Introduction
Head and neck squamous cell carcinoma (HNSCC) is the
sixth most common cancer worldwide and has a poor prognosis
(1). The overall survival rate has
not changed over the past 30 years, with >400,000 patients
diagnosed and >200,000 mortalities annually (2). Despite numerous advances in treatment
modalities and techniques, including surgery, chemotherapy,
radiation therapy and combinations of these, only 50–60% of
patients survive for >5 years (3).
The static survival rate is considered to be the result of a high
incidence of treatment resistance, resulting in local/regional
recurrences and frequent distant metastases (4). Thus, to improve the survival of patients
with HNSCC, an increased understanding of the pathophysiology and
tumorigenesis of HNSCC is urgently required.
A previous study revealed that a small subpopulation
of tumor cells, cancer stem cells (CSCs), can initiate and maintain
tumors and metastasize (5). As
conventional therapies usually target differentiated cancer cells,
CSCs are considered to be responsible for treatment failure
(6). Thus, a considerable
understanding and identification of the promoters and regulators
that control expression of HNSCC CSCs may lead to methods to
increase the prognosis of HNSCC against treatment failure.
Transforming growth factor β (TGFβ) is a well-known
and potent regulator of the extracellular matrix (7). In human cancer, TGFβ shows biphasic
roles in tumor development and progression (8). TGFβ functions as a tumor suppressor by
cellular proliferation inhibition or by promoting differentiation
of normal cells (9). Mutations of
type II receptors in the TGFβ signaling pathway have been
demonstrated for head and neck cancer (10). However, once tumor cells become
resistant to the proliferation-inhibiting function of TGFβ, they
increase their production of TGFβ. As a result, the increase in
TGFβ leads to TGFβ-mediated angiogenesis stimulation, immune
suppression and cell motility stimulation (11). Consequently, an increased TGFβ level
results in increased invasiveness of tumor cells. Previously, it
was reported that TGFβ is able to increase the proportion of
cluster of differentiation (CD)133+ (a putative marker of CSCs)
cells in liver cancer cells, suggesting a positive role for TGFβ in
the ‘stemness’ of cancer cells (12).
However, whether TGFβ stimulates the key features of this cell
subpopulation in HNSCC has not been reported.
In the present study, the effect of an increased
TGFβ level on the stemness of HNSCC CSCs was investigated. The
association with the Wnt/β-catenin signaling pathway was also
examined to elucidate the downstream effect of TGFβ on stemness
regulation in HNSCC CSCs.
Materials and methods
Cell culture and reagents
The primary sphere K3 cell line was used. This cell
was isolated from a surgical specimen from a HNSCC patient, and the
CSC properties of the cell line have been validated using a number
of functional assays, testing self-renewal capability, stem cell
marker expression, chemoresistance and in vivo
tumorigenicity, as previously reported (5). Cells were cultured in serum-free
Dulbecco's modified Eagle's medium (DMEM)-Ham's F-12 (F12) medium
supplemented with human recombinant basic fibroblast growth factor
(bFGF; 10 ng/ml; R&D Systems, Minneapolis, MN, USA), N2
supplement (1 ml per 500 ml medium; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), and epidermal growth factor
(EGF; 10 ng/ml; R&D Systems). The primary β-catenin antibody
was purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA), and the secondary anti-mouse IgG antibody was obtained from
Jackson ImmunoResearch Laboratories (West Grove, PA, USA).
Tumorsphere formation assay
To assess self-renewal in vitro, the cells
were dissociated into a single-cell suspension, seeded in a 24-well
plate at a density of 500 cells per well, and cultured in
serum-free medium, with EGF and bFGF added every other day.
Untreated cells (cells not treated with EGF or bFGF) were used as a
control. Tumorspheres were allowed to grow for 7 days, and spheres
with a diameter >30 µm were counted.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blotting
RT-qPCR and western blotting experiments were
performed as previously described (5). RT-qPCR analysis of Wnt signaling target
genes (β-catenin, c-myc and cyclin D) was subsequently performed on
an iCycler IQ real-time detection system (Bio-Rad Laboratories,
Hercules, CA, USA), using IQ Supermix with SYBR-Green (Bio-Rad
Laboratories). The sequences of the human-specific primers used
were as follows: Oct4 forward, 5′-GCAATTTGCCAAGCTCCTGAA-3′ and
reverse, 5′-GCAGATGGTCGTTTGGCTGA-3′; Sox2 forward,
5′-CCTCCGGGACATGATCAG-3′ and reverse, 5′-TTCTCCCCCCTCCAGTTC-3′;
Twist forward, 5′-CAGTCTTACGAGGAGCTGCA-3′ and reverse,
5′-TCTTGCTCAGCTTGTCCGAG-3′; Snail forward,
5′-CACCTCCAGACCCACTCAGAT-3′ and reverse,
5′-CCTGAGTGGGGTGGGAGCTTC-3′; Slug forward,
5′-CTCCATGCCTGTCATACCAC-3′ and reverse, 5′-GGAAAGAGGAGAGAGGCCAT-3′;
β-catenin forward, 5′-GGCAGTGCGTTTAGCTGGTG-3′ and reverse,
5′-AGCTTGGGGTCCACCACTAG-3′; c-myc forward,
5′-GTATGTGGACGGCTTCTCGC-3′ and reverse, 5′-GCTGCTGTCGTTGAGAGGGT-3′;
and cyclin D forward, 5′-GGAGCTGCTGCAAATGGAGC-3′ and reverse,
5′-CAGAGGGCAACGAAGGTCTG-3′.
Aldehyde dehydrogenase (ALDH) activity
assay
ALDH activity was analyzed using an Aldefluor assay
kit (Stem Cell Technologies, Vancouver, BC, Canada). Briefly, cells
were suspended in Aldefluor assay buffer containing the ALDH
substrate (1 µmol/l per 1×106 cells). A sample of cells was stained
with the specific ALDH inhibitor diethylaminobenzaldehyde (DEAB) as
a negative control. Fluorescent ALDH+ cells were detected in the
green fluorescence channel, and samples treated with the specific
ALDH inhibitor DEAB were used as the control to set the gates
defining the ALDH+ region. Flow cytometric sorting was conducted
using a FACSAria flow cytometer (Becton Dickinson, San Jose, CA,
USA).
Chemosensitivity assay
Cells were dissociated into a single-cell suspension
and then plated in a 24-well plate at a density of 1×105 cells per
well under serum-free culture conditions. The cells were treated
with cisplatin at the indicated concentrations and then cultured at
37°C in a humidified 5% CO2 atmosphere. Subsequently, 24 h later,
50 µl of MTT solution (5 mg/ml in PBS) was added to each well, and
the plate was incubated at room temperature for 2 h. Absorbance was
measured on a SpectraMax 190 device (Molecular Devices, LLC,
Sunnyvale, CA, USA) at a wavelength of 570 nm.
Optimal Tcf-binding site
(TOP)/far-from-optimal Tcf-binding site (FOP) luciferase assay
Cells were seeded in 24-well plates at a density of
1×105 cells per well. Cells were transiently transfected using
Polyexpress® (Excellgen, Rockville, MD, USA). For the
luciferase assay, cells were transfected with 0.5 µg of the
lymphoid enhancer factor-reporter pTOPFLASH/pFOPFLASH along with 50
ng of pRL-SV40. TGFβ1 treatment was initiated at 24 h
post-transfection and performed for 48 h. Luciferase activity was
monitored using the Dual-Glo luciferase assay system (Promega
Corporation, Madison, WI, USA). The transfection efficiency was
normalized to the cotransfected Renilla luciferase activity
according to the manufacturer's protocol.
Statistical analysis
Experimental data were statistically assessed using
either 2-tailed Student's t-test or analysis of variance with
Scheffe's post-hoc analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
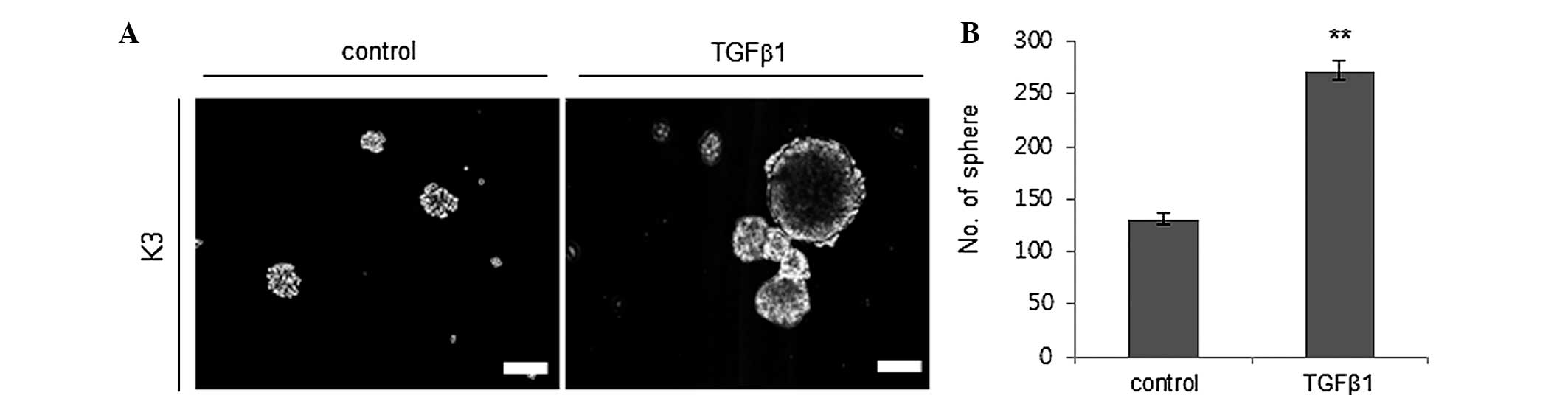
TGFβ1 increases the self-renewal
capacity of cancer stem cells
The capacity for self-renewal is one of the putative
traits of CSCs (5). Thus, it was
evaluated whether TGFβ1 treatment enhances the self-renewal
capacity of HNSCC CSCs by performing a sphere-forming assay. TGFβ1
(2 ng/ml) treatment significantly increased the sphere formation
ability of HNSCC CSCs with a diameter of >30 µm compared to the
control treatment group (Fig. 1).
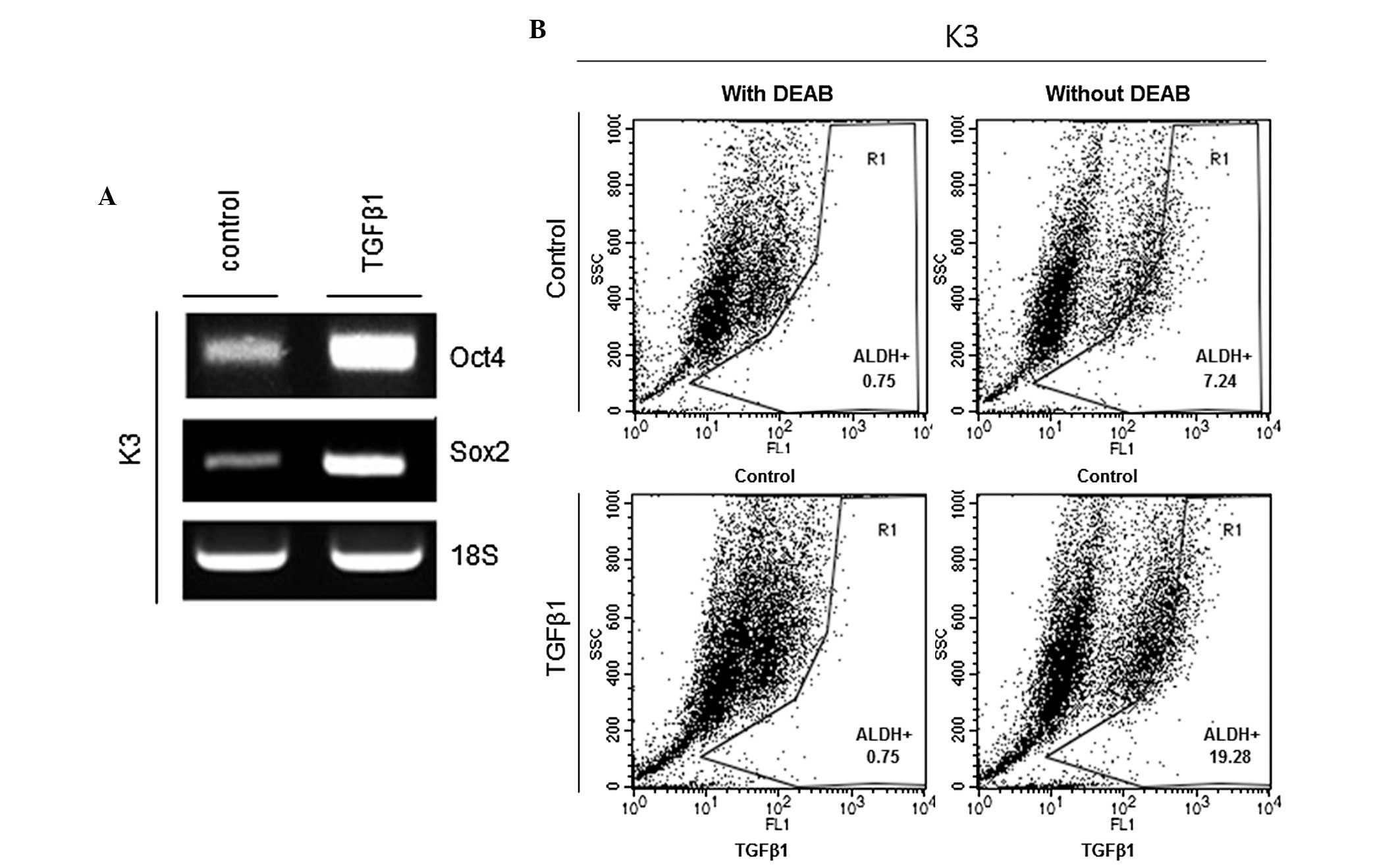
TGFβ1 increases the levels of stemness
markers of CSCs
Oct4 and Sox2 are critical regulators of
pluripotency in the mammalian embryo, and deregulated expression of
these genes can be found in HNSCC CSCs (13). Thus, whether TGFβ1 treatment increases
the transcriptional expression of Oct4 and Sox2 was examined. The
results showed that mRNA expression levels of Oct4 and Sox2 were
significantly increased in the TGFβ1 treatment group compared to
the control treatment group (Fig.
2A). ALDH is considered to be a marker of HNSCC CSCs (14). Therefore, ALDH activity was examined
in TGFβ1- and control-treated HNSCC CSCs. Fig. 2B shows that the proportion of ALDH+
cells was significantly increased in the TGFβ1 treatment group
compared with the control treatment group (7.24 vs. 19.28%).
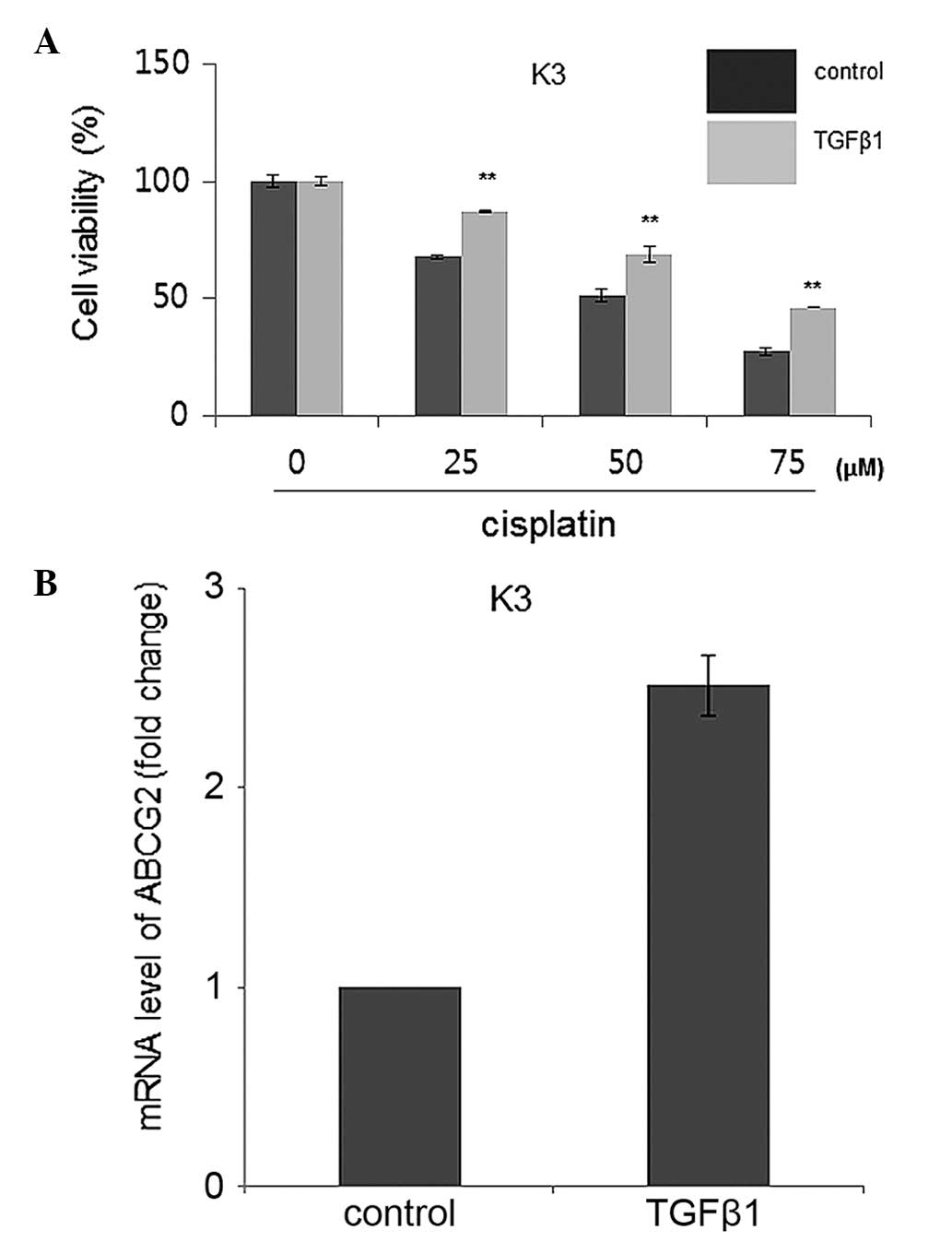
TGFβ1 increases chemoresistance to
cisplatin for HNSCC CSCs by overexpression of ATP-binding cassette
sub-family G member 2 (ABCG2)
HNSCC CSCs possess a chemoresistance ability against
anticancer drugs, and the ABCG2 transporter is responsible for this
resistance phenotype (5). Therefore,
chemoresistance levels were compared using MTT assay subsequent to
cisplatin administration at various concentrations with the TGFβ1
and control treatment groups. The results showed that TGFβ1-treated
HNSCC CSCs had significantly increased cisplatin resistance
compared with control-treated HNSCC CSCs (Fig. 3A). Furthermore, an increased ABCG2
expression level was identified in TGFβ1-treated HNSCC CSCs
compared to control-treated HNSCC CSCs, indicating that TGFβ1 may
enhance the chemoresistance of HNSCC CSCs through increased
expression of ABCG2 (Fig. 3B).
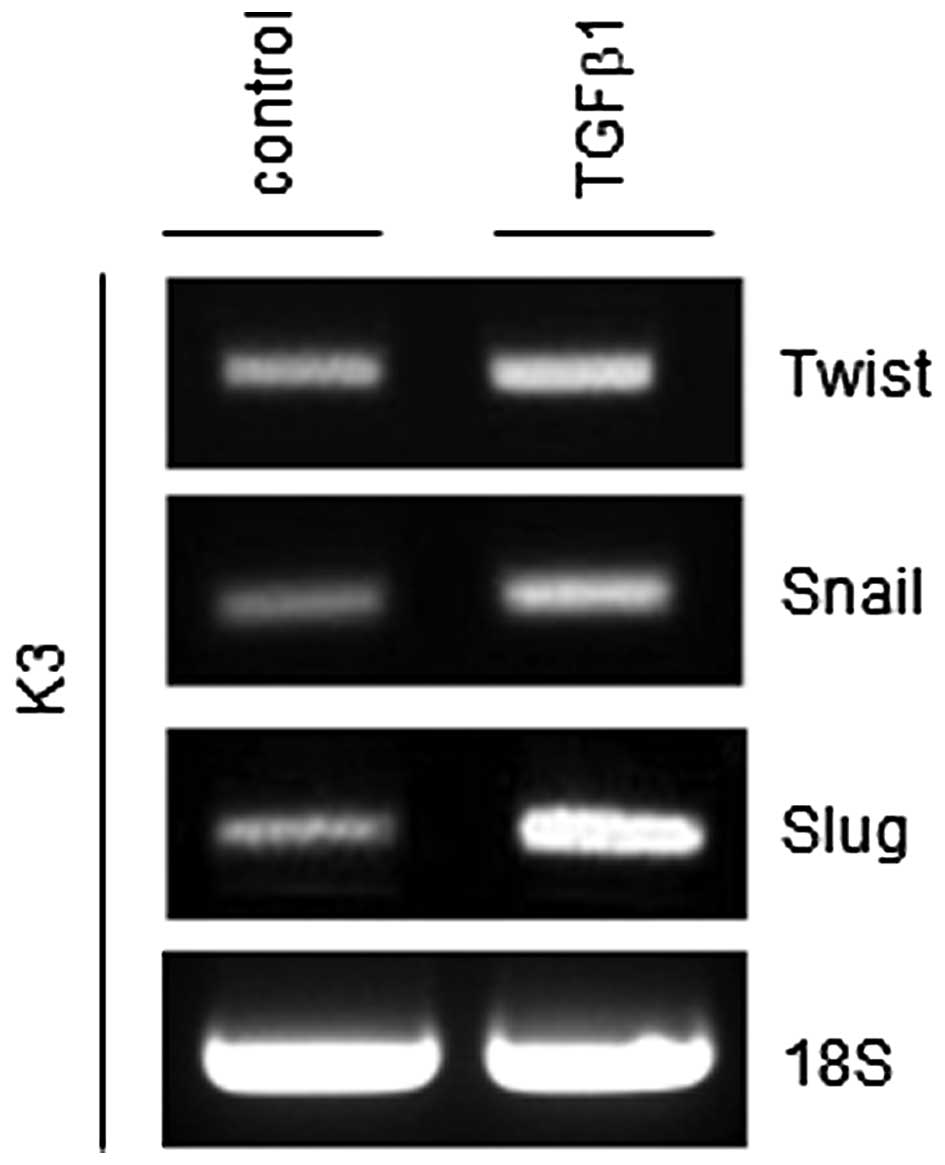
TGFβ1 increases the expression of
epithelial-mesenchymal transition (EMT) regulators
EMT is a key process in tumor invasion and
metastasis (15). Previously, it was
reported that the EMT process can generate cancer cells with a stem
cell phenotype (15). Thus, whether
TGFβ1 can increase central regulators of the EMT process, such as
Twist, Snail and Slug, was investigated. As shown in Fig. 4, mRNA expression levels of all three
EMT regulators were increased in TGFβ1-treated HNSCC CSCs compared
with control-treated HNSCC CSCs.
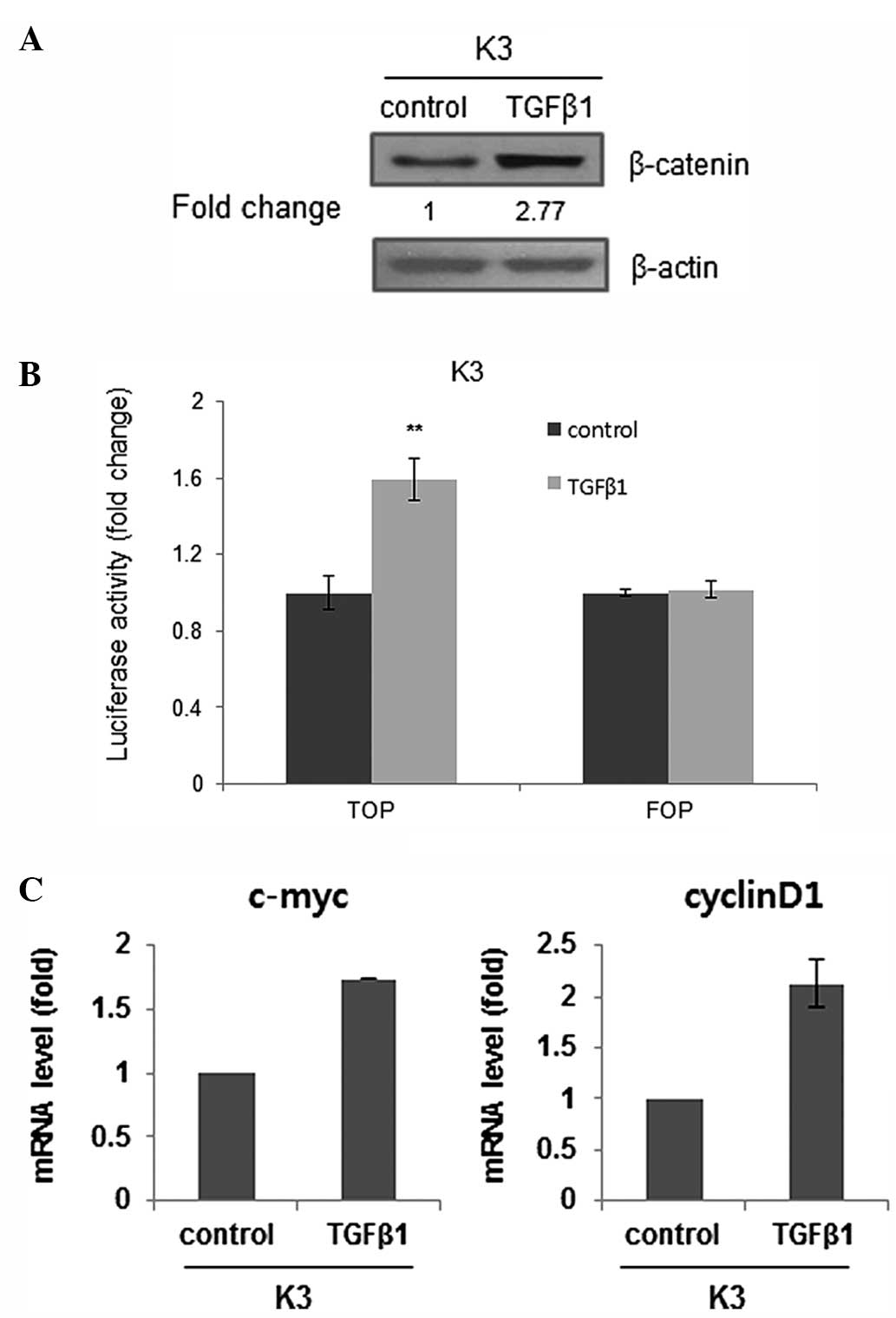
TGFβ1 activates canonical
Wnt/β-catenin signaling
It was previously suggested that Wnt/β-catenin
contributes to the stemness of HNSCC cells (16). Thus, whether TGFβ1 acts as an upstream
stimulator of Wnt/β-catenin signaling was investigated.
Administration of TGFβ1 increased the expression of β-catenin,
which is an effector of the Wnt pathway (Fig. 5A). Also, TGFβ1 treatment increased the
activity of a Wnt/β-catenin-dependent reporter as a functionally
more relevant indicator (Fig. 5B).
Furthermore, the transcript levels of Wnt/β-catenin signaling
target genes, such as the c-myc and cyclin D1 genes, were increased
in TGFβ1-treated HNSCC CSCs (Fig.
5C). Overall, these data suggest that TGFβ1-induced stemness
may be associated with Wnt pathway upregulation in HNSCC CSCs.
Discussion
Numerous previous studies have indicated that a rare
population of cells within the entire tumor bulk, termed CSCs or
cancer-initiating cells, possess capabilities for tumor initiation
and progression, and in certain cases, metastasis, which are not
found in the majority of cells (5,14,17). In clinical settings, these cells
exhibit an intrinsic resistance to popular chemotherapeutic agents,
preventing complete eradication of the tumor cells following
treatment. Therefore, an understanding of the molecular mechanism
of CSCs as a treatment target is critical for the design and
development of effective anticancer therapies against tumor
relapse.
Several signal transmission pathways regulating CSC
pathobiology have been suggested, including the Wnt, Notch and Hh
pathways (18). Previously, members
of the TGFβ cytokine family, such as TGFβs, bone morphogenetic
proteins (BMPs), Nodal and activins, were also shown to be involved
in the control of the CSC phenotype, particularly in glioblastoma
multiforme (GM) (19,20). Notably, BMP and TGFβ have conflicting
effects on GM CSC proliferation, although they are included in the
same family (TGFβ) (19,20). BMP has been shown to inhibit tumor
growth and to induce differentiation of GM CSCs (19). By contrast, TGFβ promotes self-renewal
and prevents differentiation, enhancing the oncogenic ability of GM
CSCs (21).
TGFβ1 expression is observed in ~80% of cases of
human HNSCC and correlates with more advanced disease and reduced
survival (11). In addition, TGFβ1
overexpression promotes tumorigenesis in a paracrine manner,
leading to increased inflammation and angiogenesis (11). However, different TGFβ signaling
disruptions also enhance epithelial carcinogenesis through various
mechanisms and are common in HNSCC, suggesting biphasic roles for
this signaling pathway in tumorigenesis of HNSCC (22). Also, there are extremely few previous
studies regarding the effects of TGFβ ligands on HNSCC CSCs. Thus,
the present study aimed to clarify the role of TGFβ1 in HNSCC CSC
biology.
In the present study, it was shown that TGFβ1
treatment enhances self-renewal and stemness-associated genes (Oct4
and Sox2) expression in HNSCC CSCs. In addition, this treatment was
shown to result in enrichment of the ALDH+ cell population,
expressing a putative marker of HNSCC CSCs. Furthermore, TGFβ1
treatment results in an increase of stem cell-like traits, such as
resistance to conventional chemotherapy (cisplatin) for treatment
of HNSCC patients. Collectively, the present data suggest that
TGFβ1 may be involved in the regulation of cells with stem
cell-like traits in HNSCC.
Mechanistically, it has been shown that the
regulation of GM CSC self-renewal by TGFβ is mediated by leukemia
inhibitory factor (LIF), a member of the interleukin 6 (IL6) family
of cytokines, and by signaling through a heterodimeric receptor
complex composed of the glycoprotein 130 and the LIF receptor
inducing the Janus kinase-signal transducers and activators of
transcription pathway (21). In
addition, IL6 may also be a mediator of the induction of GM CSC
self-renewal by TGFβ (23). Notably,
the present study showed that TGFβ administration increased the
expression of β-catenin, a major effector of the canonical Wnt
signal, although a detailed mechanism is not presented here. Serra
et al observed that Wnt5a, a Wnt ligand, was directly
regulated by TGFβ in primary mammary gland cells, and they
identified Smad binding sites in the Wnt5a promoter (24). Thus, crosstalk between TGFβ signaling
and Wnt signaling may be significant in HNSCC CSC biology.
Additional studies are required to show how TGFβ signaling
interacts with Wnt signaling.
Based on these data, the present findings
demonstrate that TGFβ1 may have an important role in HNSCC CSC
genesis and that its inhibitors may be valuable for eradicating
HNSCC CSCs.
Acknowledgements
This study was supported by Konkuk University
Hospital (grant no., 201403).
References
|
1
|
Jemal A, Murray T, Samuels A, Ghafoor A,
Ward E and Thun MJ: Cancer statistics, 2003. CA Cancer J Clin.
53:5–26. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Forastiere A, Koch W, Trotti A and
Sidransky D: Head and neck cancer. N Engl J Med. 345:1890–1900.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamano Y, Uzawa K, Saito K, Nakashima D,
Kasamatsu A, Koike H, Kouzu Y, Shinozuka K, Nakatani K, Negoro K,
et al: Identification of cisplatin-resistance related genes in head
and neck squamous cell carcinoma. Int J Cancer. 126:437–449. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haddad RI and Shin DM: Recent advances in
head and neck cancer. N Engl J Med. 359:1143–1154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lim YC, Oh SY, Cha YY, Kim SH, Jin X and
Kim H: Cancer stem cell traits in squamospheres derived from
primary head and neck squamous cell carcinomas. Oral Oncol.
47:83–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mannelli G and Gallo O: Cancer stem cells
hypothesis and stem cells in head and neck cancers. Cancer Treat
Rev. 38:515–539. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu L, Jiao J, Wang Y, Wu J, Huang D, Teng
W and Chen L: TGF-beta1 gene polymorphism in association with
diabetic retinopathy susceptibility: A systematic review and
meta-analysis. PLoS One. 9:e941602014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
White RA, Malkoski SP and Wang XJ: TGFβ
signaling in head and neck squamous cell carcinoma. Oncogene.
29:5437–5446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Siegel PM and Massagué J: Cytostatic and
apoptotic actions of TGF-beta in homeostasis and cancer. Nat Rev
Cancer. 3:807–821. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu SL, Herrington H, Reh D, Weber S,
Bornstein S, Wang D, Li AG, Tang CF, Siddiqui Y, Nord J, et al:
Loss of transforming growth factor-beta type II receptor promotes
metastatic head-and-neck squamous cell carcinoma. Genes Dev.
20:1331–1342. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu SL, Reh D, Li AG, Woods J, Corless CL,
Kulesz-Martin M and Wang XJ: Overexpression of transforming growth
factor beta1 in head and neck epithelia results in inflammation,
angiogenesis, and epithelial hyperproliferation. Cancer Res.
64:4405–4410. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
You H, Ding W and Rountree CB: Epigenetic
regulation of cancer stem cell marker CD133 by transforming growth
factor-beta. Hepatology. 51:1635–1644. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kashyap V, Rezende NC, Scotland KB,
Shaffer SM, Persson JL, Gudas LJ and Mongan NP: Regulation of stem
cell pluripotency and differentiation involves a mutual regulatory
circuit of the NANOG, OCT4, and SOX2 pluripotency transcription
factors with polycomb repressive complexes and stem cell microRNAs.
Stem Cells Dev. 18:1093–1108. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zou B, Sun S, Qi X and Ji P: Aldehyde
dehydrogenase activity is a cancer stem cell marker of tongue
squamous cell carcinoma. Mol Med Rep. 5:1116–1120. 2012.PubMed/NCBI
|
|
15
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SH, Koo BS, Kim JM, Huang S, Rho YS,
Bae WJ, Kang HJ, Kim YS, Moon JH and Lim YC: Wnt/β-catenin
signalling maintains self-renewal and tumourigenicity of head and
neck squamous cell carcinoma stem-like cells by activating Oct4. J
Pathol. 234:99–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dalerba P and Clarke MF: Cancer stem cells
and tumor metastasis: First steps into uncharted territory. Cell
Stem Cell. 1:241–242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Campelo MR Garcia, Curbera G Alonso,
Gallego G Aparicio, Pulido E Grande and Antón Aparicio LM: Stem
cell and lung cancer development: Blaming the Wnt, Hh and Notch
signalling pathway. Clin Transl Oncol. 13:77–83. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Piccirillo SG, Reynolds BA, Zanetti N,
Lamorte G, Binda E, Broggi G, Brem H, Olivi A, Dimeco F and Vescovi
AL: Bone morphogenetic proteins inhibit the tumorigenic potential
of human brain tumour-initiating cells. Nature. 444:761–765. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee J, Son MJ, Woolard K, Donin NM, Li A,
Cheng CH, Kotliarova S, Kotliarov Y, Walling J, Ahn S, et al:
Epigenetic-mediated dysfunction of the bone morphogenetic protein
pathway inhibits differentiation of glioblastoma-initiating cells.
Cancer Cell. 13:69–80. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Seoane J: TGFbeta and cancer initiating
cells. Cell Cycle. 8:3787–3788. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Levy L and Hill CS: Alterations in
components of the TGF-beta superfamily signaling pathways in human
cancer. Cytokine Growth Factor Rev. 17:41–58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang H, Lathia JD, Wu Q, Wang J, Li Z,
Heddleston JM, Eyler CE, Elderbroom J, Gallagher J, Schuschu J, et
al: Targeting interleukin 6 signaling suppresses glioma stem cell
survival and tumor growth. Stem Cells. 27:2393–2404. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Serra R, Easter SL, Jiang W and Baxley SE:
Wnt5a as an effector of TGFβ in mammary development and cancer. J
Mammary Gland Biol Neoplasia. 16:157–167. 2011. View Article : Google Scholar : PubMed/NCBI
|