Introduction
Colorectal cancer is the third leading cause of
mortality in the Western world (1)
and has emerged as a common malignancy in the Asian population as a
result of changes in diet and physical activity levels (2). Dietary habits have been related to the
risk of colorectal cancer (3,4). Surgery and chemotherapy are the primary
treatments for colorectal cancer. Radiotherapy is a typical
adjuvant treatment after surgery or chemotherapy for high-stage
colorectal cancers (5,6). However, colorectal carcinomas display a
wide range of radiosensitivity (7,8).
Therefore, new approaches are necessary to enhance the efficacy of
radiation treatments for colorectal cancers.
Previous epidemiological studies have shown that the
daily inclusion of fruit and vegetables in the diet decreases the
risk of colon cancer (9). In
addition, it has been reported that flavonoids, which are abundant
in numerous plants, protect against a number of tumorigenic
processes, including oxidative stress, inflammation, angiogenesis
and cell invasion (10–12). Furthermore, flavonoids induce cell
cycle arrest, apoptosis and radiosensitivity in cancer cells in
vitro (13–15). The flavonoid fisetin
(3,7,3′,4′-tetrahydroxyflavone) is a polyphenol found in numerous
plants. A number of previous reports have shown that fisetin
activates p53 activity, and represses the cyclooxygenase-2 and
Wnt/epidermal growth factor receptor/nuclear factor-B signaling
pathways in human cancer cells to promote apoptosis (14,16–18). In
addition, fisetin inhibits the spindle checkpoint response that
arrests cells in the radiosensitive G2/M phase (19,20).
However, the in vivo effects of fisetin remain unclear. As
fisetin is a natural and edible product with acceptable biosafety,
the clinical potential of this compound is of particular interest
and warrants further investigation.
Securin, which was originally isolated from rat
pituitary tumor cells, is alternatively called the pituitary tumor
transforming gene (21). Securin is a
multi-functional protein that serves a number of biological roles,
such as the regulation of cellular transformation, sister chromatid
separation (22,23), gene transcription (24) and DNA damage repair (25,26).
Notably, securin interacts with p53 and perturbs p53-mediated
transcription and apoptosis in tumor cells (22). Thus, securin is regarded as an
oncoprotein. The depletion of securin has been reported to
sensitize human colorectal cancer cells to various types of
treatment, including fisetin, butein and ionizing radiation
(27–29). However, whether these effects can be
repeated in vivo is unknown.
In the present study, tumor-bearing mice were used
to examine the effect of fisetin alone and in combination with
radiation on the growth of colorectal tumors in vivo. In
addition, tumors with defective securin expression were assessed to
investigate if securin depletion would enhance sensitivity to these
treatments.
Materials and methods
Cell culture
Murine CT-26 colon cancer cells, and human
HCT116WT, HCT116p53-/−,
HCT116securin-/− and p53-R273H mutant HT-29 colorectal
cancer cell lines (30) were cultured
in RPMI-1640 medium (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (Thermo Fisher
Scientific Inc.), 100 U/ml penicillin, 100 µg/ml streptomycin and 1
mM sodium pyruvate. Cultures were maintained at 37°C in a 95%
humidified incubator (Thermo Fisher Scientific, Inc.) with 5%
CO2 and passaged at 1:3 every 2 days.
Mouse xenograft models
A total of 62 male BALB/c nude mice (weight, 20 g;
age, 6 weeks) were purchased from the National Laboratory Animal
Center (NLAC, Nankang, Taipei, Taiwan). The mice were maintained at
22–24°C and 70% humidity under a 12-h light/dark cycle. Food and
water were available ad libitum. Five mice were kept in each
77.4×77.4-cm cage. All animal protocols were approved by the
Institutional Animal Care and Use Committee (IACUC) of National
Yang-Ming University (Taipei, Taiwan; approval no. 1001270). Prior
to tumor xenografting, mice were anesthetized with ketamine (50
mg/kg; IMALGENE®; Merial Laboratoire de Toulouse, Lyon,
France) and xylazine (15 mg/kg; Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany). Subsequently, CT-26 (1×106) or
HCT116 (2×106) cells were subcutaneously injected into
the hind legs of the mice (n=5 for injection of each cell line).
Prior to further treatment, mice were maintained until tumors
reached 100 mm3. Tumor volume was measured using a
caliper and calculated as the following: (Length ×
width2) / 2. Tumor volume was measured every 2–3 days to
draw tumor growth curves followed by measurement of body
weight.
Reagents and radiation treatments
Fisetin was purchased from Sigma-Aldrich
(Sigma-Aldrich) and intratumorally injected at a dose of 5 mg/kg on
days 0 and 7. Tumors were irradiated with 2 Gy/day for 5 days using
an X-ray machine (RS 2000 Biological Research X-ray Irradiator; Rad
Source Technologies, Inc., Suwanee, GA, USA) operating at 160 kVp
and 25 mA. The dose rate at a source to subject distance of 38 cm
was 1.83 Gy/min. For combined fisetin and radiation treatment, mice
were treated with 5 mg/kg fisetin followed by 2 Gy irradiation on
days 0 and 7. For the control group, the mice were injected with
DMSO at the same volume of dissolved fisetin on days 0 and 7. Five
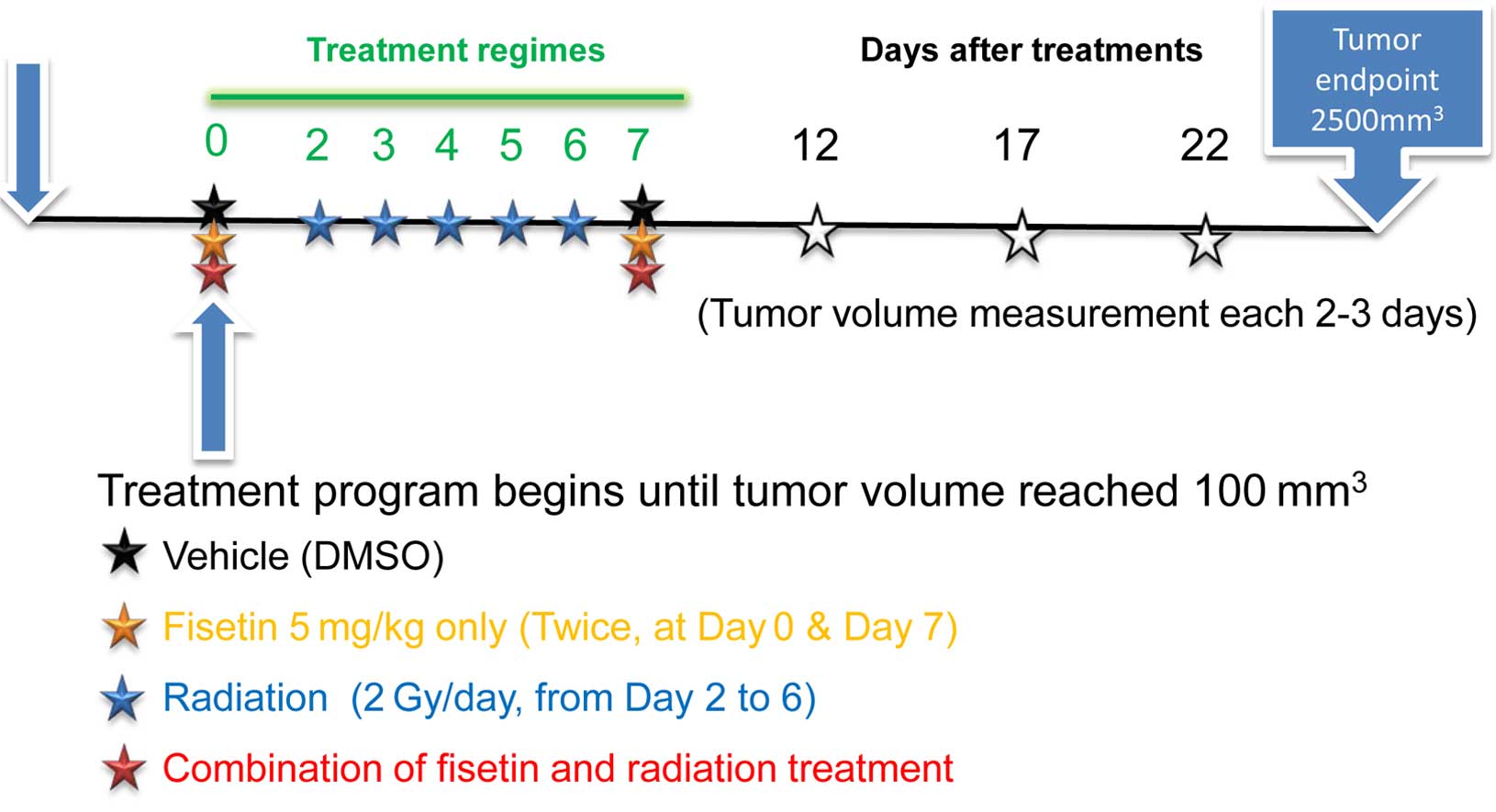
mice were used in each group. These treatments are illustrated on a
timeline in Fig. 1. Excluding the
tumor site, all areas of the mice were masked using a lead
radiation protector.
Western blotting and antibodies
Western blot analysis was performed as described
previously (29). The following
primary antibodies were used: Anti-p53 (Cat. no. GEX70214; 1:2,000;
GeneTex, Inc., Irvine, CA, USA), anti-securin (Cat. no. ab3305;
1:1,000; Abcam, Cambridge, UK) and anti-GAPDH (Cat. no. PB197650;
1:5,000; Thermo Fisher Scientific Inc.). The secondary antibodies
included a goat anti-mouse antibody (Cat. no. AP124P; 1:10,000; EMD
Millipore, Billerica, MA, USA) for detecting the anti-securin and
anti-GAPDH primary antibodies, and a goat anti-rabbit antibody
(Cat. no. AP132P; 1:10,000; EMD Millipore) for detecting the
anti-p53 primary antibody. Band intensities were measured by
densitometry using ImageJ 1.x software (National Institutes of
Health, Bethesda, MD, USA) (31).
Cell proliferation measurement
Cells (1×105) were seeded into 6-cm
culture dishes and collected for hemocytometric calculation every
day for 7 days. For each time point, the mean number of cells was
calculated from three independent cell cultures. The results were
plotted as cell proliferation curves.
Statistical analysis
Results are presented as the mean ± standard
deviation. A Student's t-test was performed to determine if
differences between groups were statistically significant. For
survival analysis, the Kaplan-Meier estimator was used and the
results were analyzed using the log-rank test. Statistical analyses
were performed using GraphPad Prism 3.0 software (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Fisetin inhibits tumor growth in a
mouse CT-26 xenograft model
To examine the effect of fisetin on colorectal
cancer growth in vivo, a mouse tumor xenograft model
generated by injection of CT-26 colon cancer cells was treated with
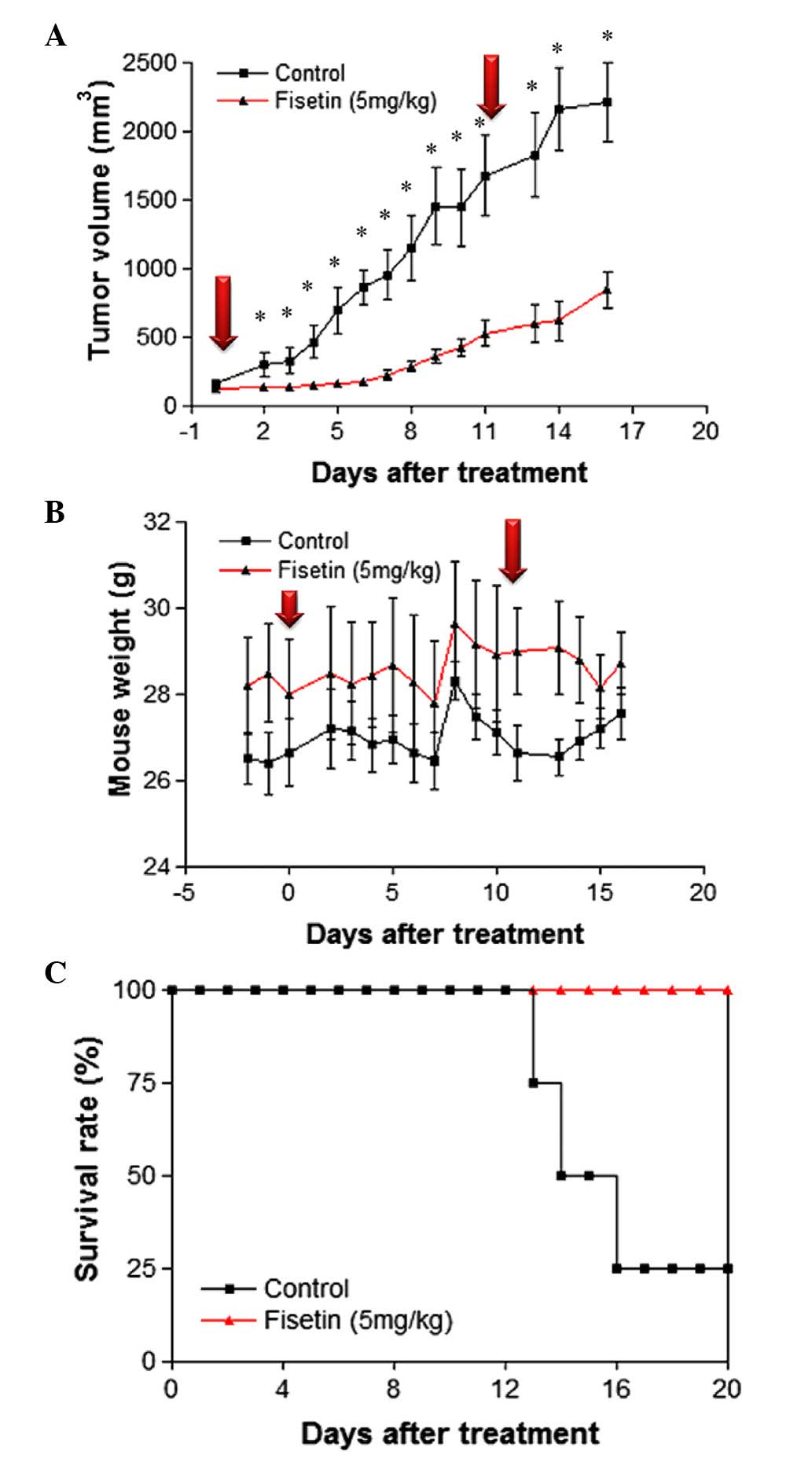
fisetin. Compared with the untreated control group, a single
intratumoral injection of 5 mg/kg fisetin significantly reduced
tumor volume for the following 10 days (Fig. 2A). As tumors appeared to re-grow 6
days after the first injection, a second dose of fisetin was
administered on day 11 to examine if tumor growth could be
inhibited. The results demonstrated that tumor growth was
significantly suppressed for 3 days following the second
administration of fisetin, as compared with the control group
(Fig. 2A), although tumor growth was
not completely inhibited. Fluctuations in body weight between the
control and fisetin-treated group were similar (Fig. 2B), indicating that the concentration
of fisetin used in this study was not cytotoxic. In addition, the
survival rate of fisetin-treated tumor-bearing mice was increased
compared with the untreated control group (Fig. 2C). These results demonstrate that
administration of fisetin suppresses in vivo tumor growth in
a mouse CT-26 xenograft model.
Combining fisetin and radiation for
the treatment of mammalian colorectal cancers
Treatment with a combination of fisetin and
radiation treatment was investigated in mouse CT-26 and
HCT116WT xenograft tumor models. The timeline of fisetin
and radiation treatment is illustrated in Fig. 1; 2-Gy X-rays were administered to the
tumor site five times between the first and second fisetin
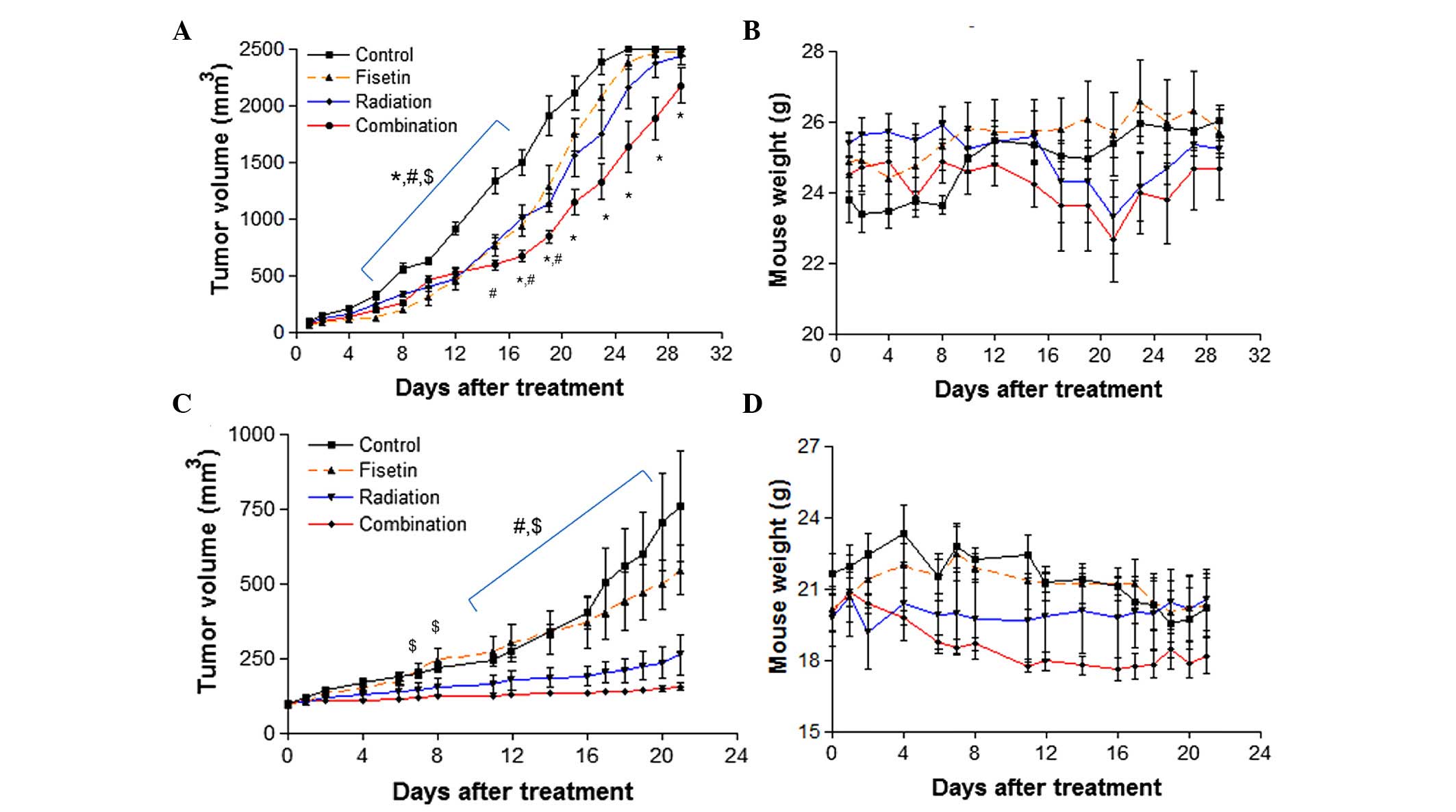
treatments to mimic a clinical regime. The results showed that
CT-26 tumor growth was suppressed by fisetin and radiation alone;
however, this effect was enhanced by combining the two treatments
at 16 to 29 days following treatment (Fig. 3A). Body weight was not significantly
different between any of the groups, suggesting that combined
fisetin/radiation treatment did not cause systemic toxicity
(Fig. 3B). In addition, HCT116 tumor
growth was completely and significantly inhibited by combined
fisetin/radiation treatment (P<0.05 vs. the control group;
Fig. 3C), without significant loss of
body weight (Fig. 3D). These results
suggest that combined fisetin and radiation treatment is highly
effective in suppressing colorectal cancer.
Fisetin induces p53 and suppresses
securin protein expression in human colorectal cancer cells
p53 is a tumor suppressor protein that is regulated
by a number of different proteins. A previous study demonstrated
that the securin oncoprotein binds to p53 and modulates its
activity (32). Although securin
protein levels have been reported to be elevated by radiation
independently of p53 activity (27),
it is unknown whether fisetin influences the expression of p53 or
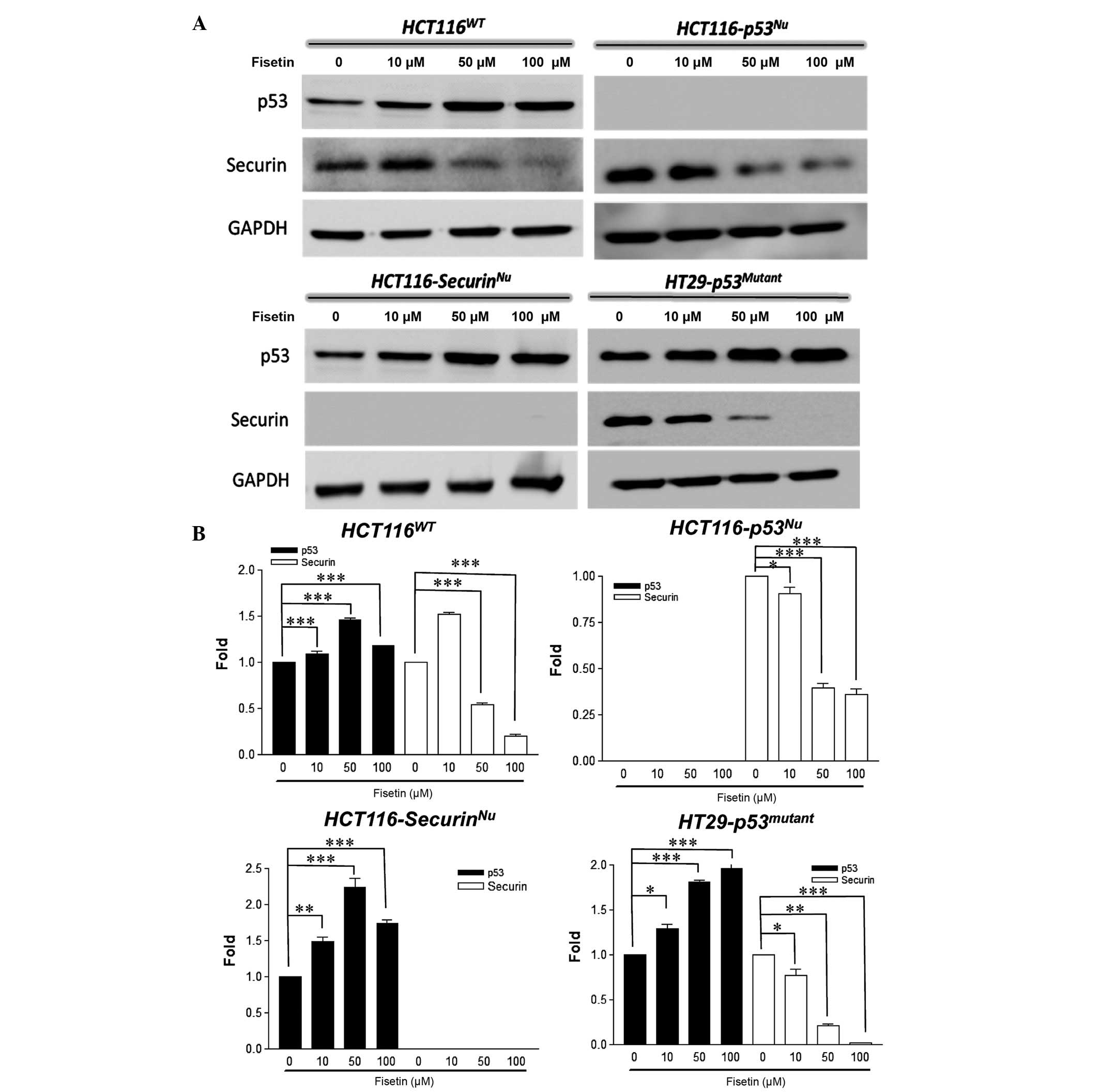
securin. In the present study, an experiment with a series of
fisetin doses was performed to investigate whether fisetin
influences the expression of p53 and securin in different human
colorectal cancer cell lines. The results identified that in
HCT116WT cells, p53 protein levels were increased and
securin protein levels were decreased following fisetin treatment
(Fig. 4A). Interestingly, fisetin
increased the expression of p53 in HCT116securin-/−
cells and decreased the expression of securin in
HCT116p53-/− cells (Fig.
4A). In p53 mutant HT-29 cells, fisetin downregulated the
expression of securin protein (Fig.
4A). Quantification of the protein bands on the Western Blot
analysis revealed the same results (Fig.
4B). These results indicate that fisetin increases the
expression of p53 and decreases the expression of securin, which
suppresses tumor growth. Furthermore, fisetin-mediated expression
of p53 and securin was independent of the expressive status of
p53/securin (null or wild-type).
Effect of fisetin and radiation
combination treatment on securin-null colorectal cell-formed
xenograft tumors
As fisetin could induce the expression of the p53
protein in the absence of securin, the ability of fisetin and
radiation treatment to suppress tumor growth in vivo was
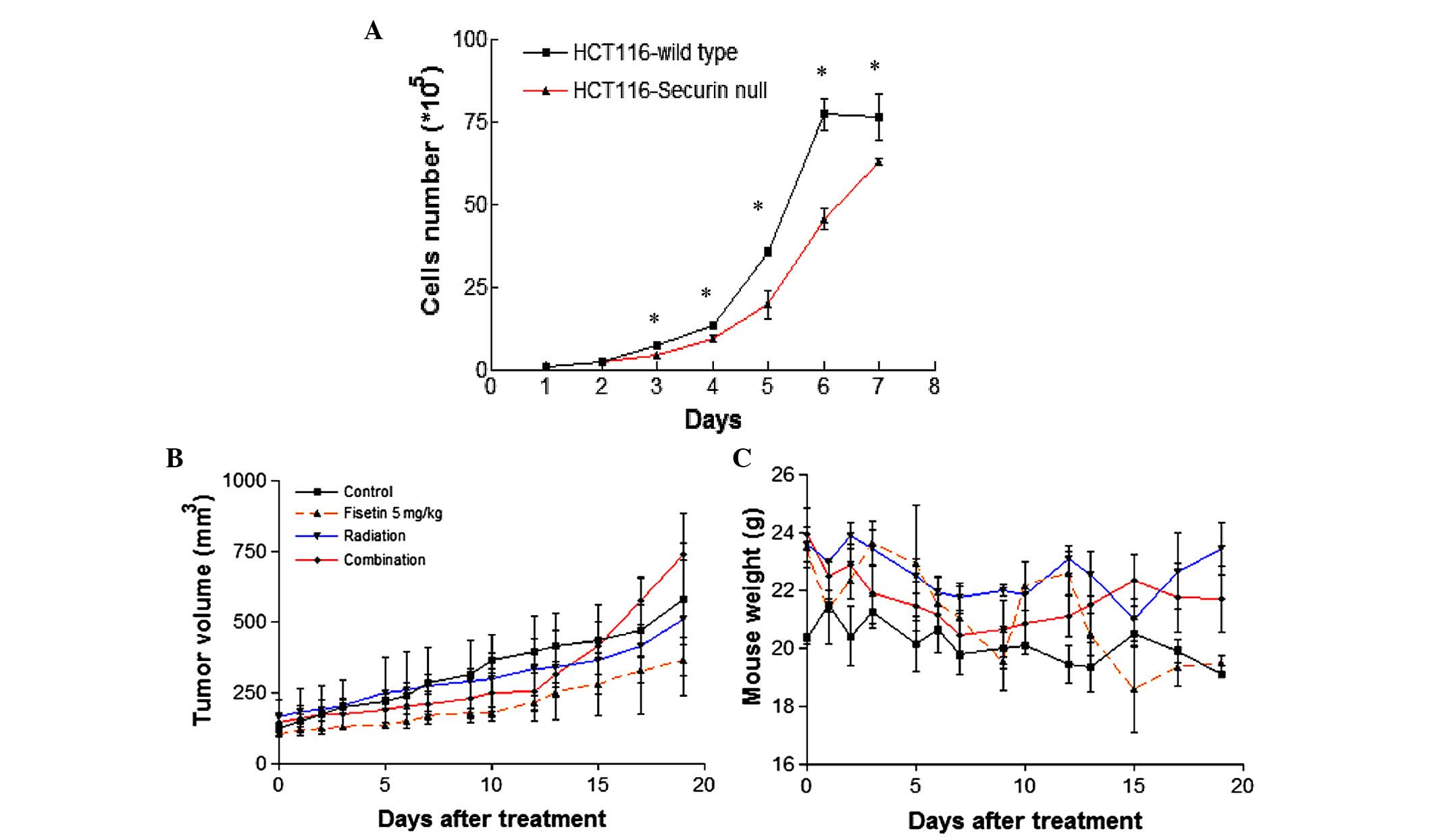
investigated. Firstly, the growth curves of HCT116WT and
HCT116securin-/− cells were compared, which showed that
the proliferation of HCT116securin-/− cells was
significantly slower than that of HCT116WT cells after 3
days of proliferation (Fig. 5A).
Tumor growth was moderately suppressed by fisetin treatment in
HCT116securin-/− cells; however, no significant
advantage was observed following fisetin and radiation combination
treatment (Fig. 5B). The body weight
of the mice following fisetin, radiation and fisetin/radiation
showed no significant difference compared with the control group
(Fig. 5C). Therefore, depletion of
securin does not enhance the effects of fisetin and radiation
combination treatment on colorectal tumors in vivo. However,
a larger sample size is necessary to validate this result.
Discussion
Previous studies have shown that fisetin possesses a
wide range of activities to suppress the growth of human cancer
cells, including breast, prostate, bladder, lung, melanoma and
colorectal cancer cells (16,33–39). Among
these types of cancer, colorectal cancer is particularly
interesting with regards to fisetin, since fisetin is a nutrient
supplement that can be administrated orally (40). It is thought that fisetin is absorbed
through the digestive tract without the need for intravascular
injection (41). Although in
vitro studies have demonstrated the efficacy of fisetin in the
treatment of colorectal cancer (15,16), few
in vivo studies have been reported. Since preclinical
studies are essential for potent therapeutic agents to be accepted
for clinical trials, the experimental animal data in the present
study is important for evaluating the application of fisetin to
colorectal cancer therapy. In addition, the current study
investigated whether fisetin treatment combined with ionizing
radiation exerted synergistic effects. A previous study reported
that a combination of cisplatin and fisetin exerted anticancer
activity in embryonic carcinoma cells in vitro and in
vivo (42). Although radiotherapy
is not typically used in the treatment of colorectal cancer, it is
frequently applied to adjuvant chemotherapy in rectal cancer
(43,44). The combination of fisetin and
radiotherapy is an interesting alternative adjuvant therapy, since
it would likely avoid the side effects associated with
chemotherapy.
In the present study, a mouse xenograft model was
used to examine the tumor-suppressive efficacy of fisetin in
vivo on colon and colorectal cancers. Animal (CT-26) and human
(HCT116) colon and colorectal cancer cells, respectively, were used
to establish xenograft tumor models. Fisetin treatment was shown to
suppress the growth of tumors formed by CT-26 cells, but not HCT116
cells. However, treatment with fisetin combined with radiation
exhibited enhanced tumor suppressive effects on CT-26 and HCT116
xenograft tumors. This finding supports the potential application
of fisetin to adjuvant radiotherapy for colorectal cancers. As
fisetin did not exert toxicity in the current study, or in other
reports (45,46), its use as an adjuvant treatment should
be feasible.
In the present study, CT-26 and HCT116 cell lines,
which express wild-type p53, formed tumors that were sensitive to
radiation treatment. Since p53 is known to be upregulated by
radiation (47), it is reasonable
that this phenomenon was observed in vivo in the present
study. Importantly, the current study used clinically comparable
fractionated radiation of 2-Gy/day/fraction for 5 days (48). Compared with this regime, the
fisetin-combined radiation treatments only irradiated tumors at 2
Gy twice in one week, yet remained more efficient than the
fractionated radiation on tumor suppression, suggesting that
fisetin enhances the tumor response to radiation in vivo.
Thus, the use of fisetin in adjuvant radiotherapy may reduce total
radiation exposure and improve the quality of life of patients
during and following treatment. Further investigations into the
optimal combination for colorectal cancer therapy are
warranted.
The genetic background of a tumor is known to
influence the prognosis (49,50). The authors of the present study were
interested in p53 and securin because of the results of our
previous studies (15,27). Briefly, fisetin was shown to enhance
the radiosensitivity of p53-mutant human HT-29 colorectal cancer
cells, and promote apoptosis in securin-depleted human HCT-116
colorectal cancer cells. Consistent with a previous report
(14), in the present study, we also
showed that fisetin could induce p53 protein expression in
wild-type and securin-null HCT116 cells. Furthermore, expression of
the securin protein was downregulated by fisetin regardless of the
p53 expression status. In addition, in the current study xenograft
securin-null colorectal tumors exhibited increased sensitivity to
fisetin compared with the untreated control. However, individual
variance in the tumor-bearing mice may have reduced the extent of
tumor suppression resulting from fisetin treatment. Therefore, an
increased sample size is required to validate the effect of fisetin
on securin-null colorectal cancers. In addition, this limitation
should be considered for the investigation of fisetin and radiation
combination treatment. In the current study, p53 protein expression
was still induced in securin-null HCT116 cells by fisetin,
indicating that p53 and securin protein expression should be
modulated by fisetin and radiation combination treatment in order
to achieve optimal tumor suppression.
In conclusion, the present study used mouse
xenograft tumor models to investigate the effect of fisetin alone
or in combination with radiation on colon and colorectal tumor
growth. The results showed that fisetin treatment alone was
sufficient to suppress murine CT-26 colon tumors, but not human
HCT116 colorectal tumors. Interestingly, a combination of fisetin
and radiation treatment enhanced the tumor suppression compared
with fractionated irradiation alone. In addition, the results of
the current study revealed that securin-null HCT116 tumors
exhibited increased sensitivity to fisetin treatment, although this
observation needs to be validated in a larger sample set.
Investigating whether securin is the key molecule in mediating the
effects of fisetin and radiation combination treatment on colon and
colorectal cancers warrants future exploration. To the best of our
knowledge, this is the first report demonstrating the therapeutic
efficacy of fisetin and radiation combination treatment on colon
and colorectal cancer in vivo. The results of the present
study provide important preclinical information for evaluating the
potential use of fisetin in adjuvant cancer radiotherapy.
Acknowledgements
The present study was supported by the Taipei City
Government Department of Health (Taipei, Taiwan; grant no.
10101-62-034) and the Ministry of Science and Technology (Taipei,
Taiwan; grant nos. 102-2628-B-010-012-MY3 and
105-2628-B-010-013-MY3).
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Davies NJ, Batehup L and Thomas R: The
role of diet and physical activity in breast, colorectal, and
prostate cancer survivorship: A review of the literature. Br J
Cancer. 105:(Suppl 1). S52–S73. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Doll R and Peto R: The causes of cancer:
Quantitative estimates of avoidable risks of cancer in the United
States today. J Natl Cancer Inst. 66:1191–1308. 1981.PubMed/NCBI
|
|
4
|
Lin J, Zhang SM, Cook NR, Rexrode KM, Liu
S, Manson JE, Lee IM and Buring JE: Dietary intakes of fruit,
vegetables, and fiber, and risk of colorectal cancer in a
prospective cohort of women (United States). Cancer Causes Control.
16:225–233. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hamaya Y, Guarinos C, Tseng-Rogenski SS,
Iwaizumi M, Das R, Jover R, Castells A, Llor X, Andreu M and
Carethers JM: Efficacy of Adjuvant 5-Fluorouracil Therapy for
Patients with EMAST-Positive Stage II/III Colorectal Cancer. PLoS
One. 10:e01275912015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Saltz LB and Minsky B: Adjuvant therapy of
cancers of the colon and rectum. Surg Clin North Am. 82:1035–1058.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Allal AS, Kähne T, Reverdin AK, Lippert H,
Schlegel W and Reymond MA: Radioresistance-related proteins in
rectal cancer. Proteomics. 4:2261–2269. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma W, Yu J, Qi X, Liang L, Zhang Y and
Ding Y, Lin X, Li G and Ding Y: Radiation-induced microRNA-622
causes radioresistance in colorectal cancer cells by
down-regulating Rb. Oncotarget. 6:15984–15994. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lock K, Pomerleau J, Causer L, Altmann DR
and McKee M: The global burden of disease attributable to low
consumption of fruit and vegetables: Implications for the global
strategy on diet. Bull World Health Organ. 83:100–108.
2005.PubMed/NCBI
|
|
10
|
Martinez-Perez C, Ward C, Cook G, Mullen
P, McPhail D, Harrison DJ and Langdon SP: Novel flavonoids as
anti-cancer agents: Mechanisms of action and promise for their
potential application in breast cancer. Biochem Soc Trans.
42:1017–1023. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Orlikova B, Menezes JC, Ji S, Kamat SP,
Cavaleiro JA and Diederich M: Methylenedioxy flavonoids: Assessment
of cytotoxic and anti-cancer potential in human leukemia cells. Eur
J Med Chem. 84:173–180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Imai M, Kikuchi H, Denda T, Ohyama K,
Hirobe C and Toyoda H: Cytotoxic effects of flavonoids against a
human colon cancer derived cell line, COLO 201: A potential natural
anti-cancer substance. Cancer Lett. 276:74–80. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu X, Jung JI, Cho HJ, Lim DY, Lee HS,
Chun HS, Kwon DY and Park JH: Fisetin inhibits the activities of
cyclin-dependent kinases leading to cell cycle arrest in HT-29
human colon cancer cells. J Nutr. 135:2884–2890. 2005.PubMed/NCBI
|
|
14
|
Lim DY and Park JH: Induction of p53
contributes to apoptosis of HCT-116 human colon cancer cells
induced by the dietary compound fisetin. Am J Physiol Gastrointest
Liver Physiol. 296:G1060–G1068. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen WS, Lee YJ, Yu YC, Hsaio CH, Yen JH,
Yu SH, Tsai YJ and Chiu SJ: Enhancement of p53-mutant human
colorectal cancer cells radiosensitivity by flavonoid fisetin. Int
J Radiat Oncol Biol Phys. 77:1527–1535. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suh Y, Afaq F, Johnson JJ and Mukhtar H: A
plant flavonoid fisetin induces apoptosis in colon cancer cells by
inhibition of COX2 and Wnt/EGFR/NF-kappaB-signaling pathways.
Carcinogenesis. 30:300–307. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li J, Cheng Y, Qu W, Sun Y, Wang Z, Wang H
and Tian B: Fisetin, a dietary flavonoid, induces cell cycle arrest
and apoptosis through activation of p53 and inhibition of NF-kappa
B pathways in bladder cancer cells. Basic Clin Pharmacol Toxicol.
108:84–93. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chou RH, Hsieh SC, Yu YL, Huang MH, Huang
YC and Hsieh YH: Fisetin inhibits migration and invasion of human
cervical cancer cells by down-regulating urokinase plasminogen
activator expression through suppressing the p38 MAPK-dependent
NF-κB signaling pathway. PloS one. 8:e719832013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Salmela AL, Pouwels J, Varis A, Kukkonen
AM, Toivonen P, Halonen PK, Perälä M, Kallioniemi O, Gorbsky GJ and
Kallio MJ: Dietary flavonoid fisetin induces a forced exit from
mitosis by targeting the mitotic spindle checkpoint.
Carcinogenesis. 30:1032–1040. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Haddad AQ, Venkateswaran V, Viswanathan L,
Teahan SJ, Fleshner NE and Klotz LH: Novel antiproliferative
flavonoids induce cell cycle arrest in human prostate cancer cell
lines. Prostate Cancer Prostatic Dis. 9:68–76. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pei L and Melmed S: Isolation and
characterization of a pituitary tumor-transforming gene (PTTG). Mol
Endocrinol. 11:433–441. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rustgi AK: Securin a new role for itself.
Nat Genet. 32:222–224. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zou H, McGarry TJ, Bernal T and Kirschner
MW: Identification of a vertebrate sister-chromatid separation
inhibitor involved in transformation and tumorigenesis. Science.
285:418–422. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tong Y and Eigler T: Transcriptional
targets for pituitary tumor-transforming gene-1. J Mol Endocrinol.
43:179–185. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim DS, Franklyn JA, Smith VE, Stratford
AL, Pemberton HN, Warfield A, Watkinson JC, Ishmail T, Wakelam MJ
and McCabe CJ: Securin induces genetic instability in colorectal
cancer by inhibiting double-stranded DNA repair activity.
Carcinogenesis. 28:749–759. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nagao K, Adachi Y and Yanagida M:
Separase-mediated cleavage of cohesin at interphase is required for
DNA repair. Nature. 430:1044–1048. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu SH, Yang PM, Peng CW, Yu YC and Chiu
SJ: Securin depletion sensitizes human colon cancer cells to
fisetin-induced apoptosis. Cancer Lett. 300:96–104. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang YT, Lin CI, Chien PH, Tang TT, Lin J
and Chao JI: The depletion of securin enhances butein-induced
apoptosis and tumor inhibition in human colorectal cancer. Chem
Biol Interact. 220:41–50. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen WS, Yu YC, Lee YJ, Chen JH, Hsu HY
and Chiu SJ: Depletion of securin induces senescence after
irradiation and enhances radiosensitivity in human cancer cells
regardless of functional p53 expression. Int J Radiat Oncol Biol
Phys. 77:566–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Haupt S, di Agostino S, Mizrahi I,
Alsheich-Bartok O, Voorhoeve M, Damalas A, Blandino G and Haupt Y:
Promyelocytic leukemia protein is required for gain of function by
mutant p53. Cancer Res. 69:4818–4826. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH Image to ImageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bernal JA, Luna R, Espina A, Lázaro I,
Ramos-Morales F, Romero F, Arias C, Silva A, Tortolero M and
Pintor-Toro JA: Human securin interacts with p53 and modulates
p53-mediated transcriptional activity and apoptosis. Nat Genet.
32:306–311. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang PM, Tseng HH, Peng CW, Chen WS and
Chiu SJ: Dietary flavonoid fisetin targets caspase-3-deficient
human breast cancer MCF-7 cells by induction of
caspase-7-associated apoptosis and inhibition of autophagy. Int J
Oncol. 40:469–478. 2012.PubMed/NCBI
|
|
34
|
Khan N, Afaq F, Syed DN and Mukhtar H:
Fisetin, a novel dietary flavonoid, causes apoptosis and cell cycle
arrest in human prostate cancer LNCaP cells. Carcinogenesis.
29:1049–1056. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li J, Qu W, Cheng Y, Sun Y, Jiang Y, Zou
T, Wang Z, Xu Y and Zhao H: The inhibitory effect of intravesical
fisetin against bladder cancer by induction of p53 and
down-regulation of NF-kappa B pathways in a rat bladder
carcinogenesis model. Basic Clin Pharmacol Toxicol. 115:321–329.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kang KA, Piao MJ and Hyun JW: Fisetin
induces apoptosis in human nonsmall lung cancer cells via a
mitochondria-mediated pathway. In Vitro Cell Dev Biol Anim.
51:300–309. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Syed DN, Chamcheu JC, Khan MI, Sechi M,
Lall RK, Adhami VM and Mukhtar H: Fisetin inhibits human melanoma
cell growth through direct binding to p70S6K and mTOR: Findings
from 3-D melanoma skin equivalents and computational modeling.
Biochem Pharmacol. 89:349–360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pal HC, Sharma S, Strickland LR, Katiyar
SK, Ballestas ME, Athar M, Elmets CA and Afaq F: Fisetin inhibits
human melanoma cell invasion through promotion of mesenchymal to
epithelial transition and by targeting MAPK and NFκB signaling
pathways. PloS one. 9:e863382014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Szliszka E, Helewski KJ, Mizgala E and
Krol W: The dietary flavonol fisetin enhances the
apoptosis-inducing potential of TRAIL in prostate cancer cells. Int
J Oncol. 39:771–779. 2011.PubMed/NCBI
|
|
40
|
Khan N, Syed DN, Ahmad N and Mukhtar H:
Fisetin: A dietary antioxidant for health promotion. Antioxid Redox
Signal. 19:151–162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sahu BD, Kumar JM and Sistla R: Fisetin, a
dietary flavonoid, ameliorates experimental colitis in mice:
Relevance of NF-κB signaling. J Nutr Biochem. 28:171–182. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tripathi R, Samadder T, Gupta S, Surolia A
and Shaha C: Anticancer activity of a combination of cisplatin and
fisetin in embryonal carcinoma cells and xenograft tumors. Mol
Cancer Ther. 10:255–268. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Breugom AJ, van Gijn W, Muller EW,
Berglund Å, van den Broek CB, Fokstuen T, Gelderblom H, Kapiteijn
E, Leer JW, Marijnen CA, et al: Adjuvant chemotherapy for rectal
cancer patients treated with preoperative (chemo)radiotherapy and
total mesorectal excision: A dutch colorectal cancer group (DCCG)
randomized phase III trial. Ann Oncol. 26:696–701. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Colorectal Cancer Collaborative Group, .
Adjuvant radiotherapy for rectal cancer: A systematic overview of
8,507 patients from 22 randomised trials. Lancet. 358:1291–1304.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Currais A, Prior M, Dargusch R, Armando A,
Ehren J, Schubert D, Quehenberger O and Maher P: Modulation of p25
and inflammatory pathways by fisetin maintains cognitive function
in Alzheimer's disease transgenic mice. Aging Cell. 13:379–390.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
da Costa MP, Bozinis MC, Andrade WM, Costa
CR, da Silva AL, de Oliveira CM Alves, Kato L, Ode F Fernandes,
Souza LK and Mdo R Silva: Antifungal and cytotoxicity activities of
the fresh xylem sap of Hymenaea courbaril L. and its major
constituent fisetin. BMC Complement Altern Med. 14:2452014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fei P and El-Deiry WS: P53 and radiation
responses. Oncogene. 22:5774–5783. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fertil B and Malaise EP: Inherent cellular
radiosensitivity as a basic concept for human tumor radiotherapy.
Int J Radiat Oncol Biol Phys. 7:621–629. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lindström LS, Hall P, Hartman M, Wiklund F
and Czene K: Is genetic background important in lung cancer
survival? PloS one. 4:e55882009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ashworth A, Lord CJ and Reis-Filho JS:
Genetic interactions in cancer progression and treatment. Cell.
145:30–38. 2011. View Article : Google Scholar : PubMed/NCBI
|