Introduction
Gastric cancer (GC) is a major health problem, and
is one of the leading causes of morbidity and mortality worldwide,
representing the second most common cause of cancer-associated
mortality (1). In 2012, GC caused
723,000 mortalities worldwide (2),
due to its poor prognosis and the limited efficacy of treatment. It
is widely accepted that environmental factors, including
Helicobacter pylori infection (3), Epstein-Barr virus infection (4), a poor diet, smoking and obesity
(5) contribute to gastric
carcinogenesis. Additionally, certain genetic factors have been
implicated in the development of GC, including somatic mutations,
gene amplifications and deletions, epigenetic inactivation of genes
and aberrant DNA methylation (4,6). Molecular
profiling of GC can be performed using gene expression microarray
analysis or DNA sequencing (7–9), which
facilitates the identification of putative biomarkers for subtype
classification, prognosis and therapeutic targets. However, the
molecular mechanisms underlying the progression of GC remain poorly
understood.
In addition to protein coding genes, microRNAs
(miRNAs/miRs) serve important roles in human carcinogenesis. miRNAs
are short (~22 nucleotides) non-coding RNAs that regulate gene
expression primarily through translational repression or
transcriptional degradation, and as such effect important cellular
processes, including cell proliferation, cell death and
tumorigenesis (10–12). Previous studies have suggested
oncogenic and tumor suppressive roles for miRNAs in cancer
(13,14). miRNAs also have the potential to be
cancer biomarkers in terms of their tissue-specific expression and
aberrant expression in cancer cells (15). miRNAs can be measured through
high-throughput microarray analysis. In GC, aberrant miRNA
expression profiles have been associated with GC progression,
prognosis and pathogenesis (16,17) by
perturbing the function of target genes (18–21). For
example, miR-148a is significantly downregulated in GC cell lines
and tissue (22–24).
Numerous aberrantly expressed miRNAs have been
identified in GC; however, only a fraction of these have been
functionally investigated and novel deregulated miRNAs in GC remain
to be explored. In the present study, two public miRNA expression
profile datasets were examined to identify novel aberrantly
expressed miRNAs in GC. One of the differentially expressed miRNAs
identified, miR-564, which was downregulated, was validated in the
tissue samples of patients with GC patients by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis. The target genes of miR-564 were then predicted and it
was demonstrated that miR-564 could bind to the 3′-untranslated
region (UTR) of transcription factor E2F3 (E2F3). Finally,
overexpression of miR-564 was found to significantly inhibit the
mRNA and protein levels of E2F3 in GC cells. In conclusion, the
results of the current study indicate that miR-564 is an important
novel potential tumor suppressor gene in gastric
carcinogenesis.
Materials and methods
Public miRNA microarray data
processing
Two public miRNA microarray datasets for GC and
normal gastric tissue were obtained from the Gene Expression
Omnibus (GEO; GEO nos. GSE23739 and GSE30070). The limma software
package (version 3.30.12; https://bioconductor.org/packages/release/bioc/html/limma.html)
was used to determine differentially expressed miRNAs in the two
data sets between GC and normal gastric tissues. The upregulated
and downregulated miRNAs from the two data sets were overlapped to
generate a consensus list of differentially expressed miRNAs, which
was visualized as a heatmap using Multiple Experiment Viewer
software (version 4.9.0; https://sourceforge.net/projects/mev-tm4/).
Prediction of miRNA targets
TargetScan (http://www.targetscan.org) was used to predict target
genes for miR-564.
Functional annotation
The enrichment of KEGG pathways for targeted genes
was determined by DAVID (25).
Cytoscape (26) and Enrichment Map
(27) were used for visualization of
the network.
Patients and tissue samples
To validate the results of the miRNA microarray
analysis, 8 pairs of GC and adjacent non-cancerous gastric tissue
samples were obtained. All samples were obtained from patients with
gastric cancer who underwent surgical resection at No. 161 Hospital
of the People's Liberation Army (Wuhan, China) between May and
October 2014. All the participants were histologically confirmed to
have gastric adenocarcinoma and did not receive any other forms of
therapy prior to the time of enrollment. Written informed consent
was obtained from all patients and the procedures used in the
present study were approved by the Institutional Review Board of
No. 161 Hospital of the People's Liberation Army.
Total RNA extraction and RT-qPCR
analysis
All tissue samples were frozen with liquid nitrogen
immediately following surgical resection. For miRNA RT-qPCR
analysis, total RNA was extracted from the samples using the
miRNeasy Mini kit (Qiagen, Inc., Valencia, CA, USA) according to
the manufacturer's protocol. For DNA RT-qPCR analysis, total RNA
was extracted with TRIzol Reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in accordance with the
manufacturer's instructions. RNU6B and β-Actin were used as
reference genes for quantification of the expression of miR-564 and
E2F3, respectively. The sequences of these primers are listed in
Table I. PCR reaction volumes used
were as follows: 2X SYBR®-Green Real-time PCR Master mix
5 µl (Toyobo Co., Ltd., Osaka, Japan); cDNA50 ng, forward primer
and reverse primer 5 µM; distilled water up to the volume of 10 µl.
qPCR was performed at 50°C for 2 min, followed by 40 cycles of 95°C
for 15 sec and 60°C for 1 min on an iQ5 Real-Time PCR Detection
System (Bio-Rad Laboratories, Inc., Hercules, CA, USA). All
reactions were carried out in triplicate. Expression was quantified
using the 2−ΔΔCq method (28).
 | Table I.Primers used for quantitative
polymerase chain reaction. |
Table I.
Primers used for quantitative
polymerase chain reaction.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| RNU6B |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
| β-Actin |
CTGGAACGGTGAAGGTGACA |
AAGGGACTTCCTGTAACAATGCA |
| miR-564 |
ACACTCCAGCTGGGAGGCACGGTGTCA |
TGGTGTCGTGGAGTCG |
| E2F3 |
GCACTACGAAGTCCAGATAGTCC |
AGACTGCAGCCCATCCATTG |
Cell culture
The GC cell line, SGC-7901 was obtained from the
Institute of Biochemistry and Cell Biology of the Chinese Academy
of Sciences (Shanghai, China). The cells were maintained in
Dulbecco's modified Eagle medium, supplemented with 10% fetal
bovine serum (Hyclone; GE Healthcare Life Sciences, Logan, UT,
USA), 100 U/ml penicillin, and 100 µg/ml streptomycin. The cells
were incubated in an atmosphere of 5% CO2 at 37°C.
Luciferase reporter assay
The 3′-UTR of E2F3 mRNA containing the predicted
miR-564 binding site using TargetScan was amplified by PCR using
the PCR Amplification kit (Takara Biotechnology Co., Ltd., Dalian,
China). The PCR product was digested and cloned into the
psiCHECK™-2 reporter vector (Promega Corporation, Madison, WI, USA)
to produce psiCHECK-2-E2F3-3′UTR reporter plasmids. Cells were
seeded in 24-well plates at an initial density of 1×105
cells/well and cultured for 24 h at 37°C in the presence of 5%
CO2. A total of 200 ng psiCHECK-2-E2F3-3′UTR and 100
nmol/l hsa-miR-564 mimic (or miRNA mimics control, NC group) were
synthesized at GenePharma Co., Ltd. (Suzhou, China) and
cotransfected into the SGC7901 cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h, the cells were lyzed by adding 100
µl Passive Lysis Buffer (Promega Corporation; E194A) and reporter
activity was determined using a Dual-Luciferase®
Reporter Assay kit (Promega Corporation; E1910) according to the
manufacturer's protocol.
Western blotting
Proteins were isolated via lysing cells in Cell
Lysis Buffer (Cell Signaling Technology, Inc., Danvers, MA, USA)
followed by centrifugation at 13,000 × g for 10 min at 4°C. Protein
(10 µg/lane) was loaded onto a 10% gel, subjected to SDS-PAGE and
subsequently transferred to a polyvinylidene difluoride membrane.
Membranes were probed with polyclonal rabbit antibodies directed
against anti-E2F3 (catalog no., ab50917; dilution, 1:500; Abcam,
Cambridge, MA, USA) overnight at 4°C, then incubated in horseradish
peroxidase-conjugated anti-rabbit secondary antibodies (catalog
no., GB23303; dilution, 1:3,000; Servicebio, Wuhan, China) for 1 h
at room temperature. An anti-β-tubulin rabbit polyclonal antibody
(catalog no., ab151318; dilution, 1:5,000; Abcam) was used as a
loading control. The blot was developed using Enhanced
Chemiluminescence Substrate Solution (Beyotime Institute of
Biotechnology, Haimen, China) and images of the blot captured using
the FluorChem™ Imaging System (ProteinSimple; Bio-Techne,
Minneapolis, MN, USA). The intensity of each spot was analyzed with
AlphaEaseFC™ 4.0 imaging software (AlphaInnotech, San Leandro, CA,
USA).
Statistical analysis
All data are presented as the mean ± standard
deviation of ≥3 independent experiments. The results were analyzed
using a two-tailed Student's t-test. P<0.05 Results were
considered statistically significant at P<0.05.
Results
Identification of novel aberrantly
expressed miRNAs in GC
To identify novel miRNAs associated with GC, two
publicly available miRNA expression profiles were analyzed. Of the
two data sets, GSE23739 contained 40 GC tissue samples and 40
normal gastric tissue samples, whereas GSE30070 contained 90 GC
tissue samples and 34 normal gastric tissue samples. Differential
expression analysis was performed on the datasets, then by
comparing and overlapping the differentially expressed miRNAs, a
list of miRNAs that were differentially expressed in GSE23739 and
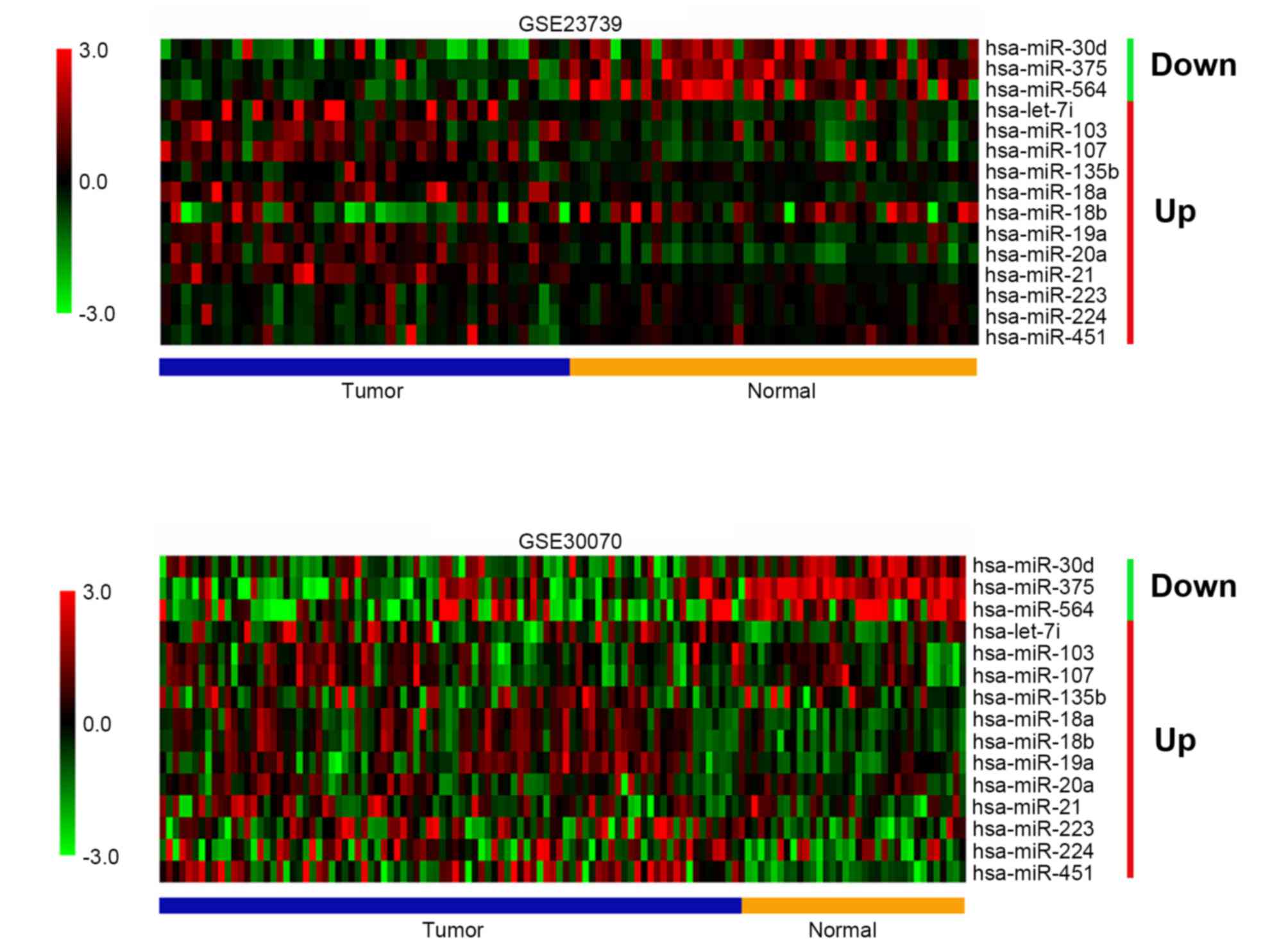
GSE30070 were obtained. As illustrated in Fig. 1, 3 miRNAs were downregulated in GCs
and 12 miRNAs were upregulated in GCs. One of the downregulated
miRNAs, miR-564, was identified to be a novel miRNA that was
dysregulated in GC, though it has been reported that miR-564 was
increased in H. pylori-positive compared with the H.
pylori-negative GC tissues (29).
miR-564 expression is downregulated in
GC
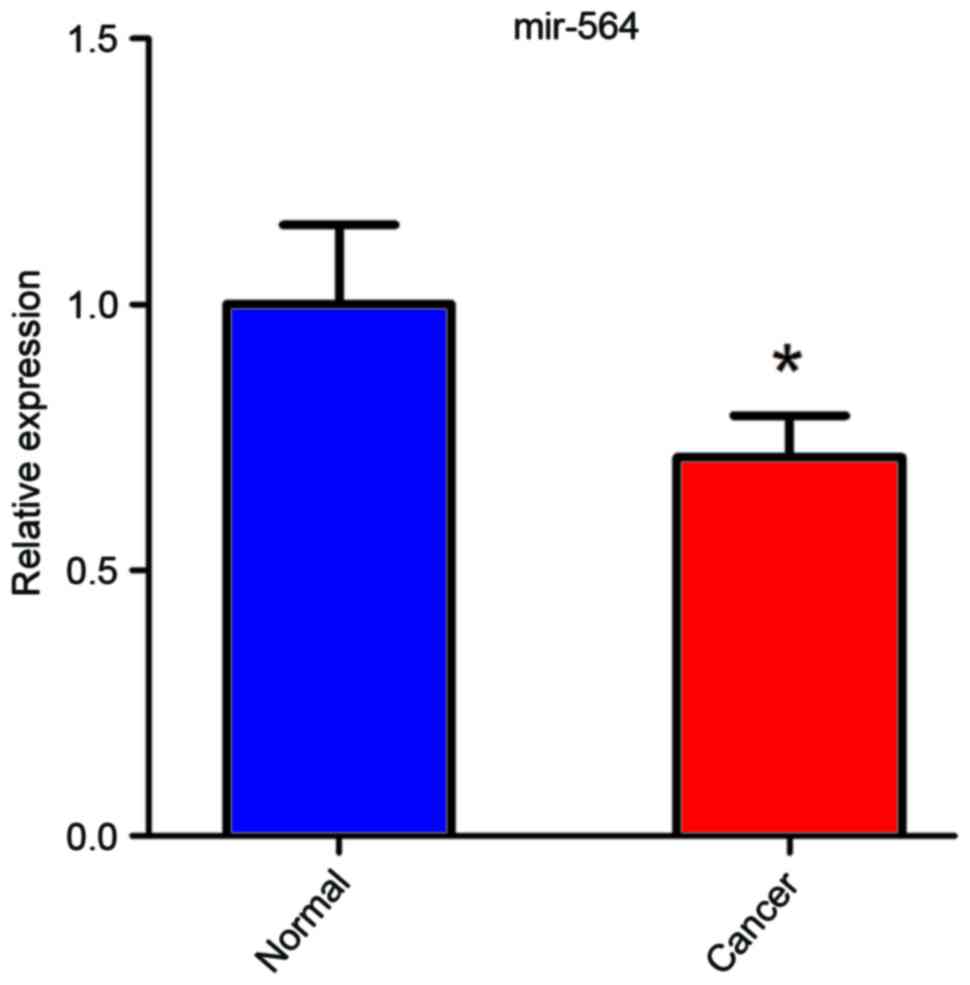
To evaluate the role of miR-564 in the development
of GC, the expression levels of miR-564 were measured in 8 human
primary GC tissues and paired normal adjacent gastric tissues. The
results revealed that the GC samples had a significantly lower
level of miR-564 expression compared with adjacent normal gastric
tissue (P<0.05; Fig. 2). These
data suggest that miR-564 is downregulated in GC and indicate that
it is a potential tumor suppressor.
Predicting the putative targets of
miR-564
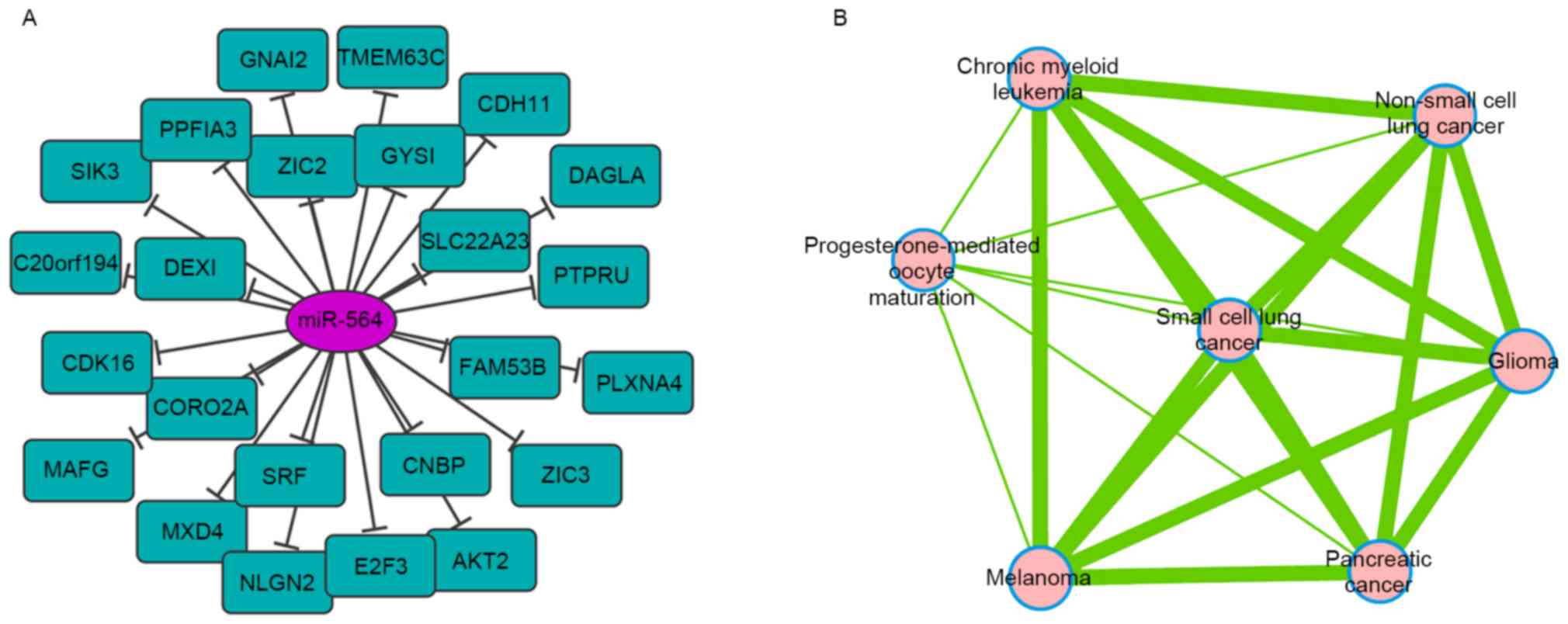
To investigate the potential role of miR-564, target
genes for this miRNA were predicted using TargetScan. As a result,
25 candidate targets were identified (Fig. 3A). Functional pathway enrichment
analysis revealed that the proteins encoded by these genes were
enriched in various types of cancer signaling pathways (Fig. 3B). One predicted target, E2F3, a
transcriptional activator of E2F-family of transcription factors,
was associated with numerous cancer-associated signaling pathways.
This indicates that miR-564 regulates GC progression through
targeting E2F3.
E2F3 is a direct target of miR-564 in
GC cells
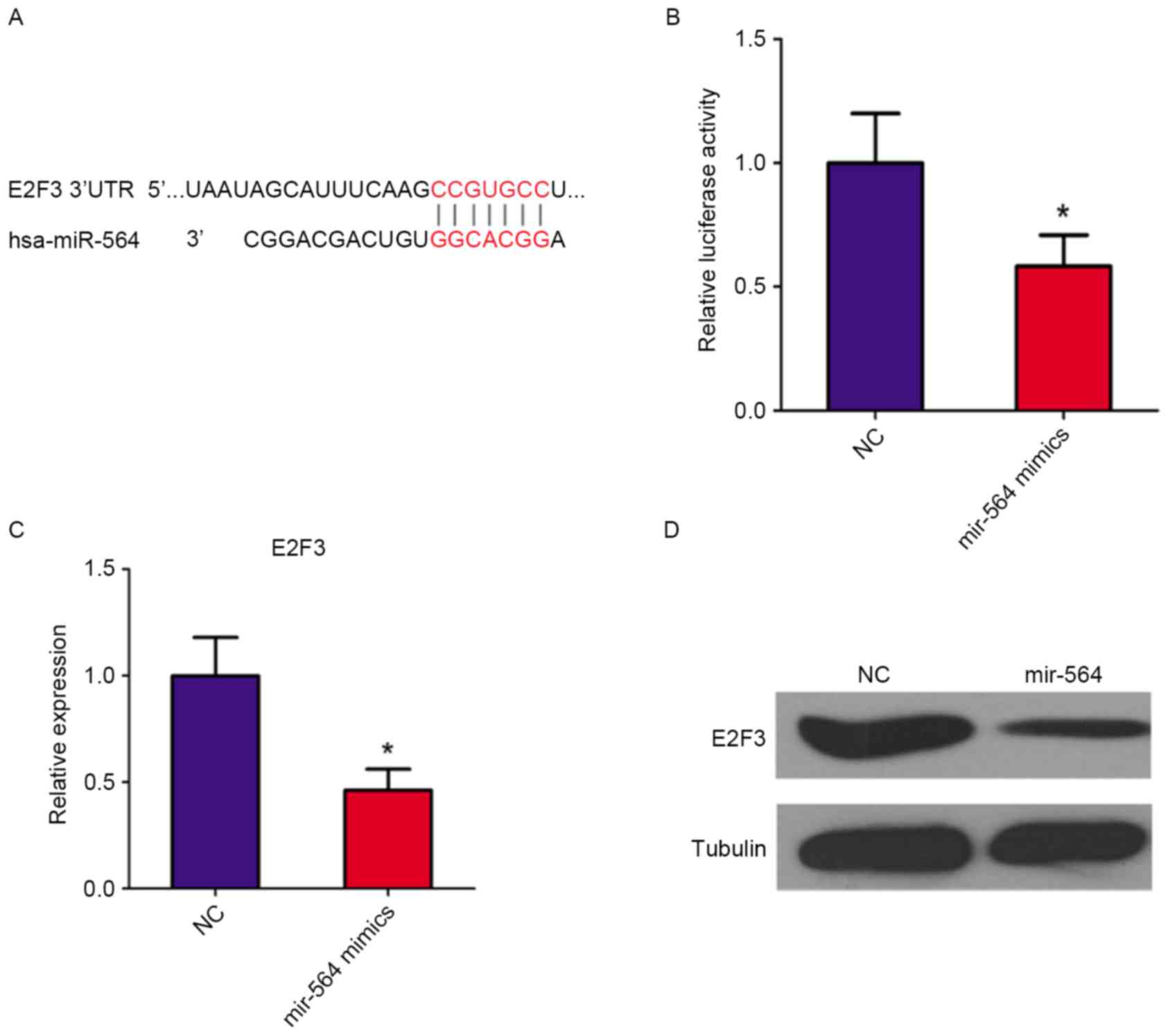
To validate E2F3 as a target of miR-564, luciferase
reporter assays were conducted using the E2F3-3′UTR in
miR-564-transfected SGC7901 cells. Position 416–422 of E2F3-3′UTR
was predicted as the target binding region for miR-564 (Fig. 4A). The luciferase activity of the
E2F3-3′UTR plasmid significantly decreased by 40% following
treatment with the miR-564 mimic (P<0.05 vs. the NC group;
Fig. 4B). These results indicate that
miR-564 suppresses gene expression through binding to the 3′UTR of
E2F3 mRNA. Moreover, RT-qPCR (Fig.
4C) and western blot (Fig. 4D)
analyses demonstrated that the expression of E2F3 mRNA and protein,
respectively, were decreased following treatment with the miR-564
mimic. These results indicate that E2F3 is a direct target of
miR-564.
Discussion
Accumulating evidence has demonstrated that miRNAs
regulate cancer development and progression by acting as tumor
suppressors or oncogenes (13,14).
Through examining two public miRNA expression profiles the present
study identified 15 aberrantly expressed miRNAs in GC, including 3
downregulated and 12 upregulated miRNAs. Some of these miRNAs, for
example miR-21, have previously been demonstrated to be upregulated
in GC (30). In addition, the present
study identified a novel aberrantly expressed miRNA, miR-564.
RT-qPCR analysis of 8 GC tissue and adjacent normal tissue samples
validated that miR-564 was downregulated in GC, which was
consistent with the results of miRNA microarray analysis. miR-564
has been reported to be aberrantly expressed in breast cancer
(31) and chronic myeloid leukemia
(32), indicating that is serves a
role in cancer development. However, the molecular mechanism
underlying the downregulation of miR-564 in GC remains unknown.
Genomic deletions in miRNAs have been reported as a mechanism for
miRNA downregulation, including that of miR-101, −15a and −16-1
(33,34). Thus, examining whether genomic loss
has occurred at chromosome 3p21.31, where miR-564 is located, in GC
may provide evidence to explain this. Other factors, including DNA
methylation, histone modification, interactions between
transcriptional suppressors and the miR-564 promoter, and
post-transcriptional regulation may also serve role in the
downregulation of miR-564 in GC. Further studies are required to
determine the mechanism underlying miR-564 downregulation in
GC.
The predicted target genes of miR-564, particularly
E2F3, were functionally enriched in the signaling pathways of
several cancers, including non-small cell lung cancer, glioma,
melanoma, pancreatic cancer, chronic myeloid leukemia and small
cell lung cancer. E2F3 is known to be a potent regulator of cell
cycle progression and apoptosis, with the capacity to stimulate
quiescent cells to proliferate or to induce cell apoptosis.
Deregulation of E2F3, either through overexpression (35) or inactivation (36) by repressor mechanisms, is a frequent
oncogenic event in human tumorigenesis. In GC, 4 miRNAs (miR-141,
−449a, −145 and −125a-5p) have been reported to inhibit cancer cell
proliferation through directly targeting E2F3 (37–40).
Furthermore, E2F3 has been demonstrated to be upregulated in human
GC tissue samples compared with paired normal tissues (38). The luciferase reporter assay performed
in the current study confirmed that miR-564 directly binds the
3′-UTR of E2F3 mRNA to inhibit E2F3 protein translation in GC
cells. In addition, overexpression of miR-564 decreased the mRNA
and protein levels of E2F3. Although no cell proliferation assay
was performed in the current study, the results obtained suggest
that the downregulation of miR-564 serves a tumor suppressor role
in GC and may inhibits cancer cell proliferation through directly
targeting E2F3.
It should be noted that although E2F3 was
demonstrated to be a direct target gene of miR-564, the potential
antioncogenic effects of miR-564 may not be fully explained by its
ability to regulate a single gene alone, particularly as a previous
study identified that single miRNAs frequently regulate numerous
genes (41). Therefore, other
predicted target genes of miR-564 should be investigated in future
studies. For example, another putative target of miR-564 identified
in the current study, AKT serine/threonine kinase 2 (AKT2) is an
essential mediator of tumorigenesis and thought to be an ideal
target for the treatment of malignancies. Previous studies have
reported that miRNA-137 and miRNA-29 s may target AKT2 to inhibit
the tumorigenesis and invasiveness of GC cells (42,43).
However, the present study had several limitations, including the
fact that there was no clinical data. Further studies are required
to determine the effects of miR-564 in patients with GC with
different stages of cancer, and to evaluate its prognostic
value.
In conclusion, the present study revealed that
miR-564 is frequently downregulated in GC and that its potential
tumor suppressor functions in GC are associated with directly
targeting E2F3. These results indicate that miR-564 represents a
novel target for GC therapy.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Uemura N, Okamoto S, Yamamoto S, Matsumura
N, Yamaguchi S, Yamakido M, Taniyama K, Sasaki N and Schlemper RJ:
Helicobacter pylori infection and the development of gastric
cancer. N Engl J Med. 345:784–789. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular characterization of gastric adenocarcinoma.
Nature. 513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang X, Ni Z, Duan Z, Xin Z, Wang H, Tan
J, Wang G and Li F: Overexpression of E2F mRNAs associated with
gastric cancer progression identified by the transcription factor
and miRNA co-regulatory network analysis. PLoS One.
10:e01169792015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in Cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan IB, Ivanova T, Lim KH, Ong CW, Deng N,
Lee J, Tan SH, Wu J, Lee MH, Ooi CH, et al: Intrinsic subtypes of
gastric cancer, based on gene expression pattern, predict survival
and respond differently to chemotherapy. Gastroenterology.
141:476–485. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lei Z, Tan IB, Das K, Deng N, Zouridis H,
Pattison S, Chua C, Feng Z, Guan YK, Ooi CH, et al: Identification
of molecular subtypes of gastric cancer with different responses to
PI3-kinase inhibitors and 5-fluorouracil. Gastroenterology.
145:554–565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang K, Kan J, Yuen ST, Shi ST, Chu KM,
Law S, Chan TL, Kan Z, Chan AS, Tsui WY, et al: Exome sequencing
identifies frequent mutation of ARID1A in molecular subtypes of
gastric cancer. Nat Genet. 43:1219–1223. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartel DP: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kent OA and Mendell JT: A small piece in
the cancer puzzle: MicroRNAs as tumor suppressors and oncogenes.
Oncogene. 25:6188–6196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gaur A, Jewell DA, Liang Y, Ridzon D,
Moore JH, Chen C, Ambros VR and Israel MA: Characterization of
microRNA expression levels and their biological correlates in human
cancer cell lines. Cancer Res. 67:2456–2468. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ueda T, Volinia S, Okumura H, Shimizu M,
Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: A microRNA expression analysis. Lancet Oncol.
11:136–146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Zhang Y, Zhang Y, Ding J, Wu K and
Fan D: Survival prediction of gastric cancer by a seven-microRNA
signature. Gut. 59:579–585. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Petrocca F, Visone R, Onelli MR, Shah MH,
Nicoloso MS, de Martino I, Iliopoulos D, Pilozzi E, Liu CG, Negrini
M, et al: E2F1-regulated microRNAs impair TGFbeta-dependent
cell-cycle arrest and apoptosis in gastric cancer. Cancer cell.
13:272–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bandres E, Bitarte N, Arias F, Agorreta J,
Fortes P, Agirre X, Zarate R, Diaz-Gonzalez JA, Ramirez N, Sola JJ,
et al: microRNA-451 regulates macrophage migration inhibitory
factor production and proliferation of gastrointestinal cancer
cells. Clin Cancer Res. 15:2281–2290. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oh HK, Tan AL, Das K, Ooi CH, Deng NT, Tan
IB, Beillard E, Lee J, Ramnarayanan K, Rha SY, et al: Genomic loss
of miR-486 regulates tumor progression and the OLFM4 antiapoptotic
factor in gastric cancer. Clin Cancer Res. 17:2657–2667. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carvalho J, van Grieken NC, Pereira PM,
Sousa S, Tijssen M, Buffart TE, Diosdado B, Grabsch H, Santos MA,
Meijer G, et al: Lack of microRNA-101 causes E-cadherin functional
deregulation through EZH2 up-regulation in intestinal gastric
cancer. J Pathol. 228:31–44. 2012.PubMed/NCBI
|
|
22
|
Chen Z, Saad R, Jia P, Peng D, Zhu S,
Washington MK, Zhao Z, Xu Z and El-Rifai W: Gastric adenocarcinoma
has a unique microRNA signature not present in esophageal
adenocarcinoma. Cancer. 119:1985–1993. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng G, Xiong Y, Xu W, Wang Y, Chen F,
Wang Z and Yan Z: A two-microRNA signature as a potential biomarker
for early gastric cancer. Oncol Lett. 7:679–684. 2014.PubMed/NCBI
|
|
24
|
Sakamoto N, Naito Y, Oue N, Sentani K,
Uraoka N, Oo H Zarni, Yanagihara K, Aoyagi K, Sasaki H and Yasui W:
MicroRNA-148a is downregulated in gastric cancer, targets MMP7 and
indicates tumor invasiveness and poor prognosis. Cancer Sci.
105:236–243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
da W Huang, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.PubMed/NCBI
|
|
26
|
Cline MS, Smoot M, Cerami E, Kuchinsky A,
Landys N, Workman C, Christmas R, Avila-Campilo I, Creech M, Gross
B, et al: Integration of biological networks and gene expression
data using Cytoscape. Nat Protoc. 2:2366–2382. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Merico D, Isserlin R, Stueker O, Emili A
and Bader GD: Enrichment map: A network-based method for gene-set
enrichment visualization and interpretation. PLoS One.
5:e139842010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chang H, Kim N, Park JH, Nam RH, Choi YJ,
Lee HS, Yoon H, Shin CM, Park YS, Kim JM and Lee DH: Different
microRNA expression levels in gastric cancer depending on
Helicobacter pylori infection. Gut Liver. 9:188–196. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Effatpanah H, Yadegarazari R, Karami M,
Majlesi A, Shabab N and Saidijam M: Expression analysis of mir-21
and mir-221 in cancerous tissues from Iranian patients with gastric
cancer. Iran Biomed J. 19:188–193. 2015.PubMed/NCBI
|
|
31
|
Wang B, Li J, Sun M, Sun L and Zhang X:
miRNA expression in breast cancer varies with lymph node metastasis
and other clinicopathologic features. IUBMB Life. 66:371–377. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rokah OH, Granot G, Ovcharenko A, Modai S,
Pasmanik-Chor M, Toren A, Shomron N and Shpilberg O: Downregulation
of miR-31, miR-155, and miR-564 in chronic myeloid leukemia cells.
PLoS One. 7:e355012012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Varambally S, Cao Q, Mani RS, Shankar S,
Wang X, Ateeq B, Laxman B, Cao X, Jing X, Ramnarayanan K, et al:
Genomic loss of microRNA-101 leads to overexpression of histone
methyltransferase EZH2 in cancer. Science. 322:1695–1699. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Calin GA, Dumitru CD, Shimizu M, Bichi R,
Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, et al:
Frequent deletions and down-regulation of micro-RNA genes miR15 and
miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci
USA. 99:15524–15529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Oeggerli M, Tomovska S, Schraml P,
Calvano-Forte D, Schafroth S, Simon R, Gasser T, Mihatsch MJ and
Sauter G: E2F3 amplification and overexpression is associated with
invasive tumor growth and rapid tumor cell proliferation in urinary
bladder cancer. Oncogene. 23:5616–5623. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miles WO, Tschöp K, Herr A, Ji JY and
Dyson NJ: Pumilio facilitates miRNA regulation of the E2F3
oncogene. Genes Dev. 26:356–368. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou X, Ji G, Ke X, Gu H, Jin W and Zhang
G: MiR-141 inhibits gastric cancer proliferation by interacting
with long noncoding RNA MEG3 and down-regulating E2F3 expression.
Dig Dis Sci. 60:3271–3282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li X, Li H, Zhang R and Liu J:
MicroRNA-449a inhibits proliferation and induces apoptosis by
directly repressing E2F3 in gastric cancer. Cell Physiol Biochem.
35:2033–2042. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chang S, Gao L, Yang Y, Tong D, Guo B, Liu
L, Li Z, Song T and Huang C: miR-145 mediates the antiproliferative
and gene regulatory effects of vitamin D3 by directly targeting
E2F3 in gastric cancer cells. Oncotarget. 6:7675–7685. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu Y, Huang Z and Liu Y: Reduced
miR-125a-5p expression is associated with gastric carcinogenesis
through the targeting of E2F3. Mol Med Rep. 10:2601–2608.
2014.PubMed/NCBI
|
|
41
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu L, Chen J, Ding C, Wei S, Zhu Y, Yang
W, Zhang X, Wei X and Han D: MicroRNA-137 contributes to dampened
tumorigenesis in human gastric cancer by targeting AKT2. PLoS One.
10:e01301242015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang H, Cheng Y, Jia C, Yu S, Xiao Y and
Chen J: MicroRNA-29s could target AKT2 to inhibit gastric cancer
cells invasion ability. Med Oncol. 32:3422015. View Article : Google Scholar : PubMed/NCBI
|