Introduction
Acute myeloid leukemia (AML) is composed of a group
of diseases with marked morphological and cytogenetic heterogeneity
that account for ~20% of childhood and adolescent acute leukemias
(1,2).
Acute promyelocytic leukemia (APL) is considered a distinct subtype
of AML, with a particular cytological morphology (M and M3 variant
in the French-American-British classification) (3). APL is characterized by a specific
chromosome translocation, t(15;17), which results in the
rearrangement of the promyelocytic leukemia (PML) gene and the
retinoic acid receptor α (RARα) gene, followed by the expression of
the PML-RARα chimeric protein (4).
Despite the fact that majority of APL cases can achieve complete
remission when treated with conventional chemotherapy by all-trans
retinoic acid (ATRA) and/or arsenic trioxide
(As2O3) (5–8), certain
clinical cases exhibit acquired resistance and APL relapse
(5,6,8), and the
detailed molecular mechanisms have not been completely
elucidated.
The small ubiquitin-like modifier
(SUMO)/sentrin-specific protease (SENP) family includes six SENPs
named SENP1, SENP2, SENP3, SENP5, SENP6 and SENP7, which have
different substrate specificities and subcellular localizations in
mammalian cells (9). It has been
reported that SENP1 is important in placental development and
erythropoiesis (10,11). In addition, SENP1 could control
adipocyte differentiation via de-SUMOylation modification (12), and is important in protecting against
myocardial ischemia/reperfusion injury via a hypoxia-inducible
factor 1α-dependent pathway (13).
Numerous studies have demonstrated that SENP1 is overexpressed and
contributes significantly to the development and progression of
tumors, including prostatic intraepithelial neoplasia, colon cancer
and pancreatic cancer (14–16). Previous studies have revealed that
deficiency or downregulation of SENP1 could induce
radiosensitization in lung cancer cells (17), promote endoplasmic reticulum (ER)
stress-induced apoptosis in MEF and HEK 293T cells (18), and increase apoptosis in Burkitt
lymphoma cells (19). Furthermore,
SENP1 is also involved in the formation of PML protein nuclear
bodies (20,21), which were originally characterized as
part of a fusion protein with PML-RARα in patients with APL
(22). However, whether SENP1 is a
potential drug target for APL treatment still remains unclear.
ER stress has been implicated in diverse diseases,
including cancer, diabetes, cerebral ischemia, and
neurodegenerative and cardiovascular diseases (23,24). In
addition to the intrinsic and extrinsic apoptosis pathways, ER
stress-induced apoptosis has recently been reported as another
major pathway mediating cell apoptosis (25), and is involved in arsenic-induced
apoptosis of osteoblasts, myoblasts, pancreatic cells, myeloma
cells, and drug-sensitive and drug-resistant leukemia cells
(26–30). It has been suggested that
As2O3 can induce human lens cell apoptosis by
the ER stress-initiated process (31).
In the present study, the NB4 cell line (an APL cell
line from a relapsed APL patient) was utilized as an in
vitro model (32). It was
demonstrated that As2O3 could induce ER
stress-initiated apoptosis in NB4 cells, which was significantly
upregulated by SENP1 knockdown. In addition, it was observed that
SENP1 specifically de-SUMOylated X-box binding protein 1 (XBP1) and
played a critical role during As2O3-induced
ER stress. Taken together, our results revealed the roles of SENP1
in APL and the potential effects of clinical APL treatment by
targeting SENP1.
Materials and methods
Antibodies and reagents
RPMI-1640 medium, Dulbecco's modified Eagle medium,
trypsin and TRIzol were obtained from Invitrogen (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Puromycin and fetal bovine
serum (FBS) were purchased from Gibco (Thermo Fisher Scientific,
Inc.). As2O3 was provided by Beijing Shuanglu
Pharmaceutical Co., Ltd. (Beijing, China). Radioimmunoprecipitation
assay lysates were purchased from Beyotime Institute of
Biotechnology (Haimen, China). ATRA, phenylmethylsulfonyl fluoride,
aprotinin, leupeptin and pepstatin were acquired from Sigma-Aldrich
(Merck Millipore, Darmstadt, Germany). Protein A/G PLUS-Agarose was
obtained from Roche Diagnostics (Indianapolis, IN, USA). Anti-XBP1
antibody (cat. no. H00007494-D01) and anti-SUMO-1 antibody (cat.
no. AJ1746a) were purchased from Abnova (Taipei City, Taiwan) and
Abgent Biotech Co., Ltd. (Suzhou, China), respectively. The FITC
Annexin V Apoptosis Detection kit and the anti-cluster of
differentiation (CD) 11b antibody (cat. no. C09-550019) were
commercially available from BD Pharmingen (San Diego, CA, USA). The
PrimeScript RT reagent kit and SYBR Green PCR Master Mix were
commercially available from Takara Bio, Inc. (Otsu, Japan).
Cell culture
Human APL NB4 cells (American Type Culture
Collection, Manassas, VA, USA) were suspended at 5×105
cells/ml in RPMI-1640 medium supplemented with 10% FBS. Retrovirus
containing SENP1 small interfering RNA (siRNA) or nonspecific
control (NC) siRNA, as described previously (10), was transfected into NB4 cells to
generate si-SENP1-transfected NB4 cells (si-SENP1) or NC
siRNA-transfected cells (si-NC) upon puromycin (0.75 µg/ml)
selection. All cells were cultured in RPMI-1640 medium with 10% FBS
at 37°C in 5% CO2.
As2O3 and ATRA
treatment
NB4 cells were seeded into a 6-well plate and then
incubated with As2O3 (1 µM) or ATRA (1 µM)
for different time periods, as indicated. A total of 106
cells were harvested at different time points subsequent to
As2O3 or ATRA treatment.
Flow cytometry
For apoptosis assay, cells treated with
As2O3 at each indicated time point were
washed twice with ice-cold phosphate-buffered saline, and the
apoptotic cells were detected with a flow cytometer (Merck
Millipore) using the FITC Annexin V Apoptosis Detection kit
according to the manufacturer's protocol. For CD11b assay, cells
were incubated with fluorescein isothiocyanate (FITC)-conjugated
anti-CD11b antibody (1:100) after treatment with ATRA for 24 h.
Appropriate isotype control IgG2b antibodies (cat. no. NB810-82278;
Novus Biologicals LLC, Littleton, CO, USA) were used. The
percentage of apoptotic cells and the differentiation marker of
cell surface expression were analyzed using Guava 1.0 software
(Guava Technologies, Inc., Hayward, CA, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated with TRIzol, and
complementary DNA was synthesized using TaKaRa RNA PCR kit (Takara
Bio, Inc.) according to the manufacturer's protocol. The sequences
of the PCR primers used in the amplification of the target genes
are shown in Table I. RT-qPCR was
performed with the double-stranded DNA dye SYBR Green PCR core
reagents using the ABI ViiA 7 system (PerkinElmer, Inc., Waltham,
MA, USA). Thermal cycler conditions were 95°C for 30 sec, and 40
cycles of 95°C for 5 sec and 60°C for 30 sec. Relative gene
expression was determined by the delta delta Cq method (Applied
Biosystems; Thermo Fisher Scientific, Inc.), with β-actin as the
endogenous control using the Applied Biosystems ViiA™ 7 Real-Time
PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
(16). Three independent experiments
were performed, and the standard deviations (SDs) representing
experimental errors were calculated. All data were analyzed using
GraphPad Prism 5 software (GraphPad Software, Inc., La Jolla, CA,
USA).
 | Table I.Sequences of the primers used in the
amplification of the target genes. |
Table I.
Sequences of the primers used in the
amplification of the target genes.
| Locus | Primers
(5′-3′) |
|---|
| β-actin | (F)
CTTTTCCAGCCTTCCTTCTTGG |
|
| (R)
CAGCACTGTGTTGGCATAGAGG |
| SENP1 | (F)
ATCAGGCAGTGAAACGTTGGAC |
|
| (R)
ATCAGGCAGTGAAACGTTGGAC |
| GADD153 | (F)
GAAACGGAAACAGAGTGGTCATTCCCC |
|
| (R)
GTGGGATTGAGGGTCACATCATTGGCA |
| ATF6 | (F)
ATGAAGTTGTGTCAGAGAAC |
|
| (R)
GGGTGCTATTGTAATGACTCA |
| GRP78 | (F)
AGTTGATATTGGAGGTGGGC |
|
| (R)
CATTGAAGTAAGCTGGTACAGTAAC |
| Erdj4 | (F)
CCCCAGTGTCAAACTGTACCAG |
|
| (R)
AGCGTTTCCAATTTTCCATAAATT |
| Sec61a | (F)
CTATTTCCAGGGCTTCCGAGT |
|
| (R)
AGGTGTTGTACTGGCCTCGGT |
| Edem | (F)
AAGCCCTCTGGAACTTGCG |
|
| (R)
AACCCAATGGCCTGTCTGG |
Immunoprecipitation
Treated cells were lysed with lysis buffer, and
immunoprecipitation using anti-XBP1 and anti-SUMO-1 antibodies was
performed as previously reported (33,34).
Briefly, cell lysates were centrifuged at 10,800 × g for 15
min at 4ºC, and the supernatant was collected. Protein
concentrations were determined by means of Lowry protein assay.
Equal amounts of protein samples (1 mg) were incubated with 2 µg
antibody for 3 h at 4ºC, followed by addition of
pre-equilibrated Protein A/G PLUS-Agarose beads (20 µl) and
incubation overnight. The immunoprecipitates were washed four times
with lysis buffer, and the bound proteins were boiled in 15 µl 2X
sodium dodecyl sulfate (SDS) sample buffer, eluted and resolved on
10% SDS-polyacrylamide gel electrophoresis. The proteins were
transferred onto a polyvinylidene fluoride membrane and probed with
horseradish peroxidase (HRP)-phytohemagglutinin-L (Thermo Fisher
Scientific, Inc.) and HRP-concanavalin A lectin (Abnova), and then
detected using an enhanced chemiluminescence kit (Pierce
Biotechnology, Inc., Rockford, IL, USA).
Statistical analyses
The data were presented as the mean ± SD.
Statistical analysis was performed using GraphPad Prism 5 software.
The paired t-test was used to compare the difference between
two different groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
SENP1 deficiency has no effect on the
spontaneous apoptosis or differentiation of NB4 cells
The fact that SENP1 was overexpressed in several
neoplastic tissues (14–16) raised the question whether SENP1 plays
any role in the pathogenesis of APL. Thus, a SENP1-knockdown NB4
cell line (si-SENP1) was generated by stably transfecting SENP1
siRNA into NB4 cells. Compared with the NB4 cells transfected with
NC scrambled siRNA (si-NC), the messenger RNA (mRNA) expression of
SENP1 was significantly downregulated to ~45% in si-SENP1 NB4 cells
(Fig. 1A). A series of flow cytometry
assays were performed to explore whether SENP1 deficiency would
promote spontaneous apoptosis and differentiation in NB4 cells.
Firstly, the percentage of apoptotic si-NC and si-SENP1 NB4 cells
was examined by flow cytometry following annexin V-FITC and
propidium iodide (PI) staining. As shown in Fig. 1B, knockdown of SENP1 had no effect on
the spontaneous apoptosis of NB4 cells compared with si-NC cells.
Next, the effects of SENP1 deficiency on NB4 cell differentiation
were examined by detecting the expression levels of a
differentiation marker, CD11b. The results from cytometry revealed
no difference in the expression of CD11b between si-NC and si-SENP1
NB4 cells, even after treatment with 1 µΜ ATRA (a differentiation
inducing agent) for 24 h (Fig. 1C).
These results suggested that the downregulation of SENP1 has no
effect on the spontaneous apoptosis or differentiation of NB4
cells.
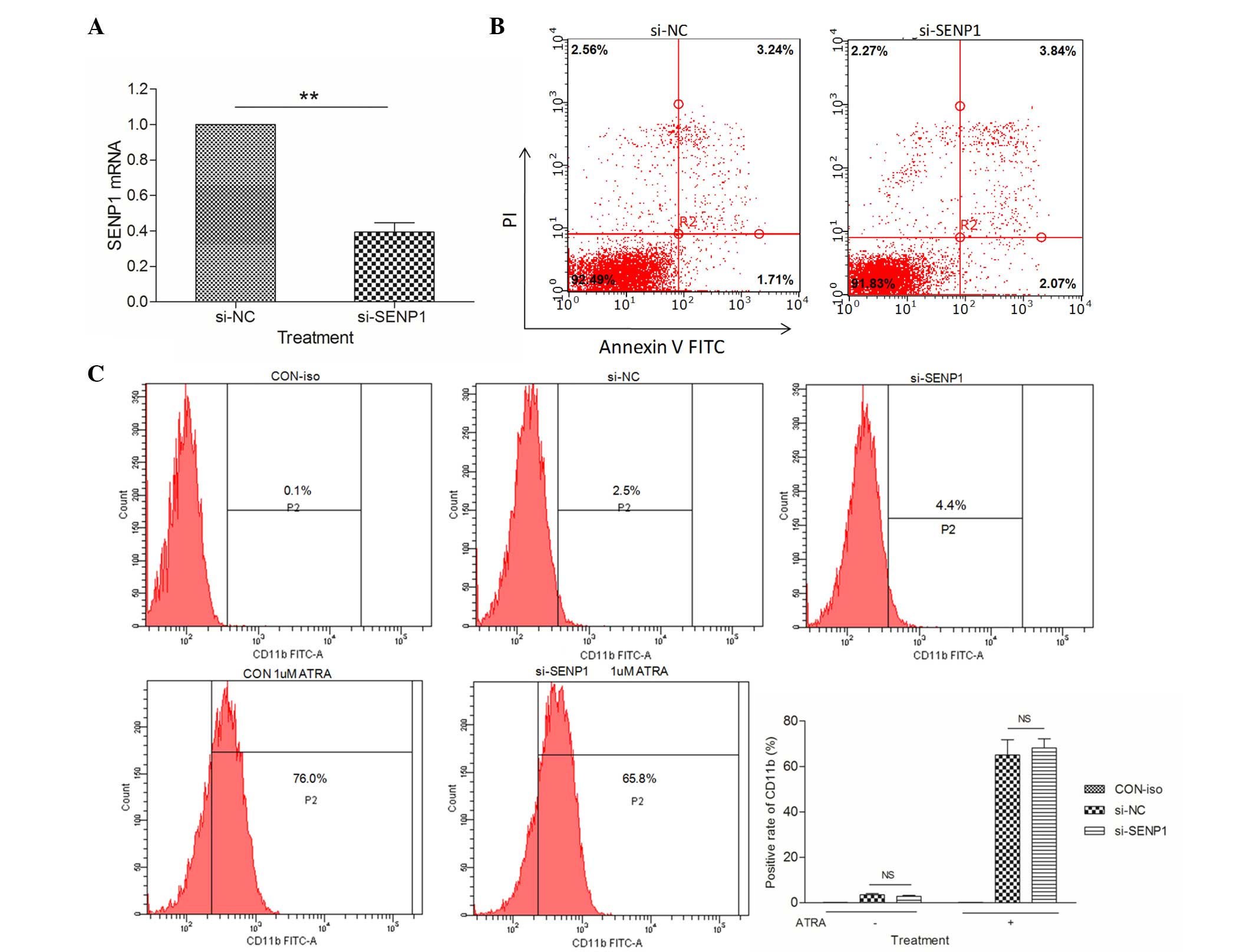 | Figure 1.SENP1 deficiency has no effect on the
spontaneous apoptosis or differentiation of NB4 cells. (A) mRNA
expression level of SENP1 in cells transfected with si-SENP1 and
cells transfected with si-NC. (B) Spontaneous apoptosis of NB4
cells transfected with si-SENP1 and si-NC. (C) Positive expression
rate of CD11b in NB4 cells induced by ATRA for 72 h. **P<0.01,
t-test. SENP1, small ubiquitin-like
modifier/sentrin-specific protease 1; mRNA, messenger RNA; NC,
nonspecific control; si, small interfering; PI, propidium iodide;
FITC, fluorescein isothiocyanate; CD, cluster of differentiation;
ATRA, all-trans retinoic acid; CON, control; iso, isotype; NS, not
significant. |
SENP1 deficiency promotes
As2O3-induced NB4 cell apoptosis
As2O3 has been successfully
used in the treatment of APL (7,35,36), and therapeutic doses of
As2O3 could effectively induce complete
molecular remission in vivo and trigger the apoptotic death
of APL cells (37). Thus, we sought
to investigate if the knockdown of SENP1 affects
As2O3-induced apoptosis in NB4 cells.
Therefore, normal and SENP1 knocked down NB4 cells treated with
As2O3 were collected and subjected to flow
cytometry assay upon annexin V-FITC and PI staining. As shown in
Fig. 2A, after treatment with
As2O3 for 3 days, the apoptotic cells in the
si-NC NB4 group only increased by 2.90-fold (from 12.92±1.00 to
37.00±2.00%), while in the si-SENP1 NB4 group, the apoptotic cells
increased by ≤4.00-fold (from 16.62±1.00 to 66.00±3.00%). The total
number of apoptotic cells was ~30% in si-SENP1 cells compared with
that in si-NC cells (Fig. 2B),
suggesting that SENP1 is essential for NB4 cell survival during
treatment with As2O3.
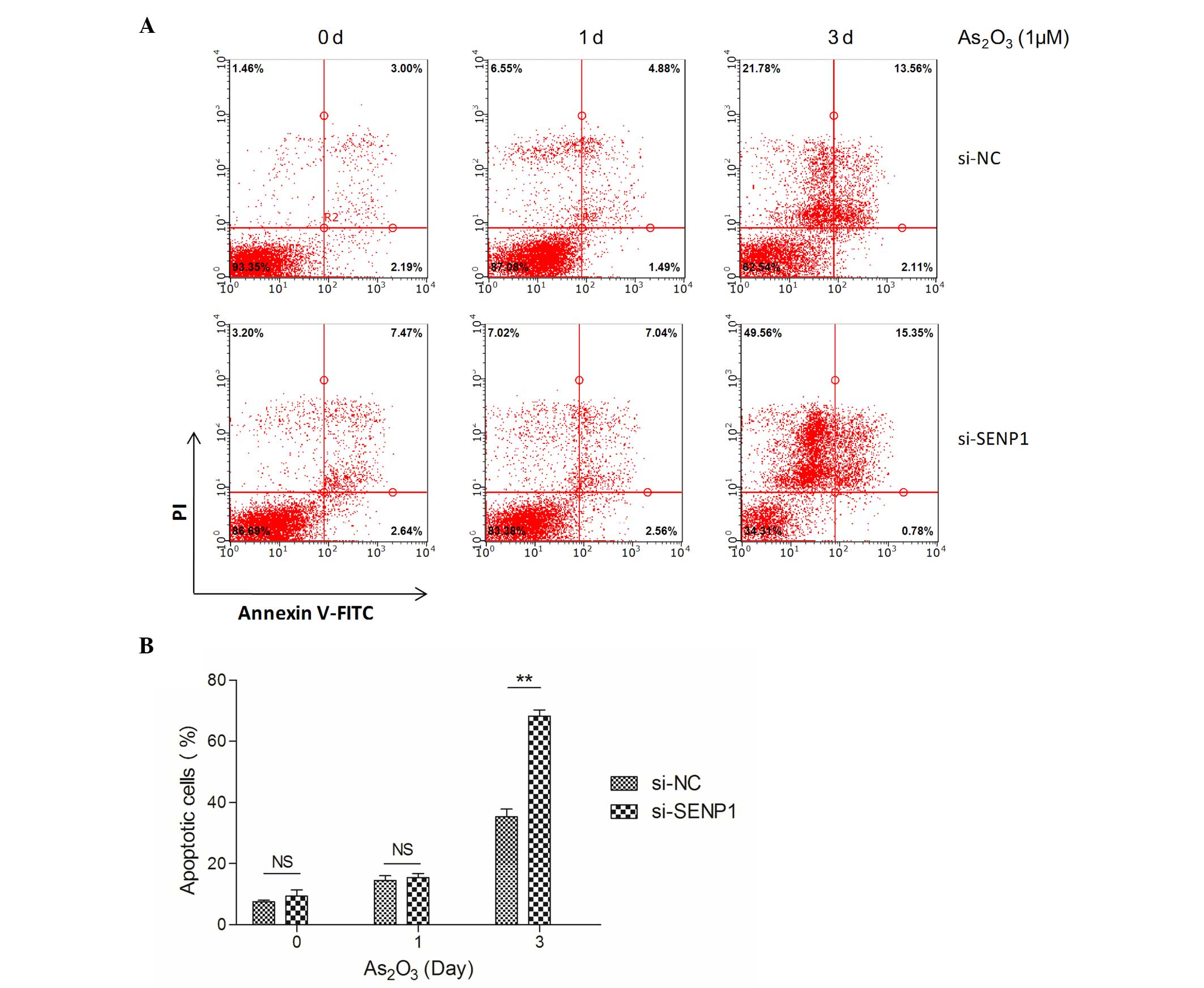 | Figure 2.SENP1 deficiency promotes
As2O3-induced NB4 cell apoptosis. (A)
Apoptosis of the si-NC and si-SENP1 NB4 cells was analyzed by flow
cytometry. (B) The number of apoptotic si-SENP1 cells was
significantly higher than that of the si-NC cells, following
treatment with As2O3 for 3 d.**P<0.01,
t-test. SENP1, small ubiquitin-like
modifier/sentrin-specific protease 1; NC, nonspecific control; si,
small interfering; PI, propidium iodide; FITC, fluorescein
isothiocyanate; NS, not significant; As2O3,
arsenic trioxide; d, day. |
SENP1 regulates the expression of XBP1
target genes in the ER stress-mediated apoptotic pathway
Numerous studies have revealed that
As2O3 induces cell apoptosis through the ER
stress pathway (28,29,31). Thus,
we assumed that As2O3-induced apoptosis may
also accompany ER stress. Several key players, including
CCAAT-enhancer-binding protein homologous protein [also known as
growth arrest and DNA damage inducible protein 153 (GADD153)],
activating transcription factor 6 (ATF6) and 78 kDa
glucose-regulated protein (GRP78), as well as XBP1 target genes,
including ER-localized DnaJ 4 (Erdj4), Sec61a and ER degradation
enhancer, mannosidase (Edem), are common markers that reflect the
unfolded protein response during ER stress (38,39).
Therefore, to investigate the effects of SENP1 on the ER
stress-mediated apoptotic pathway, the mRNA expression levels of
the above proteins were detected by RT-qPCR in si-NC and si-SENP1
NB4 cells subsequent to treatment with As2O3
for different time periods. As shown in Fig. 3, the mRNA expression levels of these
markers were highly increased by ~4.00-fold (GRP78), 1.77-fold
(GADD153), 1.80-fold (Erdj4) and 1.80-fold (Sec61a) in si-NC cells
after As2O3 treatment for 1 day, but no
effect on ATF6 or Edem (Fig. 3C and
F) were observed, and upregulation was also observed in
si-SENP1 NB4 cells, which revealed that As2O3
induced NB4 cell apoptosis through the ER stress-mediated apoptotic
pathway. Additionally, among the XBP1 target genes, the mRNA
expression levels of Erdj4 and Sec61a were significantly reduced by
knockdown of SENP1 following As2O3 treatment
in si-SENP1 NB4 cells (Fig. 3B and
D).
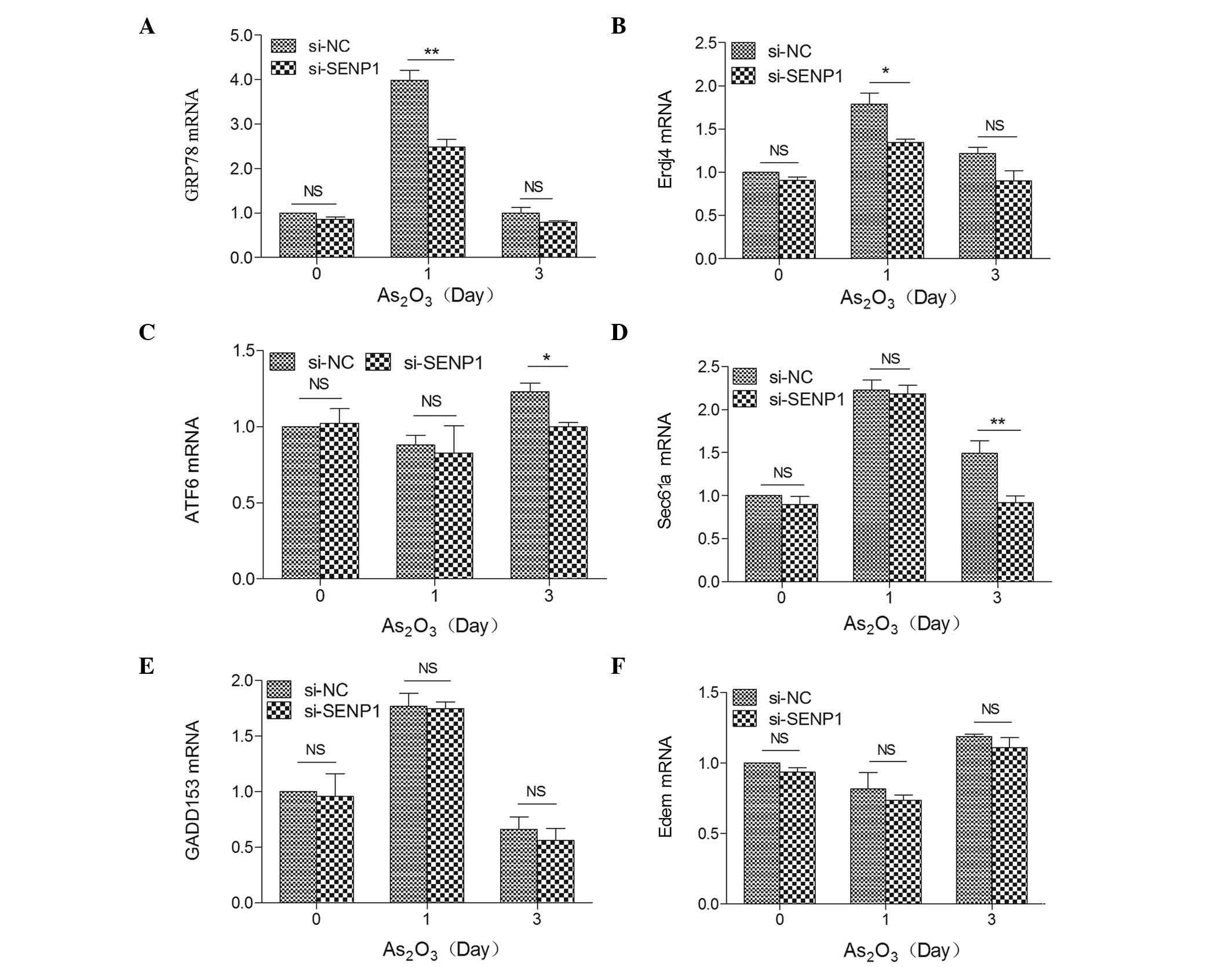 | Figure 3.Expression levels of ER
stress-related key markers (A) GRP78, (B) Erdj4, (C) ATF6, (D)
Sec61a, (E) GADD153 and (F) Edem in SENP1-deficient cells treated
with As2O3. **P<0.01; *P<0.05,
t-test. SENP1, small ubiquitin-like
modifier/sentrin-specific protease 1; GADD153, growth arrest and
DNA damage inducible protein 153; ATF6, activating transcription
factor 6; GRP78, 78 kDa glucose-regulated protein; ErdJ4,
ER-localized DnaJ 4; Edem, ER degradation enhancer, mannosidase;
NC, nonspecific control; si, small interfering; NS, not
significant; As2O3, arsenic trioxide; mRNA,
messenger RNA; ER, endoplasmic reticulum. |
SENP1 deficiency upregulates XBP1
SUMOylation in si-SENP1 NB4 cells
Since the mRNA expression levels of XBP1 target
genes, including Erdj4 and Sec61a, were significantly reduced in
si-SENP1 NB4 cells upon As2O3 treatment, the
present study next sought to explore whether XBP1 could be
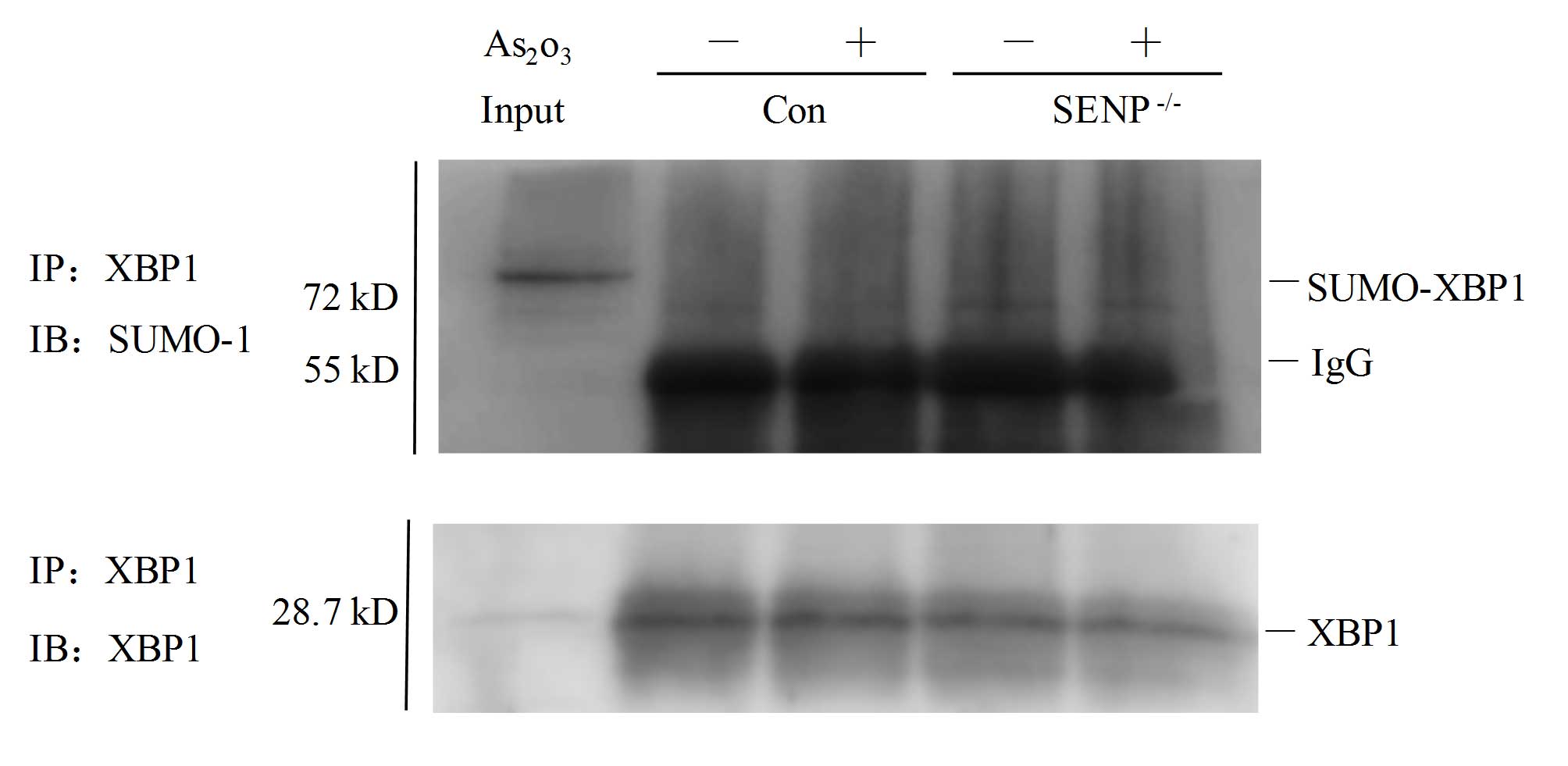
SUMOylated in si-SENP1 NB4 cells. As shown in Fig. 4, in contrast to si-NC cells (Fig. 4, lanes 2 and 3), XBP1 was partly
SUMOylated following immunoprecipitation with an anti-SUMO-1
antibody in si-SENP1 NB4 cells (Fig.
4, lanes 4 and 5). The input sample for immunoprecipitation was
directly loaded to reveal the appropriate location of the protein
bands of interest (Fig. 4, lane 1).
These findings suggested that SENP1 deficiency may downregulate
XBP1 transactivation by increasing XBP1 SUMOylation.
Discussion
APL is characterized by a specific t(15;17)
chromosomal translocation that yields the PML/RARA fusion gene
(3). It has been reported that
>97% of APL patients have the t(15;17) translocation, and
several rare variant translocations observed in the remaining APL
patients always involve RARα (3,40).
As2O3 is an effective treatment for this
disease, since it could induce SUMO-dependent ubiquitin-mediated
proteasomal degradation of PML-RARα (41). However, knockdown of SENP1 had no
effect on the SUMOylation of PML-RARα in NB4 cells, even after NB4
cells were treated with 1 µΜ As2O3 (data not
shown), suggesting the possibility that other members of the SENP
family may be involved in regulating the level of SUMOylated
PML-RARα, which remains to be further investigated. It has been
reported that knockdown of SENP1 could promote drug-induced cell
apoptosis in MEF and HEK 293T cells, as well as lung cancer and
Burkitt lymphoma cells (17–19). However, our results revealed that
downregulation of SENP1 had no effect on the spontaneous apoptosis
or differentiation of NB4 cells (Fig.
1). Furthermore, it was observed that downregulation of SENP1
had no impact on drug-induced apoptosis or differentiation in NB4
cells (Fig. 1).
The metabolism of As2O3 in APL
cells is complex (42). Production of
intracellular methylated metabolites and epigenetic changes of DNA
methylation during As2O3 metabolism may
contribute to the therapeutic efficacy of
As2O3 in APL (43). The signaling cascades and
transcription factors related to
As2O3-induced apoptosis were caused by
reactive oxygen species formation (44,45),
resulting in the stimulation of apoptosis in leukemia cells. In
contrast to the observation that inhibition of phosphoinositide
3-kinase (PI3K)/Akt could increase
As2O3-induced apoptosis of APL cells
(46,47), in the present study, it was observed
that downregulation of SENP1 could promote
As2O3-induced NB4 cell apoptosis, suggesting
that there may be pathways other than PI3K/Akt contributing to the
effect of SENP1 on As2O3-treated APL cells.
Numerous studies have demonstrated that As2O3
could induce cell apoptosis through the ER stress pathway (28,29,31,48).
Consistently, in our study, the upregulation of key markers of the
ER stress-mediated apoptotic pathway in si-SENP1 NB4 cells
indicated that As2O3 could induce NB4 cell
apoptosis through the ER stress-mediated apoptotic pathway.
Notably, the high ER stress induced by As2O3
treatment at day 1 in the present study led to significant cell
apoptosis at day 3, which may be explained by the sustained effect
of As2O3-induced apoptosis. SUMOylation is a
dynamic process that is reversed by proteins of the SENP family,
which can be catalyzed by SUMO-specific activating (E1),
conjugating (E2) and ligating (E3) enzymes (49). The modification of proteins with SUMO
plays pivotal roles in modulating the activation, function and
subcellular localization of these proteins (50). The transcription factor XBP1 is a key
component of the ER stress response and a critical transcription
factor that drives the transcription of various genes (51,52).
Additionally, XBP1 can be activated in MEF and HEK 293T cells via
the de-SUMOylation function of SENP1 (18). Therefore, it would be reasonable to
assume the existence of an intrinsic mechanism by which the
downregulation of SENP1 affects cell apoptosis in the treatment of
APL with As2O3. Our data suggested that SENP1
regulated XBP1 transactivation by regulating the expression of XBP1
target genes, and that SENP1 may play a specific role in regulating
XBP1 activity during As2O3-induced apoptosis.
Of note, SENP1 downregulation also reduced the expression of the
transcription factor GRP78, which was involved in the ER stress
response after As2O3 treatment for 1 day
(Fig. 3C), suggesting that SENP1 and
its downstream signaling pathway may affect other transcription
factors involved in ER stress.
Taken together, our study provided the first
evidence for As2O3-induced NB4 cell apoptosis
through the ER stress-mediated apoptotic pathway, and demonstrated
that SENP1 downregulation promoted
As2O3-induced NB4 cell apoptosis. The
knockdown of SENP1 also increased the levels of accumulated
SUMOylated XBP1, which was accompanied by downregulation of the
mRNA expression and transcriptional activity of XBP1 target genes,
which were involved in ER stress response and closely linked to the
apoptosis of NB4 cells. Our data also provided insight into
potential therapeutic targets for APL treatment via manipulating
XBP1 signaling during ER stress by targeting SENP1.
Acknowledgements
The present study was supported by Shanghai Health
Bureau of Scientific Research Projects (Shanghai, China; grant no.
20124Y110), the Scientific Research Foundation of XinHua Hospital
Group (Shanghai, China; grant no. 12XJ22005) and the National
Natural Science Foundation of China (Beijing, China; grant nos.
81401572 and 81201450, awarded to Q.C.).
References
|
1
|
Kömür M, Erbey F, Bayram I and Tanyeli A:
Incidence and prognostic importance of molecular genetic defects in
children with acute myeloblastic leukemia. Asian Pac J Cancer Prev.
11:1393–1395. 2010.PubMed/NCBI
|
|
2
|
Gilliland DG: Molecular genetics of human
leukemia. Leukemia. 12(Suppl 1): S7–S12. 1998.PubMed/NCBI
|
|
3
|
Rowley JD, Golomb HM and Dougherty C:
15–17 translocation, a consistent chromosome change in acute
promyelocytic leukemia. Lancet. 1:549–50. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Melnick A and Licht JD: Deconstructing a
disease: RARalpha, its fusion partners and their roles in the
pathogenesis of acute promyelocytic leukemia. Blood. 93:3167–3215.
1999.PubMed/NCBI
|
|
5
|
Burnett AK, Grimwade D, Solomon E,
Wheatley K and Goldstone AH: Presenting white blood cell count and
kinetics of molecular remission predict prognosis in acute
promyelocytic leukemia treated with all-trans retinoic acid: Result
of the Randomized MRC Trial. Blood. 93:4131–4143. 1999.PubMed/NCBI
|
|
6
|
Fenaux P, Chastang C, Chevret S, Sanz M,
Dombret H, Archimbaud E, Fey M, Rayon C, Huguet F, Sotto JJ, et al:
A randomized comparison of all transretinoic acid (ATRA) followed
by chemotherapy and ATRA plus chemotherapy and the role of
maintenance therapy in newly diagnosed acute promyelocytic
leukemia. The European APL Group. Blood. 94:1192–1200.
1999.PubMed/NCBI
|
|
7
|
Chen GQ, Zhu J, Shi XG, Ni JH, Zhong HJ,
Si GY, Jin XL, Tang W, Li XS, Xong SM, et al: In vitro studies on
cellular and molecular mechanisms of arsenic trioxide
(As2O3) in the treatment of acute
promyelocytic leukemia: As2O3 induces NB4 cell apoptosis with
downregulation of Bcl-2 expression and modulation of PML-RAR
alpha/PML proteins. Blood. 88:1052–1061. 1996.PubMed/NCBI
|
|
8
|
Lam MS and Ignoffo RJ: Arsenic trioxide
for the treatment of acute promyelocytic leukemia. Cancer Pract.
9:155–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bawa-Khalfe T and Yeh ET: SUMO losing
balance: SUMO proteases disrupt SUMO homeostasis to facilitate
cancer development and progression. Genes Cancer. 1:748–752. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheng J, Kang X, Zhang S and Yeh ET:
SUMO-specific protease 1 is essential for stabilization of
HIF1alpha during hypoxia. Cell. 131:584–595. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu L, Ji W, Zhang H, Renda MJ, He Y, Lin
S, Cheng EC, Chen H, Krause DS and Min W: SENP1-mediated GATA1
deSUMOylation is critical for definitive erythropoiesis. J Exp Med.
207:1183–1195. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu B, Wang T, Mei W, Li D, Cai R, Zuo Y
and Cheng JK: Small ubiquitin-like modifier (SUMO) protein-specific
protease 1 de-SUMOylates Sharp-1 protein and controls adipocyte
differentiation. J Biol Chem. 289:22358–22364. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gu J, Fan Y, Liu X, Zhou L, Cheng J, Cai R
and Xue S: SENP1 protects against myocardial ischaemia/reperfusion
injury via a HIF1α-dependent pathway. Cardiovasc Res. 104:83–92.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bawa-Khalfe T, Cheng J, Lin SH, Ittmann MM
and Yeh ET: SENP1 induces prostatic intraepithelial neoplasia
through multiple mechanisms. J Biol Chem. 285:25859–25866. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu Y, Li J, Zuo Y, Deng J, Wang LS and
Cheng GQ: SUMO-specific protease 1 regulates the in vitro and in
vivo growth of colon cancer cells with the upregulated expression
of CDK inhibitors. Cancer Lett. 309:78–84. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma C, Wu B, Huang X, Yuan Z, Nong K, Dong
B, Bai Y, Zhu H, Wang W and Ai K: SUMO-specific protease 1
regulates pancreatic cancer cell proliferation and invasion by
targeting MMP-9. Tumour Biol. 35:12729–12735. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang RT, Zhi XY, Zhang Y and Zhang J:
Inhibition of SENP1 induces radiosensitization in lung cancer
cells. Exp Ther Med. 6:1054–1058. 2013.PubMed/NCBI
|
|
18
|
Jiang Z, Fan Q, Zhang Z, Zou Y, Cai R,
Wang Q, Zuo Y and Cheng J: SENP1 deficiency promotes ER
stress-induced apoptosis by increasing XBP1 SUMOylation. Cell
Cycle. 11:1118–1122. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang BB, Gao QM, Liang W, Xiu B, Zhang WJ
and Liang AB: Downregulation of SENP1 expression increases
apoptosis of Burkitt lymphoma cells. Asian Pac J Cancer Prev.
13:2045–2049. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sharma P, Murillas R, Zhang H and Kuehn
MR: N4BP1 is a newly identified nucleolar protein that undergoes
SUMO-regulated polyubiquitylation and proteasomal turnover at
promyelocytic leukemia nuclear bodies. J Cell Sci. 123:1227–1234.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ohbayashi N, Kawakami S, Muromoto R, Togi
S, Ikeda O, Kamitani S, Sekine Y, Honjoh T and Matsuda T: The IL-6
family of cytokines modulates STAT3 activation by desumoylation of
PML through SENP1 induction. Biochem Biophys Res Commun.
371:823–828. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kastner P, Perez A, Lutz Y, Rochette-Egly
C, Gaub MP, Durand B, Lanotte M, Berger R and Chambon P: Structure,
localization and transcriptional properties of two classes of
retinoic acid receptor alpha fusion proteins in acute promyelocytic
leukemia (APL): Structural similarities with a new family of
oncoproteins. EMBO J. 11:629–642. 1992.PubMed/NCBI
|
|
23
|
Mollereau B, Manié S and Napoletano F:
Getting the better of ER stress. J Cell Commun Signal. 8:311–321.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gorman AM, Healy SJ, Jäger R and Samali A:
Stress management at the ER: Regulators of ER stress-induced
apoptosis. Pharmacol Ther. 134:306–316. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mathur A, Abd Elmageed ZY, Liu X,
Kostochka ML, Zhang H, Abdel-Mageed AB and Mondal D: Subverting
ER-stress towards apoptosis by nelfinavir and curcumin coexposure
augments docetaxel efficacy in castration resistant prostate cancer
cells. PLoS One. 9:e1031092014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kondo S, Hino SI, Saito A, Kanemoto S,
Kawasaki N, Asada R, Izumi S, Iwamoto H, Oki M, Miyagi H, et al:
Activation of OASIS family, ER stress transducers, is dependent on
its stabilization. Cell Death Differ. 19:1939–1949. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qin L, Wang Z, Tao L and Wang Y: ER stress
negatively regulates AKT/TSC/mTOR pathway to enhance autophagy.
Autophagy. 6:239–247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chiu HW, Tseng YC, Hsu YH, Lin YF, Foo NP,
Guo HR and Wang YJ: Arsenic trioxide induces programmed cell death
through stimulation of ER stress and inhibition of the
ubiquitin-proteasome system in human sarcoma cells. Cancer Lett.
356:762–772. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li K, Zhang L, Xiang X, Gong S, Ma L, Xu
L, Wang G, Liu Y, Ji X, Liu S, et al: Arsenic trioxide alleviates
airway hyperresponsiveness and promotes apoptosis of
CD4+ T lymphocytes: Evidence for involvement of the ER
stress-CHOP pathway. Ir J Med Sci. 182:573–583. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Doudican NA, Wen SY, Mazumder A and Orlow
SJ: Sulforaphane synergistically enhances the cytotoxicity of
arsenic trioxide in multiple myeloma cells via stress-mediated
pathways. Oncol Rep. 28:1851–1858. 2012.PubMed/NCBI
|
|
31
|
Zhang H, Duncan G, Wang L, Liu P, Cui H,
Reddan JR, Yang BF and Wormstone IM: Arsenic trioxide initiates ER
stress responses, perturbs calcium signalling and promotes
apoptosis in human lens epithelial cells. Exp Eye Res. 85:825–835.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fan YH, Chen M, Meng J, Yu L, Tu Y, Wan L,
Fang K and Zhu W: Arsenic trioxide and resveratrol show synergistic
anti-leukemia activity and neutralized cardiotoxicity. PLoS One.
9:e1058902014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu M, Yang Y, Wang C, Sun L, Mei C, Yao
W, Liu Y, Shi Y, Qiu S, Fan J, et al: The effect of epidermal
growth factor receptor variant III on glioma cell migration by
stimulating ERK phosphorylation through the focal adhesion kinase
signaling pathway. Arch Biochem Biophys. 502:89–95. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang C, Yang Y, Yang Z, Liu M, Li Z, Sun
L, Mei C, Chen H, Chen L, Wang L and Zha X: EGF-mediated migration
signaling activated by N-acetylglucosaminyltransferase-V via
receptor protein tyrosine phosphatase kappa. Arch Biochem Biophys.
486:64–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ghaffari SH, Bashash D, Dizaji MZ,
Ghavamzadeh A and Alimoghaddam K: Alteration in miRNA gene
expression pattern in acute promyelocytic leukemia cell induced by
arsenic trioxide: A possible mechanism to explain arsenic
multi-target action. Tumour Biol. 33:157–172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ghaffari SH, Momeny M, Bashash D, Mirzaei
R, Ghavamzadeh A and Alimoghaddam K: Cytotoxic effect of arsenic
trioxide on acute promyelocytic leukemia cells through suppression
of NFkβ-dependent induction of hTERT due to downregulation of Pin1
transcription. Hematology. 17:198–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao XY, Yang S, Chen YR, Li PC, Dou MM
and Zhang J: Resveratrol and arsenic trioxide act synergistically
to kill tumor cells in vitro and in vivo. PLoS One. 9:e989252014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
So AY, de la Fuente E, Walter P, Shuman M
and Bernales S: The unfolded protein response during prostate
cancer development. Cancer Metastasis Rev. 28:219–223. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yamamoto K, Tashiro E and Imoto M:
Quinotrierixin inhibited ER stress-induced XBP1 mRNA splicing
through inhibition of protein synthesis. Biosci Biotechnol Biochem.
75:284–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nasr R, Lallemand-Breitenbach V, Zhu J,
Guillemin MC and de Thé H: Therapy-induced PML/RARA proteolysis and
acute promyelocytic leukemia cure. Clin Cancer Res. 15:6321–6326.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Geoffroy MC, Jaffray EG, Walker KJ and Hay
RT: Arsenic-induced SUMO-dependent recruitment of RNF4 into PML
nuclear bodies. Mol Biol Cell. 21:4227–4239. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen GQ, Shi XG, Tang W, Xiong SM, Zhu J,
Cai X, Han ZG, Ni JH, Shi GY, Jia PM, et al: Use of arsenic
trioxide (As2O3) in the treatment of acute promyelocytic leukemia
(APL): I. As2O3 exerts dose-dependent dual effects on APL cells.
Blood. 89:3345–3353. 1997.PubMed/NCBI
|
|
43
|
Khaleghian A, Ghaffari SH, Ahmadian S,
Alimoghaddam K and Ghavamzadeh A: Metabolism of arsenic trioxide in
acute promyelocytic leukemia cells. J Cell Biochem. 115:1729–1739.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sumi D, Shinkai Y and Kumagai Y: Signal
transduction pathways and transcription factors triggered by
arsenic trioxide in leukemia cells. Toxicol Appl Pharmacol.
244:385–392. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang J, Li L, Cang H, Shi G and Yi J:
NADPH oxidase-derived reactive oxygen species are responsible for
the high susceptibility to arsenic cytotoxicity in acute
promyelocytic leukemia cells. Leuk Res. 32:429–436. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tabellini G, Cappellini A, Tazzari PL,
Falà F, Billi AM, Manzoli L, Cocco L and Martelli AM:
Phosphoinositide 3-kinase/Akt involvement in arsenic trioxide
resistance of human leukemia cells. J Cell Physiol. 202:623–634.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tabellini G, Tazzari PL, Bortul R,
Evangelisti C, Billi AM, Grafone T, Martinelli G, Baccarani M and
Martelli AM: Phosphoinositide 3-kinase/Akt inhibition increases
arsenic trioxide-induced apoptosis of acute promyelocytic and
T-cell leukaemias. Br J Haematol. 130:716–725. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Binet F, Chiasson S and Girard D: Evidence
that endoplasmic reticulum (ER) stress and caspase-4 activation
occur in human neutrophils. Biochem Biophys Res Commun. 391:18–23.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang Y and Dasso M: SUMOylation and
deSUMOylation at a glance. J Cell Sci. 122:4249–4252. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Verger A, Perdomo J and Crossley M:
Modification with SUMO. A role in transcriptional regulation. EMBO
Rep. 4:137–142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Quentin T, Steinmetz M, Poppe A and Thoms
S: Metformin differentially activates ER stress signaling pathways
without inducing apoptosis. Dis Model Mech. 5:259–269. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ibuki T, Yamasaki Y, Mizuguchi H and
Sokabe M: Protective effects of XBP1 against oxygen and glucose
deprivation/reoxygenation injury in rat primary hippocampal
neurons. Neurosci Lett. 518:45–48. 2012. View Article : Google Scholar : PubMed/NCBI
|