Introduction
Cholangiocarcinoma (CCA), the malignant tumor of
biliary epithelial cells, is the cancer of highest incidence in
northeastern Thailand (1), and has
increasing incidence and mortality worldwide (2,3). More than
70% of CCA patients have an advanced stage of the disease at the
time of diagnosis, which makes curative surgical resection
unfeasible (4). In these advanced CCA
patients, chemotherapy is the usual treatment option (5). The systemic chemotherapy choices
currently offered are 5-fluorouracil (5-FU), carboplatin plus 5-FU,
gemcitabine and paclitaxel, which have failed to improve survival,
and response rates are only ~20% (4).
New therapeutic agents for CCA are urgently required.
In the past few decades, natural bioactive
substances derived from plants have been considered as important
antitumor drug sources (6).
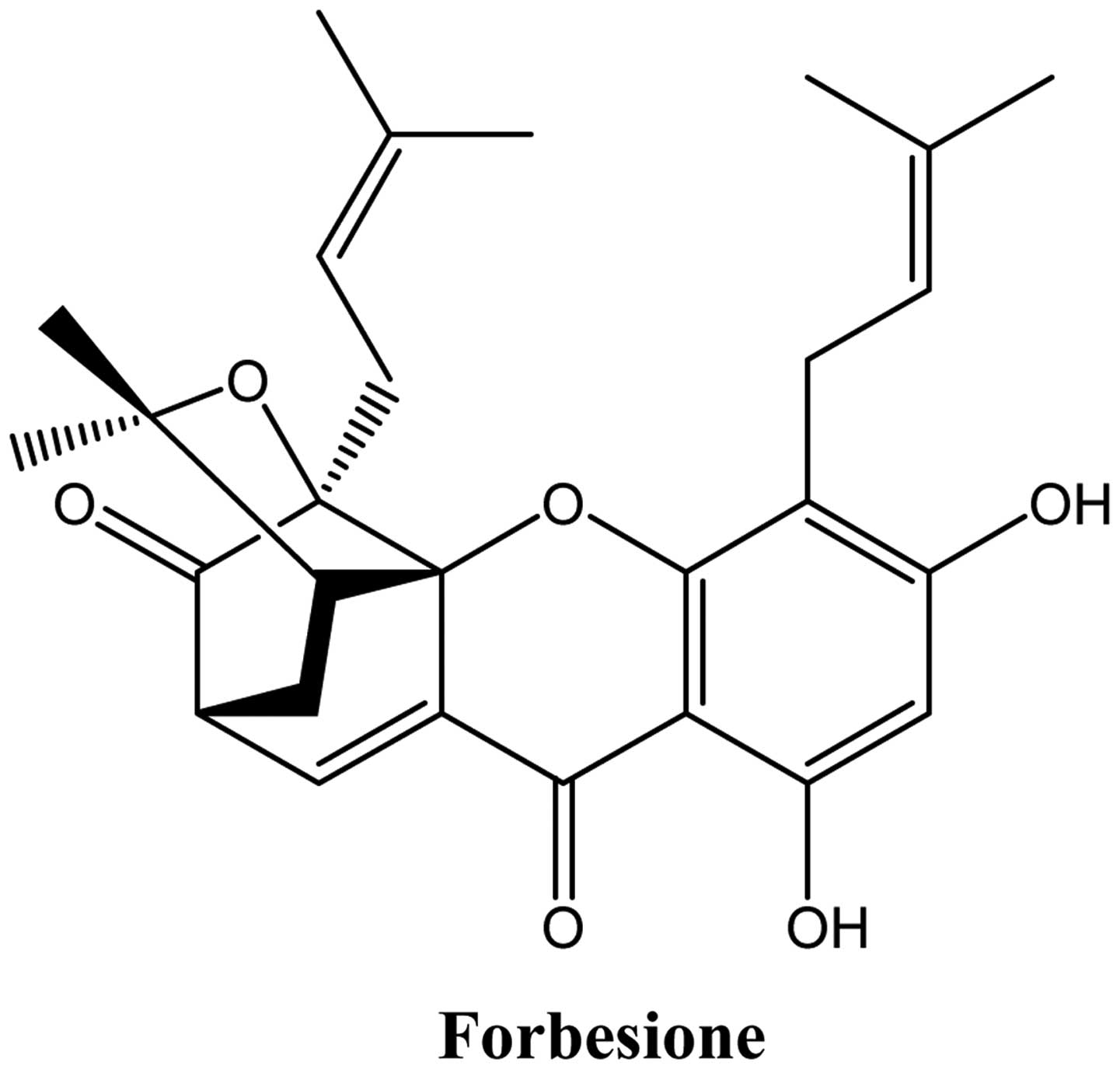
Forbesione is a caged xanthone isolated from the resin and fruits
of Garcinia hanburyi Hook. f. (family Guttiferae), which
have been used in Thai traditional medicine (6,7). Gambogic
acid, forbesione, isomorellin and isomorellinol, the caged
xanthones isolated from G. hanburyi, are reported to exhibit
antitumor activities (8) and
cytotoxic effects in several cancer cell lines (8–10).
Gambogic acid is the most intensively studied caged xanthone, and
exhibits potent antitumor activity in vitro and in
vivo (11–13) through several mechanisms, including
inhibition of topoisomerase II alpha activity (14), downregulation of telomerase (15), induction of cell cycle arrest
(16) and induction of apoptosis
(17). Due to its low toxicity
against normal tissues (18,19), gambogic acid is now approved for a
phase II clinical trial in China (20).
The results from our previous in vitro
studies demonstrated that gambogic acid, forbesione, isomorellin
and isomorellinol selectively inhibited the proliferation of the
human CCA cell lines KKU-100 and KKU-M156 by inducing apoptosis
through the mitochondrial pathway (6), and by inducing
G0/G1-phase cell cycle arrest through p53 and
the nuclear factor (NF)-κB signaling pathway (21). Our previous studies demonstrated that
combinations of isomorellin/doxorubicin and forbesione/doxorubicin
exhibited significant synergy for inhibition of cell growth and
induction of apoptosis in KKU-M156 and KKU-100 cells, respectively,
through suppression of multidrug resistance-associated protein 1,
NF-κB activation, enhanced expression of B-cell lymphoma
(Bcl)-2-like protein 4 (Bax)/Bcl-2, activation of caspase-9 and
caspase-3, and suppression of the expression of survivin,
procaspase-9 and procaspase-3 (22).
To date, the effects of caged xanthones on CCA in
vivo have not yet been reported. In the present study, the
growth-inhibitory effect of forbesione on Ham-1 CCA cells was
investigated in vitro and in vivo when grown as
allografts. It was observed that forbesione inhibits Ham-1 cell
growth in vitro and suppresses allograft tumor growth.
Furthermore, our study suggested that the possible mechanisms are
the induction of cell cycle arrest at the S phase by altering the
expression of cell cycle-regulated proteins [including cyclin E,
cyclin A, cyclin-dependent kinase 2 (Cdk2), p21, p27 and
proliferating cell nuclear antigen (PCNA)] and by inducing multiple
pathways of apoptosis [including the death receptor pathway, the
mitochondrial pathway and the endoplasmic reticulum (ER) pathway].
This which was achieved by altering the expression of genes and
proteins related to apoptosis regulation, including Fas,
Fas-associated death domain (FADD), procaspase-8, activated
caspase-3, Bcl-2, Bax, survivin, activated caspase-9, activated
caspase-12, NF-κB/p65 and inhibitor of κB-α (IκB-α).
Materials and methods
Materials
Forbesione (Fig. 1)
was extracted from G. hanburyi Hook. f. (family Guttiferae)
using bioassay-directed fractionation (10). Forbesione was dissolved in dimethyl
sulfoxide (DMSO) to create a stock solution of 1.8 mM that was
stored at −20°C. Annexin-V-FLOUS Staining kit was purchased from
Roche Applied Science (Penzberg, Germany); ethidium
bromide/acridine orange (EB/AO) mixture was purchased from
Sigma-Aldrich (Merck Millipore, Darmstadt, Germany); and
chemiluminescence reagent, RNasin Ribonuclease Inhibitor, DNase I,
oligo(dT), Moloney-murine leukemia virus (M-MLV) polymerase and
TrueStart Hot Start Taq DNA polymerase were purchased from Pierce
(Thermo Fisher Scientific, Inc., Waltham, MA, USA).
TRIzol® reagent was purchased from Invitrogen (Thermo
Fisher Scientific, Inc.). Primary antibodies against Bax, Bcl-2,
survivin, caspase-9, procaspase-3, Fas, FADD, procaspase-8, cyclin
E, cyclin A, Cdk2, p21, p27, IκB-α and NF-κB/p65, as well as
secondary antibodies, were purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). Activated caspase-3 and β-actin were
purchased from Sigma-Aldrich (Merck Millipore), while caspase-12
was purchased from Chemicon International, Inc. (Temecula, CA,
USA). Anti-cytokeratin 19 (CK19) antibody was purchased from Abcam
(Cambridge, UK), while anti-PCNA antibody was purchased from
Novocastra Laboratories Ltd. (Newcastle upon Tyne, UK). Anti-rabbit
and anti-mouse horseradish peroxidase-streptavidin conjugates were
purchased from Zymed Laboratories (Thermo Fisher Scientific, Inc.).
The primary and secondary antibodies used in the present study are
listed and characterized in Tables I
and II.
 | Table I.Primary antibodies used in western
blot analysis. |
Table I.
Primary antibodies used in western
blot analysis.
| Target | Type | Catalog no. | Host
species/targeted species | Dilution | Secondary
antibody |
|---|
| β-actin | Monoclonal | A1978 | Mouse
anti-human | 1:3,000 | Goat
anti-mouse |
| Bcl-2 | Polyclonal | sc-492 | Rabbit
anti-human | 1:200 | Goat
anti-rabbit |
| Bax | Polyclonal | sc-493 | Rabbit
anti-human | 1:200 | Goat
anti-rabbit |
| Survivin | Monoclonal | sc-17779 | Mouse
anti-human | 1:400 | Goat
anti-mouse |
| Caspase-9 | Polyclonal | sc-8355 | Rabbit
anti-human | 1:200 | Goat
anti-rabbit |
| Procaspase-3 | Polyclonal | sc-7148 | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| Activated
caspase-3 | Polyclonal | #9662s | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| Fas | Polyclonal | sc-715 | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| FADD | Polyclonal | sc-5559 | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| Procaspase-8 | Monoclonal | #9746 | Mouse
anti-human | 1:400 | Goat
anti-mouse |
| Cyclin E | Polyclonal | sc-481 | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| Cyclin A | Polyclonal | sc-751 | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| Cdk2 | Polyclonal | sc-163 | Rabbit
anti-human | 1:200 | Goat
anti-rabbit |
| p21 | Polyclonal | sc-397 | Rabbit
anti-human | 1:400 | Goat
anti-rabbit |
| p27 | Polyclonal | sc-527 | Rabbit
anti-human | 1:200 | Goat
anti-rabbit |
| IκB-α | Polyclonal | sc-371 | Rabbit
anti-human | 1:500 | Goat
anti-rabbit |
| NF-κB/p65 | Polyclonal | sc-109 | Rabbit
anti-human | 1:500 | Goat
anti-rabbit |
| Caspase-12 | Polyclonal | AB3613 | Rabbit
anti-human | 1:200 | Goat
anti-rabbit |
| CK19 | Monoclonal | ab7754 | Mouse
anti-human | 1:500 | Goat
anti-mouse |
| PCNA | Monoclonal | NCL-L-PCNA | Mouse
anti-human | 1:500 | Goat
anti-mouse |
| Table II.Primary antibodies used in
immunohistochemistry. |
Table II.
Primary antibodies used in
immunohistochemistry.
| Target | Type | Catalog no. | Host
species/targeted species | Dilution | Secondary
antibody |
|---|
| CK19 | Polyclonal | ab15463 | Rabbit
anti-human | 1:50 | Goat
anti-rabbit |
| PCNA | Monoclonal | NCL-L-PCNA | Mouse
anti-human | 1:50 | Goat
anti-mouse |
| Cyclin A | Polyclonal | sc-751 | Rabbit
anti-human | 1:50 | Goat
anti-rabbit |
| Bax | Polyclonal | ab7977 | Rabbit
anti-human | 1:100 | Goat
anti-rabbit |
| Bcl-2 | Polyclonal | sc-492 | Rabbit
anti-human | 1:50 | Goat
anti-rabbit |
| Caspase-9 | Polyclonal | ab2014 | Rabbit
anti-human | 1:100 | Goat
anti-rabbit |
| Caspase-3 | Polyclonal | ab44976 | Rabbit
anti-human | 1:100 | Goat
anti-rabbit |
Cell culture
The cell line Ham-1 (23), derived from the CCA tissue of
Opisthorchis viverrini-infected and
N-nitrosodimethylamine-treated Syrian hamsters, which was
established at the Department of Biochemistry, Faculty of Medicine,
Khon Kaen University (Khon Kaen, Thailand), was used in the present
study. Ham-1 cells were cultured in Dulbecco's modified Eagle
medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented
with 10% heat-inactivated fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin, and maintained at 37°C in a 5% CO2
humidified incubator.
Sulforhodamine B (SRB) assay for cell
number determination
Ham-1 cells were seeded into 96-well plates (Costar;
Corning Incorporated, Corning, NY, USA) at a density of
1×104 cells/well. The cytotoxicity of forbesione was
determined using the SRB assay as previously described (24).
Cell cycle analysis
Ham-1 cells (3×105 cells/well) seeded in
6-well plates were treated with various forbesione concentrations
for 24 h. Cells were harvested by trypsinization, washed twice with
cold PBS, fixed in 70% ethanol at 4°C overnight, centrifuged at
1,000 × g for 10 min at 4°C, incubated for 45 min with RNase A
(final concentration, 2 µg/ml) and then stained with propidium
iodide (PI) (final concentration, 2.4 µg/ml) for 1 h on ice in the
dark. The DNA content and cell cycle distribution of the cells were
analyzed by FACSCalibur (BD Biosciences, Franklin Lakes, NJ,
USA).
Morphological characterization of cell
death
Ham-1 cells were treated with varying concentrations
of forbesione for 24 h and then stained with 14 µl of 100 µg/ml
EB/AO mixture. Apoptotic cells with condensed or fragmented
chromatin were determined with an Eclipse Ni-U microscope (Nikon
Corporation, Tokyo, Japan).
Assessment of apoptosis using flow
cytometry
Ham-1 cells were exposed to different concentrations
of forbesione for 6, 24 or 48 h. Floating and adherent cells were
harvested by trypsinization, washed with cold PBS and centrifuged
at 1,000 × g for 10 min at 4°C. Pelleted cells were resuspended in
400 µl of PBS and then stained with 100 µl of incubation buffer
containing 2 µl of annexin V-fluorescein isothiocyanate (FITC) and
2 µl of PI for 10–15 min at room temperature in the dark, prior to
analysis by flow cytometry (BD Biosciences).
Western blot analysis
Ham-1 cells were seeded in 10-cm-diameter dishes
(Costar; Corning Incorporated) at a density of 1×106
cells/dish and incubated with various concentration of forbesione
for 24 h. Harvested cells were washed twice with cold PBS and lysed
with ice-cold radioimmunoprecipitation assay buffer [50 mM Tris-HCl
(pH 7.5), 150 mM NaCl, 0.5% Nonidet P-40, 1 mM EDTA, 1 mM
dithiothreitol, 0.5% deoxycholate and 0.1% SDS] with protease
inhibitor cocktail (Pierce; Thermo Fisher Scientific, Inc.) and
phosphatase inhibitor cocktail (Pierce; Thermo Fisher Scientific,
Inc.). The cell lysates were homogenized and clarified by
centrifugation at 15,000 × g for 30 min at 4°C. The protein
concentration of the total cell lysate was determined by the method
of Bradford (25). Cell lysates were
fractionated by 12% SDS-PAGE and transferred to nitrocellulose
membranes. Membranes were blocked with 5% nonfat milk in 0.1% Tween
20 in PBS at 37°C for 1 h, followed by incubation with the
corresponding primary β-actin (1:3,000), Bcl-2 (1:200), Bax
(1:200), survivin (1:400), caspase-9 (1:200), procaspase-3 (1:400),
activated caspase-3 (1:400), Fas (1:400), FADD (1:400),
procaspase-8 (1:400), cyclin E (1:400), cyclin A (1:400), Cdk2
(1:200), p21 (1:400), p27 (1:200), IκB-α (1:500), NF-κB/p65
(1:500), caspase-12 (1:200), CK19 (1:500) and PCNA (1:500)
antibodies at 4°C overnight. Next, membranes were washed with 0.1%
Tween 20 in PBS and incubated with the appropriate secondary
antibodies (anti-mouse or anti-rabbit; 1:10,000). The bound
secondary antibody-POD conjugates were visualized using an enhanced
chemiluminescence reagent (Pierce; Thermo Fisher Scientific, Inc.),
quantified by densitometry (ImageQuant LAS 4000; GE Healthcare Life
Sciences, Chalfont, UK) and analyzed using the program Scion Image
(Version 4.0.2; Scion Corporation, Frederick, MD, USA). Data for
each protein expressed were normalized to β-actin expression.
Experimental allografts of Ham-1 in
hamsters
A total of 12 male Syrian hamsters aged 6–8 weeks
(Animal Unit, Faculty of Medicine, Khon Kaen University, Khon Kaen,
Thailand) were maintained at 22°C and exposed to 12 h light/dark
cycles with free access to food and water. The hamsters were
injected intradermally with 2.5×105 Ham-1 cells in 50 µl
of 1% FBS in DMEM into the flank. After the tumors had become
established (~3 days), those tumors whose volume had reached 10–20
mm3 were selected. Syrian hamsters were randomly
allocated into two groups (n=6/group): i) Negative control group
(1% DMSO, oral every day); and ii) forbesione (50 mg/kg, oral every
day) for 1 month. The day of first treatment was set as day 1. The
size of the tumors was assessed every other day, and tumor volume
was determined using the following equation: Tumor volume
(mm3) = (a × b2) / 2 (where a and b refer to
the longest and shortest dimension in mm, respectively) (12). The animals were weighed, and food and
water intake was measured daily. The physical activity and
mortality of the animals was monitored daily during the
experimental period to assess the toxicity of the treatments. All
the protocols were approved by the animal ethics committee of Khon
Kaen University (Khon Kaen, Thailan; approval no. AEKKU
2/2556).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from tumor tissues with
TRIzol® solution (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Complementary DNA
(cDNA) was synthesized from 3 µg of total RNA using 0.5 µg/µl of
oligo(dT)15 primers, 20 U of RNase inhibitor, 20 U of M-MLV
(Pierce; Thermo Fisher Scientific, Inc.), 10 mM deoxynucleotides
(dNTPs) and M-MLV reverse transcriptase 5X reaction buffer. RT-qPCR
was performed using the SYBR® Green method on a CFX96
Touch™ Real-Time PCR Detection System (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) to analyze the relative abundance of each
individual messenger RNA (mRNA) relative to that of GAPDH, which
was used as an endogenous control. The reaction mixture contained 3
µl of 1:10 diluted single-strand cDNA, 2 µl of 10X HotStar Taq
polymerase buffer, 1 µl of each 5 mM dNTP, 2.4 µl of 25 mM
MgCl2, 1 µl of 5 µM primer pairs, 0.2 µl of HotStar Taq
DNA polymerase (Pierce; Thermo Fisher Scientific, Inc.) and 7.4 µl
of distilled water to a final volume of 20 µl. PCR cycling
conditions were as follows: Initital denaturation at 95°C for 10
sec followed by 35–50 cycles of 95°C for 15 sec, 52–62C for 60 sec
and 72°C for 1 min, with a final extension step at 72°C for 10 min.
The PCR primers for GAPDH (26),
CK19, apoptotic protease activating factor (Apaf)-1 (27), Bax (26), caspase-9 and caspase-3 (Integrated DNA
Technologies, Coralville, IA, USA) are summarized in Table III. Relative expression of CK19,
Apaf-1, Bax, caspase-9 and caspase-3 mRNA was calculated using the
comparative ΔΔCq method as previously described (28). All the values were reported as
fold-change over background levels detected in the untreated
control as a calibrator.
| Table III.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table III.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Primer
sequences | No. of cycles | Ta,°C | GenBank accession
no. (ref.) |
|---|
| GAPDH | Forward:
5′-GGCATTGTGGAAGGGCTCAT-3′ | 35 | 60 | (25) |
|
| Reverse:
5′-GACACATTGGGGGTAGGAACAC-3′ |
|
|
|
| CK19 | Forward:
5′-GCGGGACAAGATTCTTGGTG-3′ | 40 | 62 | NM_008471.2 |
|
| Reverse:
5′-CTTCAGGCCTTCGATCTGCAT-3 |
|
|
|
| Bax | Forward:
5′-AGCTGCAGAGGATGATTGCT-3′ | 40 | 62 | (25) |
|
| Reverse:
5′-CTCTCGGAGGAAGTCCAGTG-3′ |
|
|
|
| Apaf-1 | Forward:
5′-ATCCTGGTGCTTTGCCTCTA-3′ | 50 | 62 | (26) |
|
| Reverse:
5′-TACACCCCCTGAAAAGCAAC-3′ |
|
|
|
| Caspase-9 | Forward:
5′-CTGCACTTCCTCTCAAGGCA-3 | 40 | 52 | XM_005139261.1 |
|
| Reverse:
5′-GAAACAGCATTGGCGACCTG-3′ |
|
|
|
| Caspase-3 | Forward:
5′-TTCGAGCCACCGAGGAGATA-3′ | 40 | 54 | NM_001281582.1 |
|
| Reverse:
5′-TTGGGGACATCATCCACACG-3′ |
|
|
|
Immunohistochemical (IHC) staining for
PCNA, CK19, cyclin A, Bcl-2, Bax, caspase-9 and caspase-3
Formalin-fixed, paraffin-embedded blocks of
allograft tissues were prepared, and sections (4-µm) were
deparaffinized in xylene and rehydrated through graded alcohol
series to water. Sections were rinsed for 5 min in distilled water,
and antigen retrieval was performed by autoclaving the sections in
50 mM citrate buffer (pH 6.0) for 5 min at 121°C and treating them
for 30 min in methanol containing 3% H2O2 to
block any endogenous peroxidase activity. Upon blocking with 5%
skim milk in PBS, the sections were incubated with the primary
monoclonal antibodies against CK19 (a marker of bile duct
epithelium), PCNA (a proliferation marker), cyclin A (an S-phase
cell cycle-regulated protein), or Bcl-2, Bax, caspase-9 or
caspase-3 (apoptosis-related proteins) at 37°C for 90 min, as
described previously (29). Upon
rinsing, anti-rabbit or anti-mouse horseradish
peroxidase-conjugated secondary antibodies (dilution 1:100; Zymed
Laboratories; Thermo Fisher Scientific, Inc.) were used to detect
the primary antibodies. The sections were subsequently washed twice
with PBS. The peroxidase reaction was developed with
3-amino-9-ethylcarbazole (Sigma-Aldrich; Merck Millipore) as a
chromogen, and the sections were washed for 5 min in tap water and
counterstained with hematoxylin. The IHC results were evaluated by
two observers by semi-quantifying the percentage of
immunopositivity of ten fields per slide, and scored as
0%=negative, <25%=1+, 25–50%=2+, >50–75%=3+ and
>75%=4+.
Histopathological study
The liver, kidney and stomach of hamsters from each
group were fixed in 10% buffered formalin for 24 h and then
embedded in paraffin wax. Sections of 4-µm thickness were stained
with hematoxylin and eosin (H&E) staining. The tissues slides
were observed and photographed on a light microscope at ×10 and ×40
magnification (Eclipse Ni-U; Nikon Corporation).
Liver and kidney function assays
Plasma alanine aminotransferase (ALT) and alkaline
phosphatase (ALP), markers of liver damage, as well as blood urea
nitrogen (BUN) and creatinine levels were analyzed at the Chemistry
Room, Community Laboratory, Faculty of Associated Medical Sciences,
Khon Kaen University (Khon Kaen, Thailand) (30).
Statistical analysis
All the values are expressed as the mean ± standard
deviation. All data analysis was performed using SPSS 10.0
statistical software (SPSS, Inc., Chicago, IL, USA). Statistical
comparisons of the results were evaluated using the Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference, and the statistical significance of the
differences between the groups were indicated as *P<0.05,
**P<0.01 or ***P<0.001.
Results
Forbesione inhibits the proliferation
of Ham-1 cells in vitro
Treatment of Ham-1 cells with forbesione inhibited
cell proliferation in a concentration-dependent manner, with a half
maximal inhibitory concentration value of 3.21±0.27 µM.
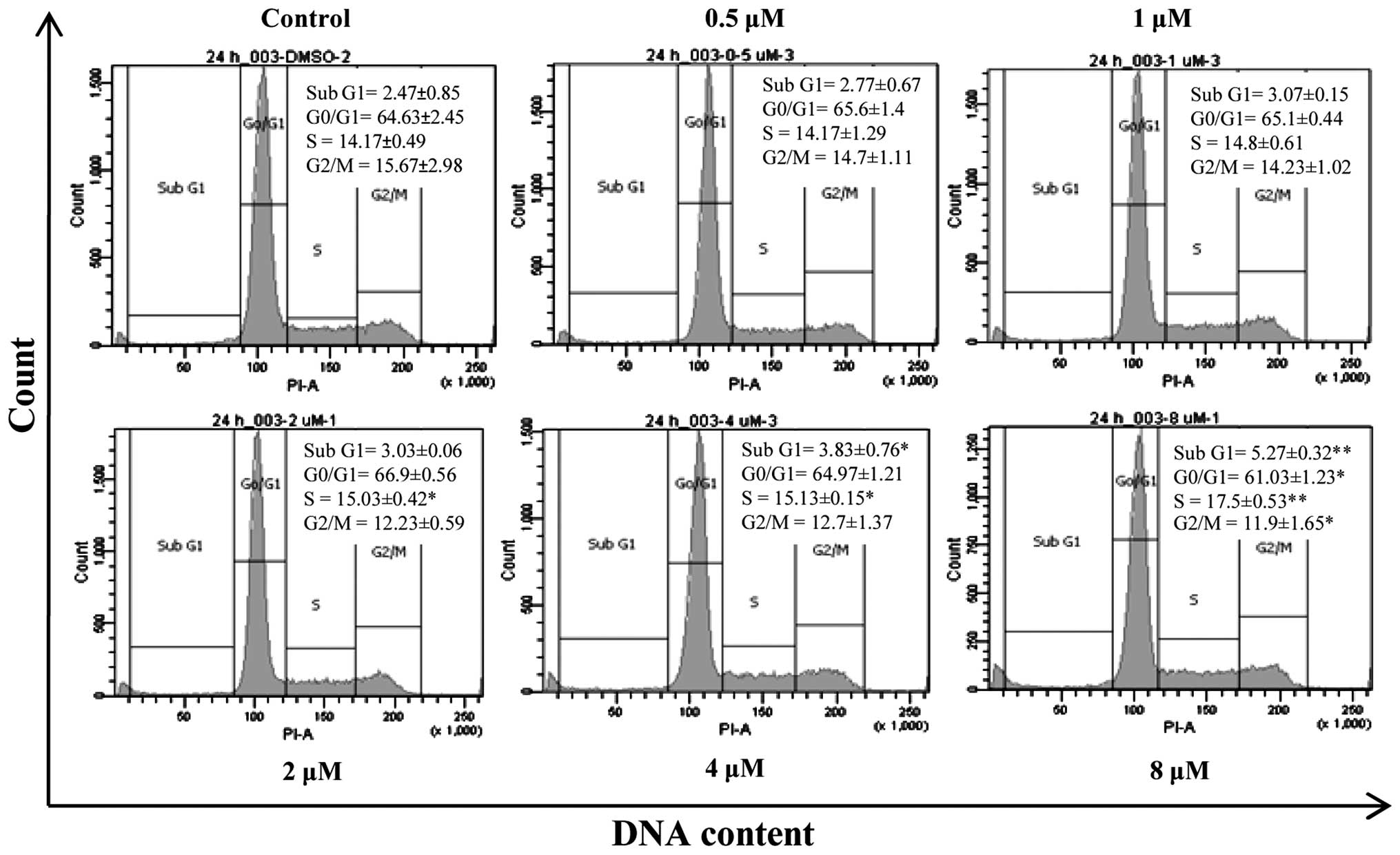
Forbesione induces S-phase cell cycle
arrest in Ham-1 cells
Since forbesione significantly reduced Ham-1 cell
numbers after 24 h, the effects of forbesione at 0.5, 1, 2, 4 and 8
µM on the cell cycle progression of Ham-1 cells were determined.
The results revealed that Ham-1 cells became arrested in the S
phase (P<0.01) in response to forbesione treatment in a
concentration-dependent manner, concomitant with a reduction in the
proportion of cells in the G0/G1 (P<0.05) and G2/M (P<0.05)
phases (Fig. 2). In addition, the
percentage of Ham-1 cells in the sub-G1 fraction (apoptotic cells)
was significantly increased in a concentration-dependent manner
(P<0.01). These results support that forbesione induces a
growth-inhibitory effect, cell cycle arrest and apoptosis.
Forbesione induces changes in the
nuclear morphology of Ham-1 cells
As forbesione significantly increased the
sub-G1 fraction, nuclear morphological features of
apoptosis were assessed in Ham-1 cells treated with various
concentrations of forbesione for 24 h followed by EB/AO staining.
The results indicated that the untreated cells displayed normal
nuclear shapes that stained uniformly green, whereas the treated
cells exhibited bright green, condensed and fragmented chromatin
(Fig. 3A), which are the hallmarks of
apoptosis (31).
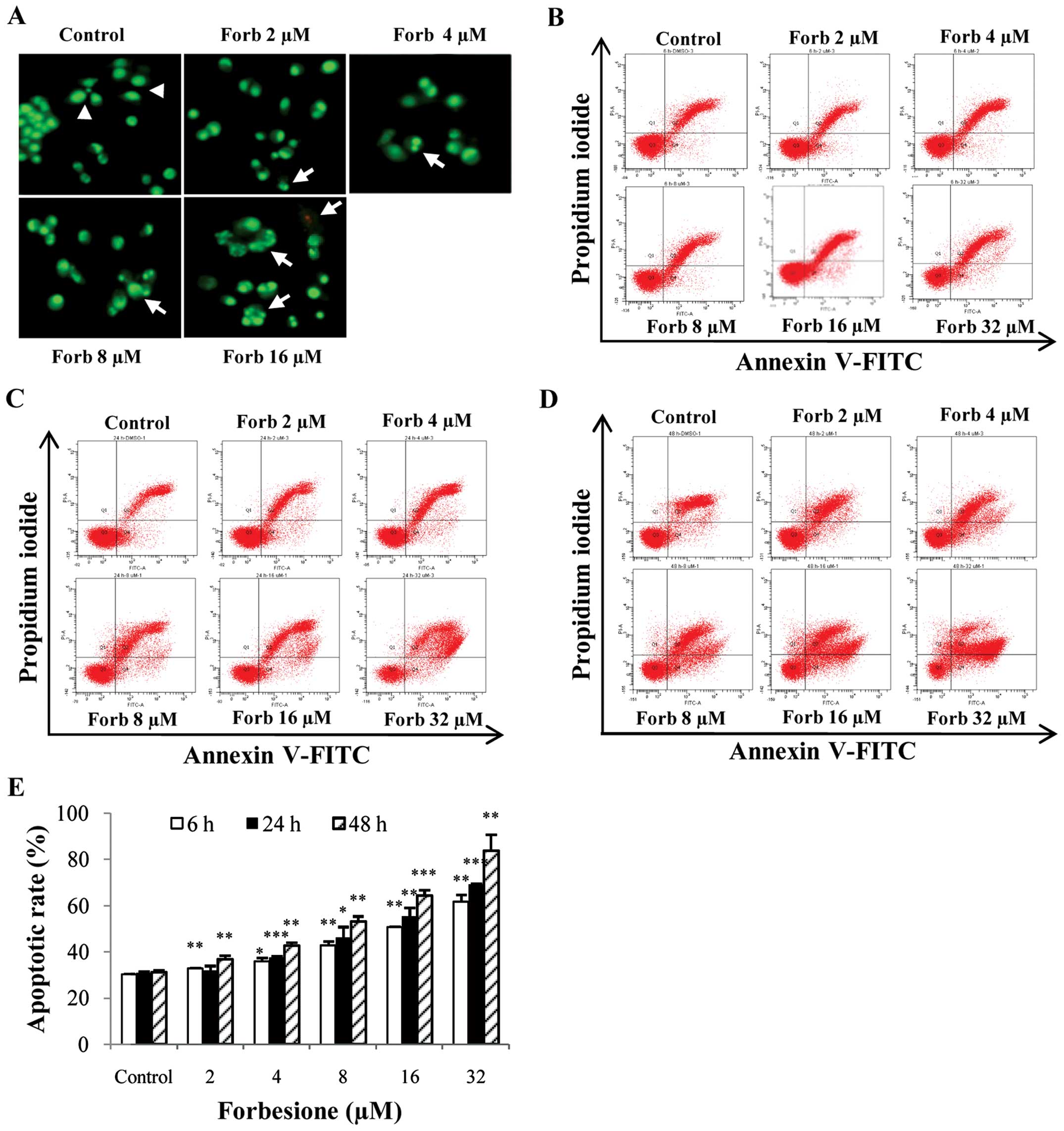 | Figure 3.Apoptotic effect in Ham-1 cells
following treatment with forbesione. (A) Nuclear morphological
change of Ham-1 cells treated with different concentrations of
forbesione (0, 2, 4, 8, 16 and 32 µM) for 24 h followed by ethidium
bromide/acridine orange staining (magnification, ×40). Nuclei of
living cells were uniformly stained green (arrowheads). Early
apoptotic cells exhibited condensed and fragmented chromatin
(arrows). Flow cytometric analysis of the induction of apoptosis in
Ham-1 cells following treatment with forbesione (0, 2, 4, 8, 16 and
32 µM) for (B) 6, (C) 24 and (D) 48 h. (E) Graph of the percentage
of apoptotic cells for the indicated concentrations at different
times. Data are presented as the mean ± standard deviation of three
separate experiments. *P<0.05, **P<0.01, ***P<0.001. Forb,
forbesione; FITC, fluorescein isothiocyanate. |
Forbesione increases the percentage of
Ham-1 cells undergoing apoptosis
Ham-1 cells were treated with 0.1% DMSO or with 2,
4, 8, 16 or 32 µM forbesione for 6, 24 and 48 h, and then the
percentages of apoptotic cells were determined by annexin V-FITC/PI
staining. The flow cytometry results revealed that the proportions
of cells undergoing apoptosis were significantly increased by
forbesione in a concentration- and time-dependent manner (Fig. 3B-E). Treatment with 32 µM forbesione
for 6, 24 and 48 h resulted in 50.75 (P<0.01), 55.57
(P<0.001) and 64.43% (P<0.001) apoptotic cells, respectively
(Fig. 3E).
Forbesione alters the expression of
proteins regulating the cell cycle and apoptosis
As forbesione induced cell cycle arrest at the S
phase, western blot analysis was used to investigate the expression
of proteins involved in cell cycle regulation at the S phase in
Ham-1 cells treated with 0.1% DMSO or with 2, 4 or 8 µM forbesione
for 24 h. The results demonstrated that cyclin E, cyclin A and Cdk2
levels were significantly decreased (P<0.01, P<0.05 and
P<0.01, respectively), while p21 and p27 levels were
significantly increased (P<0.05), in forbesione-treated cells
compared with those in the control cells (Fig. 4A).
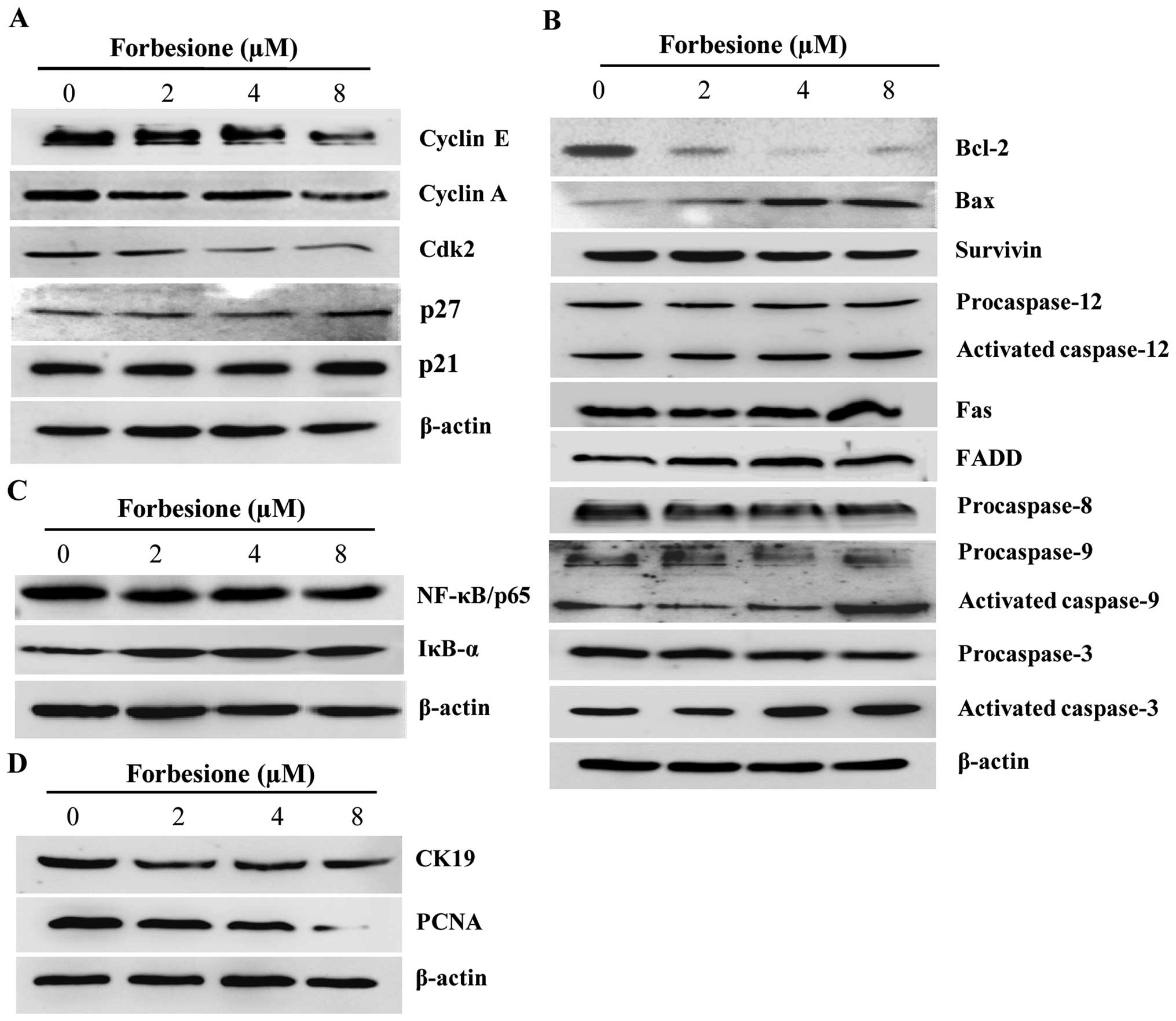 | Figure 4.Expression of proteins associated
with the regulation of cell cycle arrest, apoptosis and growth
inhibition in forbesione-treated cells. (A) Representative western
blot analysis of cell lysates after being treated with forbesione
(0, 2, 4 and 8 µM) for 24 h; the expression of cell
cycle-regulatory proteins was decreased. (B) Compared with the
control group, the expression of apoptotic-regulatory molecules
changed (increased or decreased) in the forbesione-treated groups.
(C) After 24 h of treatment, forbesione-treated cells exhibited
increased expression of NF-κB, while the expression of IκB-α was
decreased, as compared with that in the control group. (D) Compared
with the control group, the expression of CK19 and PCNA proteins
was decreased following treatment with forbesione. Bcl, B-cell
lymphoma; Bax, Bcl-2-like protein 4; FADD, Fas-associated death
domain; Cdk, cyclin-dependent kinase; IκB-α; inhibitor of κB-α; NF,
nuclear factor; CK, cytokeratin; PCNA, proliferating cell nuclear
antigen. |
Apoptosis is triggered by multiple pathways,
including the extrinsic, intrinsic and ER pathways (32). The extrinsic pathway is regulated by
the activation of death receptors on the cell surface such as Fas
(also known as apoptosis antigen-1 and cluster of differentiation
95) (33), which will activate
downstream signal transduction molecules, including FADD,
procaspase-8 and procaspase-3, subsequently leading to apoptotic
cell death (34). Treatment of Ham-1
cells with 0.1% DMSO or with 2, 4 or 8 µM forbesione for 24 h
significantly increased the level of Fas, FADD and activated
caspase-3 (P<0.05), while that of procaspase-8 and procaspase-3
was significantly decreased (P<0.05 and P<0.01, respectively;
Fig. 4B). These findings suggest that
forbesione induces apoptosis possibly through the Fas signaling
pathway.
The intrinsic pathway is controlled by the Bcl-2
family of proteins, including pro-apoptotic proteins such as Bax
and Bak, and anti-apoptotic proteins such as Bcl-2, which leads to
the activation of procaspase-9 and procaspase-3 and subsequently
apoptotic cell death (35). In the
present study, forbesione treatment significantly decreased the
expression of Bcl-2, procaspase 9 (P<0.05) and procaspase 3
(P<0.01), while significantly increasing the expression of Bax,
activated caspase-9 and activated caspase-3 (P<0.05; Fig. 4B), suggesting that forbesione-induced
apoptosis is mediated in part through the intrinsic mitochondrial
pathway.
Upon encountering ER stress, Ca2+ efflux
from the ER will activate the calcium-dependent protease m-calpain,
which activates procaspase-12 and subsequently activates
procaspase-9 and procaspase-3, thereby driving apoptotic cell death
(36,37). Forbesione treatment of Ham-1 cells
significantly decreased the protein level of procaspase-12,
procaspase-9 and procaspase-3 (P<0.05, P<0.05 and P<0.01,
respectively), while significantly increasing the protein level of
activated caspase-12, activated caspase-9 and activated caspase-3
(P<0.05; Fig. 4B), suggesting that
the induction of apoptosis occurs through the ER pathway.
NF-κB is a nuclear factor known to activate the
expression of genes involved in cell survival (anti-apoptotic
proteins such as Bcl-2 and survivin), and NF-κB activation is
regulated by IκB-α (38,39). The present results indicated that
forbesione treatment significantly decreased NF-κB/p65 expression
(P<0.01) while significantly increasing IκB-α expression
(P<0.05; Fig. 4C). These results
support that the inhibition of NF-κB activation by forbesione is
possibly mediated through decreased NF-κB expression and increased
IκB-α expression, leading to reduced expression of Bcl-2 and
survivin.
Forbesione decreases expression of
CK19 and PCNA proteins in Ham-1 cells in vitro
To further assess the changes during the growth
inhibition induced by forbesione, the expression of CK19 and PCNA
proteins in Ham-1 cells treated with 0.1% DMSO or with 2, 4 or 8 µM
forbesione for 24 h was determined by western blot analysis. The
results revealed that the CK19 and PCNA protein levels were
significantly decreased in forbesione-treated cells (P<0.01 and
P<0.05, respectively) compared with those in the control cells
(Fig. 4D).
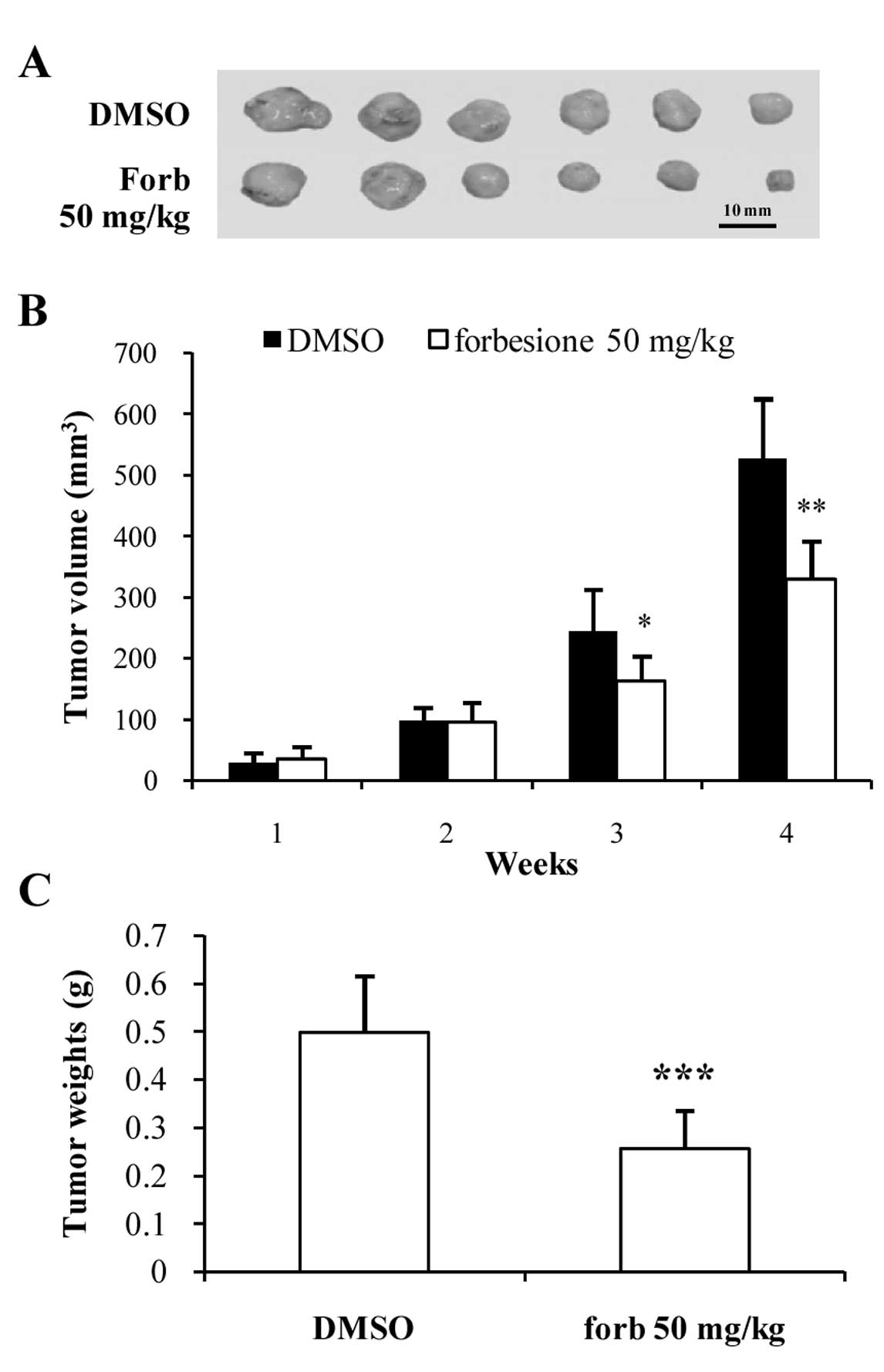
Forbesione suppresses tumor growth in
the Ham-1 allograft hamster model
To determine the effect of forbesione on the growth
of CCA in vivo, Ham-1 cells were injected intradermally in
the flanks of Syrian hamsters. Tumors were allowed to establish for
3 days prior to administration of forbesione (50 mg/kg/day orally
for 4 weeks). The control group received the same volume of diluent
control (1% DMSO in 1% FBS-DMEM). Forbesione treatment reduced the
tumor size (Fig. 5A), volume
(P<0.01; Fig. 5B) and weight
(P<0.001; Fig. 5C) when compared
to the control group. The mean tumor weight of the
forbesione-treated group (0.27 g) was significantly lower than that
of the control group (0.54 g).
Forbesione decreases the relative
level of CK19 mRNA while increasing that of Bax, Apaf-1, caspase-9
and caspase-3 in tumor allografts
To investigate the mechanisms of tumor growth
suppression mediated by forbesione, the relative changes in CK19,
Bax, Apaf-1, caspase-9 and caspase-3 mRNA were assessed in tumor
tissues derived from the control and forbesione-treated groups by
RT-qPCR. The relative expression of CK19 mRNA was significantly
decreased, while that of Bax, Apaf-1, caspase-9 and caspase-3 was
significantly increased in the tumor tissues from the
forbesione-treated group as compared with those in the control
group (P<0.05; Fig. 6A). These
findings support that apoptosis induction by forbesione leads to
reduced expression of CK19 [a marker of bile duct epithelial cells
(40)] (Fig. 6A).
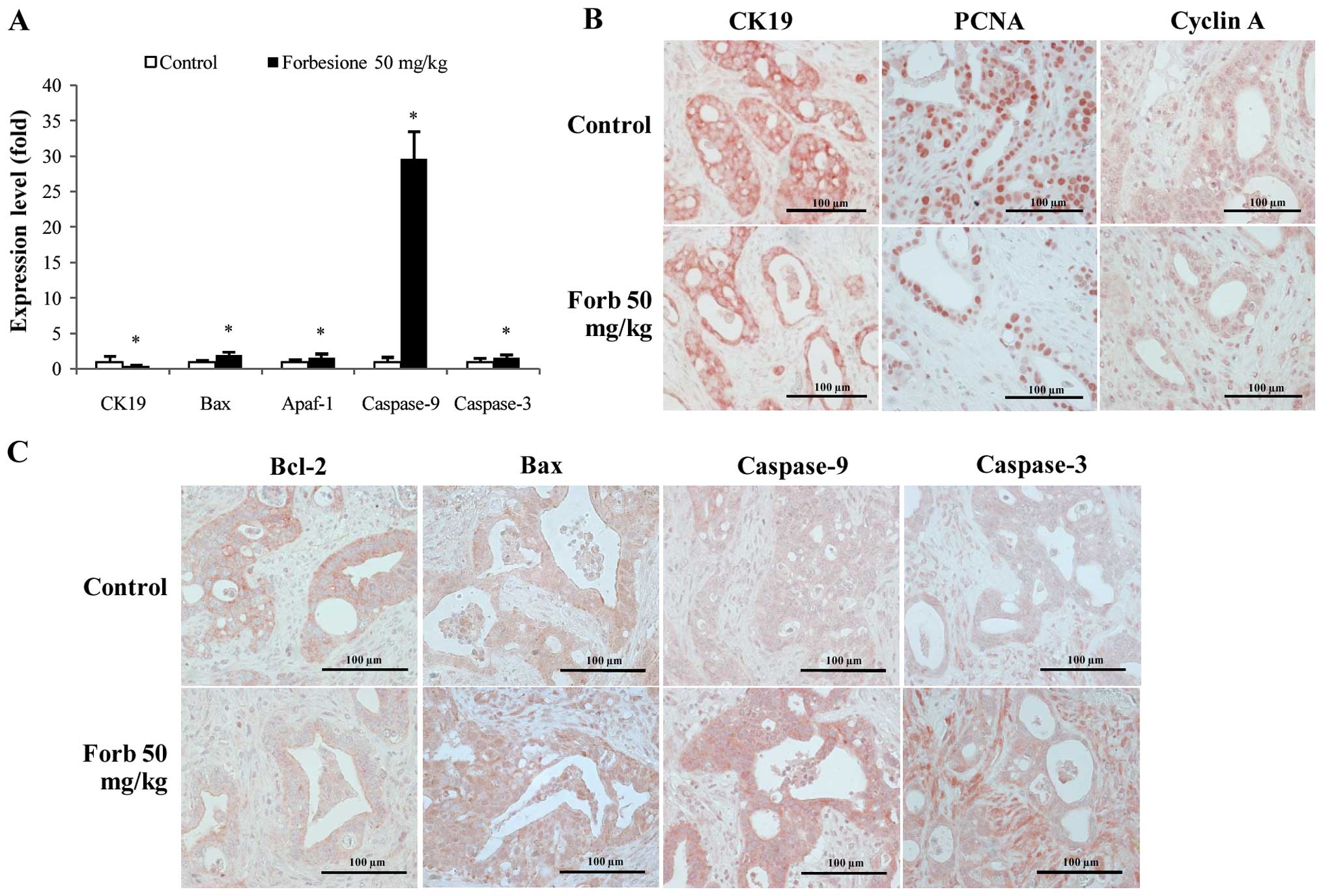 | Figure 6.Expression changes in genes and/or
proteins associated with the regulation of cell cycle and cell
apoptosis (epithelial bile duct marker CK19 and proliferation
marker PCNA). (A) Compared with the control group, the gene
expression of CK19 significantly decreased, while the gene
expression of Bax, Apaf-1, caspase-9 and caspase-3 significantly
increased, in the forbesione-treated group. Data are presented as
the mean ± standard deviation. *P<0.05. (B and C) Representative
immunohistochemical staining of Ham-1 allograft tissue
(magnification, ×40). Compared with the control group, the protein
expression of (B) CK19, PCNA, cyclin A and (C) Bcl-2 was decreased,
while the protein expression of (C) Bax, caspase-9 and caspase-3
was increased, in the forbesione-treated group. Forb, forbesione;
Apaf, apoptotic protease activating factor; Bcl, B-cell lymphoma;
Bax, Bcl-2-like protein 4; CK, cytokeratin; PCNA, proliferating
cell nuclear antigen. |
Forbesione decreases the expression of
CK19, PCNA, cyclin A and Bcl-2 proteins, and increases the
expression of Bax, caspase-9 and caspase-3 proteins in tumor
allografts
To further assess the changes during the suppression
of tumor growth mediated by forbesione, the allograft tissues
derived from the control and forbesione-treated groups were IHC
stained for CK19, PCNA, an S-phase cell cycle-regulated protein
(cyclin A) and apoptosis-related proteins (Bcl-2, Bax, caspase-9
and caspase-3). Semi-quantitative IHC scores were analyzed. As
compared with the control allograft group, allografts from the
forbesione-treated hamsters had significantly decreased expression
of CK19 (P=0.036), PCNA (P=0.032), cyclin A (P=0.028) and Bcl-2
(P=0.039), with significantly increased expression of Bax
(P=0.039), caspase-9 (P=0.028) and caspase-3 (P=0.039) (Fig. 6B and C and Table IV). These results corroborate our
in vitro study in which forbesione suppressed tumor growth
partly through cell cycle arrest (by decreasing the abundance of
the cell cycle regulatory protein cyclin A) and through induction
of apoptosis (by modulating the expression of apoptosis-related
proteins).
| Table IV.Effect of forbesione on IHC scores
for the expression of CK19, PCNA, cyclin A, Bcl-2, Bax, caspase-9
and caspase-3 in allograft tissues. |
Table IV.
Effect of forbesione on IHC scores
for the expression of CK19, PCNA, cyclin A, Bcl-2, Bax, caspase-9
and caspase-3 in allograft tissues.
|
|
| IHC score |
|
|---|
|
|
|
|
|
|---|
| Antigen | Groups | 0 n (%) | 1 n (%) | 2 n (%) | 3 n (%) | 4 n (%) | P-value |
|---|
| CK19 | Control | 0 (0) | 0 (0) | 1 (25) | 1 (25) | 2 (50) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 1 (25) | 2 (50) | 1 (25) | 0 (0) | 0.036a |
| PCNA | Control | 0 (0) | 0 (0) | 1 (25) | 3 (75) | 0 (0) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 2 (50) | 2 (50) | 0 (0) | 0 (0) | 0.032a |
| Cyclin A | Control | 0 (0) | 0 (0) | 0 (0) | 1 (25) | 3 (75) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 0 (0) | 3 (75) | 1 (25) | 0 (0) | 0.028a |
| Bcl-2 | Control | 0 (0) | 0 (0) | 2 (50) | 0 (0) | 2 (50) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 1 (25) | 3 (75) | 0 (0) | 0 (0) | 0.039a |
| Bax | Control | 0 (0) | 1 (25) | 2 (50) | 0 (0) | 1 (25) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 0 (0) | 0 (0) | 2 (50) | 2 (50) | 0.039a |
| Caspase-9 | Control | 0 (0) | 0 (0) | 2 (50) | 1 (25) | 1 (25) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 0 (0) | 0 (0) | 2 (50) | 2 (50) | 0.028a |
| Caspase-3 | Control | 0 (0) | 1 (25) | 2 (50) | 0 (0) | 1 (25) |
|
|
| Forbesione (50
mg/kg) | 0 (0) | 0 (0) | 1 (25) | 0 (0) | 3 (75) | 0.039a |
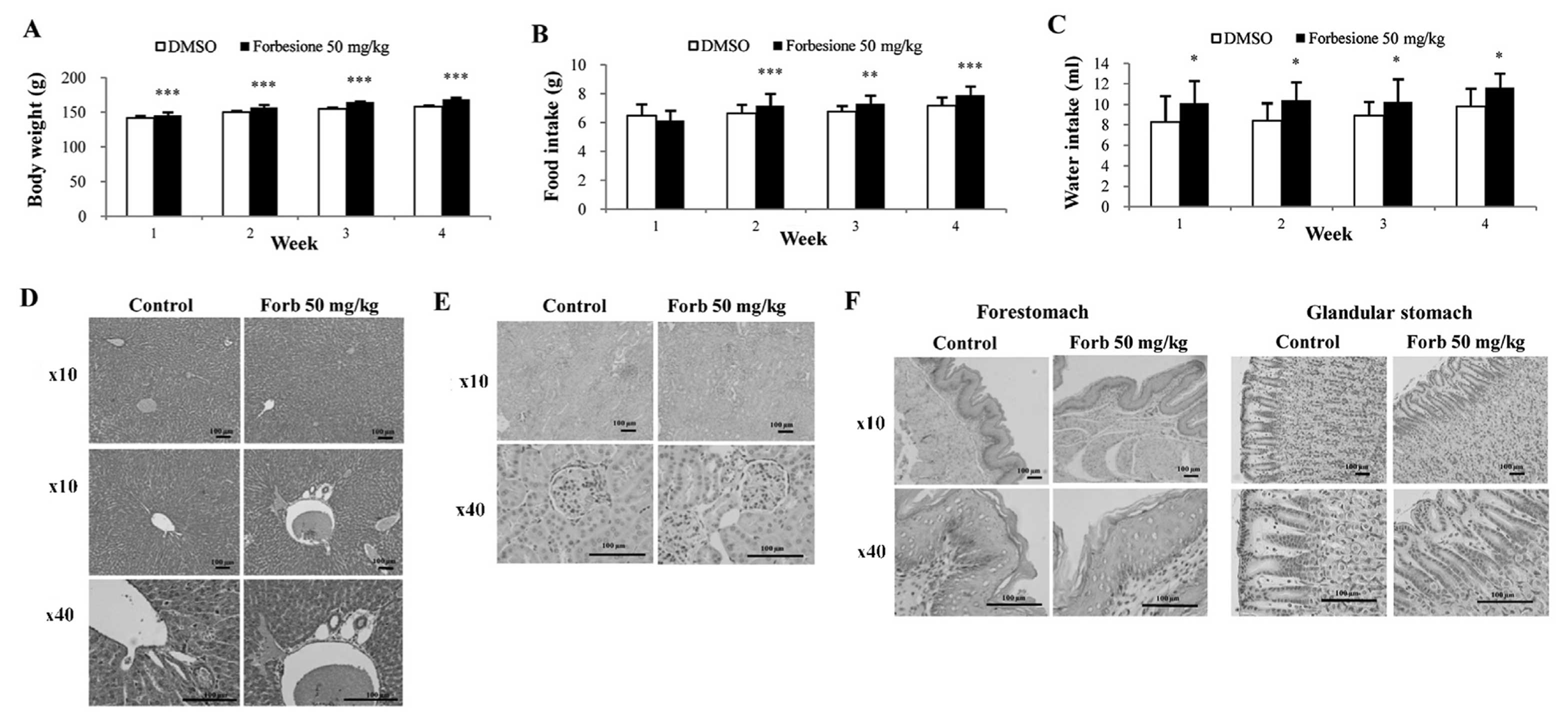
Forbesione has no side effects in
hamsters
To assess whether forbesione has side effects on
treated hamsters, several parameters were determined, including
physical activity, body weight, histopathological changes within
the liver, kidney and stomach, and liver and kidney function tests.
During 4 weeks of experiments, the physical activities were
observed, and the body weight, food intake and water intake were
measured daily. The forbesione-treated hamsters were healthy, with
no changes in normal physical activities as compared with the
control group. In the forbesione-treated group, the body weight,
food and water intake were significantly increased as compared with
those in the control hamsters (P<0.001, P<0.001 and
P<0.05, respectively; Fig.
7A-C).
At the end of the treatment, H&E-stained
sections of the liver, kidney and stomach of the control and
forbesione-treated hamsters were evaluated by two investigators
without knowledge of the treatment status. Semi-quantitative
analysis was performed to assess inflammatory cell aggregates
surrounding the hepatic bile ducts and present in the liver, kidney
and stomach (forestomach and glandular stomach) tissues. The
histopathological changes of the liver (Fig. 7D and Table
V), kidney (Fig. 7E and Table VI) and stomach (Fig. 7F and Table
VII) of the forbesione-treated hamsters were not significantly
different from those of the control hamsters.
| Table V.Histopathological features of liver
tissues and grading criteria level in each experimental group. |
Table V.
Histopathological features of liver
tissues and grading criteria level in each experimental group.
|
|
| Experimental
groups |
|---|
|
|
|
|
|---|
| Histopathology | Score | Control, n=6 n
(%) | Forbesione 50
mg/kg, n=6 n (%) |
|---|
| Periportal
inflammationa | 0 | 5 (83.3) | 5 (83.3) |
|
| 1 | 1 (16.7) | 1 (16.7) |
|
| 2 | 0 (0.0) | 0 (0.0) |
|
| 3 | 0 (0.0) | 0 (0.0) |
|
| 4 | 0 (0.0) | 0 (0.0) |
| Focal
inflammationb | 0 | 5 (83.3) | 6 (100.0) |
|
| 1 | 1 (16.7) | 0 (0.0) |
|
| 2 | 0 (0.0) | 0 (0.0) |
|
| 3 | 0 (0.0) | 0 (0.0) |
|
| 4 | 0 (0.0) | 0 (0.0) |
| Table VI.Histopathological features of kidney
tissues and grading criteria level in each experimental group. |
Table VI.
Histopathological features of kidney
tissues and grading criteria level in each experimental group.
|
|
| Experimental
groups |
|---|
|
|
|
|
|---|
| Histopathology | Score | Control, n=6 n
(%) | Forbesione 50
mg/kg, n=6 n (%) |
|---|
|
Inflammationa | 0 |
6 (100.0) |
6 (100.0) |
|
| 1 | 0 (0.0) | 0 (0.0) |
|
| 2 | 0 (0.0) | 0 (0.0) |
|
| 3 | 0 (0.0) | 0 (0.0) |
|
| 4 | 0 (0.0) | 0 (0.0) |
| Table VII.Histopathological features of stomach
tissues (forestomach and glandular stomach) and grading criteria
level in each experimental group. |
Table VII.
Histopathological features of stomach
tissues (forestomach and glandular stomach) and grading criteria
level in each experimental group.
|
|
| Experimental
groups |
|---|
|
|
|
|
|---|
| Histopathology | Score | Control, n=6 n
(%) | Forbesione 50
mg/kg, n=6 n (%) |
|---|
|
Inflammationa | 0 |
6 (100.0) | 4
(66.7) |
|
| 1 | 0 (0.0) | 2
(33.3) |
|
| 2 | 0 (0.0) | 0 (0.0) |
|
| 3 | 0 (0.0) | 0 (0.0) |
|
| 4 | 0 (0.0) | 0 (0.0) |
The sera collected from the control and
forbesione-treated hamsters at the end of the experiment were used
to determine the liver and kidney functions. The levels of liver
serum markers (ALT and ALP) and kidney serum markers (BUN and
creatinine) in the forbesione-treated group were similar to those
observed in the control group (Table
VIII), indicating no toxic effects of forbesione on the liver
or kidney in the study.
| Table VIII.Serum ALT, ALP, BUN and creatinine
levels in each experimental group. |
Table VIII.
Serum ALT, ALP, BUN and creatinine
levels in each experimental group.
|
| Kidney function
test | Liver function
test |
|---|
|
|
|
|
|---|
| Groups | BUN (mg/dl) | Creatinine
(mg/dl) | ALT (U/l) | ALP (U/l) |
|---|
| Control (n=6) | 18.80±1.29 | 0.31±0.04 | 78.33±22.87 | 97.83±11.03 |
| Forbesione 50 mg/kg
(n=6) | 18.21±2.33 | 0.28±0.04 | 66.33±15.01 | 102.50±8.75 |
| P-value | 0.279 | 0.181 | 0.157 | 0.105 |
Discussion
Forbesione, an active caged xanthone extracted from
the resin of G. hanburyi, has been reported to have a
growth-inhibitory effect against several cancer cell lines,
including CCA cell lines (6,9). Forbesione selectively inhibited cancer
cell growth with no effect on normal human peripheral blood
mononuclear cells (6). Recently, our
group has reported that forbesione induced apoptosis in CCA cell
lines through the intrinsic mitochondrial pathway (6). A synergistic effect of forbesione was
observed with doxorubicin on the induction of apoptosis in human
CCA cell lines (22). In the present
study, the growth-inhibitory effect of forbesione on Ham-1 cells
was investigated in vitro and in vivo in allografts,
and the molecular mechanisms were also clarified. It was observed
that forbesione significantly inhibited Ham-1 cell growth in
vitro and suppressed allograft growth in vivo.
Previous studies demonstrated that gambogic acid,
another caged xanthone isolated from G. hanburyi, causes cell cycle
arrest at different phases in different tumor cells (41). For example, gambogic acid induces
G0/G1 cell cycle arrest in the human CCA cell
lines KKU-100 and KKU-M156 (21), as
well as G2/M cell cycle arrest in breast carcinoma MCF-7
cells (42). Consistent with previous
studies, forbesione was observed to inhibit Ham-1 cell
proliferation by causing S-phase cell cycle arrest.
In eukaryotic cells, cell cycle progression is
regulated by Cdks and cyclins (43).
The formation of cyclin-Cdk complexes results in Cdk activation
leading to the phosphorylation of substrates involved in cell cycle
progression (44). Transition from
the G1 to S phase is regulated by cyclin E-Cdk2
complexes, while the S phase is regulated by cyclin A-Cdk2
complexes (44). Cdk inhibitor
proteins such as p21 and p27 can bind to the cyclin-Cdk complexes,
resulting in an inhibition of kinase activity and prevention of
cell cycle progression (45).
PCNA is involved in DNA repair, DNA replication and
cell cycle regulation (46). PCNA is
a transducer and a target of positive and negative signals
(46). Binding of PCNA to cyclin-Cdk
complexes may bring these regulatory proteins to the targets and
promote cell proliferation, whereas binding of p21 to PCNA prevents
PCNA from binding to the cell cycle or DNA replication machinery,
thus leading to inhibition of DNA replication and cell cycle arrest
(46). Our in vitro study
demonstrated that the decreased expression of cyclin E, cyclin A
and Cdk2 that occurred with increased expression of p21 and p27
could be a possible explanation for Ham-1 cells undergoing S-phase
cell cycle arrest in response to forbesione. These results are
partly confirmed by our in vivo study, in which forbesione
treatment decreased the expression of cyclin A in tumor allograft
tissues. In addition, the expression of PCNA and CK19 in
forbesione-treated Ham-1 cells in vitro and tumor tissues
from forbesione-treated hamsters was decreased compared with that
in the control group. This may possibly be due to high levels of
p21 in forbesione-treated cells causing downregulation of PCNA,
which results in decreased tumor cell proliferation (47). The present results are consistent with
those of our previous study (21),
which demonstrated that the isomorellin-induced
G0/G1 phase cell cycle arrest of human CCA
cell lines is mediated through inhibition of NF-κB activation,
upregulation of p53, p21 and p27, and downregulation of cyclin D1,
cyclin E, Cdk2 and Cdk4 protein expression. However, the mechanisms
of cell cycle arrest induced by forbesione are only partially
understood, and additional experiments are required to provide
conclusive answers.
Apoptosis is a highly regulated process that
involves the activation of a series of molecular events that lead
to a form of cell death (48).
Apoptosis is one of the mechanisms occurring in response to cancer
therapies (49). Our previous study
demonstrated that forbesione induces apoptosis of human CCA cell
lines through the mitochondrial pathway (6). In the present study, it was observed
that forbesione treatment of Ham-1 cells caused nuclear
morphological changes (chromatin condensation and nuclear
fragmentation), increased the proportion of cells in the
sub-G1 fraction (representing apoptotic cells) and
increased the percentage of apoptotic cells.
DNA fragmentation is the hallmark of apoptosis
(50). There are three steps of DNA
degradation, identified as a high-molecular weight DNA
fragmentation (0.05–1.00 Mb), an intermediate DNA fragmentation
(~300 Kb) and an internucleosomal DNA fragmentation (mono or
oligonucleosomal-size fragments) (51). Apoptotic cells, containing
internucleosomal-sized fragments, can be identified on DNA content
frequency histograms using a flow cytometric assay as cells with
fractional ‘sub-G1’ DNA content or ‘sub-G1’ cells, which represent
late apoptotic cells (51,52). However, during apoptosis, DNA
fragmentation may be terminated at 50–300 Kb fragments (53), which cannot be identified as ‘sub-G1’
cells (54). Furthermore, apoptotic
G2/M- or S-phase cells with fractional DNA content may appear at
the G1- or early S-position on DNA content histograms, but may not
be present at the ‘sub-G1’ location (55). Therefore, the sub-G1 DNA content
cannot be used to detect all apoptotic cells.
In the early events of apoptosis, externalization of
phosphatidylserine to the outer membrane of the cells occurs, which
can be detected by annexin V staining (56). PI has been used to differentiate
between early apoptotic (annexin V+ and PI-) and late apoptotic or
necrotic (annexin V+ and PI+) cells, depending on the integrity of
the cell membrane, which remains intact during early apoptosis
whilst being disrupted during necrosis (56).
The results in Fig. 3
represent both early and late apoptotic cells, which exhibit a
higher percentage of apoptotic cells than that of late apoptotic
cells in the sub-G1 fraction, which is shown in Fig. 2.
The regulation of apoptosis may be divided into
three main pathways: i) The extrinsic pathway or death receptor
pathway; ii) the intrinsic mitochondrial pathway; and iii) the ER
pathway (57). In the extrinsic
pathway, binding of Fas ligand to Fas on the cell surface will
recruit the adapter protein FADD and procaspase-8 to form
death-inducing signaling complex (DISC) (58,59). In
DISC, procaspase-8 will become activated, forming activated
caspase-8, which can directly activate effector caspases-3, −6 and
−7, leading to apoptotic cell death (58,59). Our
in vitro study demonstrated that forbesione increased the
protein expression of Fas, FADD, procaspase-8 and activated
caspase-3, suggesting apoptosis induction through the extrinsic
pathway.
The mitochondrial apoptotic pathway is regulated by
the Bcl-2 family of proteins, including the anti-apoptotic members
Bcl-2 and Bcl-extra large and the pro-apoptotic members Bax and Bak
(60). The equilibrium between the
levels of pro-apoptotic and anti-apoptotic proteins is important
for cell survival and death (60). An
increased ratio of Bax/Bcl-2 can trigger the release of cytochrome
c from mitochondria into the cytoplasm (61). Cytochrome c will form a complex with
Apaf-1 and procaspase-9, resulting in the activation of caspase-9
and caspase-3, leading to apoptotic cell death (61). Survivin, one of the human inhibitors
of apoptotic proteins, has been reported to inhibit activated
caspase-7 and caspase-3 (62,63). In accord with our previous study
(24), the present study demonstrated
that the forbesione-mediated induction of apoptosis in Ham-1 cells
was associated with decreased levels of Bcl-2 and survivin and with
increased levels of Bax, activated caspase-9 and activated
caspase-3, supporting that apoptosis induction is occurring through
the mitochondrial pathway. These results agree with the results of
our in vivo study, in which forbesione-mediated suppression
of tumor growth occurred through reduced expression of Bcl-2 and
increased expression of Bax, caspase-9 and caspase-3.
Upon encountering ER stress, the ER pathway is
triggered by altered conformation of Bax and Bak in the ER
membrane, resulting in Ca2+ efflux that activates the
calcium-dependent protease m-calpain in the cytoplasm (36). Subsequently, m-calpain cleaves and
activates the ER-resident protein procaspase-12, which subsequently
activates procaspase-9 and procaspase-3, thus promoting apoptotic
cell death (37). In the present
study, forbesione was able to decrease the expression of
procaspase-12 while increasing the expression of activated
caspase-12, suggesting that the induction of apoptosis was mediated
in part through the ER pathway.
NF-κB is a transcription factor that controls the
expression of several genes involved in the control of cell cycle
progression and apoptosis (64).
NF-κB consists of a complex of p50 and p65 subunits, and is present
in an inactive form when bound to an endogenous inhibitor, IκB, in
the cytoplasm (64). In response to
stimuli, IκB-α kinase (IKK) phosphorylates IκB, leading to its
degradation, and consequently NF-κB translocates into the nucleus
to activate the transcription of its target genes (40,65).
Previous studies demonstrated that isomorellin and gambogic acid
inhibit NF-κB activation through decreased NF-κB protein
expression, and that the inhibition of IKK activation leads to
suppressed phosphorylation and degradation of IκB-α as well as to
the suppression of the nuclear translocation of NF-κB/p65 (17,21). The
role of NF-κB in cell cycle progression has been previously
reported (61). NF-κB is able to
induce the expression of activators of cell cycle progression such
as cyclin D3, cyclin E and cyclin A (66). NF-κB also induces the expression of
anti-apoptotic genes such as Bcl-2 and survivin (67). Consistent with those studies, the
present study identified a decrease in NF-κB protein expression and
an increase in IκB-α protein expression, which could possibly cause
the inactivation of NF-κB, resulting in a decrease in protein
expression of cyclin E and cyclin A, thus leading to cell cycle
arrest in S phase and to a decrease in protein expression of Bcl-2
and survivin, thus resulting in apoptosis.
Our in vivo study revealed that forbesione
treatment significantly inhibited the growth of Ham-1 allografts.
The forbesione-treated hamsters were healthy (with normal physical
activities and eating habits) and gained weight. Examination of
tissue sections demonstrated no obvious histopathological changes
in the vital organs (liver, kidney and stomach) of the hamsters
treated daily with 50 mg/kg (orally). The liver and kidney
functions were normal. Our results confirm those of a previous
study in which gambogic acid at doses of 30 and 60 mg/kg
(intragastrically) exhibited potent antitumor effects without
obvious side effects or toxicity in preclinical experiments
(8). However, gambogic acid at a dose
of 120 mg/kg in a long-term follow-up study on rats exhibited toxic
effects in the liver and kidney (8).
In conclusion, our study demonstrates for the first
time that forbesione inhibits the growth of Ham-1 CCA cells in
vitro and suppresses allograft tumor growth in vivo. We
propose that forbesione acts via multiple mechanisms, including
induction of S-phase cell cycle arrest and multiple pathways of
apoptosis (including the death receptor pathway, the mitochondrial
pathway and the ER pathway), by altering the expression of genes
and proteins that are related to cell cycle and apoptosis
regulation. There were no observed side effects or toxicity in
hamsters treated with forbesione for 4 weeks, suggesting that
forbesione may represent a promising therapeutic drug for the
treatment of cancer. Additional studies leading to clinical trials
are merited.
Acknowledgements
The present study was supported in part by grants
from the Center of Excellence for Innovation in Chemistry,
Commission on Higher Education (Bangkok, Thailand; grant no.
48-03-3-00-144), Khon Kaen University Research Fund (Khon Kaen,
Thailand; grant no. 573301582701) and Khon Kaen University under
the Incubation Researcher Project, Khon Kaen University (Khon Kaen,
Thailand; grant no. 563902).
References
|
1
|
Sripa B and Pairojkul C:
Cholangiocarcinoma: Lessons from Thailand. Curr Opin Gastroenterol.
24:349–356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Welzel TM, McGlynn KA, Hsing AW, O'Brien
TR and Pfeiffer RM: Impact of classification of hilar
cholangiocarcinomas (Klatskin tumors) on the incidence of intra-and
extrahepatic cholangiocarcinoma in the United States. J Natl Cancer
Inst. 98:873–875. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
West J, Wood H, Logan R, Quinn M and
Aithal G: Trends in the incidence of primary liver and biliary
tract cancers in England and Wales 1971–2001. Br J Cancer.
94:1751–1758. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Patt YZ, Hassan MM, Aguayo A, Nooka AK,
Lozano RD, Curley SA, Vauthey JN, Ellis LM, Schnirer II, Wolff RA,
et al: Oral capecitabine for the treatment of hepatocellular
carcinoma, cholangiocarcinoma, and gallbladder carcinoma. Cancer.
101:578–586. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thongprasert S: The role of chemotherapy
in cholangiocarcinoma. Ann Oncol. 16:ii93–ii96. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hahnvajanawong C, Boonyanugomol W,
Nasomyon T, Loilome W, Namwat N, Anantachoke N, Tassaneeyakul W,
Sripa B, Namwat W and Reutrakul V: Apoptotic activity of caged
xanthones from Garcinia hanburyi in cholangiocarcinoma cell lines.
World J Gastroenterol. 16:2235–2243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saralamp P, Chuakul W, Temsiririrkkul R
and Clayton T: Medicinal Plants in Thailand. 1. Amarin Printing and
Publishing Public Co., Ltd.; Bangkok: 1996
|
|
8
|
Wu ZQ, Guo QL, You QD, Zhao L and Gu HY:
Gambogic acid inhibits proliferation of human lung carcinoma SPC-A1
cells in vivo and in vitro and represses telomerase activity and
telomerase reverse transcriptase mRNA expression in the cells. Bio
Pharm Bull. 27:1769–1774. 2004. View Article : Google Scholar
|
|
9
|
Han QB, Wang YL, Yang L, Tso TF, Qiao CF,
Song JZ, Xu LJ, Chen SL, Yang DJ and Xu HX: Cytotoxic
polyprenylated xanthones from the resin of Garcinia hanburyi. Chem
Pharm Bull. 54:265–267. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reutrakul V, Anantachoke N, Pohmakotr M,
Jaipetch T, Sophasan S, Yoosook C, Kasisit J, Napaswat C, Santisuk
T and Tuchinda P: Cytotoxic and anti-HIV-1 caged xanthones from the
resin and fruits of garcinia hanburyi. Planta Med. 73:33–40. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo QL, You QD, Wu ZQ, Yuan ST and Zhao L:
General gambogic acids inhibited growth of human hepatoma SMMC-7721
cells in vitro and in nude mice. Acta Pharmacol Sin. 25:769–774.
2004.PubMed/NCBI
|
|
12
|
Liu W, Guo QL, You QD, Zhao L, Gu HY and
Yuan ST: Anticancer effect and apoptosis induction of gambogic acid
in human gastric cancer line BGC-823. World J Gastroenterol.
11:3655–3659. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao L, Guo QL, You QD, Wu ZQ and Gu HY:
Gambogic acid induces apoptosis and regulates expressions of Bax
and Bcl-2 protein in human gastric carcinoma MGC-803 cells. Bio
Pharm Bull. 27:998–1003. 2004. View Article : Google Scholar
|
|
14
|
Qin Y, Meng L, Hu C, Duan W, Zuo Z, Lin L,
Zhang X and Ding J: Gambogic acid inhibits the catalytic activity
of human topoisomerase IIalpha by binding to its ATPase domain. Mol
Cancer Ther. 6:2429–2440. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu J, Guo QL, You QD, Lin SS, Li Z, Gu HY,
Zhang HW, Tan Z and Wang X: Repression of telomerase reverse
transcriptase mRNA and hTERT promoter by gambogic acid in human
gastric carcinoma cells. Cancer Chemother Pharmacol. 58:434–443.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu J, Guo QL, You QD, Zhao L, Gu HY, Yang
Y, Zhang HW, Tan Z and Wang X: Gambogic acid-induced G2/M phase
cell-cycle arrest via disturbing CDK7-mediated phosphorylation of
CDC2/p34 in human gastric carcinoma BGC-823 cells. Carcinogenesis.
28:632–638. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pandey MK, Sung B, Ahn KS, Kunnumakkara
AB, Chaturvedi MM and Aggarwal BB: Gambogic acid, a novel ligand
for transferrin receptor, potentiates TNF-induced apoptosis through
modulation of the nuclear factor-KappaB signaling pathway. Blood.
110:3517–3525. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo Q, Qi Q, You Q, Gu H, Zhao L and Wu Z:
Toxicological studies of gambogic acid and its potential targets in
experimental animals. Basic Clin Pharmacol Toxicol. 99:178–184.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qi Q, You Q, Gu H, Zhao L, Liu W, Lu N and
Guo Q: Studies on the toxicity of gambogic acid in rats. J
Ethnopharmacol. 117:433–438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qi Q, Gu H, Yang Y, Lu N, Zhao J, Liu W,
Ling H, You QD, Wang X and Guo Q: Involvement of matrix
metalloproteinase 2 and 9 in gambogic acid induced suppression of
MDA-MB-435 human breast carcinoma cell lung metastasis. J Mol Med
(Berl). 86:1367–1377. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hahnvajanawong C, Ketnimit S,
Pattanapanyasat K, Anantachoke N, Sripa B, Pinmai K, Seubwai W and
Reutrakul V: Involvement of p53 and nuclear factor-kappaB signaling
pathway for the induction of G1-phase cell cycle arrest of
cholangiocarcinoma cell lines by isomorellin. Bio Pharm Bull.
35:1914–1925. 2012.
|
|
22
|
Hahnvajanawong C, Wattanawongdon W,
Chomvarin C, Anantachoke N, Kanthawong S, Sripa B and Reutrakul V:
Synergistic effects of isomorellin and forbesione with doxorubicin
on apoptosis induction in human cholangiocarcinoma cell lines.
Cancer Cell Int. 14:682014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Puthdee N, Vaeteewoottacharn K, Seubwai W,
Wonkchalee O, Kaewkong W, Juasook A, Pinlaor S, Pairojkul C,
Wongkham C, Okada S, et al: Establishment of an allo-transplantable
hamster cholangiocarcinoma cell line and its application for in
vivo screening of anti-cancer drugs. Korean J Parasitol.
51:711–717. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pinmai K, Chunlaratthanabhorn S,
Ngamkitidechakul C, Soonthornchareon N and Hahnvajanawong C:
Synergistic growth inhibitory effects of Phyllanthus emblica and
Terminalia bellerica extracts with conventional cytotoxic agents:
Doxorubicin and cisplatin against human hepatocellular carcinoma
and lung cancer cells. World J Gastroenterol. 14:1491–1497. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boonmars T, Srisawangwong T, Srirach P,
Kaewsamut B, Pinlaor S and Sithithaworn P: Apoptosis-related gene
expressions in hamsters re-infected with Opisthorchis viverrini and
re-treated with praziquantel. Parasitol Res. 102:57–62. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Boonmars T, Wu Z, Nagano I and Takahashi
Y: Expression of apoptosis-related factors in muscles infected with
Trichinella spiralis. Parasitology. 128:323–332. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee SO, Nadiminty N, Wu XX, Lou W, Dong Y,
Ip C, Onate SA and Gao AC: Selenium disrupts estrogen signaling by
altering estrogen receptor expression and ligand binding in human
breast cancer cells. Cancer Res. 65:3487–3492. 2005.PubMed/NCBI
|
|
29
|
Wu Z, Boonmars T, Nagano I, Boonjaraspinyo
S, Pinlaor S, Pairojkul C, Chamgramol Y and Takahashi Y: Alteration
of galectin-1 during tumorigenesis of Opisthorchis viverrini
infection-induced cholangiocarcinoma and its correlation with
clinicopathology. Tumour Biol. 33:1169–1178. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wonkchalee O, Boonmars T, Aromdee C,
Laummaunwai P, Khunkitti W, Vaeteewoottacharn K, Sriraj P,
Aukkanimart R, Loilome W, Chamgramol Y, et al: Anti-inflammatory,
antioxidant and hepatoprotective effects of Thunbergia laurifolia
Linn. on experimental opisthorchiasis. Parasitol Res. 111:353–359.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Baskić D, Popović S, Ristić P and
Arsenijević NN: Analysis of cycloheximide-induced apoptosis in
human leukocytes: Fluorescence microscopy using annexin V/propidium
iodide versus acridin orange/ethidium bromide. Cell Biol Int.
30:924–932. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grimm S and Brdiczka D: The permeability
transition pore in cell death. Apoptosis. 12:841–855. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cryns V and Yuan J: Proteases to die for.
Genes Dev. 12:1551–1570. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Burlacu A: Regulation of apoptosis by
Bcl-2 family proteins. J Cell Mol Med. 7:249–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Szegezdi E, Logue SE, Gorman AM and Samali
A: Mediators of endoplasmic reticulum stress-induced apoptosis.
EMBO Rep. 7:880–885. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu J and Kaufman R: From acute ER stress
to physiological roles of the unfolded protein response. Cell Death
Differ. 13:374–384. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guttridge DC, Albanese C, Reuther JY,
Pestell RG and Baldwin AS Jr: NF-kappaB controls cell growth and
differentiation through transcriptional regulation of cyclin D1.
Mol Cell Biol. 19:5785–5799. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Perkins ND: The Rel/NF-kappa B family:
Friend and foe. Trends Biochem Sci. 25:434–440. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sriraj P, Aukkanimart R, T Juasook A
Boonmars, Sudsarn P, Wonkchalee N, Pairojkul C, Waraasawapati S,
Laummaunwai P and Boonjaraspinyo S: Does a combination of
opisthorchiasis and ethyl alcohol consumption enhance early
cholangiofibrosis, the risk of cholangiocarcinoma? Parasitol Res.
112:2971–2981. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao W, Zhou SF, Zhang ZP, Xu GP, Li XB
and Yan JL: Gambogic acid inhibits the growth of osteosarcoma cells
in vitro by inducing apoptosis and cell cycle arrest. Oncol Rep.
25:1289–1295. 2011.PubMed/NCBI
|
|
42
|
Chen J, Gu HY, Lu N, Yang Y, Liu W, Qi Q,
Rong JJ, Wang XT, You QD and Guo QL: Microtubule depolymerization
and phosphorylation of c-Jun N-terminal kinase-1 and p38 were
involved in gambogic acid induced cell cycle arrest and apoptosis
in human breast carcinoma MCF-7 cells. Life Sci. 83:103–109. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Graña X and Reddy EP: Cell cycle control
in mammalian cells: Role of cyclins, cyclin dependent kinases
(CDKs), growth suppressor genes and cyclin-dependent kinase
inhibitors (CKIs). Oncogene. 11:211–219. 1995.PubMed/NCBI
|
|
44
|
King K and Cidlowski J: Cell cycle
regulation and apoptosis 1. Annu Rev Physiol. 60:601–617. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yadav V, Sultana S, Yadav J and Saini N:
Gatifloxacin induces S and G2-phase cell cycle arrest in pancreatic
cancer cells via p21/p27/p53. PLoS One. 7:e477962012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Maga G and Hubscher U: Proliferating cell
nuclear antigen (PCNA): A dancer with many partners. J Cell Sci.
116:3051–3060. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Engel FB, Hauck L, Boehm M, Nabel EG,
Dietz R and von Harsdorf R: P21(CIP1) controls proliferating cell
nuclear antigen level in adult cardiomyocytes. Mol Cell Bio.
23:555–565. 2003. View Article : Google Scholar
|
|
48
|
Steller H: Mechanisms and genes of
cellular suicide. Science. 267:1445–1449. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nagata S: Apoptotic DNA fragmentation. Exp
Cell Res. 256:12–18. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kajstura M, Halicka HD, Pryjma J and
Darzynkiewicz Z: Discontinuous fragmentation of nuclear DNA during
apoptosis revealed by discrete ‘sub-G1’ peaks on DNA content
histograms. Cytometry A. 71:125–131. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Franz S, Frey B, Sheriff A, Gaipl US, Beer
A, Voll RE, Kalden JR and Herrmann M: Lectins detect changes of the
glycosylation status of plasma membrane constituents during late
apoptosis. Cytometry A. 69:230–239. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Oberhammer F, Wilson J, Dive C, Morris ID,
Hickman JA, Wakeling AE, Walker PR and Sikorska M: Apoptotic death
in epithelial cells: Cleavage of DNA to 300 and/or 50 kb fragments
prior to or in the absence of internucleosomal fragmentation. EMBO
J. 12:3679–3984. 1993.PubMed/NCBI
|
|
54
|
Darzynkiewicz Z, Juan G, Li X, Gorczyca W,
Murakami T and Traganos F: Cytometry in cell necrobiology: Analysis
of apoptosis and accidental cell death (necrosis). Cytometry.
27:1–20. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Huang X, Halicka HD, Traganos F, Tanaka T,
Kurose A and Darzynkiewicz Z: Cytometric assessment of DNA damage
in relation to cell cycle phase and apoptosis. Cell Prolif.
38:223–243. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Jakubikova J, Bao Y and Sedlak J:
Isothiocyanates induce cell cycle arrest, apoptosis and
mitochondrial potential depolarization in HL-60 and
multidrug-resistant cell lines. Anticancer Res. 25:3375–3386.
2005.PubMed/NCBI
|
|
57
|
Du Y, Wang K, Fang H, Li J, Xiao D, Zheng
P, Chen Y, Fan H, Pan X, Zhao C, Zhang Q, et al: Coordination of
intrinsic, extrinsic, and endoplasmic reticulum-mediated apoptosis
by imatinib mesylate combined with arsenic trioxide in chronic
myeloid leukemia. Blood. 107:1582–1590. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Choi C and Benveniste EN: Fas ligand/Fas
system in the brain: Regulator of immune and apoptotic responses.
Brain Res Rev. 44:65–81. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chatfield K and Eastman A: Inhibitors of
protein phosphatases 1 and 2A differentially prevent intrinsic and
extrinsic apoptosis pathways. Biochem Biophys Res Commun.
323:1313–1320. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liang X, Wu A, Xu Y, Xu K, Liu J and Qian
X: B1, a novel naphthalimide-based DNA intercalator, induces cell
cycle arrest and apoptosis in HeLa cells via p53 activation. Invest
New Drugs. 29:646–658. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fulda S and Debatin K: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Deveraux QL and Reed JC: IAP family
proteins-suppressors of apoptosis. Genes Dev. 13:239–252. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shin S, Sung BJ, Cho YS, Kim HJ, Ha NC,
Hwang JI, Chung CW, Jung YK and Oh BH: An anti-apoptotic protein
human survivin is a direct inhibitor of caspase-3 and-7.
Biochemistry. 40:1117–1123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Baldwin AS Jr: The NF-kappaB and I kappaB
proteins: New discoveries and insights. Annu Rev Immunol.
14:649–683. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ghosh S and Karin M: Missing pieces in the
NF-kappaB puzzle. Cell. 109:(Suppl). S81–S96. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hsia CY, Cheng S, Owyang AM, Dowdy SF and
Liou HC: C-Rel regulation of the cell cycle in primary mouse B
lymphocytes. Int Immunol. 14:905–916. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tracey L, Perez-Rosado A, Artiga MJ,
Pérez-Rosado A, Artiga MJ, Camacho FI, Rodríguez A, Martínez N,
Ruiz-Ballesteros E, Mollejo M, et al: Expression of the NF-kappaB
targets BCL2 and BIRC5/Survivin characterizes small B-cell and
aggressive B-cell lymphomas, respectively. J Pathol. 206:123–134.
2005. View Article : Google Scholar : PubMed/NCBI
|