Introduction
MicroRNAs (miRNAs) are small (18–25 nt), non-coding
RNAs of endogenous origin. To date, >2,500 different mature
miRNA species have been detected in humans (1). Following maturation, double-stranded
miRNAs are immediately bound to Argonaute proteins and unwound. In
the single-stranded conformation, they build up the active
component of the RNA-induced silencing complex (RISC) (2,3). miRNAs
bind to complementary sequences in the 3′untranslated region (UTR)
of their target mRNAs, thus acting as translational repressors.
This action is primarily performed by a hexamer sequence at the
5′UTRs called the seed sequence and the subsequent translocation to
the P-bodies (processing bodies), which are specialized cellular
components for RNA silencing and decay (4,5). Due to
the simple target recognition sequence and the vast number of
different species of miRNA, it is estimated that ~60% of the human
protein-coding genes are regulated post-transcriptionally by miRNAs
(6). Due to their high abundance and
immense impact on cellular gene expression profiles, miRNAs serve
an important role in cellular proliferation and differentiation
processes. Furthermore, they contribute to the adaptation to
cellular stress, including stress caused by an acidic or hypoxic
environment (7,8).
Hypoxia, the state of decreased oxygen saturation in
a tissue, is a major problem in the treatment of solid tumors. Due
to rapid growth, regions in solid tumors tend to be cut off from
oxygen maintenance by blood vessels. The tumor cells must adapt to
this new environmental challenge, leading to the clonal selection
of successful tumor cells, increased aggressiveness and decreased
chemo- and radiosensitivity of the tumor.
There are several studies describing the effect of
hypoxia on miRNA expression. Kulshreshthra et al (9,10) have
demonstrated that the miRNAs miR-210, miR-23a, miR-24-1, miR-26b
and miR-107 are overexpressed in hypoxic conditions, while the
miRNAs miR-19a, miR-122a, miR-141, miR-101, miR-186, miR-374,
miR-424, miR-422b, miR-320, miR-29b and miR-197 are all
downregulated.
The most prominent hypoxia-induced miRNA is the
ubiquitously expressed miR-210 (11).
In the majority of cell lines studied so far, hypoxia triggers the
upregulation of miR-210 (12).
Furthermore, high expression of miR-210 is associated with a poorer
outcome for patients with mammary carcinoma, head and neck
carcinoma, pancreatic adenocarcinoma or soft tissue sarcoma
(13–16). Thus, under hypoxic conditions, miR-210
may act as an oncomir, promoting cell survival and adaptation to
the changed environment and leading to increased tumor
aggressiveness. Hypoxia-inducible miRNAs, including miR-210, may be
antagonized by hypoxia-downregulated microRNAs, such as miR-199a.
miR-199a is located within a cluster together with miR-214, which
is regulated concordantly by diverse transcription factors,
including TWIST1 (17).
miR-199a-5p regulates hypoxia-inducible factor 1α (HIF-1α)
and Sirt mRNA, thus suppressing adaptation to persistent
hypoxic conditions (18).
Furthermore, expression of the homologous miR-199b is inversely
correlated to HIF-1α protein expression in hepatocellular carcinoma
(HCC) and its elevated expression predicts an improved outcome for
patients with HCC (19). Recently, a
self-regulatory network in testicular germ cell tumors was
investigated and it was demonstrated that p53 is activated
and aids in the positive regulation of miR-199a-2/miR-214 by
repressing DNMT1 (20).
In the current study, the impact of miR-199a-5p
expression on the clinical parameters and outcome of patients with
soft tissue sarcoma was evaluated. Several studies have highlighted
the association of miR-199a-5p downregulation with the cellular
hypoxia response (18,21–23).
Furthermore, hypoxia is an important factor in sarcomagenesis and
the increasing aggressiveness of sarcoma; therefore, the present
study evaluated the impact of miR-199a-5p on hypoxia-related genes
by retesting the regulation of HIF-1α and VEGF
through miR-199a-5p and additionally proposing oxidative stress
induced growth inhibitor 2 (OSGIN2) as a novel miR-199a-5p
target gene.
Materials and methods
Patients
The cohort consisted of 96 patients with soft tissue
sarcoma who underwent surgical tumor resection between March 1998
and April 2001 in the Department of Surgery, University of Leipzig
(Leipzig, Germany). None of the patients had received adjuvant
treatment prior to resection. Patients and tissue samples have been
described in previous studies (24,25).
Tissues were snap-frozen in liquid nitrogen immediately following
resection and stored at −80°C. The present study was approved by
the local ethics committee of the Martin Luther University
Halle-Wittenberg. All patients gave written informed consent.
RNA isolation
RNA was isolated using TriZol reagent (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA USA) according to the
manufacturer's protocol. Briefly, the tissue was cut in a cryotome
and 20–30 slices (30-µm thick) were lysed in TriZol reagent. The
tissue protein and DNA were precipitated by centrifugation. DNAse I
(Qiagen GmbH, Hilden, Germany) digestion was performed on the
flow-through for 30 min to eliminate remaining traces of DNA. RNA
was precipitated by isopropanol (Sigma-Aldrich, St. Louis, Mo, USA)
overnight at −20°C and washed twice with ice-cold ethanol prior to
elution in RNAse-free water.
miRNA synthesis and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
A total of 10 ng RNA was used for cDNA synthesis of
miR-199a-5p and U18 small nucleolar (sno) RNA (reference gene)
using stem-loop primers of the TaqMan® microRNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.). cDNA synthesis was performed by MuLV Reverse Transcriptase
kit (Fermentas; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. qPCR was performed with the HotStartTaq
DNA Polymerase kit (Qiagen GmbH) on a Rotorgene Cycler (LTF
Labortechnik, Wasserburg, Germany). U18 snoRNA values were uniform
and served as an internal reference. Cq values were
obtained and transformed via the ΔΔCq method (26) and the lowest Cq value
served as a calibrator (27).
Luciferase reporter assay
3′UTR regions of HIF-1α, OSGIN2 and
VEGF were amplified using the following primers:
HIF-1α, forward, 5′-AAACTCGAGTGGCATGTAGACTGCTGGGGCAA-3′ and
reverse, 5′-AAACTCGAGTGGCTACCACGTACTGCTGGCAA-3′; OSGIN2:
forward, 5′-AAACTCGAGTGGGGTTTTGCAGTGTACTGGCT-3′ and reverse,
5′-AAACTCGATGGACCCCACCCCCAGTTATACA-3′; VEGF forward,
5′-AAACTCGAGTGGACCACACCATCACCATCGAC-3′ and reverse,
5′-AAACTCGAGTGGCGTCTGACCTGGGGTAGAGA-3′. The 3′UTR sequences with
the putative miRNA binding sites were cloned into a psiCheck™-2
vector carrying a renilla luciferase and a constitutively expressed
firefly luciferase gene (Promega Corporation, Madison, WI, USA) by
applying restriction digestion with XhoI (Fermentas; Thermo
Fisher Scientific, Inc.) and ligation with Ligase I (Fermentas;
Thermo Fisher Scientific, Inc.). Cells from the human osteosarcoma
SAOS-2 cell line were cultivated for 24 h in Dulbecco's modified
Eagle's medium (DMEM) in 5% CO2, transfected with
psiCheck2-3′UTR-constructs and mimics for miR-199a-5p (Ambion;
Thermo Fisher Scientific, Inc.). miR-199a-5p mimics were
transfected as double-stranded RNA according to the manufacturer's
protocol, with the complementary miR-199a-3p strand inactivated by
appropriate chemical modifications. Luciferase activity
measurements with the Dual-Glo® Luciferase assay
(Promega Corporation) were performed after 24 h in a Tecan X50
reader (Tecan Austria GmbH, Grödig, Austria), to determine relative
chemiluminescence. Firefly luciferase activity served as an
internal reference. Relative values were standardized on the
luminescence values of the non-mimic transfected control.
Western blot analyses
For the analysis of miR-199a-5p-mediated translation
inhibition of the selected target genes, two sarcoma cell lines
were chosen as representative sarcoma systems. Rhabdomyosarcoma
(RD) and SAOS-2 cells were transfected with miR-199a-5p mimics or
scrambled RNA (Ambion; Thermo Fisher Scientific, Inc.),
respectively and cultivated for 24 h under hypoxic or normoxic
conditions in DMEM in 5% CO2. Cells were harvested on
ice immediately following reoxygenation using a cell scraper and
protein was isolated according to the RIPA method. For western blot
analyses, 20 ng of whole protein was separated on a sodium dodecyl
sulphate-polyacrylamide gel electrophoresis gel (4–12%; Thermo
Fisher Scientific, Inc.), blotted on a polyvinylidene fluoride
membrane and blocked with 5% dry milk/phosphate-buffered saline
(Carl Roth GmbH & Co. KG, Karlsruhe, Germany). Membranes were
incubated with murine anti-human OSGIN2 (dilution 1:500; #ab88829;
Abcam, Cambridge, UK), murine anti-human HIF-1α (dilution 1:1,000;
#610958; BD Transduction Laboratories™; BD Biosciences) or murine
anti-β-actin antibody (dilution 1:2,000; #A2228; Sigma-Aldrich) and
tagged with a secondary horseradish peroxidase-coupled antibody
(dilution 1:10,000; #P0260; Dako, Glostrup, Denmark). Detection was
performed with enhanced chemiluminescent Detection substrate
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol.
Statistical analysis
Statistical analyses were performed using SPSS 19.0
(IBM SPSS, Armonk, NY, USA). In detail, bivariate correlations
(Spearman-Rho), Kaplan-Meier analyses, uni- and multivariate Cox's
regression analyses (the latter was adjusted to tumor entity, tumor
localisation, resection type and tumor stage) were performed to
evaluate possible correlations between miR-199a-5p and clinical
parameters and outcome. To evaluate the luciferase reporter assay,
unpaired Student's t-tests were performed. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-199a-5p is associated with
clinical and molecular factors in soft tissue sarcoma
miR-199a-5p expression was quantified in 96 patients
with soft tissue sarcoma: 37 of these patients experienced ≥1
relapses, 29 experienced metastasis and 50 died of the tumor
(Table I). Bivariate correlation
analyses according to Spearman-Rho were performed to detect
significant associations between miR-199a-5p expression and
relevant clinical or molecular factors (Table II). Intriguingly, it was demonstrated
that miR-199a-5p expression is associated with the number and
interval of relapses (rs=0.39, P=0.00009 and
rs=0.52, P=3.6×10−8, respectively) as well as
the tumor-specific survival time (rs=0.36, P=0.00024).
There was an association between miR-199a-5p expression and
OSGIN2 mRNA expression, although this was not statistically
significant (rs=−0.23; P=0.09). Furthermore, there was a
correlatiojn between miR-199a-5p and HIF-1α mRNA expression
(r =0.27, P=0.018). These findings may suggest the existence of a
feedback-loop, induced by increased HIF-1α levels, leading to a
downregulation through miR-199a.
 | Table I.Clinical and histopathological
parameters and miR-199a-5p expression level. |
Table I.
Clinical and histopathological
parameters and miR-199a-5p expression level.
| Patient
characteristic | No. cases | miR-199a-5p
≤2.24 | miR-199a-5p
>2.24 | χ2 test
(P-value) |
|---|
| Total | 96 |
|
|
| Gender |
|
|
|
|
|
Male | 47 | 12 | 35 |
|
|
Female | 49 | 21 | 28 | n.s. |
| Histological
subtype |
|
|
|
|
| LS | 18 | 7 | 11 |
|
| FS | 7 | 1 | 6 |
|
|
RMS | 7 | 3 | 4 |
|
|
LMS | 17 | 9 | 8 |
|
| NS | 11 | 4 | 7 |
|
|
Syn | 7 | 0 | 7 |
|
|
NOS | 22 | 7 | 15 |
|
|
Other | 7 | 1 | 6 | n.s. |
| Tumor size |
|
|
|
|
| T1 | 21 | 3 | 18 |
|
| T2 | 75 | 29 | 46 | n.s. |
| Tumor
gradea |
|
|
|
|
| I | 13 | 6 | 7 |
|
| II | 41 | 9 | 32 |
|
|
III | 42 | 17 | 25 | n.s. |
| Tumor
stageb |
|
| I | 10 | 4 | 6 |
|
| II | 42 | 10 | 32 |
|
|
III | 34 | 12 | 22 |
|
| IV | 10 | 6 | 4 | n.s. |
| Complete
resection |
|
|
|
|
| Radical
(R0) | 62 | 20 | 42 |
|
| Not
radical (R1) | 34 | 12 | 22 | n.s. |
| Location |
|
|
|
|
|
Extremities | 61 | 18 | 43 |
|
| Trunk
wall | l1 | 3 | 8 |
|
|
Head/neck | 3 | 1 | 2 |
|
|
Abdomen/retro-peritoneum | 19 | 9 | 10 |
|
|
Multiple locations | 2 | 1 | 1 | n.s. |
| Number of
relapses |
|
|
|
|
| 0 | 59 | 22 | 37 |
|
| 1 | 17 | 5 | 12 |
|
|
>2 | 20 | 5 | 15 | n.s. |
| Metastasis |
|
|
|
|
|
Yes | 29 | 10 | 19 |
|
| No | 67 | 22 | 45 | n.s. |
| Patient status |
|
|
|
|
|
Alive | 46 | 11 | 35 |
|
|
Succumbed | 50 | 21 | 29 | n.s. (P=0.06) |
| Table II.Bivariate correlations (Spearman's
Rho test) between miR-199a-5p expression in tumor tissues and
different clinical and molecular factors of soft tissue sarcoma
patients. |
Table II.
Bivariate correlations (Spearman's
Rho test) between miR-199a-5p expression in tumor tissues and
different clinical and molecular factors of soft tissue sarcoma
patients.
| Parameter | rs | P-value | n |
|---|
| Age | −0.21 | 0.04a | 98 |
| Tumor-specific
survival time | 0.36 |
0.00024a | 98 |
| Number of
relapses | 0.39 |
0.00009a | 98 |
| Time interval of
relapse | 0.52 |
(3.6×10−8)a | 98 |
| HIF-1α mRNA | 0.266 | 0.018a | 85 |
Survival analyses
miR-199a-5p expression was transformed according to
a tercentile distribution (low expression, <2.24; medium
expression, 2.24–10.3; elevated expression, >10.4). A
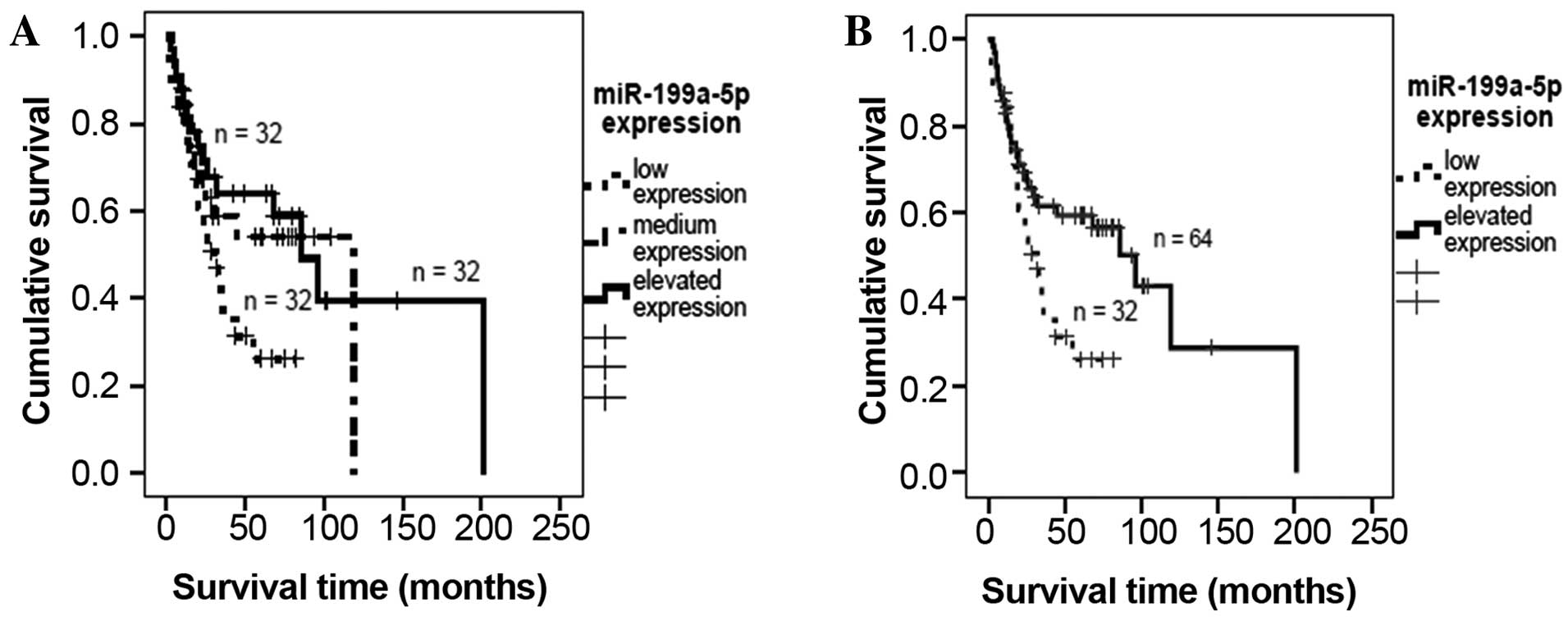
Kaplan-Meier analysis indicated that low miR-199a-5p expression was
associated with decreased patient survival time, although this
association was not significant (P=0.07, mean survival times: Low
expression, 38.3 months; medium expression, 72.2 months; elevated
expression, 105.9 months; Fig. 1A).
To investigate the effect of lower levels of miR-199a-5p expression
in detail, the groups of patients exhibiting medium and elevated
expression were combined. It was demonstrated that lower
miR-199a-5p expression was associated with a significant decrease
in survival time (P=0.026; median survival time for low expression,
38.3 months; for elevated expression, 95.6 months; Fig. 1B).
Furthermore, a univariate Cox's regression analysis
demonstrated that patients with low miR-199a expression exhibited
significantly poorer outcomes and were 1.92 times more likely to
experience tumor-associated mortality [P=0.029; 95% confidence
interval (CI), 1.07–3.44]. However, in a multivariate Cox's
regression analysis (adjusted to resection type, tumor stage,
entity and localisation), no correlation was detected between
miR-199a-5p expression and tumor-specific survival (P=0.66;
relative risk=1.16; 95% CI, 0.6–2.22).
OSGIN2 and HIF-1α are target-genes of
miR-199a-5p in vitro
To analyse possible linkages between miR-199a
expression and hypoxia-related target genes, an in silico
database search for putative 3′-UTR mRNA interactions of miR-199a
was performed. Beside others, TargetScan (www.targetscan.org) suggested HIF-1α,
VEGF and OSGIN2 mRNA as putative target genes of
miR-199a-5p (Fig. 2). The 3′UTR
sequences of these genes, including the predicted miR-199a-5p
target binding site, were cloned into psiCheck2 vectors and
luciferase reporter assays were performed to validate regulation by
miR-199a-5p. Co-transfection of SAOS-2 cells with 20 or 50 nM
miR-199a-5p mimics induced a significant downregulation of
HIF-1α and OSGIN2 3′UTR coupled luciferase expression
(Fig. 3A and B; 35.4 and 26.2%
respectively, compared to controls treated with scrambled mimics;
P=0.008 and P=0.01, respectively; Student's t-test).
However, no significant change in VEGF-3′UTR coupled
luciferase expression (105.6% compared to control, Fig. 3C) was detected.
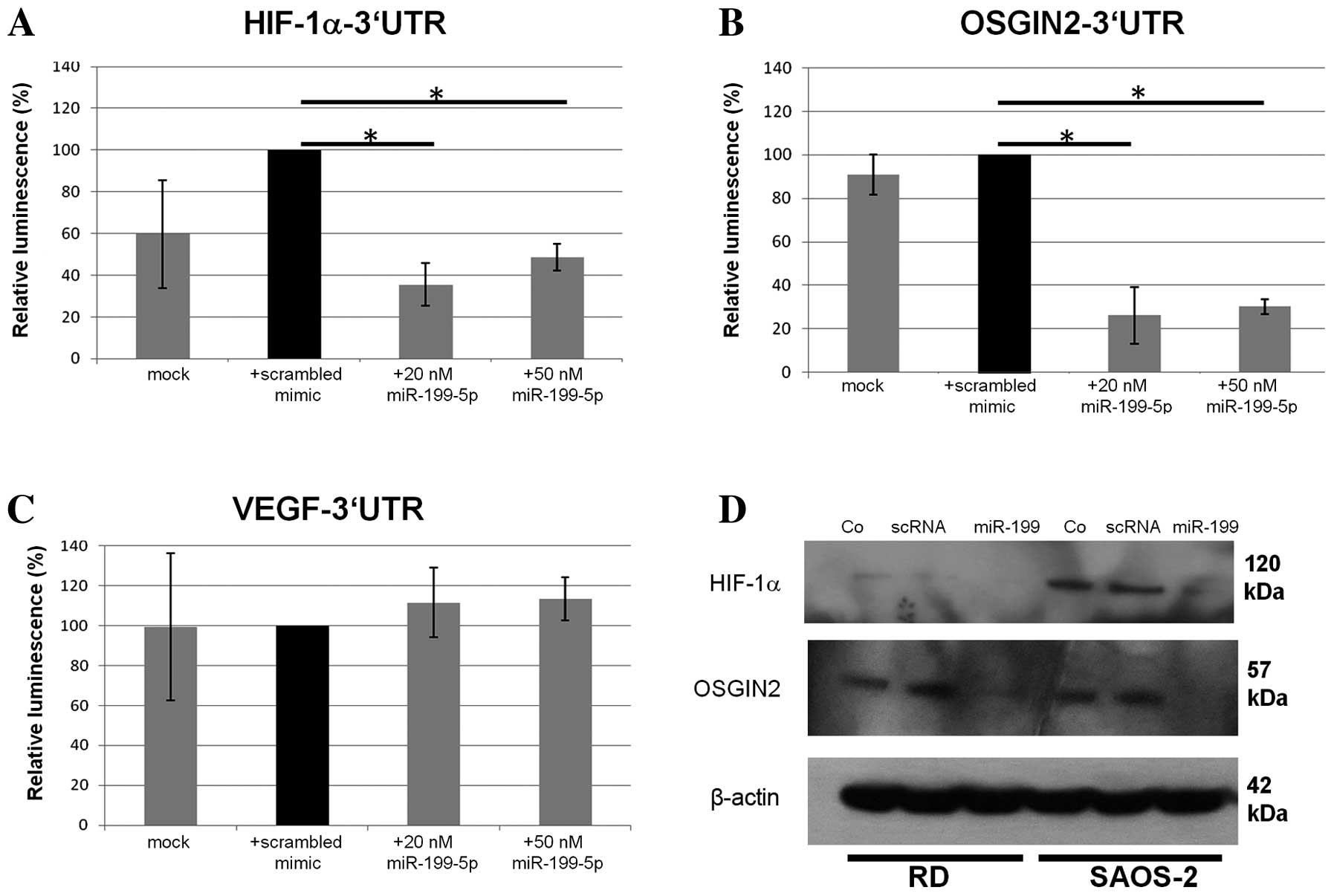 | Figure 3.Luciferase reporter assays with
reporter constructs containing (A) HIF-1α (B) OSGIN2
and (C) VEGF 3′UTRs. Relative luminescence coefficients were
standardized on a scrambled mimic transfected control.
Co-transfection with 20 nM miR-199a-5p yielded a significant
downregulation of the luciferase-coupled HIF-1α-3′UTR
construct (35.4%, P=0.041, Student's t-test) and the
OSGIN2-3′UTR construct (26.2%; P=0.029, Student's
t-test, error bars defining standard derivation) compared
with controls (n=3; *P<0.05). No such downregulation was
observed in VEGF 3′UTRs. (D) Western blot analyses presented a
downregulation of HIF-1α and OSGIN2 expression following
miR-199a-5p (20 nM) overexpression. HIF-1α, hypoxia-inducible
factor 1α OSGIN2, oxidative stress induced growth inhibitor 2;
VEGF, vascular endothelial growth factor; UTR, untranslated region;
scRNA, scrambled RNA; RD, Rhabdomyosarcoma cells. |
Furthermore, western blot analyses of HIF-1α and
OSGIN2 protein expression following miR-199a-5p transfection were
performed in RD and SAOS-2 cells under hypoxic conditions. HIF-1α
and OSGIN2 proteins were expressed in hypoxic conditions, whereas
no visible bands were detectable under normoxic conditions (data
not shown). In hypoxic conditions, transfection of miR-199a-5p
mimics resulted in the downregulation of HIF-1α and OSGIN2 protein
in comparison to untreated control cells or cells treated with
negative mimics only (Fig. 3D). Taken
together, these results suggest that miR-199a-5p downregulation
contributes to the adaptation of cells to hypoxia.
Discussion
The present study evaluated the impact of
miR-199a-5p on the survival of patients with soft tissue sarcoma
and validated HIF-1α and OSGIN2 as miR-199a-5p target
genes. Interestingly, OSGIN2 was linked for the first time
to miR-199a-5p regulation and may be an interesting target for
analyses of sarcomagenesis and progression, tumor hypoxia and
patient outcome.
The effect of the expression of several well-known
cancer genes on the mRNA and protein level is well known, however,
the data about the impact of most types of miRNA on the outcome of
patients with soft tissue sarcoma remains scarce. To the best of
our knowledge, the present study is first to report a 1.92-fold
increased risk of tumor-associated mortality for soft tissue
sarcoma patients (33% percentile) with low miR-199a-5p expression
(univariate Cox's regression analysis, P=0.029; 95% CI: 1.07–3.44).
Following multivariate Cox's Regression analysis, no effect of
miR-199a-5p expression on patients' survival was detected, however,
non-parametric correlation analyses demonstrated a correlation with
tumor entity (P=0.014, Kruskal-Wallis test). This is concordant
with the results of a previous study by Guled et al
(28), describing a diagnostic role
of miR-199-5p for the identification of undifferentiated
pleomorphic sarcoma compared to leiomyosarcoma. It has also been
demonstrated that low miR-199a-5p expression is associated with a
poorer outcome in serous ovarian carcinoma (29), non-small cell lung carcinoma measured
in patients' sera (30) and renal
cell carcinoma (31). Concordantly,
decreased expression of the complementary miRNA sequence
miR-199a-3p is associated with a shorter time to recurrence in
hepatocellular carcinoma (32).
Furthermore, multivariate Cox's regression analyses demonstrated
that low miR-199a-3p expression was significantly associated with
poorer overall survival in patients with osteosarcoma (33). However, previous studies have linked
the elevated expression of miR-199a-3p with poorer overall patient
survival in colorectal cancer (34)
and esophageal adenocarcinoma (35),
pointing towards a differential expression signature and possibly
altered regulation mechanisms of miR-199a processing and RISC
incorporation in tumor cells.
Several studies have described miR-199a as a type of
miRNA associated with hypoxia-adaptation (36–39). It
was demonstrated that miR-199a-5p downregulation is
AKT-dependent (39,40) and may be antagonized by the
β-adrenergic receptor (40). In the
current study, in order to facilitate luciferase reporter assays,
the interactions between miR-199a-5p and the 3′UTRs of three
putative hypoxia-related target genes, namely HIF-1α,
OSGIN2 and VEGF, were investigated. These target
genes were chosen following a literature search demonstrating the
potential link between miR-199a-5p expression and hypoxia. The
results of the present study demonstrated that miR-199a-5p
post-transcriptionally regulated HIF-1α and OSGIN2,
but not VEGF. Downregulation of miR-199a-5p in cardiac
myocytes mimics hypoxia preconditioning by lowering the
miR-199a-5p-mediated suppression of HIF-1α mRNA translation
(18). Furthermore, ethanol
downregulates miR-199a expression in liver-sinusoidal endothelial
liver cells independently of hypoxic stress, and this
downregulation coincides with an increase in HIF-1α and
Endothelin-1 mRNA (41). By contrast,
overexpression of miR-199a-5p and miR-199a-3p has been detected in
advanced liver fibrosis in samples from mouse and human tissue
(42). On a molecular level
miR-199a-5p targets the prominent oncogenes ERBB2 and
ERBB3 directly in ovarian and lung carcinoma cell lines
(43). Additionally, it was observed
that miR-199a-5p overexpression in ovarian carcinoma cells
significantly decreased their ability to induce angiogenesis, an
effect mediated via ERBB2 and ERBB3 signalling
(21). Moreover, overexpression of
miR-199a-5p in multiple myeloma cells significantly impaired
migration by downregulating the expression of adhesion molecules,
including vascular cell adhesion molecule-1 (VCAM-1) and
intracellular adhesion molecule 1 (ICAM-1) (39). By contrast, the complementary
miR-199a-3p transcript regulates the tumor-associated mammalian
target of rapamycin (mTOR) and c-Met in
hepatocellular carcinoma cells and its overexpression restores
hypoxia and doxorubicin sensitivity (32). Recently, Kinose et al (44) demonstrated that miR-199a-3p serves as
tumor suppressor gene in ovarian cancer by directly repressing its
target gene c-Met and subsequently inhibiting proliferation,
adhesion and invasiveness (44). The
present study validated miR-199a-5p as regulator of HIF-1α,
but not VEGF. These results are concordant with those from
previous studies demonstrating that VEGF is an indirect
target gene of miR-199a, which is regulated by HIF-1α and
ERBB2/3 downregulation rather than by direct
miRNA-medicated VEGF translation inhibition and mRNA
destabilization (21).
Furthermore, the results of the present study
suggested that OSGIN2 is a putative target of miR-199a-5p
and suggested a possible functional link between a hypoxic tumor
environment and the miRNA-induced derepression of stress genes
protecting tumor as well as normal cells from stress-induced cell
death. OSGIN2 is localized on chromosome 8q21.3 in the
neighbourhood of Nijmegen breakage syndrome gene 1 (45). The transcript is translated into a
protein 505 amino acids long and there is an additional shorter
transcript variant translated to a protein with a distinct
N-terminus (45). OSGIN2 is a
still poorly characterized homolog of OSGIN1, with 49%
identity and 62% similarity (46). A
previous study demonstrated that OSGIN2 was upregulated in
the liver biopsies of transplants exhibiting initial poor graft
function (47). Furthermore, in a
high-throughput assessment for known and novel breast cancer
candidate genes, OSGIN2 was mapped to a chromosomal region
demonstrating a significant amplification gain in breast cancer
cell lines and tumor tissue (48).
Additionally, it has been identified that OSGIN2 is
overexpressed following activation of PGC-1 related
coactivator, a protein maintaining mitochondrial homeostasis and
linking mitochondrial status to cell cycle (49). However, the physiological function of
OSGIN2, and its potential association with cancer genesis
and progression remain elusive and further studies are necessary to
clarify the impact of this gene on tumor hypoxia.
In conclusion, the present study identified a
correlation between low miR-199a-5p expression and a poorer outcome
of patients with soft tissue sarcoma. Low miR-199a-5p expression
increased the risk of tumor-associated mortality by 1.92-fold.
Furthermore, HIF-1α and OSGIN2 may be target genes of
miR-199a-5p. However, the precise role of miR-199a-5p in cancer
remains ambiguous, due to its diverging functions in different cell
types and stress situations. Thus, further studies, particularly on
the regulation and impact of this hypoxia-associated miRNA are
required.
Acknowledgements
The authors wish to thank Ms. Gabriele Thomas and
Ms. Katrin Theile for their excellent technical assistance. Thomas
Greither's work was supported by a junior research group grant of
Wilhelm-Roux program of the Medical faculty of Martin Luther
University Halle-Wittenberg (grant no. FKZ 25/43).
Glossary
Abbreviations
Abbreviations:
|
HIF-1α
|
hypoxia-inducible factor 1α
|
|
OSGIN2
|
oxidative stress induced growth
inhibitor 2
|
|
VEGF
|
vascular endothelial growth factor
|
|
RR
|
relative risk of tumor-related
death
|
|
p
|
probability
|
|
CI
|
95% confidence interval
|
|
rS
|
Spearman's rank correlation
coefficient
|
|
h
|
hour
|
|
UTR
|
untranslated region
|
References
|
1
|
Griffiths-Jones S: The microRNA registry.
Nucleic Acids Res. 32:D109–D111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim Y and Kim VN: MicroRNA factory: RISC
assembly from precursor microRNAs. Mol Cell. 46:384–386. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meister G, Landthaler M, Patkaniowska A,
Dorsett Y, Teng G and Tuschl T: Human Argonaute2 mediates RNA
cleavage targeted by miRNAs and siRNAs. Mol Cell. 15:185–197. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fabian MR, Mathonnet G, Sundermeier T,
Mathys H, Zipprich JT, Svitkin YV, Rivas F, Jinek M, Wohlschlegel
J, Doudna JA, et al: Mammalian miRNA RISC recruits CAF1 and PABP to
affect PABP-dependent deadenylation. Mol Cell. 35:868–880. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Emde A and Hornstein E: miRNAs at the
interface of cellular stress and disease. EMBO J. 33:1428–1437.
2014.PubMed/NCBI
|
|
8
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kulshreshtha R, Davuluri RV, Calin GA and
Ivan M: A microRNA component of the hypoxic response. Cell Death
Differ. 15:667–671. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kulshreshtha R, Ferracin M, Negrini M,
Calin GA, Davuluri RV and Ivan M: Regulation of microRNA
expression: The hypoxic component. Cell Cycle. 6:1426–1431. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang X and Zuo J: Emerging roles of
miR-210 and other non-coding RNAs in the hypoxic response. Acta
Biochim Biophys Sin (Shanghai). 46:220–232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Devlin C, Greco S, Martelli F and Ivan M:
miR-210: More than a silent player in hypoxia. IUBMB Life.
63:94–100. 2011.PubMed/NCBI
|
|
13
|
Camps C, Buffa FM, Colella S, Moore J,
Sotiriou C, Sheldon H, Harris AL, Gleadle JM and Ragoussis J:
Hsa-miR-210 is induced by hypoxia and is an independent prognostic
factor in breast cancer. Clin Cancer Res. 14:1340–1348. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gee HE, Camps C, Buffa FM, Patiar S,
Winter SC, Betts G, Homer J, Corbridge R, Cox G, West CM, et al:
Hsa-mir-210 is a marker of tumor hypoxia and a prognostic factor in
head and neck cancer. Cancer. 116:2148–2158. 2010.PubMed/NCBI
|
|
15
|
Greither T, Grochola LF, Udelnow A,
Lautenschläger C, Würl P and Taubert H: Elevated expression of
microRNAs 155, 203, 210 and 222 in pancreatic tumors is associated
with poorer survival. Int J Cancer. 126:73–80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Greither T, Würl P, Grochola L, Bond G,
Bache M, Kappler M, Lautenschläger C, Holzhausen HJ, Wach S, Eckert
AW and Taubert H: Expression of microRNA 210 associates with poor
survival and age of tumor onset of soft-tissue sarcoma patients.
Int J Cancer. 130:1230–1235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baumgarten A, Bang C, Tschirner A,
Engelmann A, Adams V, von Haehling S, Doehner W, Pregla R, Anker
MS, Blecharz K, et al: TWIST1 regulates the activity of ubiquitin
proteasome system via the miR-199/214 cluster in human end-stage
dilated cardiomyopathy. Int J Cardiol. 168:1447–1452. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rane S, He M, Sayed D, Vashistha H,
Malhotra A, Sadoshima J, Vatner DE, Vatner SF and Abdellatif M:
Downregulation of miR-199a derepresses hypoxia-inducible
factor-1alpha and Sirtuin 1 and recapitulates hypoxia
preconditioning in cardiac myocytes. Circ Res. 104:879–886. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang C, Song B, Song W, Liu J, Sun A, Wu
D, Yu H, Lian J, Chen L and Han J: Underexpressed microRNA-199b-5p
targets hypoxia-inducible factor-1α in hepatocellular carcinoma and
predicts prognosis of hepatocellular carcinoma patients. J
Gastroenterol Hepatol. 26:1630–1637. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen BF, Suen YK, Gu S, Li L and Chan WY:
A miR-199a/miR-214 self-regulatory network via PSMD10, TP53 and
DNMT1 in testicular germ cell tumor. Sci Rep. 4:64132014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He J, Jing Y, Li W, Qian X, Xu Q, Li FS,
Liu LZ, Jiang BH and Jiang Y: Roles and mechanism of miR-199a and
miR-125b in tumor angiogenesis. PLoS One. 8:e566472013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ding G, Huang G, Liu HD, Liang HX, Ni YF,
Ding ZH, Ni GY and Hua HW: MiR-199a suppresses the hypoxia-induced
proliferation of non-small cell lung cancer cells through targeting
HIF1α. Mol Cell Biochem. 384:173–180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang X, Lei S, Long J, Liu X and Wu Q:
MicroRNA-199a-5p inhibits tumor proliferation in melanoma by
mediating HIF-1α. Mol Med Rep. 13:5241–5247. 2016.PubMed/NCBI
|
|
24
|
Kappler M, Köhler T, Kampf C,
Diestelkötter P, Würl P, Schmitz M, Bartel F, Lautenschläger C,
Rieber EP, Schmidt H, et al: Increased survivin transcript levels:
An independent negative predictor of survival in soft tissue
sarcoma patients. Int J Cancer. 95:360–363. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Würl P, Kappler M, Meye A, Bartel F,
Köhler T, Lautenschläger C, Bache M, Schmidt H and Taubert H:
Co-expression of survivin and TERT and risk of tumour-related death
in patients with soft-tissue sarcoma. Lancet. 359:943–945. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shi R and Chiang VL: Facile means for
quantifying microRNA expression by real-time PCR. Biotechniques.
39:519–525. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guled M, Pazzaglia L, Borze I, Mosakhani
N, Novello C, Benassi MS and Knuutila S: Differentiating soft
tissue leiomyosarcoma and undifferentiated pleomorphic sarcoma: A
miRNA analysis. Genes Chromosomes Cancer. 53:693–702. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nam EJ, Yoon H, Kim SW, Kim H, Kim YT, Kim
JH, Kim JW and Kim S: MicroRNA expression profiles in serous
ovarian carcinoma. Clin Cancer Res. 14:2690–2695. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sanfiorenzo C, Ilie MI, Belaid A, Barlési
F, Mouroux J, Marquette CH, Brest P and Hofman P: Two panels of
plasma microRNAs as non-invasive biomarkers for prediction of
recurrence in resectable NSCLC. PLoS One. 8:e545962013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Si T, Liu C, Xu K and Gui Y: Association
of miR-199a expression with clinicopathologic characteristics and
prognosis of renal cell carcinoma. Nan Fang Yi Ke Da Xue Xue Bao.
32:1568–1571. 2012.(In Chinese). PubMed/NCBI
|
|
32
|
Fornari F, Milazzo M, Chieco P, Negrini M,
Calin GA, Grazi GL, Pollutri D, Croce CM, Bolondi L and Gramantieri
L: MiR-199a-3p regulates mTOR and c-Met to influence the
doxorubicin sensitivity of human hepatocarcinoma cells. Cancer Res.
70:5184–5193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tian R, Xie X, Han J, Luo C, Yong B, Peng
H, Shen J and Peng T: miR-199a-3p negatively regulates the
progression of osteosarcoma through targeting AXL. Am J Cancer Res.
4:738–750. 2014.PubMed/NCBI
|
|
34
|
Wan D, He S, Xie B, Xu G, Gu W, Shen C, Hu
Y, Wang X, Zhi Q and Wang L: Aberrant expression of miR-199a-3p and
its clinical significance in colorectal cancers. Med Oncol.
30:3782013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feber A, Xi L, Pennathur A, Gooding WE,
Bandla S, Wu M, Luketich JD, Godfrey TE and Litle VR: MicroRNA
prognostic signature for nodal metastases and survival in
esophageal adenocarcinoma. Ann Thorac Surg. 91:1523–1530. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang Y, Zhu Y, Wang X, Gong J, Hu C, Guo
B, Zhu B and Li Y: Temporal regulation of HIF-1 and NF-κB in
hypoxic hepatocarcinoma cells. Oncotarget. 6:9409–9419. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Joshi HP, Subramanian IV, Schnettler EK,
Ghosh G, Rupaimoole R, Evans C, Saluja M, Jing Y, Cristina I, Roy
S, et al: Dynamin 2 along with microRNA-199a reciprocally regulate
hypoxia-inducible factors and ovarian cancer metastasis. Proc Natl
Acad Sci USA. 111:5331–5336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mizuno S, Bogaard HJ, Gomez-Arroyo J,
Alhussaini A, Kraskauskas D, Cool CD and Voelkel NF:
MicroRNA-199a-5p is associated with hypoxia-inducible factor-1α
expression in lungs from patients with COPD. Chest. 142:663–672.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Raimondi L, Amodio N, Di Martino MT,
Altomare E, Leotta M, Caracciolo D, Gullà A, Neri A, Taverna S,
D'Aquila P, et al: Targeting of multiple myeloma-related
angiogenesis by miR-199a-5p mimics: In vitro and in vivo anti-tumor
activity. Oncotarget. 5:3039–3054. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rane S, He M, Sayed D, Yan L, Vatner D and
Abdellatif M: An antagonism between the AKT and beta-adrenergic
signaling pathways mediated through their reciprocal effects on
miR-199a-5p. Cell Signal. 22:1054–1062. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yeligar S, Tsukamoto H and Kalra VK:
Ethanol-induced expression of ET-1 and ET-BR in liver sinusoidal
endothelial cells and human endothelial cells involves
hypoxia-inducible factor-1alpha and microrNA-199. J Immunol.
183:5232–5243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Murakami Y, Toyoda H, Tanaka M, Kuroda M,
Harada Y, Matsuda F, Tajima A, Kosaka N, Ochiya T and Shimotohno K:
The progression of liver fibrosis is related with overexpression of
the miR-199 and 200 families. PLoS One. 6:e160812011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
He J, Xu Q, Jing Y, Agani F, Qian X,
Carpenter R, Li Q, Wang XR, Peiper SS, Lu Z, et al: Reactive oxygen
species regulate ERBB2 and ERBB3 expression via miR-199a/125b and
DNA methylation. EMBO Rep. 13:1116–1122. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kinose Y, Sawada K, Nakamura K, Sawada I,
Toda A, Nakatsuka E, Hashimoto K, Mabuchi S, Takahashi K, Kurachi
H, et al: The hypoxia-related microRNA miR-199a-3p displays tumor
suppressor functions in ovarian carcinoma. Oncotarget.
6:11342–11356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tauchi H, Matsuura S, Isomura M, Kinjo T,
Nakamura A, Sakamoto S, Kondo N, Endo S, Komatsu K and Nakamura Y:
Sequence analysis of an 800-kb genomic DNA region on chromosome
8q21 that contains the Nijmegen breakage syndrome gene, NBS1.
Genomics. 55:242–247. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ong CK, Ng CY, Leong C, Ng CP, Foo KT, Tan
PH and Huynh H: Genomic structure of human OKL38 gene and its
differential expression in kidney carcinogenesis. J Biol Chem.
279:743–754. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Defamie V, Cursio R, Le Brigand K,
Moreilhon C, Saint-Paul MC, Laurens M, Crenesse D, Cardinaud B,
Auberger P, Gugenheim J, et al: Gene expression profiling of human
liver transplants identifies an early transcriptional signature
associated with initial poor graft function. Am J Transplant.
8:1221–1236. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kao J, Salari K, Bocanegra M, Choi YL,
Girard L, Gandhi J, Kwei KA, Hernandez-Boussard T, Wang P, Gazdar
AF, et al: Molecular profiling of breast cancer cell lines defines
relevant tumor models and provides a resource for cancer gene
discovery. PLoS One. 4:e61462009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Raharijaona M, Le Pennec S, Poirier J,
Mirebeau-Prunier D, Rouxel C, Jacques C, Fontaine JF, Malthiery Y,
Houlgatte R and Savagner F: PGC-1-related coactivator modulates
mitochondrial-nuclear crosstalk through endogenous nitric oxide in
a cellular model of oncocytic thyroid tumours. PLoS One.
4:e79642009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
van Unnik JA, Coindre JM, Contesso C,
Lutter CE Albus, Schiodt T, Sylvester R, Thomas D, Bramwell V and
Mouridsen HT: Grading of soft tissue sarcomas: Experience of the
EORTC soft tissue and bone sarcoma group. Eur J Cancer 29A.
2089–2093. 1993. View Article : Google Scholar
|
|
51
|
Weiss SW and Goldblum JR: Malignant soft
tissue tumors of uncertain typeEnzinger and Weiss's Soft Tissue
Tumors. 5th. Mosby Elsevier; Philadelphia, PA: pp. 3–8. 2008
|