Introduction
Epstein-Barr virus (EBV), also known as human herpes
virus 4, is one of the most common human viruses. The majority of
individuals are infected during infancy or childhood, such that the
majority of adults have been infected and have established lifelong
latent infections (1). EBV was the
first human oncovirus to be described, which was identified from a
Burkitt's lymphoma cell line in 1964 (2). Subsequent studies revealed that EBV
caused infectious mononucleosis and many different human
malignancies including nasopharyngeal carcinoma, Hodgkin's
lymphoma, extranodal natural killer/T-cell lymphoma,
lymphoproliferative disorders in immunocompromised hosts and a
number of types of gastric cancer (1).
EBV-associated gastric carcinoma (EBVaGC or EBV(+)
GC) is a distinct subtype of gastric cancer that accounts for
<10% of GC (gastric carcinoma). EBVaGC is defined by the
monoclonal proliferation of carcinoma cells with latent EBV
infection, as demonstrated using EBV-encoded small RNA (EBER) in
situ hybridization (1). EBVaGC
has characteristic clinicopathological features including a
lymphoepithelioma-like histology and favorable prognosis (3). It has been histologically subdivided
into two types: A lymphoepithelioma-like carcinoma (LELC) type and
conventional type adenocarcinoma. The LELC type is also referred to
as ‘GC with lymphoid stroma (GCLS)’. In total, >80% of GCLS
cases have been identified to exhibit EBV(+) tumors (3,4).
A previous study highlighted the important role of
activation-induced cytidine deaminase (AID), a nucleotide-editing
enzyme that is essential for the somatic hypermutation and
class-switch recombination of the immunoglobulin gene, as a genomic
modulator in inflammation-associated cancer development in
digestive organs including Helicobacter pylori-associated
gastric cancer, hepatitis C virus-positive hepatocellular carcinoma
and colitis-associated colon cancers (5). Consistent with the finding that a number
of transcription factors, including nuclear factor-κB (NF-κB),
mediate the expression of AID in B cells, the proinflammatory
cytokine stimulation of numerous types of gastrointestinal
epithelial cells, including gastric, colonic, hepatic and biliary
epithelia, induced aberrant AID expression through the NF-κB
signaling pathway (6). NF-κB is a key
molecule in inflammation-associated carcinogenesis, and NF-κB
activation in epithelial cells and malignant cells is involved in
generating genomic instability through aberrant AID expression,
cell growth, proliferation, survival, angiogenesis and
epithelial-mesenchymal transition (7).
The expression and activity of AID are tightly
regulated at the levels of transcription, post-transcription and
enzymatic function. Four distinct DNA regions (regions I to IV) of
the AID gene locus contain binding sites for multiple
transcription factors. Region I functions as a promoter containing
the binding site for NF-κB, a transcriptional activator. In B and
non-B cells, enhancer elements in region II bind to the activators
paired box 5 (PAX5) and transcription factor E2A, whereas silencer
elements in region II bind to the repressor proteins c-Myb
proto-oncogene (c-Myb) and transcription factor E2F in order to
counter the activities of transcriptional activators (8).
Honjo et al (9)
described pathogen-induced AID expression in gastric cancer (caused
by H. pylori), adult T cell leukemia/lymphoma (caused by
human T lymphotrophic virus 1), hepatoma (caused by hepatitis C
virus) and Burkitt's lymphoma (caused by EBV), but not in classical
Hodgkin's lymphoma (caused by EBV) (9). However, to the best of our knowledge,
aberrant AID expression in EBVaGC has not yet been elucidated. The
majority of EBVaGCs exhibit marked lymphoid stroma surrounding
cancer cells, which suggests pathogen-induced chronic inflammation.
Chronic inflammation, regardless of the infectious agent involved,
serves important roles in the development of various types of
cancer, particularly in digestive organs (5). Therefore, the aim of the present study
was to determine whether the inflammation-associated cancer
development theory through aberrant AID expression via a
pathogen-related NF-κB signal also applies to EBVaGC and GCLS.
In order to elucidate the question of whether or not
EBV-induced AID expression is a causative factor of carcinogenesis
in EBVaGC, the aberrant expression of AID and AID-regulatory
factors including NF-κB (activator), PAX5 (activator) and c-Myb
(repressor) were immunohistochemically evaluated and compared
between EBVaGC and EBV-non-associated GC, and also between GCLS and
GC without LS.
Materials and methods
Tumor samples
Formalin-fixed paraffin-embedded (FFPE) tumor
samples were obtained from 30 patients who had undergone surgical
resection at Tottori University Hospital between January 2005 and
December 2014, and who were histopathologically diagnosed with GCLS
(n=15; 14 males and 1 female) or age-sex-stage-matched other
gastric cancers (GC without LS) (n=15; 14 males and 1 female). The
mean age ± standard deviation (SD) was 64.7±9.4 years (range, 45–77
years). The tumor location was defined as the upper third, middle
third and lower third of the tissue or remnant. All samples were
histopathologically classified according to the Japanese
Classification of Gastric Carcinoma (10). The present study was approved by the
Institutional Ethics Committee of Tottori University (Yonago,
Japan; no. 2483).
Evaluation of surrounding mucosal
inflammation
The grading of gastritis in the non-cancerous mucosa
adjacent to a tumor was evaluated and scored according to the
updated Sydney System (USS) including H. pylori infection
(11). Gimenez staining was used to
detect H. pylori.
Immunohistochemical staining of AID,
NF-κB, c-Myb, PAX5 and latent membrane protein 1 (LMP-1)
Immuno-histochemical staining was performed using
the following primary antibodies: Anti-AID (#39-2500; dilution,
1:200; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), NF-κB (#ab86299, polyclonal; dilution, 1:100; Abcam,
Cambridge, MA, USA), PAX5 (#M7307; dilution, 1:30; Dako; Agilent
Technologies, Inc., Santa Clara, CA, USA), c-Myb (#LS-C49755;
dilution, 1:150; LifeSpan BioSciences, Inc., Seattle, WA, USA) and
LMP-1 (#M0897; dilution, 1:100; Dako; Agilent Technologies, Inc.).
Sections were cut to a thickness of 4 µm, deparaffinized in xylene
and dehydrated in descending dilutions of ethanol. Antigen
retrieval was performed using a pressure cooker at 120°C with
citrate buffer (pH 6.0) for 10 min (AID and c-Myb) or with Target
Retrieval Solution (pH 9.0; Dako; Agilent Technologies, Inc.) for
20 min (NF-κB, PAX5 and LMP-1). Endogenous peroxidase activity was
blocked by a 5-min incubation at room temperature with 3% hydrogen
peroxidase. Sections were incubated with primary antibodies and
antibody diluents with background-reducing components (Dako;
Agilent Technologies, Inc.) at room temperature for 120 min (AID,
NF-κB and PAX5) or at 4°C overnight (c-Myb and LMP-1). Horseradish
peroxidase (HRP) -conjugated goat anti-mouse IgG (EnVision™+/HRP;
#K4000; prediluted; AID, NF-κB, PAX5 and LMP-1) or goat anti-rabbit
IgG (EnVision™+/HRP; #K4002; prediluted, c-Myb) was applied as the
secondary antibody (both from Dako; Agilent Technologies, Inc.).
Sections were incubated with secondary antibody at room temperature
for 30 min prior to washing in PBS. Diaminobenzidine was used as
the chromogen. Sections were counterstained with Mayer's
hematoxylin and eosin.
Immunostaining densities were assessed by
pathologists using the ECLIPSE 80i light microscope and NIS
Elements D 3.2 imaging software (both from Nikon Corporation,
Tokyo, Japan) and the H-score method, as described previously
(12): H-score = (% tumor cells
unstained × 0) + (% tumor cells weakly stained × 1) + (% tumor
cells moderately stained × 2) + (% tumor cells strongly stained ×
3). H-scores ranged between 0 (100% negative tumor cells) and 300
(100% strongly positive tumor cells).
For AID, lymphocytes from germinal centers in
lymphoid follicles were used as internal positive controls because
they were primarily activated B cells and were strongly stained
throughout the specimens. H-scores were calculated for AID, NF-κB,
PAX5 and c-Myb for each sample. The following threshold values were
adopted: 105 for AID, 125 for NF-κB, 70 for PAX5 and 100 for
c-Myb.
In situ hybridization (ISH) of
EBER1
ISH was performed using a fluorescein
isothiocyanate-labeled probe complementary to an EBER1 sequence as
described previously (13). Sections
of FFPE tissue (thickness, 4 µm) were used for ISH and were imaged
using a light microscope.
Statistical analysis
Statistical analyses were performed using Fisher's
exact test, χ2 test, Mann-Whitney U test and
Kaplan-Meier survival estimates using SPSS software (version
23.0.0.0; IBM SPSS, Armonk, NY, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Clinicopathological findings
Representative histology, ISH and immunostaining
images of EBV(+) GCLS, EBV(−) GC without LS and EBV(−) GCLS are
presented in Fig. 1.
Clinicopathological data are presented in Tables I–III.
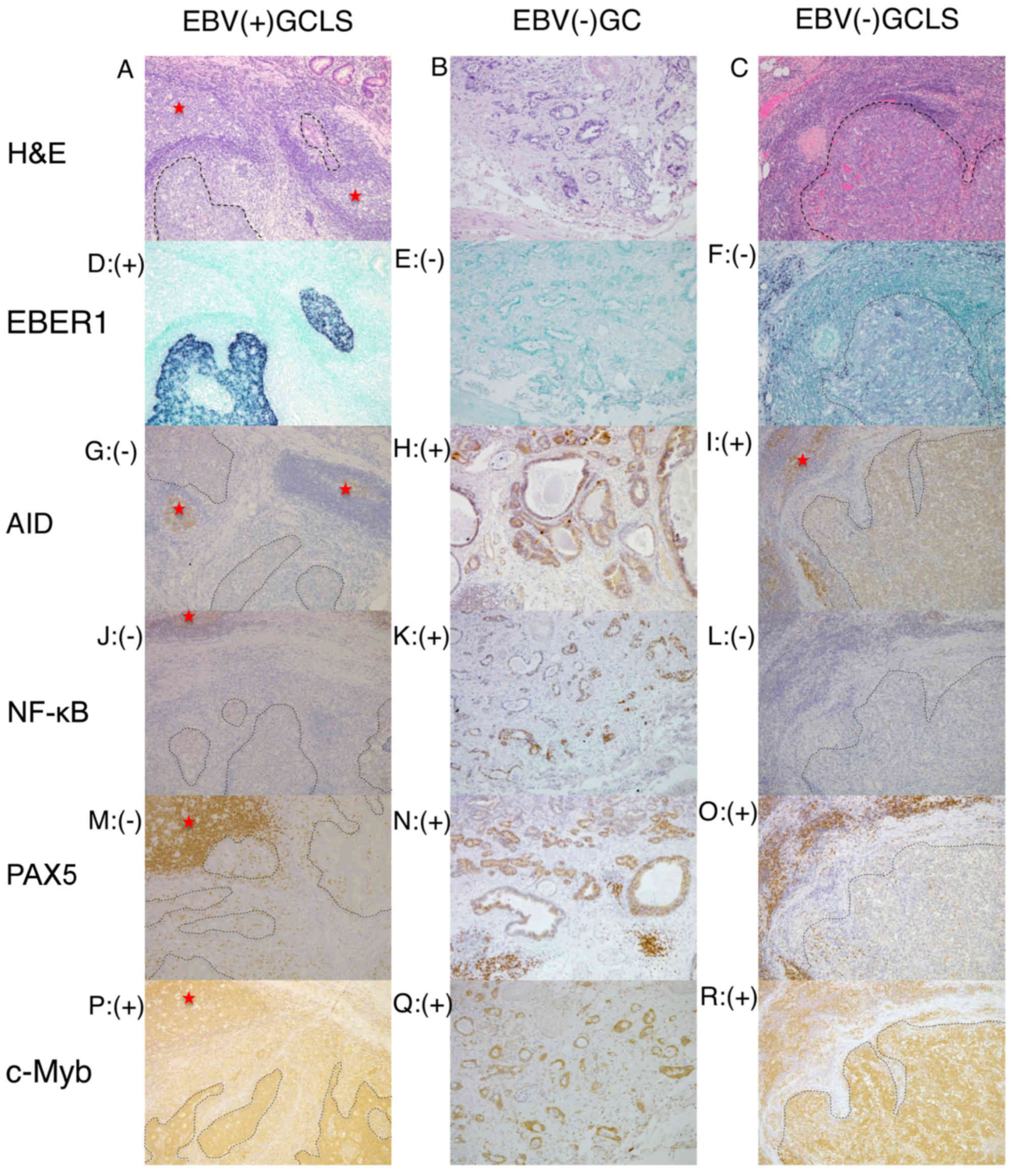 | Figure 1.Representative histology, ISH and
immunostaining images of EBV(+) GCLS, EBV(−) GC without LS and
EBV(−) GCLS. H&E staining of (A) EBV(+) GCLS, (B) EBV(−) GC
without LS and (C) EBV(−) GCLS. EBER1 ISH of (D) EBV(+) GCLS, (E)
EBV(−) GC without LS and (F) EBV(−) GCLS. AID immunostaining of (G)
EBV(+) GCLS, (H) EBV(−) GC without LS and (I) EBV(−) GCLS. NF-κB
immunostaining of (J) EBV(+) GCLS, (K) EBV(−) GC without LS and (L)
EBV(−) GCLS. PAX5 immunostaining of (M) EBV(+) GCLS, (N) EBV(−) GC
without LS and (O) EBV(−) GCLS. c-Myb immunostaining of (P) EBV(+)
GCLS, (Q) EBV(−) GC without LS and (R) EBV(−) GCLS. Magnification,
×100. (G, I) Red stars indicate AID expression as the positive
internal control observed in the lymphoid cells of the germinal
center. ISH, in situ hybridization; EBV, Epstein-Barr virus;
GCLS, GC with LS; GC, gastric carcinoma; LS, lymphoid stroma;
H&E, hematoxylin and eosin; EBER1, EBV-encoded small RNA 1;
AID, activation-induced cytidine deaminase; NF-κB, nuclear factor
κB; PAX5, paired box 5; c-Myb, c-Myb proto-oncogene. |
 | Table I.Comparative analysis of clinical and
histopathological characteristics between EBV(+) and EBV(−) GC. |
Table I.
Comparative analysis of clinical and
histopathological characteristics between EBV(+) and EBV(−) GC.
| Case | Sex | Age | T | ly | v | N | M | Stage | Histo-pathological
classification | Location | State | Survival time,
days | EBER1 | EBV-LMP-1 | AID H-score | NF-κB H-score | PAX5 H-score | c-Myb H-score | Mean USSa (Helico-bacter pylori) |
|---|
| EBV(+) GC |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 | Male | 48 | 1b | 1 | 0 | 0 | 0 | IA | GCLS | UT | NED | 3,469 | + | − | 20 | 190 |
0 | 70 | 1.6 (−) |
|
2 | Male | 68 | 3 | 1 | 1 | 0 | 0 | IIA | GCLS | UT | NED | 1,350 | + | − | 100 | 19 |
0 | 80 | 1.8 (+) |
|
3 | Male | 52 | 2 | 1 | 3 | 0 | 0 | IB | GCLS | MT | NED | 1,525 | + | − | 70 | 70 |
0 | 10 | 2.2 (+) |
|
4 | Male | 53 | 3 | 1 | 3 | 1 | 0 | IIB | GCLS | MT | NED | 1,839 | + | − |
3 | 15 | 65 | 190 | 2.2 (+) |
|
5 | Male | 74 | 1b | 1 | 0 | 1 | 0 | IB | GCLS | UT | NED | 1,512 | + | − |
5 | 15 |
0 | 40 | 1.8 (+) |
|
6 | Male | 63 | 1b | 0 | 0 | 0 | 0 | IA | GCLS | UT | NED | 845 | + | − | 80 | 80 |
0 | 70 | 2.4 (+) |
|
7 | Male | 77 | 1b | 0 | 0 | 0 | 0 | IA | GCLS | MT | NED | 700 | + | − | 90 | 90 | 35 | 225 | 2.0 (−) |
|
8 | Male | 54 | 1b | 2 | 1 | 0 | 0 | IA | GCLS | MT | NED | 675 | + | − | 30 | 75 | 60 | 110 | 1.4 (+) |
|
9 | Male | 45 | 1b | 0 | 0 | 0 | 0 | IA | GCLS | UT | NED | 657 | + | − | 90 | 85 |
0 | 160 | 1.4 (+) |
| 10 | Male | 60 | 1b | 0 | 0 | 0 | 0 | IA | GCLS | MT | NED | 391 | + | − | 30 | 24 |
0 | 60 | 0.2 (−) |
| 11 | Male | 58 | 3 | 2 | 2 | 2 | 0 | IIIA | por2>tub2 | LT | NED | 706 | + | − | 53 | 120 | 110 | 220 | 1.4 (−) |
| EBV(−) GC |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| 12 | Male | 77 | 2 | 1 | 1 | 0 | 0 | IB | GCLS | LT | NED | 1,115 | − | − | 60 |
8 | 140 | 70 | 1.8 (−) |
| 13 | Male | 73 | 3 | 2 | 3 | 2 | 0 | IIIA | GCLS | R | STD | 2,250 | − | − | 170 | 10 | 60 | 210 | 1.6 (−) |
| 14 | Male | 73 | 1b | 0 | 0 | 0 | 0 | IA | GCLS | MT | NED | 773 | − | − | 120 | 50 |
0 | 260 | 2.0 (+) |
| 15 | Female | 68 | 2 | 1 | 2 | 0 | 0 | IB | GCLS | LT/MT | NED | 2,161 | − | − | 10 | 55 | 105 | 125 | 2.0 (+) |
| 16 | Male | 76 | 1b | 1 | 0 | 0 | 0 | IA | GCLS | R | NED | 2,943 | − | − | 110 | 17 |
0 | 90 | 2.0 (+) |
| 17 | Male | 77 | 1b | 2 | 2 | 0 | 0 | IA | tub2>tub1 | MT | SAC | 127 | − | − |
9 | 140 | 150 | 40 | 2.0 (+) |
| 18 | Male | 71 | 2 | 1 | 1 | 0 | 0 | IB | sig>por2 | MT | NED | 2,876 | − | − |
3 | 140 |
0 | 20 | 2.0 (+) |
| 19 | Male | 69 | 2 | 2 | 1 | 0 | 0 | IB | tub1>
tub2>muc | UT | NED | 187 | − | − | 135 | 240 | 100 | 16 | 2.4 (+) |
| 20 | Male | 66 | 1b | 1 | 1 | 0 | 0 | IA | tub>por2 | MT | NED | 2,610 | − | − | 30 | 295 | 120 | 80 | 1.6 (+) |
| 21 | Female | 58 | 1b | 2 | 2 | 0 | 0 | IA | tub2 | MT | NED | 2,511 | − | − | 70 | 280 | 120 | 95 | 2.2 (+) |
| 22 | Male | 57 | 2 | 1 | 1 | 0 | 0 | IB | sig | MT | NED | 2,530 | − | − | 150 | 110 |
0 | 60 | 1.2 (+) |
| 23 | Male | 49 | 1b | 1 | 1 | 0 | 0 | IA | por2>tub2 | UT | NED | 1,078 | − | − | 45 | 295 | 100 | 95 | 2.4 (+) |
| 24 | Male | 59 | 1b | 1 | 0 | 0 | 0 | IA | tub1>tub2 | MT | NED | 2,493 | − | − | 30 | 255 | 70 | 24 | 2.2 (+) |
| 25 | Male | 67 | 1b | 1 | 0 | 0 | 0 | IA | por2>sig | MT | NED | 2,435 | − | − | 105 | 140 | 10 | 75 | 1.8 (+) |
| 26 | Male | 69 | 1b | 1 | 1 | 0 | 0 | IA | tub1>tub2 | UT | NED | 2,211 | − | − | 90 | 230 | 100 | 180 | 2.0 (+) |
| 27 | Male | 77 | 1b | 1 | 3 | 0 | 0 | IA | tub2 | R | NED | 1,634 | − | − | 80 | 125 | 90 | 170 | 0.8 (−) |
| 28 | Male | 70 | 1b | 2 | 2 | 1 | 0 | IB | tub2 | UT | SAC | 403 | − | − | 70 | 180 | 60 | 170 | 1.8 (+) |
| 29 | Male | 63 | 3 | 1 | 1 | 0 | 0 | IIA | tub1 | MT | NED | 1,520 | − | − | 35 | 150 | 100 | 75 | 1.8 (+) |
| 30 | Male | 70 | 3 | 2 | 3 | 1 | 0 | IIB | tub2 | MT | NED | 898 | − | − | 145 | 40 |
0 |
5 | 2.2 (+) |
| Table III.Association between EBV infection and
expression of AID and AID-regulatory factors in GC with or without
LS. |
Table III.
Association between EBV infection and
expression of AID and AID-regulatory factors in GC with or without
LS.
| GC type | AID(+) % | P-value | NF-κB (+) (AID
activator) (%) | P-value | PAX5 (+) (AID
activator) (%) | P-value | c-Myb (+) (AID
silencer) (%) | P-value |
|---|
|
|
| 0.025a |
| 0.005a |
| 0.01a |
| 0.354 |
| EBV(+) GC
(n=11) | 0 (0) |
| 1 (9) |
| 1 (9) |
| 5 (45) |
|
| EBV(−) GC
(n=19) | 7
(37) |
| 12 (63) |
| 11 (58) |
| 6 (32) |
|
|
|
| NA |
| NA |
| NA |
| NA |
| EBV(+) LS(+) GC
(n=10) | 0 (0) |
| 1 (10) |
| 0 (0) |
| 4 (40) |
|
| EBV(−) LS(+) GC
(n=5) | 3
(60) |
| 0 (0) |
| 2 (40) |
| 3 (60) |
|
|
|
| 0.500 |
|
<0.001a |
| 0.003a |
| 0.256 |
| LS(+) GC
(n=15) | 3 (20) |
| 1 (7) |
| 2
(13) |
| 7 (47) |
|
| LS(−) GC
(n=15) | 4 (27) |
| 12 (80) |
| 10 (67) |
| 4 (27) |
|
Detection of EBV markers in tissue
samples suspected of EBVaGC
The expression of EBER1 was only detected in 10/15
GCLS samples, the majority of which were histopathologically
suspected of EBVaGC. On the other hand, 1/15 GC without LS samples
tested positive for EBER1. All 30 samples were negative for LMP-1
(data not shown).
Grading of atrophic gastritis in
sample tissue backgrounds
Background atrophic gastritis was evaluated and
scored in sample tissues according to the USS. The mean USS scores
(± SD) in EBV (+)GC (n=11) and in EBV (−)GC (n=19) were 1.67
(±0.57) and 1.88 (±0.38), respectively, and were not significantly
different (P=0.328).
EBV(−) GC is more likely to express
AID and AID-regulatory factors
AID H-scores of ≥105 were considered to be positive.
Similarly, NF-κB, PAX5 and c-Myb H-scores of ≥125, ≥70 and ≥100,
respectively, were considered to be positive.
The positive H-score rates of AID and its regulatory
factors were compared between EBV(+) GC (n=11) and EBV(−) GC
(n=19). The rates were 0/11 vs. 7/19 (0 vs. 37%; P=0.025) for AID,
1/11 vs. 12/19 (9 vs. 63%; P=0.005) for NF-κB, 1/11 vs. 11/19 (9
vs. 58%; P=0.01) for PAX5 and 5/11 vs. 6/19 (45 vs. 32%; P=0.35)
for c-Myb for EBV(+) GC and EBV(−) GC, respectively. The positive
rates of AID, NF-κB and PAX5, but not c-Myb, were significantly
increased in EBV(−) GC compared with in EBV(+) GC (Table III).
No association between the expression of AID and USS
scores was identified (P=0.623; Table
II).
| Table II.Association between expression of AID
and AID-regulatory factors and clinicopathological parameters in
gastric cancers (n=30) regardless of EBV infection. |
Table II.
Association between expression of AID
and AID-regulatory factors and clinicopathological parameters in
gastric cancers (n=30) regardless of EBV infection.
| Factor | AID (+) | P-value | NF-κB (+) (AID
activator) | P-value | PAX-5 (+) (AID
activator) | P-value | c-Myb (+) (AID
silencer) | P-value |
|---|
| T |
| 0.572 |
| 0.215 |
| 0.831 |
| 0.465 |
|---|
| 1b
(n=18) | 3 |
| 10 |
| 7 |
| 7 |
|
| 2
(n=6) | 2 |
| 2 |
| 3 |
| 1 |
|
| 3
(n=6) | 2 |
| 1 |
| 2 |
| 3 |
|
| ly |
| 0.539 |
| 0.100 |
| 0.134 |
| 0.218 |
| 0
(n=5) | 1 |
| 0 |
| 0 |
| 3 |
|
| 1
(n=17) | 3 |
| 9 |
| 8 |
| 4 |
|
| 2
(n=8) | 3 |
| 4 |
| 4 |
| 4 |
|
| v |
| 0.456 |
| 0.317 |
| 0.022a |
| 0.292 |
| 0
(n=10) | 3 |
| 3 |
| 1 |
| 3 |
|
| 1
(n=10) | 2 |
| 6 |
| 6 |
| 2 |
|
| 2
(n=5) | 0 |
| 3 |
| 4 |
| 3 |
|
| 3
(n=5) | 2 |
| 1 |
| 1 |
| 3 |
|
| N |
| 0.642 |
| 0.285 |
| 0.213 |
| 0.114 |
| 0
(n=24) | 5 |
| 12 |
| 11 |
| 7 |
|
| 1
(n=4) | 1 |
| 1 |
| 0 |
| 2 |
|
| 2
(n=2) | 1 |
| 0 |
| 1 |
| 2 |
|
| Stage |
| 0.663 |
| 0.368 |
| 0.806 |
| 0.264 |
| IA
(n=16) | 3 |
| 9 |
| 7 |
| 6 |
|
| IB
(n=8) | 2 |
| 3 |
| 3 |
| 2 |
|
| IIA
(n=2) | 0 |
| 1 |
| 1 |
| 0 |
|
| IIB
(n=2) | 1 |
| 0 |
| 0 |
| 1 |
|
| IIIA
(n=2) | 1 |
| 0 |
| 1 |
| 2 |
|
| Location |
| 0.179 |
| 0.387 |
| 0.172 |
| 0.389 |
| Upper
third (n=9) | 1 |
| 5 |
| 3 |
| 3 |
|
| Middle
third (n=15) | 4 |
| 7 |
| 5 |
| 4 |
|
| Lower
third (n=3) | 0 |
| 0 |
| 3 |
| 2 |
|
| Remnant
GC (n=3) | 2 |
| 1 |
| 1 |
| 2 |
|
| Age, years |
| 0.089 |
| 0.462 |
| 0.590 |
| 0.421 |
| <65
(n=13) | 1 |
| 5 |
| 5 |
| 4 |
|
| ≥65
(n=17) | 6 |
| 8 |
| 7 |
| 7 |
|
| Mean USS |
| 0.623 |
| 0.515 |
| 0.073 |
| 0.310 |
| ≥2.2
(n=22) | 5 |
| 9 |
| 11 |
| 7 |
|
| <2.2
(n=8) | 2 |
| 4 |
| 1 |
| 4 |
|
Certain cases of EBV(−) GCLS express
AID and PAX5 (AID activator)
H-scores of AID and its regulatory factors were
compared between EBV(+) GCLS (n=10) and EBV(−) GCLS (n=5). The
positive H-score rate of AID and PAX5 was increased in EBV(−) GCLS
(3/5, 60% for AID and 2/5, 40% for PAX5) compared with in EBV(+)
GCLS (both 0/10, 0%). However, no significant differences were
identified for AID or AID-regulatory factors between EBV(+) GCLS
and EBV(−) GCLS, owing to an insufficient number of cases examined
(Table III).
Expression of NF-κB and PAX5 is
decreased in GCLS compared with in GC without LS
The aberrant expression of NF-κB and PAX5 was
significantly decreased in GCLS [1/15 (7%) and 2/15 (13%);
P<0.001] compared with in GC without LS [12/15 (80%) and 10/15
(67%); P=0.003] (Table III).
AID and AID-regulatory factors except
PAX5 are not associated with clinicopathological parameters
The relationships of the expression of AID and its
regulatory factors with clinicopathological parameters including
tumor category, lymphatic invasion, venous invasion, node category,
stage, location of GC, age and USS were investigated. No
association between AID and its regulatory factors except PAX5 and
the clinical parameters examined in EBV(+) and EBV(−) GCs was
identified (n=30), whereas PAX5 expression was significantly
associated with venous invasion (P=0.022; Table II).
Association between AID and
AID-regulatory factors and survival rates
Among 30 patients who participated in the present
study, only one succumbed to GC (Table
I, case 13). Therefore, in the survival analysis, it was not
possible to conclude whether AID and AID-regulatory factors
affected the survival rate.
Discussion
Previous comprehensive genome analyses using
next-generation sequencing of various cancer tissues have revealed
that cancer cells have numerous nucleotide alterations (14). Cancer cells are considered to be
generated from the stepwise accumulation of genetic alterations in
various genes in inflammation-associated carcinogenesis (7). AID, a nucleotide-editing enzyme that is
essential for somatic hypermutation and class-switch recombination
of the immunoglobulin gene, is also known to serve a role as a
genomic mutator in carcinogenesis (9). Shimizu et al (7) demonstrated that pathogenic bacterial or
viral factors and subsequent inflammatory reactions in H.
pylori-related gastritis, chronic viral hepatitis, Barrett's
esophagus and inflammatory bowel disease lead to the aberrant
expression of AID in various epithelial cells via NF-κB activation,
which causes the accumulation of genetic alterations in
tumor-related genes. The aberrant expression of AID was observed in
the gastric epithelium and gastric cancer cells with an H.
pylori-infected background (10,15,16).
EBVaGC, a distinct type of GC, the majority of cases of which have
lymphoid stroma, is an EBV-infected and inflammation-associated
cancer. The results of the present study identified that the
aberrant expression of AID and NF-κB was significantly decreased in
EBVaGC compared with in EBV-non-associated GC, and also that EBV
was not likely to be involved in pathogen-induced AID expression
via NF-κB activation in gastric cancers. These results suggested
that the genomic modulation of tumor-related genes by EBV-induced
AID expression is not a primary causative mechanism in the
carcinogenesis of EBVaGC. Unexpectedly, the aberrant expression of
NF-κB was less frequent in EBVaGC with LS or GCLS compared with in
GC without LS. A potential reason for the decreased expression of
NF-κB is that EBV-LMP-1, a strong activator of NF-κB, was not
expressed in EBVaGC with latency type I or II EBV infection.
However, this does not explain the decreased expression of NF-κB in
GCLS compared with in GC without LS regardless of EBV infection
because LS is a hallmark of chronic inflammation. Furthermore, no
significant differences were identified in the expression of AID
between GCLS and GC without LS. These results suggest that LS, a
histological characteristic of chronic inflammation, does not
necessarily reflect NF-κB-inducing conditions, and that NF-κB and
homeobox protein C4 may promote the transcription of AID (8). The activation of NF-κB is a complex
process induced by various stimuli, including microbial and viral
products, cytokines, DNA damage, oxidative damage and radiation
(17). Aberrant NF-κB activity has
been demonstrated to contribute to a number of human diseases that
possess an immune or inflammatory component, including
tumorigenesis and H. pylori infection (17). Other stimuli, including H.
pylori infection, appear to be more important in the induction
of aberrant AID expression via NF-κB in the stomach compared with
EBV infection (15). In the present
study, no significant differences were observed in mean USS scores
or H. pylori-infected rates between EBVaGC and
EBV-non-associated GC. No association was identified between EBV
infection and H. pylori infection in gastric cancer patients
from northern Brazil (18).
Furthermore, no association between the expression of AID and USS
scores was identified; however, H. pylori infection is
generally considered to induce atrophic gastritis and intestinal
metaplasia with the aberrant expression of AID, which subsequently
leads to gastric cancers through the accumulation of genetic
mutations (19).
PAX5, known as a B cell-specific activator protein,
serves a role as an activator for AID (9). In the present study, the aberrant
expression of PAX5 and AID was more frequently observed in
EBV-non-associated GC compared with in EBVaGC (P=0.01 and P=0.025,
respectively). The expression of PAX5 was also increased in GC
without LS compared with in GCLS (P=0.003). This was attributed to
the aberrant expression of PAX5 contributing to the induction of
AID expression in EBV-non-associated GC, the majority of cases of
which exhibited no LS background. PAX5 has been identified
as a tumor suppressor gene and the epigenetic inactivation of
PAX5 through the direct up-regulation of p53 is associated
with a poor prognosis in gastric cancer patients (20). However, it is noteworthy that PAX5
expression was significantly associated with venous invasion of GC
in that study.
The cellular oncogene c-Myb, a v-Myb
avian myeloblastosis viral oncogene homolog, has been implicated in
leukemogenesis, and c-Myb was demonstrated to be increased in acute
myeloid leukemia and acute lymphoblastic leukemia samples (21). c-Myb also serves a role as a repressor
or silencer of the expression of AID (8). The aberrant expression of c-Myb in the
present study was not significantly different between EBVaGC and
EBV-non-associated GC, or between GCLS and GC without LS. The
expression of c-Myb as a silencer for the induction of AID appears
to serve certain roles in the expression of AID in gastric cancers,
regardless of the EBV status.
The Cancer Genome Atlas Research Network recently
provided a comprehensive molecular characterization of gastric
adenocarcinoma and defined EBVaGC as a specific subtype, which
displays recurrent mutations in phosphoinositide 3-kinase catalytic
α polypeptide (PIK3CA) and AT-rich-interaction domain 1A
(ARID1A), extreme DNA hypermethylation, and the
amplification of genes encoding c-Jun-associated kinase 2, cluster
of differentiation 274 (also known as programmed death ligand 1)
and programmed cell death 1 ligand 2 (also known as programmed
death ligand 2) with the extremely rare tumor protein 53 gene
mutation (22). Furthermore,
mutations in PIK3CA and ARID1A may precede EBV
infection because mutation events have been detected in chronically
inflamed and H. pylori-infected gastric mucosa (23). EBVaGC is characterized by extreme DNA
hypermethylation and relatively rare somatic gene mutations. The
decreased aberrant expression of AID observed in EBVaGCs in the
present study is consistent with the relatively small number of
somatic gene mutations detected using the comprehensive molecular
analysis in this specific subtype of GC.
The prognostic effects of EBV positivity on gastric
cancer are controversial (23).
However, a recent meta-analysis of 4599 GC cases identified a
decreased mortality rate in EBVaGC following adjustment for the
cancer stage and other factors (24).
The 10-year disease-specific rate of patients with EBVaGCLS (89.1%)
was previously demonstrated to be significantly increased compared
with that of EBV-non-associated GCLS (66.9%) (P=0.009) (4). These results suggest that the EBV
infection status in GC is an important prognostic factor. The
aberrant expression of AID and AID-regulatory factors (NF-κB and
c-Myb) in the present study was not identified to be associated
with clinicopathological parameters, whereas only PAX5 expression
was significantly associated with venous invasion. The relevance of
AID and its regulatory factors for the survival rate of patients
with GC was not demonstrated in the present study because only 1/30
patients succumbed to GC during the observation period.
In inflammation-associated cancer development, in
addition to infectious agents including H. pylori, a number
of intrinsic mediators of inflammation, including proinflammatory
cytokines, growth factors, and reactive oxygen and nitrogen
species, have the ability to induce the genetic and epigenetic
modulation of tumor-related genes through various mechanisms,
including enhancements in cell growth, mobility and angiogenesis,
the inhibition of apoptosis and somatic gene mutations or
rearrangements, and DNA methylation (6,7).
Furthermore, inflammation modulates the expression of microRNAs
that influence the production of numerous tumor-related mRNAs or
proteins (5). On the other hand, in
the development of EBVaGC, mechanisms other than the AID-induced
genetic mutations described above, particularly DNA methylation,
may have more important functions as the mechanisms underlying
somatic gene mutations through EBV-induced aberrant AID expression
were not demonstrated in EBVaGC in the present study.
In the present study, significant aberrant
expression of AID was not observed in EBVaGC and is considered
irrelevant to the mechanisms underlying carcinogenesis in this
specific subtype of GC, whereas aberrant AID expression contributes
to carcinogenesis in certain cases of EBV-non-associated GC.
Acknowledgements
The authors thank former Professor Masahide Ikeguchi
of the First Surgery Department, Tottori University Hospital
(Yonago, Japan) for providing the clinical samples.
References
|
1
|
Jha HC, Banerjee S and Robertson ES: The
role of Gammaherpesviruses in cancer pathogenesis. Pathogens.
5(pii): E182016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Epstein MA, Achong BG and Barr YM: Virus
particle in cultured lymphoblasts from Burkitt's Lymphoma. Lancet.
1:702–703. 1964. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shinozaki-Ushiku A, Kunita A and Fukayama
M: Update on Epstein-Barr virus and gastric cancer (review). Int J
Oncol. 46:1421–1434. 2015.PubMed/NCBI
|
|
4
|
Lim H, Park YS, Lee JH, Son da H, Ahn JY,
Choi KS, Kim do H, Choi KD, Song HJ, Lee GH, et al: Features of
gastric carcinoma with lymphoid stroma associated with Epstein-Barr
virus. Clin Gastroenterol Hepatol. 13:1738–1744.e2. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chiba T, Marusawa H and Ushijima T:
Inflammation-associated cancer development in digestive organs:
Mechanisms and roles for genetic and epigenetic modulation.
Gastroenterology. 143:550–563. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marusawa H, Takai A and Chiba T: Role of
activation-induced cytidine deaminase in inflammation-associated
cancer development. Adv Immunol. 111:109–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shimizu T, Marusawa H, Endo Y and Chiba T:
Inflammation-mediated genomic instability: Roles of
activation-induced cytidine deaminase in carcinogenesis. Cancer
Sci. 103:1201–1206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zan H and Casali P: Regulation of Aicda
expression and AID activity. Autoimmunity. 46:83–101. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Honjo T, Kobayashi M, Begum N, Kotani A,
Sabouri S and Nagaoka H: The AID dilemma: Infection, or cancer? Adv
Cancer Res. 113:1–44. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kawata S, Yashima K, Yamamoto S, Sasaki S,
Takeda Y, Hayashi A, Matsumoto K, Kawaguchi K, Harada K and
Murawaki Y: AID, p53 and MLH1 expression in early gastric neoplasms
and the correlation with the background mucosa. Oncol Lett.
10:737–743. 2015.PubMed/NCBI
|
|
11
|
Dixon MF, Genta RM, Yardley JH and Correa
P: Classification and grading of gastritis. The updated Sydney
System. International Workshop on the Histopathology of Gastritis,
Houston 1994. Am J Surg Pathol. 20:1161–1181. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iwasaki T, Matsushita M, Nonaka D, Nagata
K, Kato M, Kuwamoto S, Murakami I and Hayashi K: Lower expression
of CADM1 and higher expression of MAL in Merkel cell carcinomas are
associated with Merkel cell polyomavirus infection and better
prognosis. Hum Pathol. 48:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takashima K, Ohashi M, Kitamura Y, Ando K,
Nagashima K, Sugihara H, Okuno K, Sairenji T and Hayashi K: A new
animal model for primary and persistent Epstein-Barr virus
infection: Human EBV-infected rabbit characteristics determined
using sequential imaging and pathological analysis. J Med Virol.
80:455–466. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meyerson M, Gabriel S and Getz G: Advances
in understanding cancer genomes through second-generation
sequencing. Nat Rev Genet. 11:685–696. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsumoto Y, Marusawa H, Kinoshita K, Endo
Y, Kou T, Morisawa T, Azuma T, Okazaki IM, Honjo T and Chiba T:
Helicobacter pylori infection triggers aberrant expression of
activation-induced cytidine deaminase in gastric epithelium. Nat
Med. 13:470–476. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goto A, Hirahashi M, Osada M, Nakamura K,
Yao T, Tsuneyoshi M, Takayanagi R and Oda Y: Aberrant
activation-induced cytidine deaminase expression is associated with
mucosal intestinalization in the early stage of gastric cancer.
Virchows Arch. 458:717–724. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Le Negrate G: Viral interference with
innate immunity by preventing NF-κB activity. Cell Microbiol.
14:168–181. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
de Souza CR, de Oliveira KS, Ferraz JJ,
Leal MF, Calcagno DQ, Seabra AD, Khayat AS, Montenegro RC, Alves
AP, Assumpção PP, et al: Occurrence of Helicobacter pylori and
Epstein-Barr virus infection in endoscopic and gastric cancer
patients from Northern Brazil. BMC Gastroenterol. 14:1792014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shimizu T, Marusawa H, Matsumoto Y,
Inuzuka T, Ikeda A, Fujii Y, Minamiguchi S, Miyamoto S, Kou T,
Sakai Y, et al: Accumulation of somatic mutations in TP53 in
gastric epithelium with Helicobacter pylori infection.
Gastroenterology. 147:407–417.e3. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li X, Cheung KF, Ma X, Tian L, Zhao J, Go
MY, Shen B, Cheng AS, Ying J, Tao Q, et al: Epigenetic inactivation
of paired box gene 5, a novel tumor suppressor gene, through direct
upregulation of p53 is associated with prognosis in gastric cancer
patients. Oncogene. 31:3419–3430. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shetzline SE, Rallapalli R, Dowd KJ, Zou
S, Nakata Y, Swider CR, Kalota A, Choi JK and Gewirtz AM:
Neuromedin U: A Myb-regulated autocrine growth factor for human
myeloid leukemias. Blood. 104:1833–1840. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular characterization of gastric adenocarcinoma.
Nature. 513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abe H, Kaneda A and Fukayama M:
Epstein-barr virus-associated gastric carcinoma: Use of host cell
machineries and somatic gene mutations. Pathobiology. 82:212–223.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Camargo MC, Kim WH, Chiaravalli AM, Kim
KM, Corvalan AH, Matsuo K, Yu J, Sung JJ, Herrera-Goepfert R,
Meneses-Gonzalez F, et al: Improved survival of gastric cancer with
tumour Epstein-Barr virus positivity: An international pooled
analysis. Gut. 63:236–243. 2014. View Article : Google Scholar : PubMed/NCBI
|














