Introduction
Hepatocellular carcinoma (HCC) occurs in the liver
and is related to infection by hepatitis B or C virus (1). HCC cells maintain their proliferative
potential in fibrous tissues with limited amounts of glucose and
arginine (2), despite glucose being
indispensable for cell survival (3).
Hepatocytes produce energy from galactose via glycolysis (4,5) and
arginine is an essential amino acid in this process (6). Hepatocytes produce arginine from
ornithine via the urea cycle. Hepatocyte selection medium (HSM) is
deficient in glucose and arginine, and is supplemented with
galactose and ornithine (7). HSM
enriches hepatocytes from co-culture with human induced pluripotent
stem cells (8). HSM is also
applicable to a condition similar to fibrous tissues (9).
α-fetoprotein (AFP) is a marker of immature
hepatocytes (10). HCC cells are
immature hepatocytes and produce AFP (11,12). CD44
is a receptor of hyaluronan (13) and
is used as a marker of cancer stem cells (14). In a previous study, CD44 was not
identified to be expressed in surgical specimens of HCC because the
presence of cancer stem cells is rare (15). In another previous study, HCC cells
were demonstrated to be heterogeneously positive for AFP and CD44
(16). These studies suggest that the
positivity of CD44 in HCC cells remains controversial. Alterations
in the expression patterns of AFP and CD44 are not known in HCC
cells cultured without glucose or arginine.
Therefore, in the present study, the expression
patterns of AFP and CD44 were investigated in HCC cells cultured in
HSM to examine the alterations in the characteristics of HCC cells
grown in the absence of glucose and arginine.
Materials and methods
Cell culture and determination of cell
numbers
The HCC cell lines HLF and PLC/PRF/5 cells (RIKEN
Cell Bank, Tsukuba, Japan) were cultured in Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10% fetal bovine serum
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) in 6-well plates
(AGC Techno Glass Co., Ltd., Shizouka, Japan) at 37°C in a
humidified chamber containing 5% CO2. Cells
(1×104) were spread onto each well of the 6-well plates,
and viable cell numbers were determined on days 0 and 7 following
culture using the trypan blue dye exclusion test.
Hepatocyte selection medium
HSM was prepared from amino acid powders, using the
formulation of Leibovitz L-15 medium (Thermo Fisher Scientific,
Inc.), omitting arginine, tyrosine, glucose and sodium pyruvate,
but with the addition of 900 mg/l galactose, 1 mM ornithine, 5 mM
glycerol and 260 mM proline (all Wako Pure Chemical Industries,
Osaka, Japan). Proline was included in the medium as it is required
for DNA synthesis (17). Knockout
serum replacement (Thermo Fisher Scientific, Inc.) was used in
place of FBS to establish defined xenobiotic-free conditions and
was added at a final concentration of 10%.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cells were cultured in DMEM or HSM in 6-well plates.
On days 0, 4 and 7, RNA was isolated using Isogen (Nippon Gene Co.,
Ltd., Tokyo, Japan). Total RNA (5 µg) was subjected to cDNA
synthesis using a SuperScript III First-Strand Synthesis system
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. qPCR was performed using Fast SYBR Green Master Mix
(Thermo Fisher Scientific, Inc.) with the MiniOpticon (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) for 40 cycles consisted of
two steps of denaturation at 95°C for 5 sec and annealing-extension
at 60°C for 5 sec. Table I lists the
primer sequences used. Ribosomal protein L19 (RPL19), a
constitutively expressed housekeeping gene, was used as an internal
control (18). RPL19 was used as an
endogenous control to monitor the amount of mRNA because the gene
is a constitutively expressed housekeeping gene (18). The expression levels of the genes were
analyzed automatically by the Mini Opticon system (Bio-Rad
Laboratories, Inc.) based on delta-delta cycle quantification
(ΔΔCq) method (19). Relative
expression level of a gene was calculated as expression level of a
gene divided by expression level of RPL19.
 | Table I.Sequences of primers used in the
quantitative polymerase chain reaction. |
Table I.
Sequences of primers used in the
quantitative polymerase chain reaction.
| Primer name | Sequence | Description | Product size
(bp) | GenBank®
accession no. |
|---|
| OMC317 |
5-ACACAAAAAGCCCACTCCAG-3′ | AFP, forward | 147 | NM_001134 |
| OMC318 |
5′-GGTGCATACAGGAAGGGATG-3′ | AFP, reverse | 147 |
|
| OMC321 |
5′-CGAATGCCAGAGAAGGTCAC-3′ | RPL19, forward | 157 | BC095445 |
| OMC322 |
5′-CCATGAGAATCCGCTTGTTT-3′ | RPL19, reverse | 157 |
|
| OMC755 |
5′-AGAAGGTGTGGGCAGAAGAA-3′ | CD44, forward | 116 | BC004372 |
| OMC756 |
5′-AAATGCACCATTTCCTGAGA-3′ | CD44, reverse |
|
|
Immunostaining
HLF and PLC/PRF/5 cells were spread onto 8-well
chamber slides (Matsunami Glass Ind., Ltd., Kishiwada, Japan). The
cells were cultured in DMEM or HSM. Following 7 days of culture,
the cells were fixed with 4% paraformaldehyde (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) for 30 min at 4°C. Endogenous peroxidase
was inactivated by incubation with 0.1% hydrogen peroxide in 100%
methanol for 30 min at 4°C. Specimens were incubated with 2% fetal
bovine serum in PBS (wash buffer) at 4°C for 30 min. Specimens were
incubated with mouse anti-human AFP (catalog no. M225) (Takara Bio,
Inc., Otsu, Japan) or mouse anti-human CD44 (catalog no. 3570s;
Cell Signaling Technology, Inc., Danvers, MA, USA) antibody diluted
1:1,000 in wash buffer overnight at 4°C. Specimens were rinsed with
PBS three times, and incubated with a 1:1,000 dilution of
horseradish peroxidase-labeled anti-mouse antibody (catalog no.
NA934-100UL; GE Healthcare Bio-Sciences, Pittsburg, PA, USA) at 4°C
for 3 h. Diaminobenzidine (Dako; Agilent Technologies, Inc., Santa
Clara, CA, USA) was applied, and the nuclei were stained with
hematoxylin (Muto Pure Chemicals Co., Ltd., Tokyo, Japan) for 15
sec. Specimens were observed and images were captured using an AX80
photomicroscope (Olympus Corporation, Tokyo, Japan).
Statistical analysis
A one-way analysis of variance was applied for
statistical analysis using JMP software (version 10.0.2; SAS
Institute, Cary, NC, Inc.), and Tukey's test was used as a post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
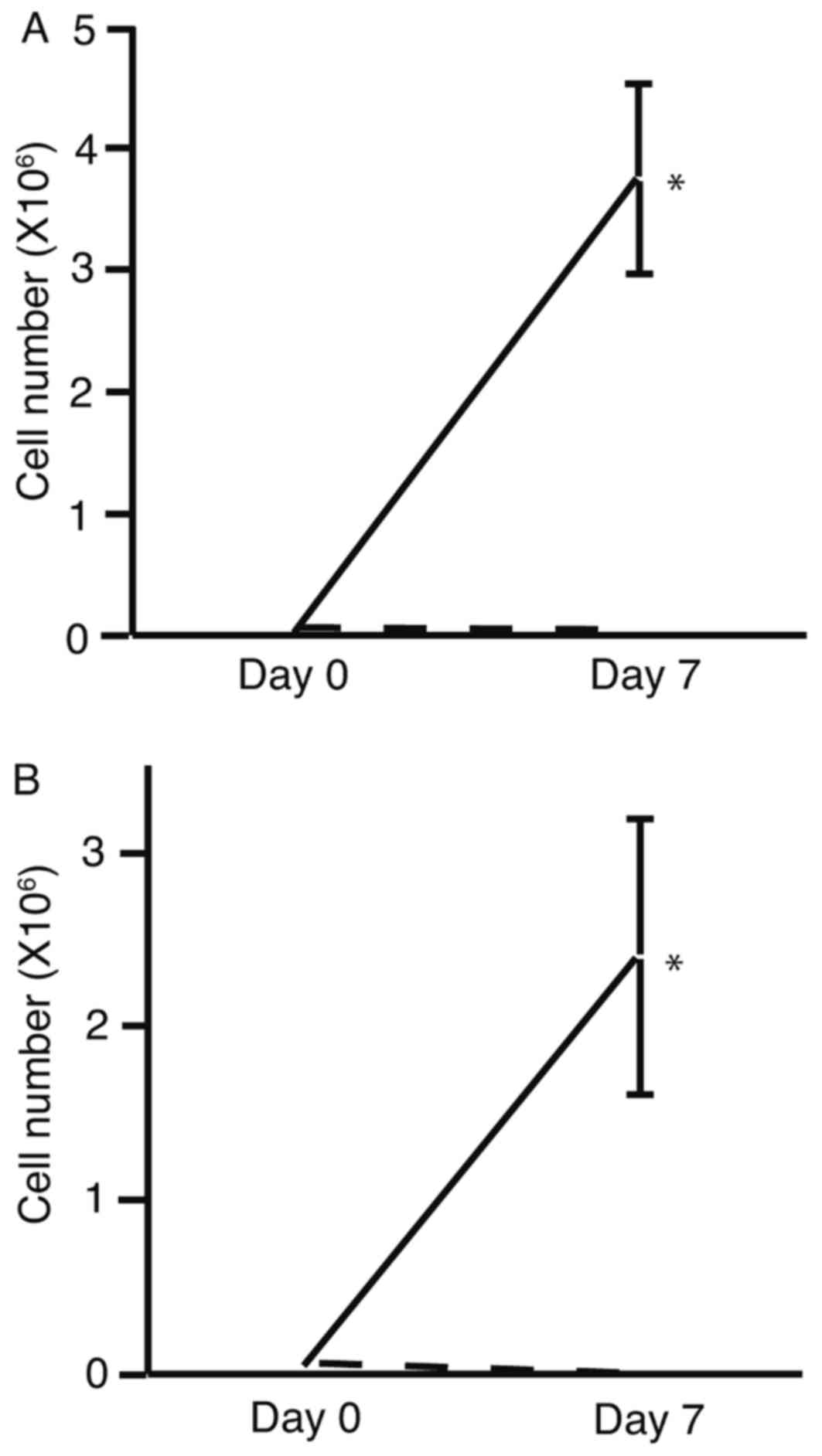
To determine the effects of HSM on cell
proliferation, the number of HLF (Fig.
1A) and PLC/PRF/5 (Fig. 1B)
viable cells were determined following culture in DMEM or HSM.
Viable cell numbers were observed to increase towards day 7 of
culture in DMEM, whereas a decrease in viable cell number was
observed towards day 7 of culture in HSM. Viable cell numbers were
significantly increased following culture in DMEM compared with
culture in HSM. These results suggest that HMS significantly
decreased cell proliferation.
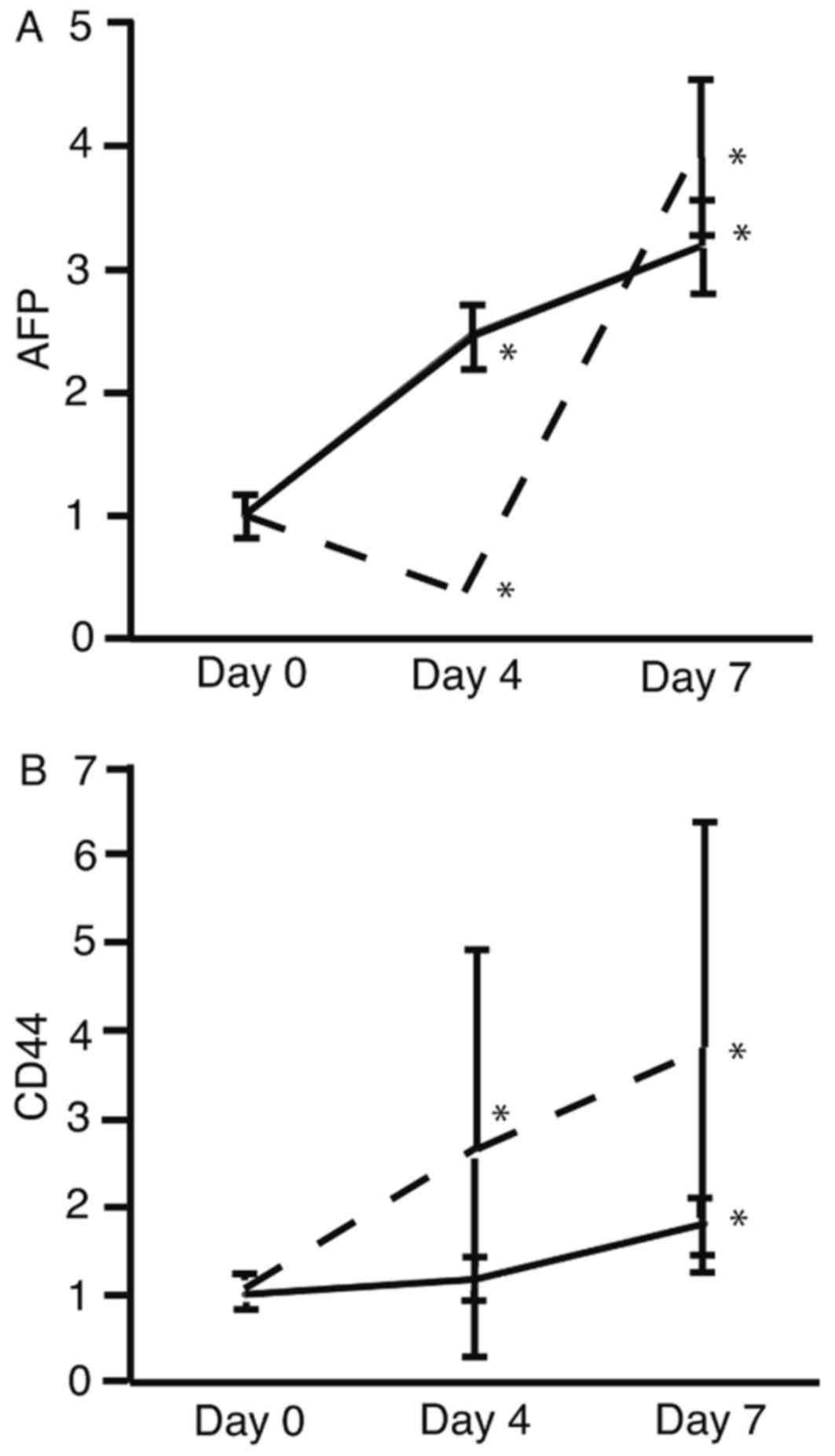
The mRNA expression levels of AFP (Fig. 2A) and CD44 (Fig. 2B) were analyzed using RT-qPCR. The
expression levels of AFP and CD44 were significantly increased on
day 7 in HLF and PLC/PRF/5 cells (P<0.05; Fig. 2). These results suggest that HSM
upregulated the expression of AFP and CD44.
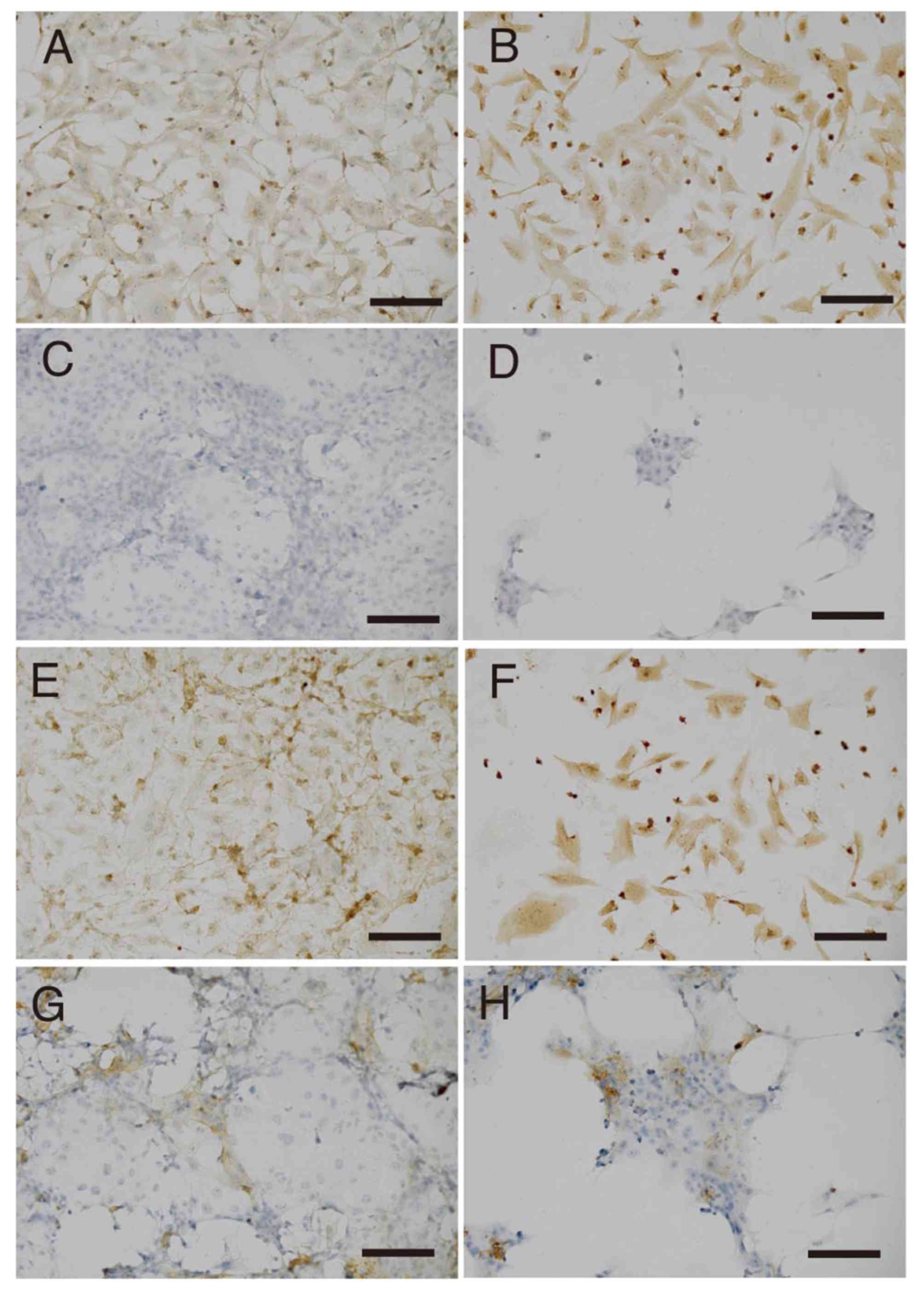
The expression levels of AFP and CD44 proteins were
analyzed by immunostaining. The expression of AFP was identified to
increase in HLF cells between day 0 (Fig.
3A) and day 7 (Fig. 3B), whereas
its expression in PLC/PRF/5 cells did not appear to alter during
this same timeframe (Fig. 3C and D).
The expression level of CD44 was also increased in HLF cells
between day 0 (Fig. 3E) and day 7
(Fig. 3F), but was not apparently
altered in PLC/PRF/5 cells (Fig. 3G and
H).
Discussion
Stem cells of HCC cloned with CD34 express AFP and
CD44 (20). AFP is expressed by HCC
cells through dedifferentiation (21). These studies suggest that HCC cells
positive for AFP and CD44 are markers of cancer stem cells of HCC.
In the present study, the expression levels of AFP and CD44 were
identified to increase following incubation in HSM. To the best of
our knowledge, there are currently no reported studies on the
culture of HCC cells in media lacking glucose. The biological
significance of the results of the present study is unclear;
however, there are two possibilities. One is that the
transcriptional activity of the AFP and CD44 genes was activated.
The other is that cancer stem cells were purified with HSM. To
determine which hypothesis may be true, the promoter activities of
AFP and CD44 should be analyzed. HCC cells positive for AFP exhibit
the characteristics of cancer stem cells and are associated with
poor prognosis (22,23). The results of these previous studies
and those of the present study suggest that HCC cells enriched in
HSM have stem cell-like characteristics. Suppressing the expression
of CD44 may be useful as a treatment for cells cultured in HSM
(24).
The cancer microenvironment is typically lacking in
glucose because the supply of glucose to the tumor is not
sufficient (25,26). In the present study, HCC cells were
cultured in HSM, a medium deficient in glucose, to mimic the
microenvironment of cancer. HSM was identified to be potentially
useful to create an experimental cancer microenvironment model,
such as fibrous tissues surrounding HCC (2). The results of the present study suggest
that cancer stem cells from HCC were enriched in HSM, which is a
potentially useful medium to enrich cancer stem cells from HCC.
A limitation of the present study was that the
resultant cells cultured in HSM may not definitively be classified
as stem cells. It is possible that these cells only have partial
cancer stem cell characteristics.
In future studies, cells obtained from culture in
HSM require propagation and investigation for cancer stem cell
characteristics including analysis of cell-surface markers and
spheroid formation assay (27,28).
The results of the present study identified that HSM
decreased the number of HCC cells, but the expression levels of AFP
and CD44 were increased in the cells obtained from culture in HSM.
Therefore, HSM is potentially useful for the enrichment of stem
cells of HCC.
Acknowledgements
The present study was supported by a Grant-in-Aid
for Scientific Research (C) from the Japan Society for the
Promotion of Science (grant no. 15K09032).
References
|
1
|
Cameron AM: Screening for viral hepatitis
and hepatocellular cancer. Surg Clin North Am. 95:1013–1021. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tomizawa M, Kondo F and Kondo Y: Growth
patterns and interstitial invasion of small hepatocellular
carcinoma. Pathol Int. 45:352–358. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leffert HL and Paul D: Studies on primary
cultures of differentiated fetal liver cells. J Cell Biol.
52:559–568. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ohira RH, Dipple KM, Zhang YH and McCabe
ER: Human and murine glycerol kinase: Influence of exon 18
alternative splicing on function. Biochem Biophys Res Commun.
331:239–246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ai Y, Jenkins NA, Copeland NG, Gilbert DH,
Bergsma DJ and Stambolian D: Mouse galactokinase: Isolation,
characterization, and location on chromosome 11. Genome Res.
5:53–59. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wheatley DN, Scott L, Lamb J and Smith S:
Single amino acid (arginine) restriction: Growth and death of
cultured HeLa and human diploid fibroblasts. Cell Physiol Biochem.
10:37–55. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tomizawa M, Toyama Y, Ito C, Toshimori K,
Iwase K, Takiguchi M, Saisho H and Yokosuka O: Hepatoblast-like
cells enriched from mouse embryonic stem cells in medium without
glucose, pyruvate, arginine, and tyrosine. Cell Tissue Res.
333:17–27. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tomizawa M, Shinozaki F, Sugiyama T,
Yamamoto S, Sueishi M and Yoshida T: Survival of primary human
hepatocytes and death of induced pluripotent stem cells in media
lacking glucose and arginine. PLoS One. 8:e718972013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghosn MG, Tuchin VV and Larin KV:
Depth-resolved monitoring of glucose diffusion in tissues by using
optical coherence tomography. Opt Lett. 31:2314–2316. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tomizawa M, Garfield S, Factor V and
Xanthopoulos KG: Hepatocytes deficient in CCAAT/enhancer binding
protein alpha (C/EBP alpha) exhibit both hepatocyte and biliary
epithelial cell character. Biochem Biophys Res Commun. 249:1–5.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tomizawa M, Wang YQ, Ebara M, Saisho H,
Watanabe K, Nakagawara A and Tagawa M: Decreased expression of the
CCAAT/enhancer binding protein alpha gene involved in hepatocyte
proliferation in human hepatocellular carcinomas. Int J Mol Med.
9:597–600. 2002.PubMed/NCBI
|
|
12
|
Guzman G, Alagiozian-Angelova V,
Layden-Almer JE, Layden TJ, Testa G, Benedetti E, Kajdacsy-Balla A
and Cotler SJ: p53, Ki-67, and serum alpha feto-protein as
predictors of hepatocellular carcinoma recurrence in liver
transplant patients. Mod Pathol. 18:1498–1503. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Orian-Rousseau V and Sleeman J: CD44 is a
multidomain signaling platform that integrates extracellular matrix
cues with growth factor and cytokine signals. Adv Cancer Res.
123:231–254. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hiraga T, Ito S and Nakamura H: Cancer
stem-like cell marker CD44 promotes bone metastases by enhancing
tumorigenicity, cell motility and hyaluronan production. Cancer
Res. 73:4112–4122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim R, Kim SB, Cho EH, Park SH, Park SB,
Hong SK and Chae G: CD44 expression in patients with combined
hepatocellular cholangiocarcinoma. Ann Surg Treat Res. 89:9–16.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Friemel J, Rechsteiner M, Frick L, Böhm F,
Struckmann K, Egger M, Moch H, Heikenwalder M and Weber A:
Intratumor heterogeneity in hepatocellular carcinoma. Clin Cancer
Res. 21:1951–1961. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakamura T, Teramoto H, Tomita Y and
Ichihara A: L-proline is an essential amino acid for hepatocyte
growth in culture. Biochem Biophys Res Commun. 122:884–891. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Davies B and Fried M: The L19 ribosomal
protein gene (RPL19): Gene organization, chromosomal mapping, and
novel promoter region. Genomics. 25:372–380. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tam S, Clavijo A, Engelhard EK and
Thurmond MC: Fluorescence-based multiplex real-time RT-PCR arrays
for the detection and serotype determination of foot-and-mouth
disease virus. J Virol Methods. 161:183–191. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park SC, Zeng C, Tschudy-Seney B, Nguyen
NT, Eun JR, Zhang Y, Ramsamooj R, Zhang Y, Zhao M, Theise ND, et
al: Clonogenically culturing and expanding CD34+ liver cancer stem
cells in vitro. Stem Cells Dev. 24:1506–1514. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mu X, Español-Suñer R, Mederacke I, Affò
S, Manco R, Sempoux C, Lemaigre FP, Adili A, Yuan D, Weber A, et
al: Hepatocellular carcinoma originates from hepatocytes and not
from the progenitor/biliary compartment. J Clin Invest.
125:3891–3903. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao X, Parpart S, Takai A, Roessler S,
Budhu A, Yu Z, Blank M, Zhang YE, Jia HL, Ye QH, et al: Integrative
genomics identifies YY1AP1 as an oncogenic driver in EpCAM(+)
AFP(+) hepatocellular carcinoma. Oncogene. 34:5095–5104. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Benzoubir N, Mussini C, Lejamtel C, Dos
Santos A, Guillaume C, Desterke C, Samuel D, Bréchot C, Bourgeade
MF and Guettier C: Gamma-smooth muscle actin expression is
associated with epithelial-mesenchymal transition and stem-like
properties in hepatocellular carcinoma. PLoS One. 10:e01305592015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao Y, Ruan B, Liu W, Wang J, Yang X,
Zhang Z, Li X, Duan J, Zhang F, Ding R, et al: Knockdown of CD44
inhibits the invasion and metastasis of hepatocellular carcinoma
both in vitro and in vivo by reversing epithelial-mesenchymal
transition. Oncotarget. 6:7828–7837. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheng GM and To KK: Adverse cell culture
conditions mimicking the tumor microenvironment upregulate ABCG2 to
mediate multidrug resistance and a more malignant phenotype. ISRN
Oncol. 2012:7460252012.PubMed/NCBI
|
|
26
|
Menendez JA, Oliveras-Ferraros C, Cufi S,
Corominas-Faja B, Joven J, Martin-Castillo B and Vazquez-Martin A:
Metformin is synthetically lethal with glucose withdrawal in cancer
cells. Cell Cycle. 11:2782–2792. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liao J, Qian F, Tchabo N,
Mhawech-Fauceglia P, Beck A, Qian Z, Wang X, Huss WJ, Lele SB,
Morrison CD and Odunsi K: Ovarian cancer spheroid cells with stem
cell-like properties contribute to tumor generation, metastasis and
chemotherapy resistance through hypoxia-resistant metabolism. PLoS
One. 9:e849412014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu CP, Wang AQ, Zhang HH, Wan XS, Yang
XB, Chen SG and Zhao HT: Research progress and prospects of markers
for liver cancer stem cells. World J Gastroenterol. 21:12190–12196.
2015. View Article : Google Scholar : PubMed/NCBI
|