDNA point mutation (5,23). However, it is challenging to identify
the optimal allele-specific primers and mode for qPCR to ensure
correct hybridization of allele-specific primers (26). The present study designed several
pairs of primers, as reported in previous studies (14,15,18), and
each amplified PCR product was identified by sequencing and
electrophoresis to ensure allele specificity. Subsequently, the
primers with strict specificity were selected for use inARMS-qPCR.
The mtDNA 10400 was included in the sequence amplified by the
specific primers with presence of mutation, which caused the
difference in the site located two bases prior to mtDNA 10398 (T
and C, Fig. 1B and C). However, the
site was located downstream of mtDNA 10398, not in the primer
binding region, and therefore, did not influence the study of mtDNA
10398 mutation heteroplasmy.
In conclusion, the present study investigated the
association between the heteroplasmic mtDNA A10398G mutation and
the prognosis of patients with NSCLC. There was no association
between the proportion of mutant mtDNA, and gender, smoking status,
tumor stage, EGFR mutation status or mtDNA content. Furthermore,
the present study confirmed that a low proportion of mutant mtDNA
10398G may be a marker of poor prognosis in patients with NSCLC.
However, the mtDNA A10398G mutation and the accumulation of this
mutation may result in mitochondrial dysfunction, sequentially
affecting biological behaviors and the sensitivity to anticancer
treatment, leading to an alteration in the prognosis of patients.
The small size of the patient cohort in the present study was also
a limitation of the study. Therefore, additional studies are
required to verify the prognostic value of mtDNA A10398G
heteroplasmy in various types of cancer, and identify the threshold
value that causes a clinical phenotype.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (Beijing, China; grant no.
81472798).
References
|
1
|
Wallace DC, Fan W and Procaccio V:
Mitochondrial energetics and therapeutics. Annu Rev Pathol.
5:297–348. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sanchez-Aragó M, Chamorro M and Cuezva JM:
Selection of cancer cells with repressed mitochondria triggers
colon cancer progression. Carcinogenesis. 31:567–576. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johns DR: Seminars in medicine of the Beth
Israel Hospital, Boston. Mitochondrial DNA and disease. N Engl J
Med. 333:638–644. 1995.PubMed/NCBI
|
|
4
|
Schon EA: Mitochondrial genetics and
disease. Trends Biochem Sci. 25:555–560. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bai RK and Wong LJ: Detection and
quantification of heteroplasmic mutant mitochondrial DNA by
real-time amplification refractory mutation system quantitative PCR
analysis: A single-step approach. Clin Chem. 50:996–1001. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Larsen NB, Rasmussen M and Rasmussen LJ:
Nuclear and mitochondrial DNA repair: Similar pathways?
Mitochondrion. 5:89–108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rozwodowska M, Drewa G, Zbytniewski Z,
Woźniak A, Krzyzyńska-Malinowska E and Maciak R: Mitochondrial
diseases. Med Sci Monit. 6:817–822. 2000.PubMed/NCBI
|
|
8
|
Wallace DC: The mitochondrial genome in
human adaptive radiation and disease: On the road to therapeutics
and performance enhancement. Gene. 354:169–180. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wallace DC: Mitochondrial DNA mutations in
disease and aging. Environ Mol Mutagen. 51:440–450. 2010.PubMed/NCBI
|
|
10
|
Lin CS, Wang LS, Tsai CM and Wei YH: Low
copy number and low oxidative damage of mitochondrial DNA are
associated with tumor progression in lung cancer tissues after
neoadjuvant chemotherapy. Interact Cardiovasc Thorac Surg.
7:954–958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Choi SJ, Kim SH, Kang HY, Lee J, Bhak JH,
Sohn I, Jung SH, Choi YS, Kim HK, Han J, et al: Mutational hotspots
in the mitochondrial genome of lung cancer. Biochem Biophys Res
Commun. 407:23–27. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pezzotti A, Kraft P, Hankinson SE, Hunter
DJ, Buring J and Cox DG: The mitochondrial A10398G polymorphism,
interaction with alcohol consumption and breast cancer risk. PLoS
One. 4:e53562009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jin X, Zhang J, Gao Y, Ding K, Wang N,
Zhou D, Jen J and Cheng S: Relationship between mitochondrial DNA
mutations and clinical characteristics in human lung cancer.
Mitochondrion. 7:347–353. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zu-Jian C, Bin Y, Qi-Cai L, Lin J, Jing C
and Qi-Shui O: Quantification of mitochondrial DNA with the A1555G
mutation in deaf patients using real-time amplification refractory
mutation system-quantitative PCR. J Mol Diagn. Dec 3–2009.(Epub
ahead of print). PubMed/NCBI
|
|
15
|
Newton CR, Graham A, Heptinstall LE,
Powell SJ, Summers C, Kalsheker N, Smith JC and Markham AF:
Analysis of any point mutation in DNA. The amplification refractory
mutation system (ARMS). Nucleic Acids Res. 17:2503–2516. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Beasley MB, Brambilla E and Travis WD: The
2004 World Health Organization classification of lung tumors. Semin
Roentgenol. 40:90–97. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Venegas V and Halberg MC: Quantification
of mtDNA mutation heteroplasmy (ARMS qPCR). Methods Mol Biol.
837:313–326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kulawiec M, Owens KM and Singh KK: mtDNA
G10398A variant in African-American women with breast cancer
provides resistance to apoptosis and promotes metastasis in mice. J
Hum Genet. 54:647–654. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Canter JA, Kallianpur AR, Parl FF and
Millikan RC: Mitochondrial DNA G10398A polymorphism and invasive
breast cancer in African-American women. Cancer Res. 65:8028–8033.
2005.PubMed/NCBI
|
|
21
|
Lee HC, Chang CM and Chi CW: Somatic
mutations of mitochondrial DNA in aging and cancer progression.
Ageing Res Rev. 9(Suppl 1): S47–S58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu H, He W, Jiang HG, Zhao H, Peng XH, Wei
YH, Wei JN, Xie CH, Liang C, Zhong YH, et al: Prognostic value of
mitochondrial DNA content and G10398A polymorphism in non-small
cell lung cancer. Oncol Rep. 30:3006–3012. 2013.PubMed/NCBI
|
|
23
|
Wong LJ and Bai RK: Real-time quantitative
polymerase chain reaction analysis of mitochondrial DNA point
mutation. Methods Mol Biol. 335:187–200. 2006.PubMed/NCBI
|
|
24
|
Zhu Y, Huang S, Kang D, Han M, Wang G,
Yuan Y, Su Y, Yuan H, Zhai S and Dai P: Analysis of the
heteroplasmy level and transmitted features in hearing-loss
pedigrees with mitochondrial 12S rRNA A1555G mutation. BMC Genet.
15:262014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Warowicka A, Kwasniewska A and
Gozdzicka-Jozefiak A: Alterations in mtDNA: A qualitative and
quantitative study associated with cervical cancer development.
Gynecol Oncol. 129:193–198. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sobenin IA, Mitrofanov KY, Zhelankin AV,
Sazonova MA, Postnov AY, Revin VV, Bobryshev YV and Orekhov AN:
Quantitative assessment of heteroplasmy of mitochondrial genome:
Perspectives in diagnostics and methodological pitfalls. Biomed Res
Int. 2014:2920172014. View Article : Google Scholar : PubMed/NCBI
|