Introduction
Laryngeal carcinoma is the 11th most common form of
cancer in humans worldwide (1).
Pathologically, >95% of all laryngeal malignancies are squamous
cell carcinomas (SCCs) (2). It has
been reported that >75% of cases of laryngeal SCC (LSCC) are
attributable to cigarette smoking and alcohol consumption.
Cigarette smoking is associated with an increased risk of LSCC
compared with that of individuals who have never smoked, and heavy
alcohol intake is an independent risk factor of LSCC (3). LSCC remains difficult to treat;
furthermore, treatment can cause severe long-term side effects. For
patients who are not cured by surgery and/or chemo-radiotherapy,
there are few effective treatment options (4). Thus, targeted therapies and predictive
biomarkers are urgently required in the perspective of LSCCs to
improve the management and minimize the treatment toxicity, as well
as to allow the selection of patients who are likely to benefit
from both non-selective and targeted therapies (5).
Previous studies have confirmed that tumor immune
inflammation is important in the tumor microenvironment (6). Cluster of differentiation (CD)
4+ T cells are essential organizers of cell-mediated
immunity, participating in each stage of the immune response. It
has been confirmed that naïve (uncommitted) CD4+ T cells
can be induced to different specific lineages according to the
local cytokine, including towards T helper (Th)1, Th2, Th17 and T
regulatory (Treg) cells (7).
Interleukin (IL)17-producing Th17 cells, which are different from
Th1 and Th2 cells, have been described as serving critical roles in
inflammation and autoimmune diseases, as well as in cancer
development (8–11). Considering these facts, it could be
concluded that pro-inflammatory Th17 cells may have extensive
effects on LSCC pathogenesis and anti-tumor response.
The present study revealed that patients with LSCC
have elevated levels of Th17 cells in their primary tumors and
peripheral blood compared with those in healthy controls. In
addition, the LSCC microenvironment was identified as a strong
Th17-cell inducer.
Materials and methods
Study subjects
A total of 70 tumors and 70 adjacent control tissues
(pathologically confirmed normal mucosa) were obtained from
patients with LSCC undergoing surgery. Peripheral blood was
obtained from another 36 patients with LSCC and from 16 healthy
individuals, who served as controls. The patients were registered
for treatment at the Eye, Ear, Nose and Throat Hospital of Fudan
University (Shanghai, China) from September 2013 to January 2015.
For LSCC samples, patients with a known history of LSCC were
enrolled, and malignancy was assessed by a pathological examination
of the biopsies. The tumor staging of the patients was performed in
accordance with the American Joint Committee on Cancer tumor-node
metastasis (TNM) classification (12). All donors participated on a voluntary
basis and provided written informed consent. The protocol was
approved (no. KJ2008-01) by the ethics committee of the Eye, Ear,
Nose and Throat Hospital of Fudan University (Shanghai, China) and
was in agreement with this institutions ethical guidelines.
Tissue microarray (TMA)
construction
TMA technology is a method where a large number of
tissue samples are placed on a microscopic glass slide, which
facilitates the transition from basic research to clinical
applications (13). Briefly, the most
representative tumor and adjacent tissue samples were selected in
pairs. The corresponding areas of each donor paraffin blocks were
perforated using a trephine needle (Quick Ray®; Unitma
Co., Ltd., Seoul, Korea) with a size of 1.5 mm. The 1.5-mm-sized
tissue cores were transferred and embedded into the recipient block
with 140 empty 1.5-mm-sized holes. All study specimens were
obtained with both tumor and adjacent tissue samples from each
donor block. Multiple 4-µm-thick sections were cut with a microtome
and transferred to poly-L-lysine-coated slides (#22247-1; Hannotech
Co., Ltd., Dongguan, China). TMA blocks were constructed, each
containing one sample from all tumors and adjacent tissues
(Fig. 1).
Immunohistochemical staining
Two TMA blocks with 70 tumors and 70 adjacent
tissues samples were used for immunostaining, followed by standard
procedures for the avidin-biotin-peroxidase method (14). The following anti-human monoclonal
antibodies were used: Rabbit anti-IL17 (#bs-2140R; BIOSS, Beijing,
China) and rabbit anti-IL17A receptor (IL17R; #bs-2606R; BIOSS).
Briefly, the color reaction was developed with a
3,3′-diaminobenzidine solution at 20°C for 5 min, and antibodies
were diluted to 1:200 and incubated at 37°C for 1 h. The cells were
counterstained with hematoxylin.
The slices were evaluated by two pathologists
without knowledge of the clinical outcome. The percentage of
immunoreactive cells was graded on a scale of 0 to 4, as follows:
No staining was scored as 0; 1–10% of stained cells was scored as
1; 11–50% of stained cells was scored as 2; 51–80% of stained cells
was scored as 3; and 81–100% of stained cells was scored as 4. The
staining intensities were graded from 0 to 3, as follows: 0 was
defined as negative; 1 as weak; 2 as moderate; and 3 as strong. The
raw data were converted into an immunohistochemical score (IHS) by
multiplying the quantity and intensity scores. An IHS score of 9–12
was considered to represent strong immunoreactivity (+++); 5–8 was
considered as moderate (++); 1–4 was considered as weak (+); and 0
was considered as negative immunoreactivity (−). On the final
analysis, the cases that had an IHS <1 were considered as
negative, and those with an IHS ≥1 were regarded as positive.
Flow cytometry
Peripheral blood mononuclear cells (PBMCs) were
isolated from 5 ml of freshly obtained peripheral blood by
centrifugation (800 × g, 20°C, 20 min) on a Ficoll Hypaque density
gradient (Ficoll PM 400; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). Prior to intracellular staining, the isolated PBMCs were
stimulated for 5 h with 2 µl/ml Cell Stimulation Cocktail
(#00-4970; eBioscience, Inc., San Diego, CA, USA), a cocktail of
phorbol 12-myristate 13-acetate (PMA; eBioscience, Inc.) and
ionomycin (eBioscience, Inc.) in the presence of Protein Transport
Inhibitor Cocktail (#560751; BD Biosciences, Franklin Lakes, NJ,
USA). Briefly, cells were fixed and permeabilised using the BD
Cytofix/Cytoperm™ Buffer (Fixation and Permeabilization Solution;
#560751; BD Biosciences) according to manufacturer's protocol.
Subsequently, the isolated PBMCs were intracellularly stained with
the Human Th1/Th2/Th17 Phenotyping Cocktail (560751; BD
Biosciences). Flow cytometry was performed on a BD FACSCalibur (BD
Biosciences) and the data were evaluated using FlowJo software
version 7.6.1 (TreeStar, Inc., Ashland, OR, USA). To determine the
percentage of Th17 and Th1 cells, lymphocytes were gated by
plotting forward vs. side scatter followed by gating on
CD4+ T cells. The gated cells were then analyzed for
IL-17A as phycoerythrin and interferon (IFN)-γ as fluorescein
isothiocyanate expression.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from patients' frozen
tissues using TRIzol reagent (15596-018; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the manufacturer's protocol.
Complementary DNA (cDNA) was synthesized from 1 µg total RNA in a
20-µl reaction system using PrimeScript RT Reagent kit (Perfect
Real Time; DRR063A; Takara, Bio, Inc., Otsu, Japan). The cDNA was
then diluted with sterile water and stored at −20°C. The RT
procedure using the PrimeScript RT Reagent kit was performed
according to the manufacturer's protocol.
RT-qPCR for IL17, IL23 and RAR-related orphan
receptor (ROR) γt was performed on an Applied Biosystems 7500 Fast
Real-Time PCR System (Thermo Fisher Scientific, Inc.), and the data
were analyzed using the Applied Biosciences Real-Tiem PCR system
7500 software version 2.0.6 (Applied Biosystems; Thermo Fisher
Scientific, Inc.). In brief, 2 µl cDNA was added to a 20-µl
reaction mixture containing 10 µl 2X SYBR Premix Ex Taq, 0.4 µl
forward primer (10 µM), 0.4 µl reverse primer (10 µM), 0.4 µl ROX
reference dye and 6.8 µl sterile water (all #DRR063A; Takara, Bio,
Inc., Otsu, Japan). All primers were designed using the Primer
Premier version 5.0 software (Premier Biosoft International, Palo
Alto, CA, USA), with their specificity confirmed by the Basic Local
Alignment Search Tool on the National Center for Biotechnology
Information website (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Detailed
information on the aforementioned primers is as follows: IL-17,
forward 5′-TCCCACGAAATCCAGGATGC-3′ and reverse
5′-GGATGTTCAGGTTGACCATCAC-3′; IL-23, forward
5′-TGCTCCCTGATAGCCCTGTGG-3′ and reverse
5′-GCTGGGACTGAGGCTTGGAAT-3′; and RORγt, forward
5′-GTGGGGACAAGTCGTCTGG-3′ and reverse 5′-AGTGCTGGCATCGGTTTCG-3′.
The PCR conditions were: 95°C for 30 sec, followed by 40 cycles at
95°C for 5 sec and 60°C for 25 sec. All samples were processed in
triplicate. The experiment was repeated for three times.
Relative gene expression was calculated using the
comparative quantification cycle (Cq) method (15). The messenger RNA expression levels of
the target genes were normalized by β-actin, and the
2−ΔCq values were represented. For data analysis, the
2−ΔCq method was used to calculate the fold-change in
expression, in which ΔCq represented the difference between the Cq
value of the target gene and that of β-actin (Cqtarget
gene-Cq β-actin).
Statistical analysis
The data were reported as the mean ± standard
deviation or the mean ± standard error. IL-17 and IL-17R expression
in tumors and adjacent tissues was evaluated by χ2 test.
The percentage of Th17 and Th17/Th1 cells in the peripheral blood
of patients with cancer compared with that of healthy controls was
assessed using t-tests. Cytokines IL17, IL23 and RORγt in LSCC
tissues compared with that of non-cancerous tissues were assessed
using one-way analysis of variance. All statistical calculations
were performed using SPSS version 19 (IBM SPSS, Armonk, NY, USA).
P<0.05 was considered to indicate a statistical significant
difference.
Results
Descriptive features of the study
groups
Table I shows the
descriptive characteristics of the study subjects, comprising 116
patients with LSCC and 16 healthy controls. Pathologically, all
patients had SCC. Of a total of 116 patients, 115 patients (99.14%)
were males and only 1 (0.86%) was female. The control group had a
similar sex distribution, including 16 males (100.00%).
 | Table I.Characteristic features of the
patients in the current study. |
Table I.
Characteristic features of the
patients in the current study.
|
Characteristics | Patients (fresh
tissue) (n=70), n (%) | Patients (blood)
(n=36), n (%) | Controls (blood)
(n=16), n (%) |
|---|
| Age, years |
|
|
|
| Mean
(range) | 60.63 (38–84) | 59.10 (43–73) | 59.50 (52–72) |
| Sex |
|
|
|
|
Male | 69 (98.57) | 36 (100.00) | 16 (100.00) |
|
Female | 1 (1.43) | 0 (0.00) | 0 (0.00) |
| Site |
|
|
|
|
Supraglottic | 23 (32.86) | 6 (16.67) |
|
|
Glottic | 40 (57.14) | 26 (72.22) |
|
|
Infraglottic | 7 (10.00) | 4 (11.11) |
|
| cT stage |
|
|
|
|
T1+T2 | 17 (24.29) | 25 (69.44) |
|
|
T3+T4 | 53 (75.71) | 11 (30.56) |
|
| pN stage |
|
|
|
|
N0 | 33 (47.14) | 31 (86.11) |
|
|
N1+N2 | 37 (52.86) | 5 (13.89) |
|
| Clinical stage |
|
|
|
| Early
stage (I+II) | 6 (8.57) | 26 (72.22) |
|
| Late
stage (III+IV) | 64 (91.43) | 10 (27.78) |
|
| Smoking
history |
|
|
|
|
Smokers | 62 (88.57) | 25 (69.44) |
|
|
Non-smokers | 8 (11.43) | 11 (30.56) |
|
| Alcohol
consumption |
|
|
|
|
Drinkers | 34 (48.57) | 17 (47.22) |
|
|
Non-drinkers | 36 (51.43) | 19 (52.78) |
|
Immunohistochemical observations
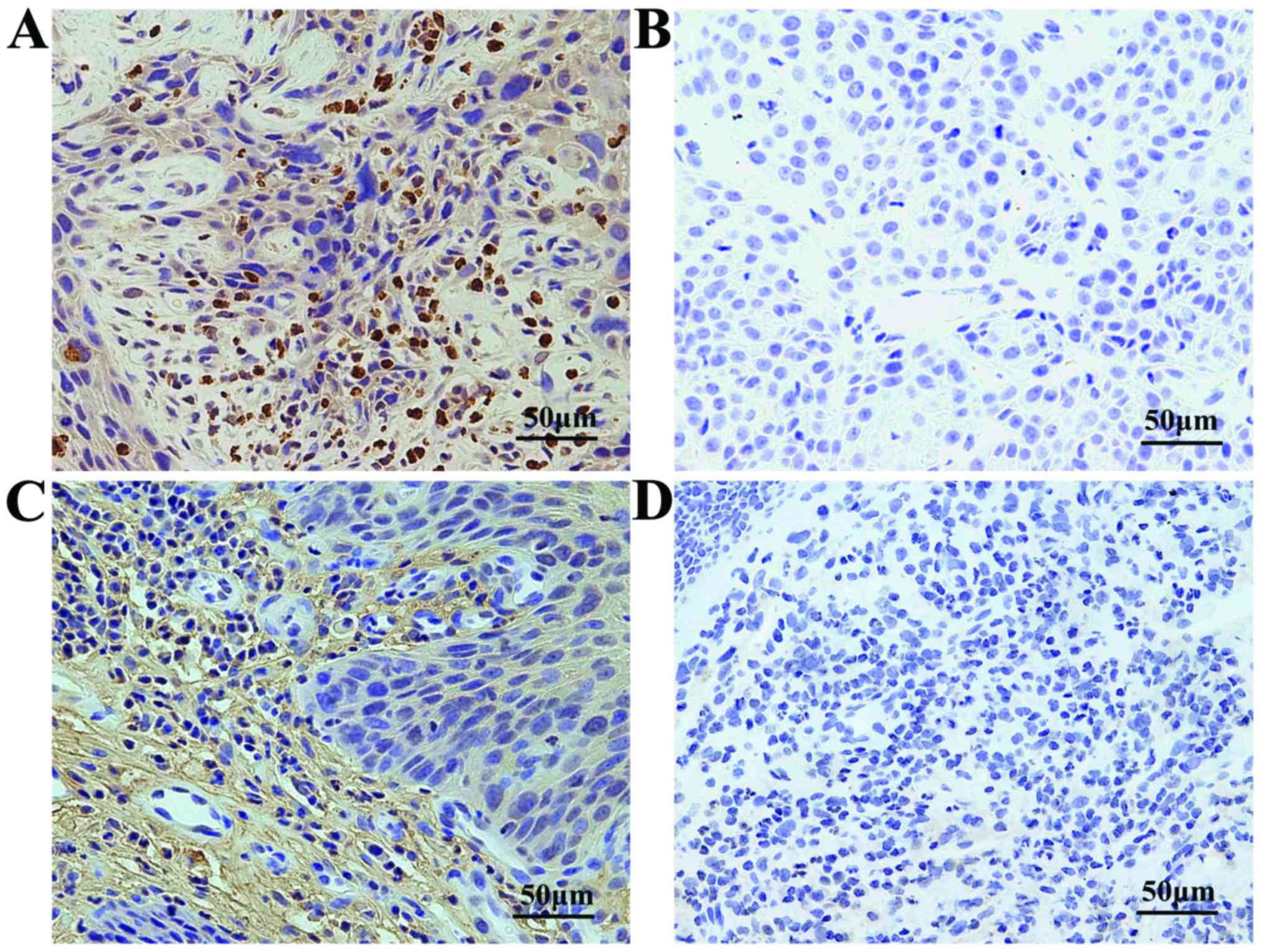
To study the expression of IL17 and IL17R in
vivo, the tumors and adjacent tissues were stained for IL17 and
IL17R using immunohistochemistry (Fig.
2). Differential IL-17 and IL-17R expression in tumors and
adjacent tissues was confirmed.
Overall, positive staining for IL17 was noted in 57
(81.43%) of 70 tumors with the following scores: 0, 13 (18.57%) of
70; 1–4, 16 (22.86%) of 70; 5–8, 27 (38.57%) of 76; and 9–12, 14
(20.00%) of 70 samples. Among the controls, positive staining for
IL17 was noted in 26 (37.14%) of 70 samples. IL17R positive
staining was observed in 64 (91.43%) of 70 cases. These included 0,
6 (8.57%) of 70; 1–4, 22 (31.43%) of 70; 5–8, 28 (40.00%) of 70;
and 9–12, 14 (20.00%) of 70 cases. Among the controls, positive
staining for IL17R was noted in 49 (70.00%) of 70 samples (Table II).
| Table II.Positive results of IL17/IL17R
immunohistochemistry in tumors and controls. |
Table II.
Positive results of IL17/IL17R
immunohistochemistry in tumors and controls.
| Molecule | Tumors, n (%) | Controls, n
(%) | P-value |
|---|
| IL17 | 57 (81.43) | 26 (37.14) | <0.05 |
| IL17R | 64 (91.43) | 49 (70.00) | <0.05 |
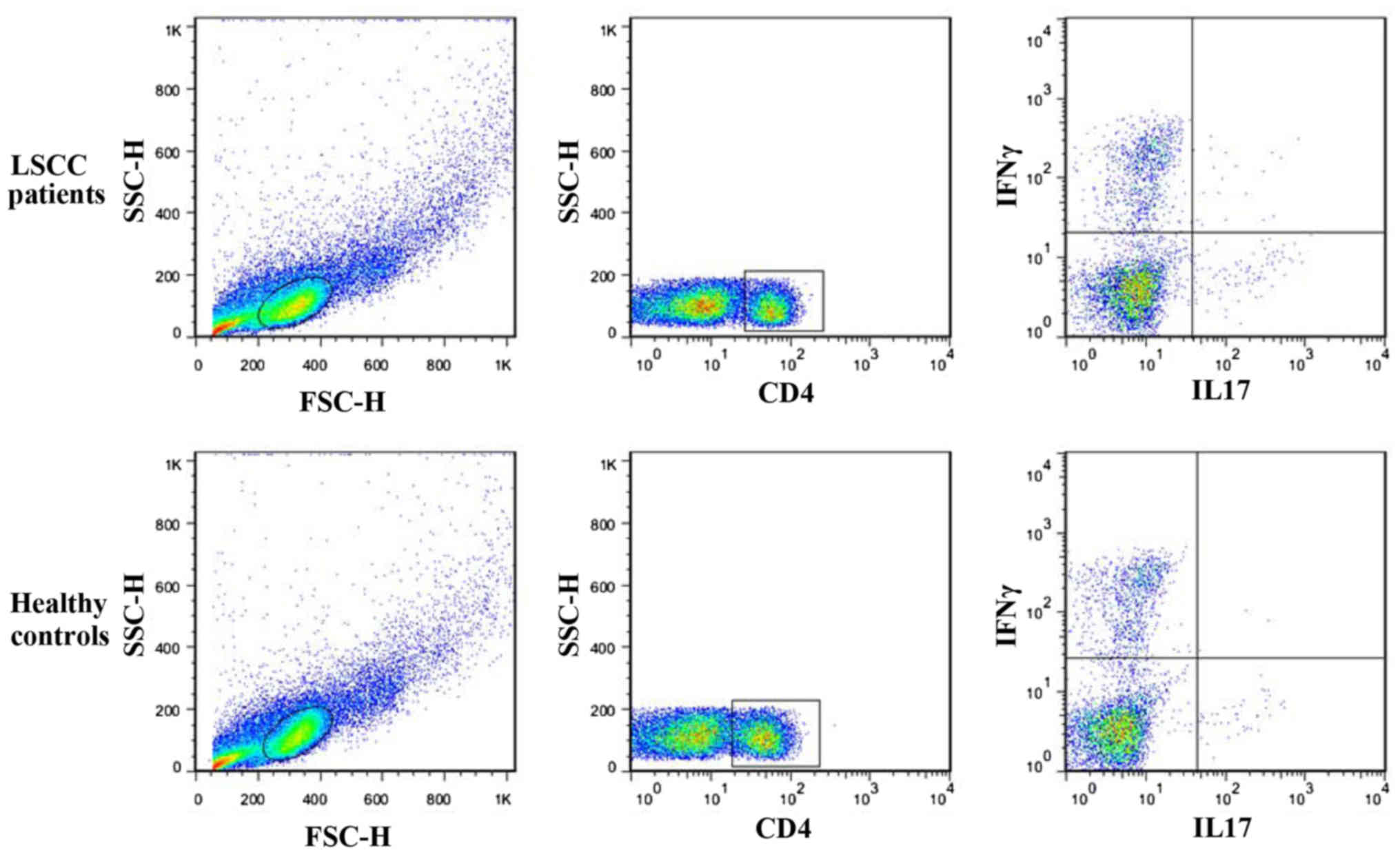
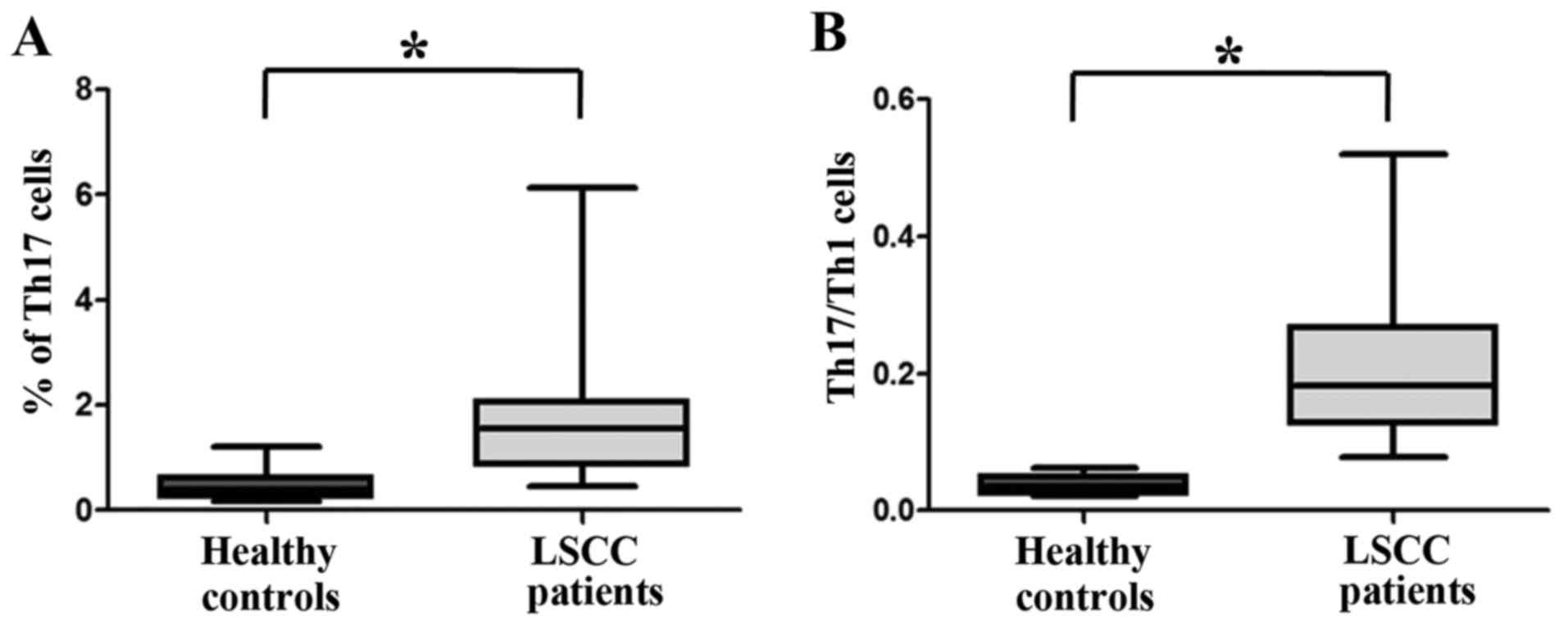
Elevated levels of Th17 cells in the
peripheral blood of patients with LSCC
The prevalence of Th17 cells in the peripheral blood
of 36 patients with LSCC was compared with that of 16 healthy
controls to address whether there was an increased prevalence of
Th17 cells in patients with LSCC. PBMCs were isolated and
stimulated with PMA and ionomycin for 5 h in the presence of
Protein Transport Inhibitor, and Th17 and Th1 cells were next
quantified by flow cytometry. Th17 cells were identified as a
CD4+IL17+ cell population. Th1 cells were
identified as a CD4+IFN-γ+ cell population.
As shown in Figs. 3 and 4A, there was a statistically significantly
higher percentage of Th17 cells in the peripheral blood of patients
with cancer (1.6860±0.1866%, n=36) compared with that of healthy
controls (0.4963±0.7862%, n=16; P<0.05).
Th17/Th1 cells are increased in the
peripheral blood of patients with LSCC
Th17/Th1 cells, which simultaneously produce the
Th17 cytokine IL17 and the Th1 cytokine IFN-γ (16), accounted for ~3.714% in healthy
individuals (3.714±0.3487%; n=16). In the peripheral blood of
patients with LSCC, this subpopulation was significantly increased
(21.03±1.750%; n=36; P<0.05; Fig.
4B).
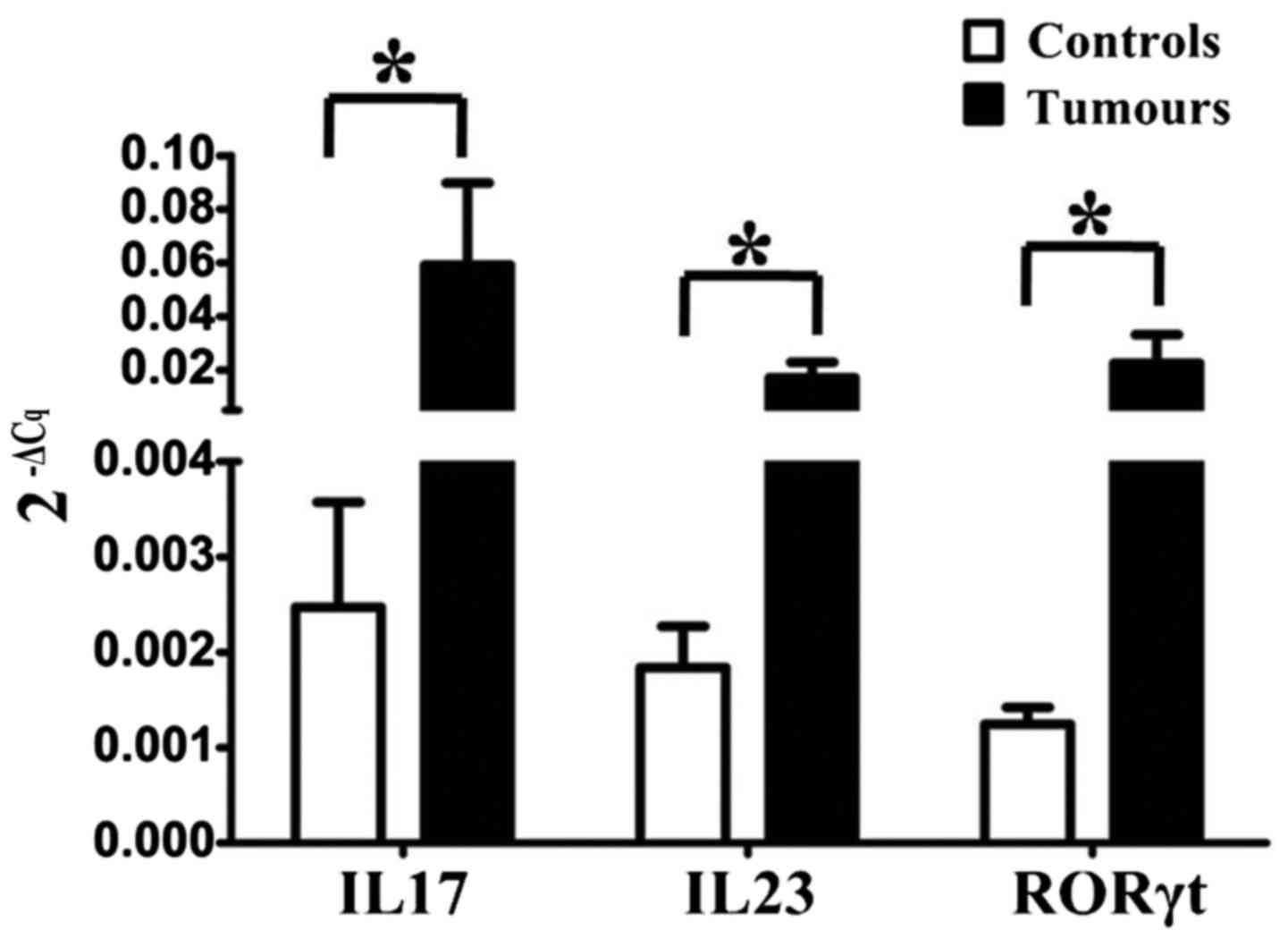
Upregulation of intracellular
cytokines IL17, IL23 and RORγt in Th17 cells
RT-qPCR revealed that the Th17-associated
intracellular cytokines and transcription factors IL17, IL23 and
RORγt in LSCC tissues were upregulated compared with their levels
in matched adjacent non-cancerous tissues (Fig. 5). This indicated that Th17 cells
expanded in the LSCC tissue cell population, and may be involved in
LSCC development and metastasis.
Discussion
Tumors grow in a complex and active
microenvironment. Within or nearby the tumor nests, lymphocytes as
well as endothelial, stromal and innate cells are present, which
interact with each other to form the tumor microenvironment
(17). Inflammation is causally
associated with cancer development through processes that involve
genotoxicity, aberrant tissue repair, proliferative responses,
invasion and metastasis (17). Tumors
modulate the inflammatory environment through the secretion of
soluble growth factors and chemoattractants, which render
inflammatory cells suppressive against anti-cancer T cell responses
(17). CD4+ Th cells, as a
highly heterogeneous population, serve critical roles in tumor
immunological responses (18).
Initially, immunologists considered that CD4+ T cells
mainly included two types, namely Th1 and Th2 (19); however, at least four distinct
CD4+ T cell subsets have been confirmed to exist,
specifically Th1, Th2, Th17 and Treg cells (19).
LSCCs produce various immunosuppressive and
tumor-promoting cytokines, leading to an impaired anti-tumor
response (20). In previous
cytogenetic studies, it was observed that multiple signaling
pathways were activated in LSCC, including the p53, vascular
endothelial growth factor, epidermal growth factor receptor,
transforming growth factor (TGF)-β and nuclear factor-κB signaling
pathways (21). In addition, it has
been well established that inflammation is closely connected to
LSCC development due to the induction of chronic inflammation
caused by exposure to irritants in inhaled air, particularly
cigarette smoke. Such irritants enhance the accumulation of viruses
and airborne microbes, thereby promoting tumor growth (22). Considering these facts, it can be
concluded that pro-inflammatory Th17 cells have extensive effects
on LSCC pathogenesis and anti-tumor response.
It was previously shown that Th17 cells are involved
in tissue inflammation by inducing the release of various
cytokines, including IL6, IL21, IL23, IL1β and TGF-β, by
neighboring tumor cells, tumor-derived fibroblasts and
antigen-presenting cells (23). Human
Th17 cells mainly release the pro-inflammatory cytokine IL17, and
one important role of IL17 appears to be the regulation of local
inflammation through the upregulation of other pro-inflammatory
cytokines and chemokines (24). In
this regard, it is reasonable to propose the impact of Th17 cells
on cancer pathogenesis and progression. In gastric cancer, there
was both an elevation of Th17 frequency in peripheral blood and
tumor-draining lymph nodes (25).
Furthermore, in ovarian cancer, it was shown that Th17 cells were
increased in tumor tissue, but not in peripheral blood (15). However, the function of Th17 cells and
their influence on LSCC development in humans remain unknown.
Several studies demonstrated that human Th17 cells
produce IL17A and exhibit RORγt expression (26,27). These
cells also express IL17R, which is also expressed by Th1 cells
(26,27). Human Th17 cells appear to exclusively
originate from a small subset of T-cell precursors, which
constitutively express RORγt and IL-17R, and develop into Th17
cells in response to IL1β and IL23 in the apparent absence of TGF-β
(28). Furthermore, even established
Th17 cells can be induced to produce IFN-γ in addition to IL17A
(Th17/Th1 cells), suggesting a common developmental association
between Th17 and at least a subset of Th1 cells (28). Notably, numerous studies have reported
increased expression of IL17/Th17 cells in different cancers,
including colorectal (29), prostate
(30), gastric (31), lung (32), ovarian (33), oral (34) and head and neck (35) carcinomas.
The present study revealed that patients with LSCC
have elevated levels of Th17 cells in their primary tumors and
peripheral blood compared with those exhibited by healthy controls.
The LSCC microenvironment was identified as a strong Th17-cell
inducer. First, histopathological characterization was performed,
and immunohistochemistry staining was used to detect IL17 and IL17R
expression in patients with LSCC. Overexpression of both IL17 and
IL17R was observed in tumors compared with the expression detected
in adjacent tissues. Next, whether Th17 cells have any functional
implications for LSCC development was evaluated. The frequencies of
Th17 and Th1 cells in the peripheral blood of patients with LSCC
were investigated, and a higher percentage of Th17 cells was
detected in the peripheral blood of patients with LSCC compared
with that in healthy controls. No association was observed between
TNM staging and Th17 cell frequency, which suggests that Th17 cells
were consistently elevated in patients with LSCC, independently of
tumor stage. This may represent a link between inflammation and
cancer; however, the exact mechanisms by which elevated levels of
pro-inflammatory Th17 cells and the resultant secretion of
cytokines contribute to inflammatory processes in cancer remain to
be elucidated.
The present study demonstrated that Th17 cells can
be induced by PBMCs of patients with LSCC. Upon incubation of
isolated naïve CD4+ T cells in LSCC PBMCs in
vitro, a significantly elevated number of Th17 cells was
detected. Therefore, it can be proposed that the LSCC
microenvironment is able to induce a Th17 lineage commitment. It
was also observed that Th1 cells were downregulated in peripheral
blood. It is known that Th1 cytokines, such as IFN-γ, are
diminished in LSCC (36), whereas
IL17 is upregulated; thus, this may be the mechanism responsible
for Th17 and Th1/Th17 cell modulation in LSCC. However, the outcome
of these Th1/Th17 cells under tumor influence (whether they simply
attenuate their IFN-γ secretion or change into another T-cell
population) remains to be investigated. Our hypothesis is that Th1
cells are functionally modulated by the tumor microenvironment and
are converted into Th17 cells.
RT-qPCR demonstrated that the levels of
Th17-associated intracellular cytokines and transcription factors
(including IL17, IL23 and RORγt) of LSCC tissue were upregulated.
Since it was shown that IL23 and RORγt lead to Th17 expansion
(37), as well as to selective
enrichment of IL17-producing cells by modulating the proliferation
of memory T cells (38), the present
study was able to show that IL23 and RORγt levels in LSCC lead to a
strong enhancement of Th17 cell expansion directly at the tumor
site. There have been different reports about the influence of Th17
on tumor progression, a number of which depict a positive influence
of Th17 on tumor proliferation (39,40), while
others describe an inhibitory influence on tumor growth (41,42). Ciree
et al (43) demonstrated that
Th17 cells are upregulated in the T-cell lymphomas mycosis
fungoides and Sézary syndrome, and may act as a tumor
growth-promoting or -inhibiting factor. In addition, the authors
observed an association between Th17 expression and
polymorphonuclear neutrophil infiltration. Muranski et al
(44) reported that Th17 cells in a
mouse model were able to eradicate melanomas. This association was
affirmed by Garcia-Hernandez et al (45); in their description, neutrophils were
attracted to the tumor microenvironment by a Th17-dependent
mechanism, and indicated that depletion of neutrophils resulted in
a diminished capacity to control tumor growth. In addition,
Honorati et al (46) reported
an increased susceptibility of osteosarcoma cells to natural killer
cells under the influence of Th17.
Altogether, the present study demonstrated that Th17
cells are highly present in LSCC. However, this appears to act as a
double-edged sword: On one hand, Th17 cells accelerate metastasis
and appear to be, therefore, beneficial to tumors; on the other
hand, Th17 cells appear to be beneficial to the host due to their
proliferation-reducing activity. Therefore, it is important to
study the function of Th17 cells in malignant diseases in depth,
and to attempt to elucidate their mechanism of action and their
modulation by the tumor microenvironment. The present results raise
a further issue to investigate, namely whether Th17 cells express
different molecules or secrete different cytokines in patients with
LSCC compared with those in healthy individuals. Future studies
should consider whether Th17 cells can recognize tumor cells and if
they are at all able to impair tumor growth or metastasis in
vivo; in addition, their exact potential mechanism of action
must be elucidated.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 30801283 and
30972691), the Shanghai Science and Technology Development Funds
(grant nos. 09QA1401000, 10QA1405900 and 14411961900), the Training
Program of the Excellent Young Talents of the Shanghai Municipal
Health System (grant nos. XYQ2011055 and XYQ2011015) and the
Shanghai Municipal Science and Technology Foundation (grant no.
11JC1410802).
References
|
1
|
Marioni G, Marchese-Ragona R, Cartei G,
Marchese F and Staffieri A: Current opinion in diagnosis and
treatment of laryngeal carcinoma. Cancer Treat Rev. 32:504–515.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Almadori G, Bussu F, Cadoni G, Galli J,
Paludetti G and Maurizi M: Molecular markers in laryngeal squamous
cell carcinoma: Towards an integrated clinicobiological approach.
Eur J Cancer. 41:683–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hashibe M, Brennan P, Benhamou S,
Castellsague X, Chen C, Curado MP, Dal Maso L, Daudt AW, Fabianova
E, Fernandez L, et al: Alcohol drinking in never users of tobacco,
cigarette smoking in never drinkers and the risk of head and neck
cancer: Pooled analysis in the International Head and Neck Cancer
Epidemiology Consortium. J Natl Cancer Inst. 99:777–789. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brandstorp-Boesen J, Falk RS, Boysen M and
Brøndbo K: Long-term trends in gender, T-stage, subsite and
treatment for laryngeal cancer at a single center. Eur Arch
Otorhinolaryngol. 271:3233–3239. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suh Y, Amelio I, Urbano T Guerrero and
Tavassoli M: Clinical update on cancer: Molecular oncology of head
and neck cancer. Cell Death Dis. 23:e10182014. View Article : Google Scholar
|
|
6
|
Qi W, Huang X and Wang J: Correlation
between Th17 cells and tumour microenvironment. Cell Immunol.
285:18–22. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Middleton GW, Annels NE and Pandha HS: Are
we ready to start studies of Th17 cell manipulation as a therapy
for cancer? Cancer Immunol Immunother. 61:1–7. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ye J, Livergood RS and Peng G: The role
and regulation of human Th17 cells in tumour immunity. Am J Pathol.
182:10–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maddur MS, Miossec P, Kaveri SV and Bayry
J: Th17 cells: Biology, pathogenesis of autoimmune and inflammatory
diseases, and therapeutic strategies. Am J Pathol. 181:8–18. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wilke CM, Bishop K, Fox D and Zou W:
Deciphering the role of Th17 cells in human disease. Trends
Immunol. 32:603–611. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kimura A and Kishimoto T: Th17 cells in
inflammation. Int Immunopharmacol. 11:319–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Paleri V, Mehanna H and Wight RG: TNM
classification of malignant tumours 7th edition: What's new for
head and neck? Clin Otolaryngol. 35:270–272. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Torhorst J, Bucher C, Kononen J, Haas P,
Zuber M, Köchli OR, Mross F, Dieterich H, Moch H, Mihatsch M, et
al: Tissue microarrays for rapid linking of molecular changes to
clinical endpoints. Am J Pathol. 159:2249–2256. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vosse BA, Seelentag W, Bachmann A, Bosman
FT and Yan P: Background staining of visualization systems in
immunohistochemistry: Comparison of the Avidin-Biotin Complex
system and the EnVision+ system. Appl Immunohistochem Mol Morphol.
15:103–107. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Miyahara Y, Odunsi K, Chen W, Peng G,
Matsuzaki J and Wang RF: Generation and regulation of human CD4+
IL-17-producing T cells in ovarian cancer. Proc Natl Acad Sci USA.
105:pp. 15505–15510. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Llosa NJ, Geis AL, Orberg E Thiele and
Housseau F: Interleukin-17 and type 17 helper T cells in cancer
management and research. Immunotargets Ther. 3:39–54.
2014.PubMed/NCBI
|
|
17
|
Elinav E, Nowarski R, Thaiss CA, Hu B, Jin
C and Flavell RA: Inflammation-induced cancer: Crosstalk between
tumours, immune cells and microorganisms. Nat Rev Cancer.
13:759–771. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu J and Paul WE: Heterogeneity and
plasticity of T helper cells. Cell Res. 20:4–12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu J and Paul WE: CD4 T cells: Fates,
functions, and faults. Blood. 112:1557–1569. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Johnson SD, De Costa AM and Young MR:
Effect of the premalignant and tumour microenvironment on immune
cell cytokine production in head and neck cancer. Cancers (Basel).
6:756–770. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Görögh T and Beier UH: Gene alterations in
head and neck carcinomas and their role in promoting malignant
behavior (Review). Int J Oncol. 36:525–532. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Douglas WG, Tracy E, Tan D, Yu J, Hicks WL
Jr, Rigual NR, Loree TR, Wang Y and Baumann H: Development of head
and neck squamous cell carcinoma is associated with altered
cytokine responsiveness. Mol Cancer Res. 2:585–593. 2004.PubMed/NCBI
|
|
23
|
Chung Y, Chang SH, Martinez GJ, Yang XO,
Nurieva R, Kang HS, Ma L, Watowich SS, Jetten AM, Tian Q and Dong
C: Critical regulation of early Th17 cell differentiation by
interleukin-1 signaling. Immunity. 30:576–587. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park H, Li Z, Yang XO, Chang SH, Nurieva
R, Wang YH, Wang Y, Hood L, Zhu Z, Tian Q and Dong C: A distinct
lineage of CD4 T cells regulates tissue inflammation by producing
interleukin 17. Nat Immunol. 6:1133–1141. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang B, Rong G, Wei H, Zhang M, Bi J, Ma
L, Xue X, Wei G, Liu X and Fang G: The prevalence of Th17 cells in
patients with gastric cancer. Biochem Biophys Res Commun.
374:533–537. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Acosta-Rodriguez EV, Rivino L, Geginat J,
Jarrossay D, Gattorno M, Lanzavecchia A, Sallusto F and Napolitani
G: Surface phenotype and antigenic specificity of human interleukin
17-producing T helper memory cells. Nat Immunol. 8:639–646. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Annunziato F, Cosmi L, Liotta F, Maggi E
and Romagnani S: The phenotype of human Th17 cells and their
precursors, the cytokines that mediate their differentiation and
the role of Th17 cells in inflammation. Int Immunol. 20:1361–1368.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Romagnani S, Maggi E, Liotta F, Cosmi L
and Annunziato F: Properties and origin of human Th17 cells. Mol
Immunol. 47:3–7. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wägsäter D, Löfgren S, Hugander A and
Dimberg J: Expression of interleukin-17 in human colorectal cancer.
Anticancer Res. 26:4213–4216. 2006.PubMed/NCBI
|
|
30
|
Sfanos KS, Bruno TC, Maris CH, Xu L,
Thoburn CJ, DeMarzo AM, Meeker AK, Isaacs WB and Drake CG:
Phenotypic analysis of prostate-infiltrating lymphocytes reveals
TH17 and Treg skewing. Clin Cancer Res. 14:3254–3261. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang B, Rong G, Wei H, Zhang M, Bi J, Ma
L, Xue X, Wei G, Liu X and Fang G: The prevalence of Th17 cells in
patients with gastric cancer. Biochem Biophys Res Commun.
374:533–537. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Koyama K, Kagamu H, Miura S, Hiura T,
Miyabayashi T, Itoh R, Kuriyama H, Tanaka H, Tanaka J, Yoshizawa H,
et al: Reciprocal CD4+ T-cell balance of effector CD62Llow CD4+ and
CD62LhighCD25+ CD4+ regulatory T cells in small cell lung cancer
reflects disease stage. Clin Cancer Res. 14:6770–6779. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kryczek I, Banerjee M, Cheng P, Vatan L,
Szeliga W, Wei S, Huang E, Finlayson E, Simeone D, Welling TH, et
al: Phenotype, distribution, generation, and functional and
clinical relevance of Th17 cells in the human tumour environments.
Blood. 114:1141–1149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gaur P, Qadir GA, Upadhyay S, Singh AK,
Shukla NK and Das SN: Skewed immunological balance between Th17
(CD4(+)IL17A (+)) and Treg (CD4 (+)CD25 (+)FOXP3 (+)) cells in
human oral squamous cell carcinoma. Cell Oncol (Dordr). 35:335–343.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kesselring R, Thiel A, Pries R, Trenkle T
and Wollenberg B: Human Th17 cells can be induced through head and
neck cancer and have a functional impact on HNSCC development. Br J
Cancer. 103:1245–1254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bose A, Chakraborty T, Chakraborty K, Pal
S and Baral R: Dysregulation in immune functions is reflected in
tumour cell cytotoxicity by peripheral blood mononuclear cells from
head and neck squamous cell carcinoma patients. Cancer Immun.
8:102008.PubMed/NCBI
|
|
37
|
Chizzolini C, Chicheportiche R, Alvarez M,
De Rham C, Roux-Lombard P, Ferrari-Lacraz S and Dayer JM:
Prostaglandin E2 synergistically with interleukin-23 favors human
Th17 expansion. Blood. 112:3696–3703. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Napolitani G, Acosta-Rodriguez EV,
Lanzavecchia A and Sallusto F: Prostaglandin E2 enhances Th17
responses via modulation of IL-17 and IFN-gamma production by
memory CD4+ T cells. Eur J Immunol. 39:1301–1312. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Benchetrit F, Ciree A, Vives V, Warnier G,
Gey A, Sautès-Fridman C, Fossiez F, Haicheur N, Fridman WH and
Tartour E: Interleukin-17 inhibits tumour cell growth by means of a
T-cell-dependent mechanism. Blood. 99:2114–2121. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Numasaki M, Watanabe M, Suzuki T,
Takahashi H, Nakamura A, McAllister F, Hishinuma T, Goto J, Lotze
MT, Kolls JK and Sasaki H: IL-17 enhances the net angiogenic
activity and in vivo growth of human non-small cell lung cancer in
SCID mice through promoting CXCR-2-dependent angiogenesis. J
Immunol. 175:6177–6189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nam JS, Terabe M, Kang MJ, Chae H, Voong
N, Yang YA, Laurence A, Michalowska A, Mamura M, Lonning S, et al:
Transforming growth factor beta subverts the immune system into
directly promoting tumour growth through interleukin-17. Cancer
Res. 68:3915–3923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kryczek I, Wei S, Szeliga W, Vatan L and
Zou W: Endogenous IL-17 contributes to reduced tumour growth and
metastasis. Blood. 114:357–359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ciree A, Michel L, Camilleri-Bröet S,
Louis F Jean, Oster M, Flageul B, Senet P, Fossiez F, Fridman WH,
Bachelez H and Tartour E: Expression and activity of IL-17 in
cutaneous T-cell lymphomas (mycosis fungoides and Sezary syndrome).
Int J Cancer. 112:113–120. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Muranski P, Boni A, Antony PA, Cassard L,
Irvine KR, Kaiser A, Paulos CM, Palmer DC, Touloukian CE, Ptak K,
et al: Tumour-specific Th17-polarized cells eradicate large
established melanoma. Blood. 112:362–373. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Garcia-Hernandez Mde L, Hamada H, Reome
JB, Misra SK, Tighe MP and Dutton RW: Adoptive transfer of
tumour-specific Tc17 effector T cells controls the growth of B16
melanoma in mice. J Immunol. 184:4215–4227. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Honorati MC, Neri S, Cattini L and
Facchini A: IL-17 enhances the susceptibility of U-2 OS
osteosarcoma cells to NK cell lysis. Clin Exp Immunol. 133:344–349.
2003. View Article : Google Scholar : PubMed/NCBI
|