Introduction
Osteosarcoma (OS) is the most common type of primary
malignant bone tumor in children and adolescents. OS accounts for
almost 60% of malignant bone tumors in individuals <20 years old
(1). Although the diagnosis and
treatment of OS with adjuvant chemotherapy and local excision is
improving, pulmonary metastasis occurs in 40–50% of patients with
OS (2). When metastatic disease
develops after radiotherapy with curative intent, the prognosis is
particularly poor (3). Tumor
metastasis remains a critical obstacle to the treatment of OS.
Arsenic trioxide (As2O3) has been effective
at inducing the apoptosis of certain types of cancer cells and
solid tumors (4,5). We previously reported that
As2O3 suppressed the growth of the human OS
MG63 cell line, and reversed the doxorubicin (ADM) resistance of
ADM-resistant MG63 (MG63/dox) cells by inducing apoptosis and
downregulating stathmin expression in vitro (6). However, the effects of
As2O3 on OS cell invasion and metastasis have
yet to be clarified. Thus, the present study aimed to elucidate the
role of As2O3 and stathmin in regulating the
invasive activity of OS cells, in vitro and in
vivo.
Stathmin (also known as oncoprotein 18) is a
microtubule-destabilizing protein. Microtubules serve a critical
role in the mediation of cell motility, which is inhibited if
microtubules are stabilized. Stathmin is involved in tumor cell
proliferation, invasion and motility (7).
The mitogen-activated protein kinase (MAPK) pathway
is associated with a series of signaling cascades, mediating
diverse intracellular responses. One potential target of MAPKs is
stathmin, a protein that serves an important role in the regulation
of microtubules, which are required for cell motility (8). In the present study, the MAPK pathway
was demonstrated to be associated with the cellular response to
As2O3. E-cadherin has been associated with
metastatic progression in several types of cancer. The dissociation
of the E-cadherin/β-catenin adhesion complex represents a key step
in the epithelial-mesenchymal transition (EMT), a major mechanism
for tumor cell metastasis. Repressing the expression of E-cadherin
may promote EMT and induce cancer invasion (9,10). Thus,
the effect of As2O3 on E-cadherin expression,
and the association between stathmin and cell invasion, were also
investigated in the present study.
Materials and methods
Cell lines
Human embryonic kidney HEK293T cells, the human OS
MG63 cell line and the normal human osteoblast hFoB1.19cell line
were purchased from the Shanghai Institute of Biochemistry and Cell
Biology (Chinese Academy of Sciences, Shanghai, China). The human
OSMG63/dox multidrug-resistant (MDR) cell line was provided by Dr
Yoshio Oda (Graduate School of Medical Sciences, Kyushu University,
Fukuoka, Japan). Cells were cultured in high-glucose Dulbecco's
modified Eagle's medium (DMEM; Hyclone; GE Healthcare, Logan, UT,
USA) and supplemented with 10% fetal bovine serum (FBS; Hyclone; GE
Healthcare), 100 U/ml penicillin and 100 g/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 37°C in a
humidified, 5% CO2 atmosphere.
Reagents and antibodies
As2O3 was obtained from
Beijing SL Pharmaceutical Co., Ltd. (Beijing, China) and ADM from
Pfizer, Inc. (New York, NY, USA). PD098095 (cat. no. 9900) and
SP600125 (cat. no. 8177) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Antibodies against MAPK 3/1
(ERK1/2, cat. no. 5013) and 8 (JNK; cat. no. 9252),
phosphorylated-ERK1/2 (P-ERK; cat. no. 4370) and phosphorylated-JNK
(P-JNK, cat. no. 9255) were purchased from Cell Signaling
Technology, Inc. Antibodies against E-cadherin (cat. no. ab15148)
and β-catenin (cat. no. ab22656) were purchased from Abcam
(Cambridge, UK). The antibody against β-actin (cat. no. cw0096 m)
was purchased from Beijing Co Win Bioscience Co., Ltd (Beijing,
China).
Invasion assay
Invasion assays were performed using a Transwell
system (Corning Inc., Corning, NY, USA). The polycarbonate membrane
of the chamber was coated with 250 µg/ml Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA). The bottom chambers were filled with DMEM
containing 10% FBS, and serum-free DMEM was added to the top
chambers. A total of 2×104 cells treated with 2 µM
As2O3 and/or 200 ng/ml ADM per well were
added into the top chamber, followed by 24 h of incubation at 37°C
with 5% CO2. MG63/dox cells incubated without drugs
acted as the control group. The cells attached to the upper side of
the filter were removed with a cotton swab. The cells invaded from
the Matrigel onto the lower side of the Transwell inserts were
fixed with 100% methanol, stained with crystal violet and counted
under a light microscope.
Wound healing assay
A total of 5×105 cells were seeded into
6-well plates and incubated at 37°C for 6 h. A linear wound was
created in the cellular monolayer using a sterile pipette tip, and
then washing occurred with DMEM to remove cellular debris and yield
an acellular area in each well. The cells were incubated with for
48 h with2 µM As2O3, 200 ng/ml ADM, both or
neither. Images were captured of the wound areas and the location
in the well was noted in terms of the distance between cells.
Western blotting
Cells were harvested using Trypsin-EDTA solution
(Beyotime Institute of Biotechnology, Jiangsu, China), washed with
PBS twice (15,000 × g, 4°C, 5 min) and lysed with
radioimmunoprecipitaiton lysis buffer (Beyotime Institute of
Biotechnology, Jiangsu, China) on ice. The proteins from the lysed
cells were extracted with centrifugation (15,000 × g, 4°C, 10 min).
The protein concentrations were determined using an Evolution 60S
UV-Visible spectrophotometer (Thermo Fisher Scientific, Inc.). A
sample of 30 µg from each protein extract was separated using 8–12%
SDS-PAGE, followed by western blot analysis using the previously
described antibodies (dilution of antibodies: ERK1/2, JNK and
β-actin: 1:1,000, P-ERK and P-JNK: 1:2,000, E-cadherin and
β-catenin: 1:500). The membranes were incubated with primary
antibody overnight at 4°C and washed with PBST three times for 10
min each time. Next the membranes were incubated with secondary
antibody (Beijing CoWin Bioscience Co., Ltd.) for 1 h at room
temperature. The relative protein level in the groups was
normalized to a β-actin loading control. The gels were captured by
Image Lab 3.0 software (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The mean gray values were quantified using Gel-Pro Analyzer
4.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Lentiviral-mediated small hairpin RNA
(shRNA) silencing of stathmin
The stathmin-specific targeting sequence designed by
Shanghai Bioneer Science & Technology Co., Ltd. (Shanghai,
China) was 5′-GUGUUGGUCUUUCUAAUGU-3′. pGCSIL-GFPshSTMN with
pHelper1.0 and pHelper2.0 plasmids applied by Genomeditech Co., Ltd
(Shanghai, China) were co-transfected into HEK293T cells and
cultured overnight, then replaced with DMEM medium. At 48 h after
transfection, the supernatant containing the virus was collected by
centrifugation for 20 min at 1,000 × g. 4×107 TU/ml
lentivirus containing shRNA against STMN (Lv-shSTMN) and a negative
control (NC) virus were transfected into 2×106 MG63/dox
cells at a multiplicity of infection of 20. Cells were subcultured
at a 1:5 dilution in DMEM containing 300 µg/ml Geneticin (cat. no.
A1720, Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Positive
stable transfectants were selected and cultured for further study.
The clone in which the Lv-shSTMN vector was transfected was
designated as the Lv-shSTMN group, and the NC vector-transfected
clone was designated as the NC group.
Xenograft nude mouse model
Female, 6-week-old, immunodeficient, nude mice
(BALB/c) were bred in the laboratory animal facility of the
Children's Hospital of Suzhou University, housed individually in
micro-isolator ventilated cages with free access to water and food.
A total of 18 nude mice were randomly assigned to three groups:
Transfected (Lv-shSTMN) or NC vector-transfected cells
(5×106) suspended in 100 µl PBS were injected into the
bone marrow cavity of the left tibia of each mouse. An equal number
of age-matched animals received PBS in a volume of 100 µl were
included as controls. The resulting tumor growths were measured
with Vernier calipers on the 5, 8 and 12th and 16th day after
xenograft formation. Tumor volume was calculated according to the
following formula: Volume (mm3)=a2xb/2, where
a and b represent the shortest and longest diameters, respectively.
At the end of the experiment, the mice were sacrificed, and the
tumors were removed and examined for E-cadherin expression.
Immunohistochemical staining of
E-cadherin
The nude mouse tissue samples were fixed in 10%
formalin, embedded in paraffin and sectioned into 4-µm thick
slices. Following heat-induced antigen retrieval,
immunohistochemical staining was performed using a rabbit
monoclonal antibody against E-cadherin (dilution: 1:30; cat. no.
ab15148). All procedures were performed according to the antibody
manufacturer's protocol.
Statistical analysis
Statistical analysis was performed with SPSS
software version 17.0 (SPSS, Inc., Chicago, IL, USA). Data are
presented as the mean ± standard deviation of three independent
experiments. The statistical significance between groups was
analyzed using the Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of stathmin in human OS
cell lines
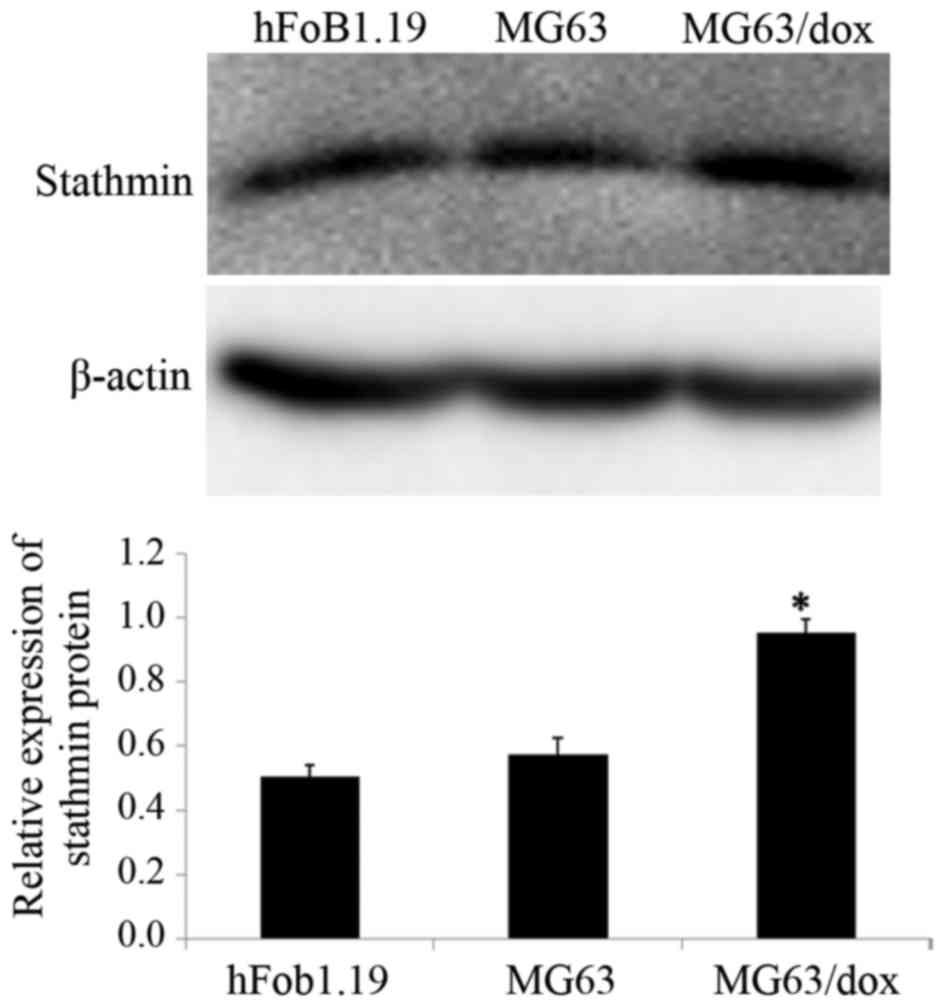
The expression of stathmin in human OSMG63 and
MG63/dox, and human osteoblast hFoB1.19 cell lines was
investigated. Western blotting results revealed that compared with
the hFoB1.19 and MG63 cells, the MG63/dox cells exhibited a higher
expression of stathmin; MG63/dox was therefore selected for the
subsequent experiments (P<0.05; Fig.
1).
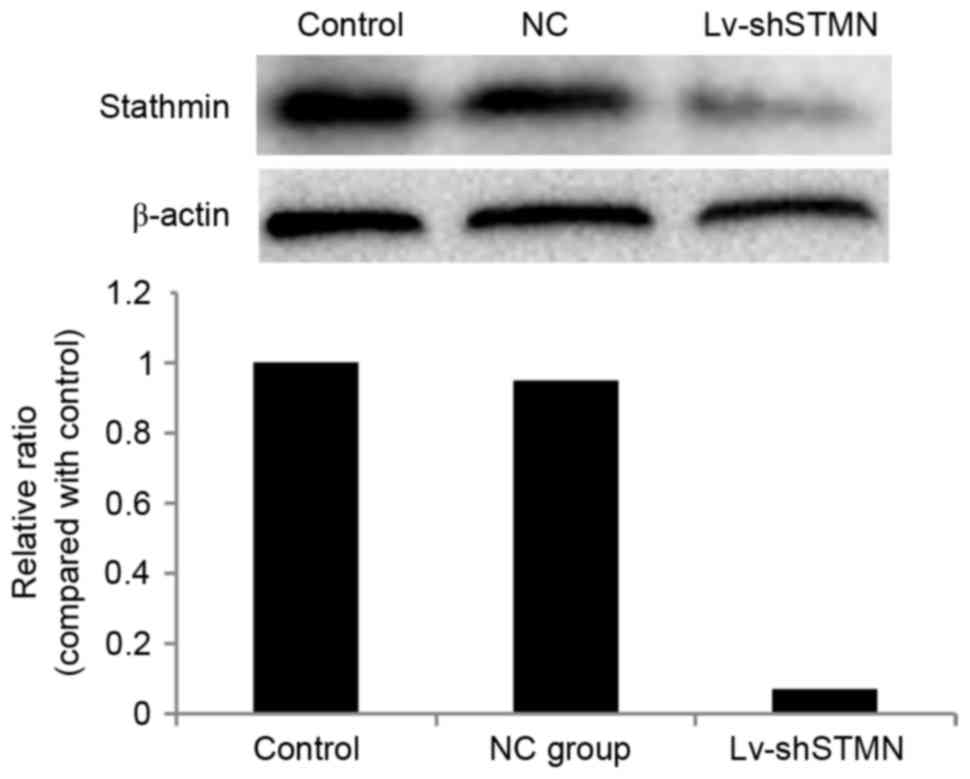
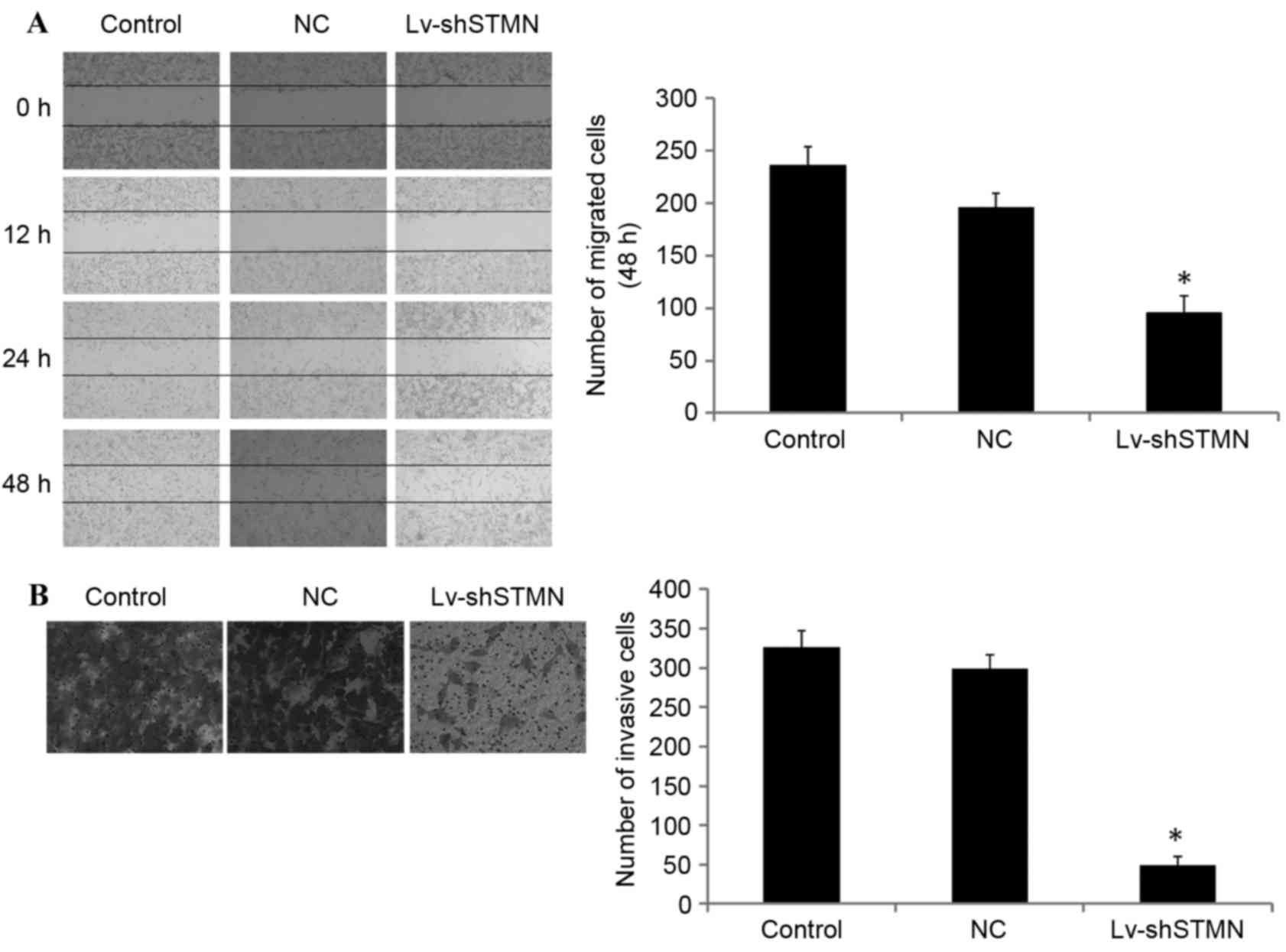
Effect of stathmin suppression on cell
invasion
Lv-shSTMN was transfected into the MG63/dox cells to
suppress the expression of stathmin (Fig.
2). The results indicated that the migration and invasion
abilities were significantly reduced by stathmin suppression in the
MG63/dox cells (P<0.05; Fig.
3).
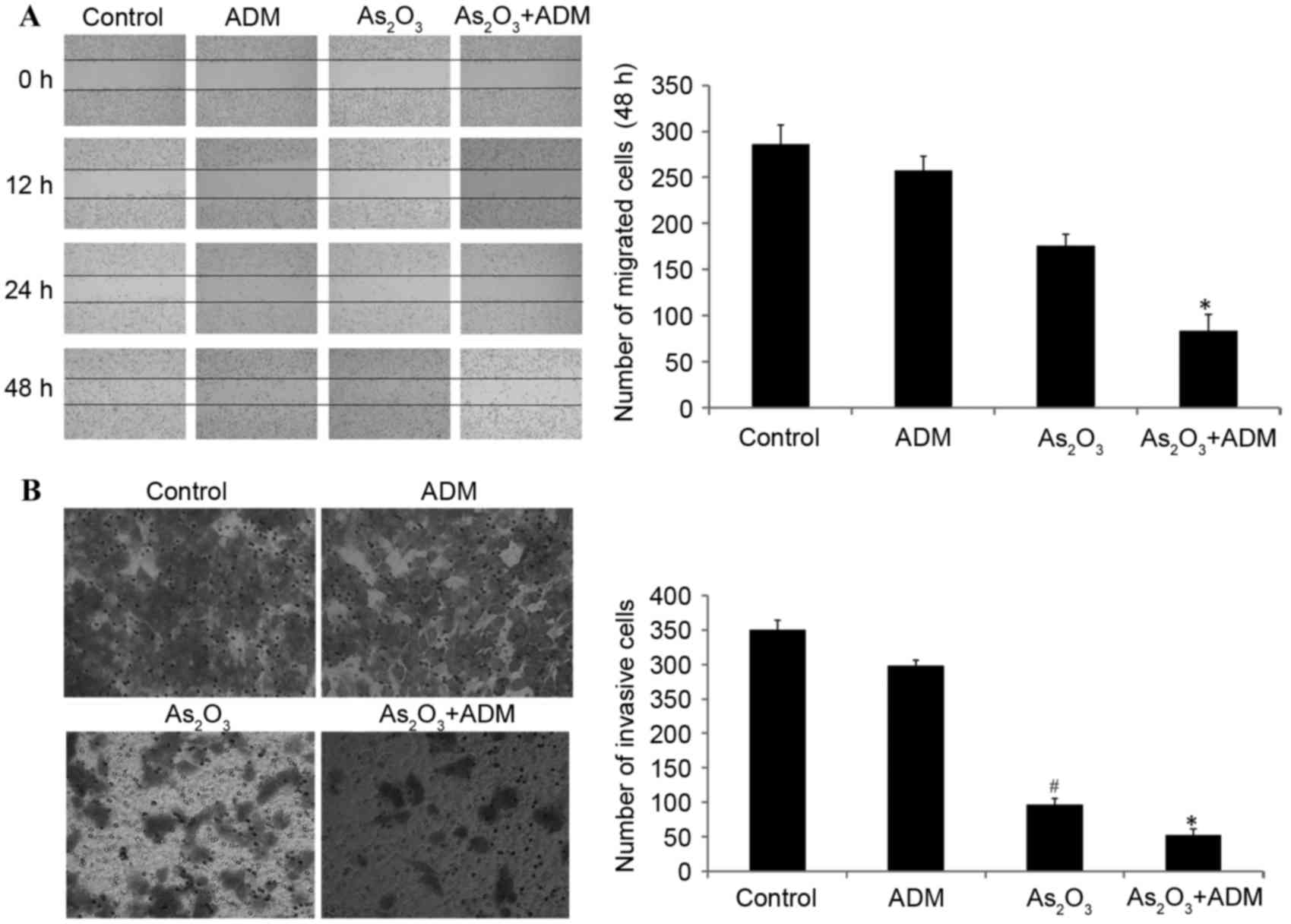
We previously reported that administration of
As2O3 with ADM suppressed stathmin expression
(6). The wound healing assay
additionally demonstrated that the administration of
As2O3 with ADM significantly inhibited cell
migration compared with the control group (P<0.05; Fig. 4A).
Transwell invasion assays were conducted to
investigate the effect of stathmin suppression on cell invasion.
The result demonstrated that As2O3 alone or
with ADM markedly inhibited tumor invasion potential compared with
the control group (P<0.05; Fig.
4B).
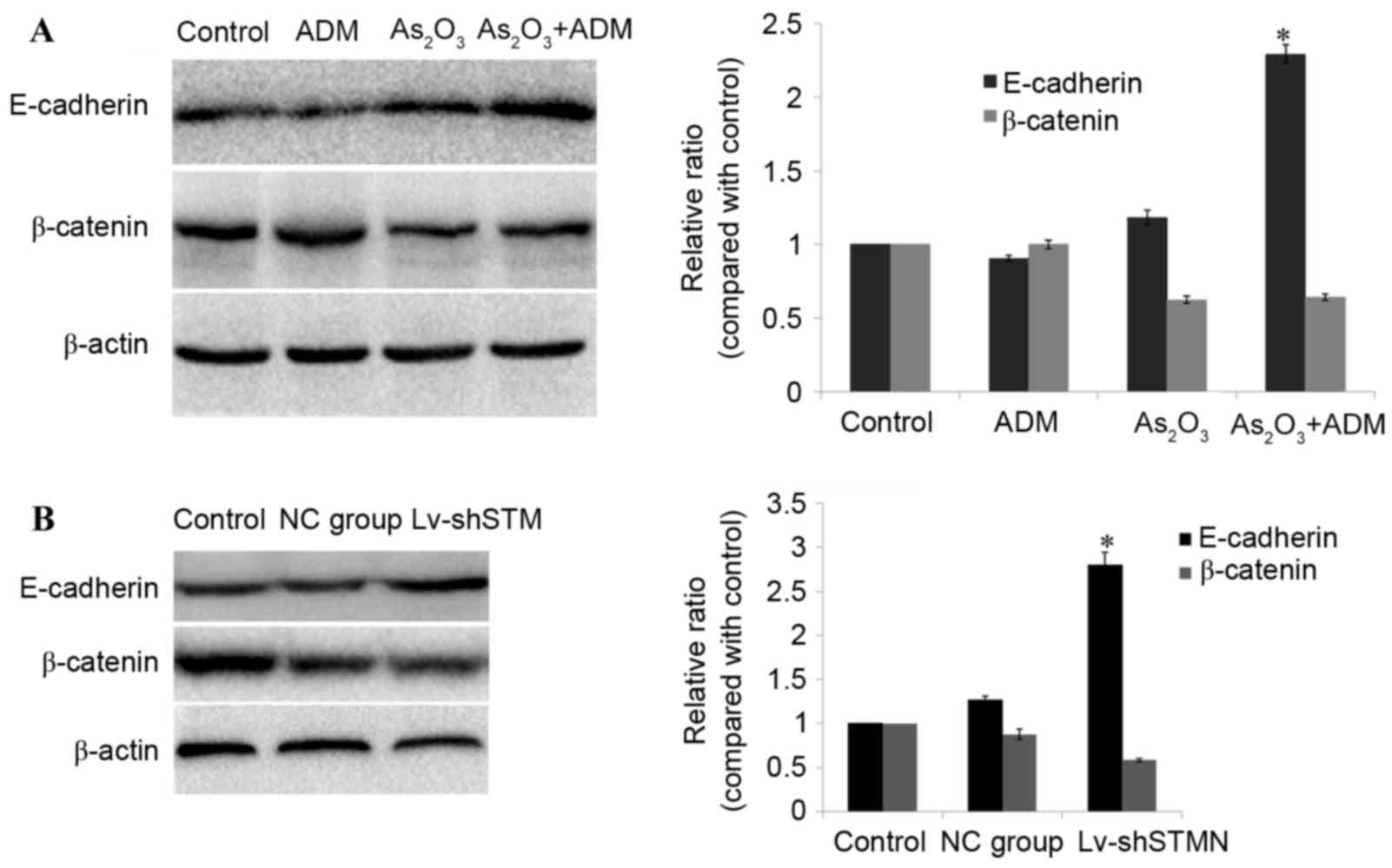
Effect of combined
As2O3 and ADM treatment on E-cadherin and
β-catenin expression levels
To investigate the molecular basis for
As2O3 efficacy, MG63/dox cells were treated
with As2O3, and the expression levels of
E-cadherin and β-catenin were analyzed by western blotting. The
results revealed that incubation with As2O3
and ADM simultaneously resulted in a significant upregulation of
E-cadherin (P<0.05), and downregulation of β-catenin in MG63/dox
cells (P=0.06; Fig. 5A).
The effect of shRNA-induced stathmin suppression on
E-cadherin expression was also detected in MG63/dox cells. As
presented in Fig. 5B, the expression
level of E-cadherin was significantly increased (P<0.05) and the
expression of β-catenin was reduced, but not significantly (P=0.08;
Fig. 5B).
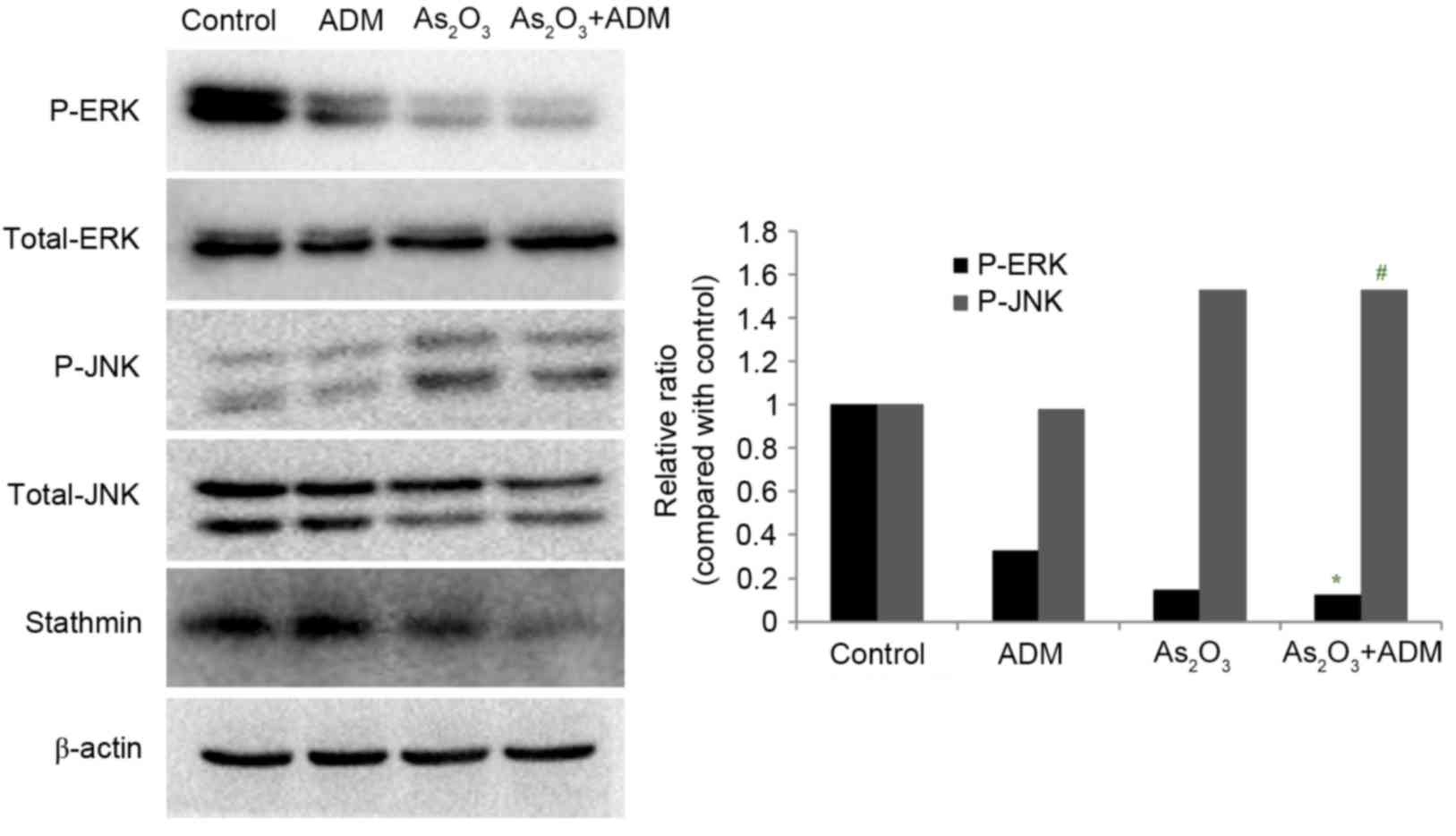
Effect of As2O3
and ADM combination treatment on JNK and ERK1/2 expression
levels
In order to evaluate the effect of
As2O3 and ADM on the MAPK pathway, MG63/dox
cells were incubated with As2O3 and ADM, and
the expression levels of P-ERK and P-JNK were determined by western
blot analysis, normalized to total ERK1/2 and JNK. The relative
abundance of P-ERK was markedly reduced (P=0.015) whereas P-JNK
expression was significantly increased (P=0.032) (Fig. 6).
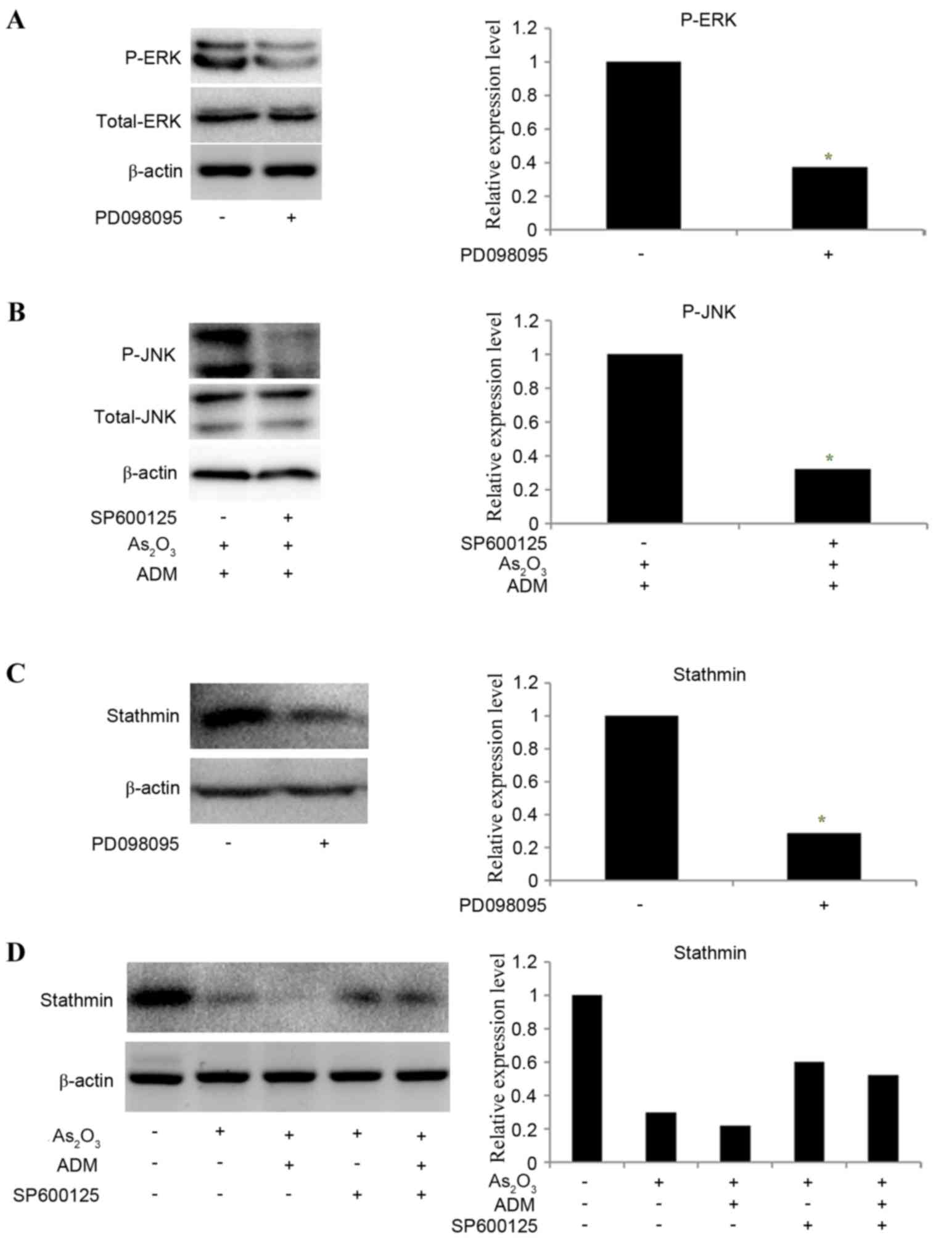
To determine whether ERK and JNK mediated the effect
of As2O3 on the expression of stathmin,
PD098095 and SP600125 were used to inhibit ERK and JNK expression,
respectively. MG63/dox cells were incubated with
As2O3 and ADM first to stimulate the JNK
phosphorylation and then treated with SP600125. The results
revealed that the relative levels of P-ERK (Fig. 7A) and P-JNK (Fig. 7B) were significantly decreased
following inhibitor treatment (P-ERK, P=0.035; P-JNK P=0.025). The
expression of stathmin was decreased following treatment
withPD098095 (P=0.02; Fig. 7C).
SP600125 partially reversed the effect of
As2O3 and ADM on stathmin expression level
(Fig. 7D). With the decline in
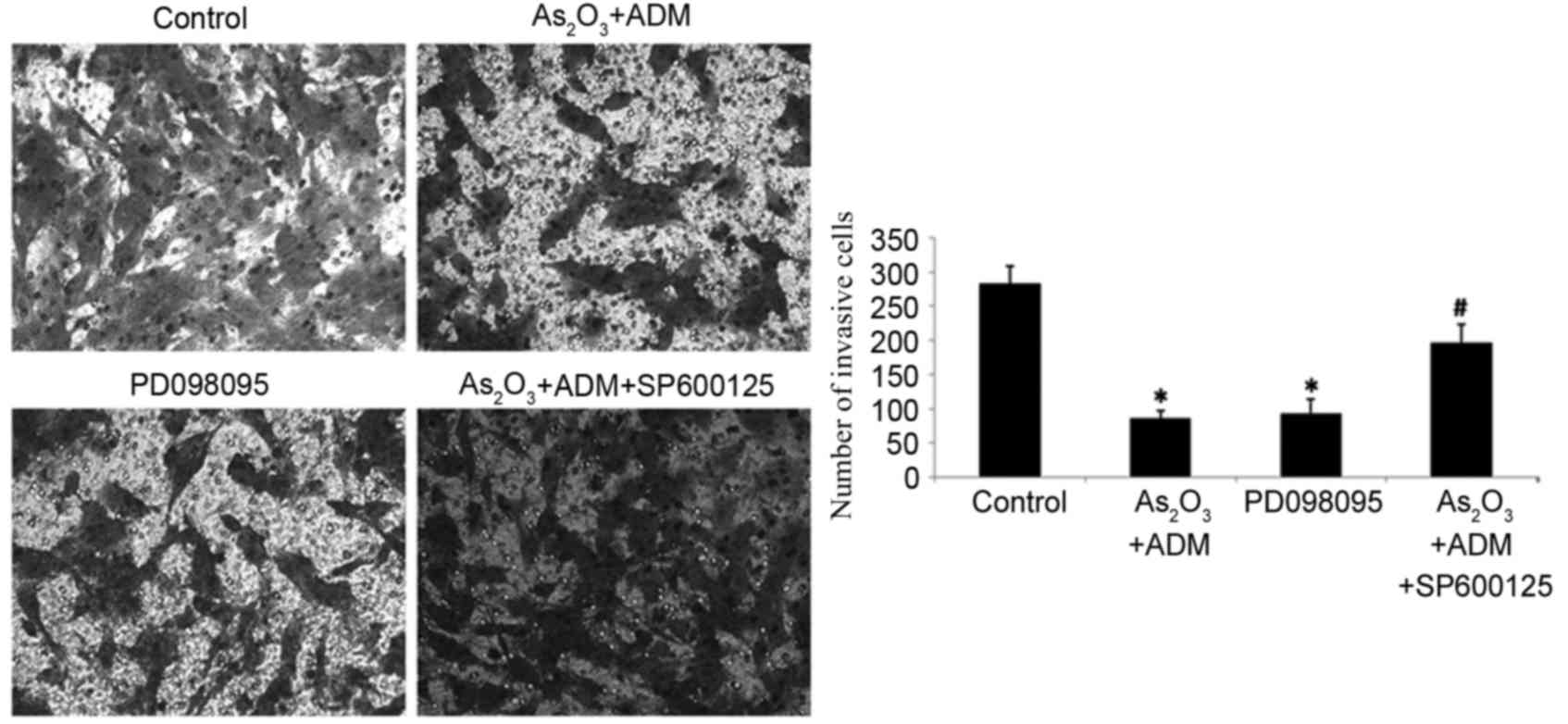
stathmin expression, PD098095 also inhibited cell invasion, as
determined by a Transwell assay, while SP600125 increased the
number of invading cells following treatment with
As2O3 and ADM (P<0.05; Fig. 8).
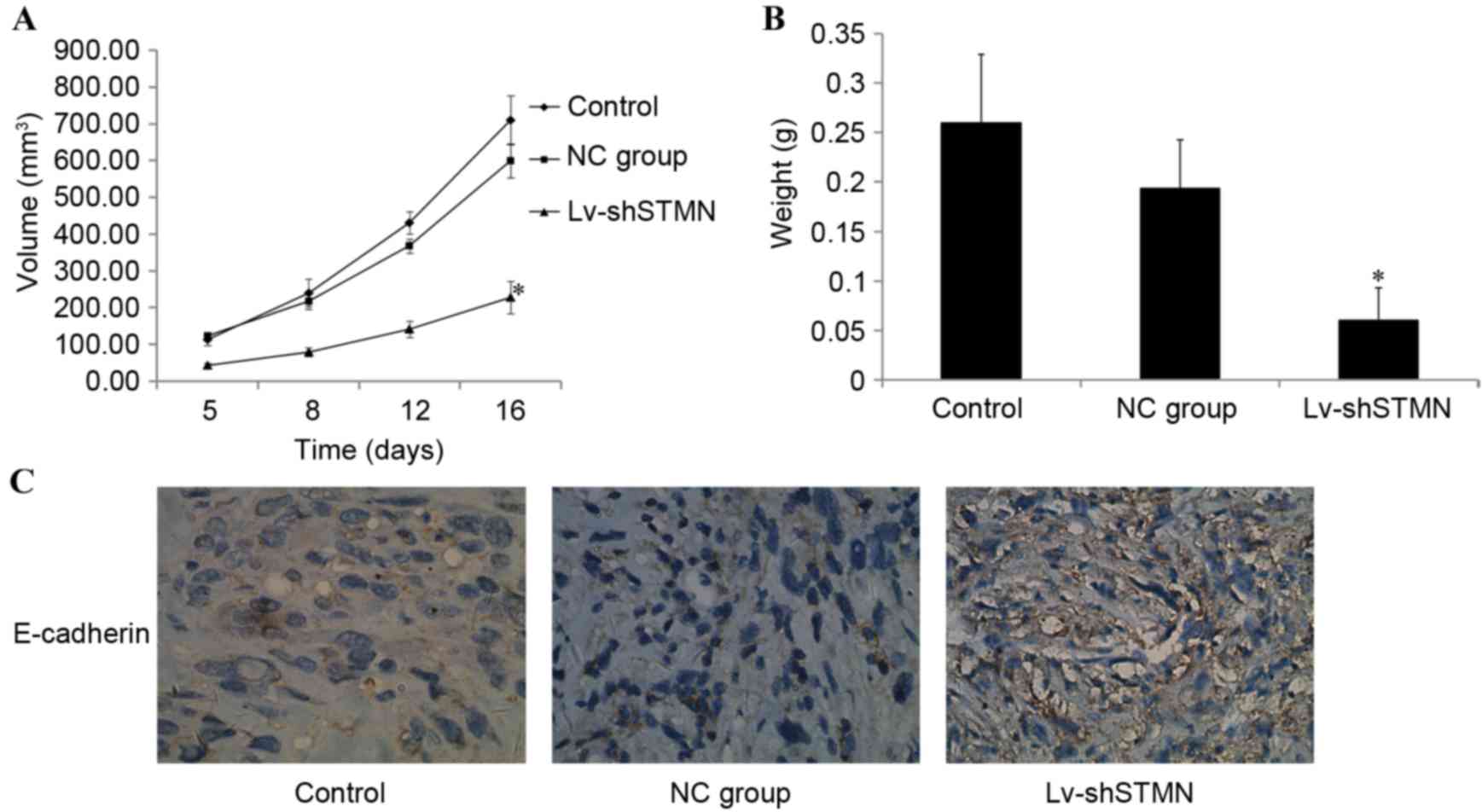
Effect of stathmin-knockdown on
xenograft tumor growth
The in vitro experiments demonstrated the inhibitory
effect of stathmin-knockdown on OS cell invasion and migration. The
effect of stathmin on MG63/dox xenograft tumor growth in vivo was
then investigated. Tumors that developed from Lv-shSTMN-transfected
cells grew more slowly when compared with the control and NC groups
(P<0.05; Fig. 9A). When the tumors
were harvested, the mean weight of the tumors in the Lv-shSTMN
group was significantly less than that in the NC and control groups
(P<0.05; Fig. 9B).
Immunohistochemistry analysis revealed that E-cadherin expression
was markedly increased in the Lv-shSTMN group (Fig. 9C).
Discussion
Cancer metastasis is a complex process that involves
several molecular mechanisms. In the present study, the expression
levels of stathmin in human OS and osteoblast cell lines were
examined. Stathmin expression was increased in the OS cells
compared with the normal cells. MG63/dox cells, which exhibit high
stathmin expression, were selected and transfected with Lv-shSTMN
to suppress stathmin. The results of the wound healing and
Transwell invasion assays demonstrated that stathmin suppression
inhibited cell migration and invasion. In order to further
elucidate the function of stathmin in OS cell invasion, we intend
to knock down stathmin expression in MG63 cells and construct
stathmin overexpressing OS cells in the future.
Types of cancer with stathmin overexpression have
been demonstrated to exhibit a more aggressive phenotype and a
worse prognosis. Baldassarre et al (11) previously observed that in sarcomas,
stathmin expression was higher in metastases than in primary
tumors. Stathmin downregulation in sarcoma-derived cell lines
inhibited migration, whereas overexpression increased FN-directed
motility. Li et al (12)
illustrated that stathmin expression promoted EMT through the
regulation of microtubule dynamics, whereas Siva1 apoptosis
inducing factor impeded cell migration and EMT by inhibiting
stathmin. Liu et al (13)
demonstrated that the knockdown of stathmin with small interfering
RNA inhibited cell migration in esophageal squamous cell carcinoma.
These results encouraged the present investigation of the effect of
stathmin on the migration and invasion of MG63/dox cells. Stathmin
expression was knocked down in MG63/dox cells using shRNA. A
Transwell invasion assay revealed that stathmin suppression reduced
the number of invasive cells. In our previous study, we
demonstrated that treatment with As2O3 and
ADM inhibited OS cell proliferation by downregulating stathmin
expression (5). The wound healing and
Transwell assays from the present study additionally demonstrated
that As2O3 combined with ADM significantly
inhibited cell migration and invasion via the inhibition of
stathmin.
As stathmin interacts with proteins from various
signaling pathways, the effect of the MAPK pathway on stathmin
function was investigated. MAPKs are serine/threonine kinase
proteins that have been identified in all eukaryotes. The MAPKs
regulate a variety of oncogenic phenotypes, including
proliferation, invasion, angiogenesis and inflammatory response
(8). Chandhanayingyong et al
(14) demonstrated the clinical
benefit of MAPK/ERK-targeted therapy for patients with unresectable
or metastatic OS. Miao et al (15) reported that the knockdown of
endogenous galanin expression in MG63 cells reduced proliferation
and invasion of the tumor cells via the suppression of MAPK/ERK
expression. The present study revealed that treatment with
As2O3 and ADM increased the phosphorylation
of JNK and decreased ERK1/2 phosphorylation, which may suggest that
As2O3 and ADM effect stathmin expression
through the MAPK pathway. In order to verify this result, PD098095
and SP600125 were used to inhibit ERK and JNK, respectively. It was
revealed that PD098095 decreased stathmin expression and the number
of invasive cells. SP600125 partially reversed the effect of
As2O3 on stathmin expression. This indicated
that As2O3 inhibited stathmin
mediated-invasion via the ERK/JNK MAPK signaling pathways. In
accordance with the in vitro results, knockdown of stathmin
inhibited the growth of OS cell in vivo, suggesting that
stathmin may present an effective target for antitumor therapy. The
regulating mechanism of the ERK/JNKMAPK signal pathways on stathmin
is to be investigated in greater detail in further studies.
E-cadherin/β-catenin signaling mediates the contact
inhibition of normal epithelial cells. The abnormal expression of
E-cadherin and β-catenin in cancer causes a decline in cell
adhesion. Cancer cells escape the normal regulatory mechanism of
contact inhibition to become more proliferative and metastasize
(16,17). Thus, E-cadherin is an important tumor
and metastasis suppressor gene. In the present study, the
expression level of E-cadherin was quantified. The result revealed
that As2O3 and ADM treatment increased
E-cadherin expression, decreasing cell invasion potential
accordingly. To simulate the growth conditions of OS, a model was
produced with the in situ vaccination of mice. Although no
attempt was made to detect metastatic foci in this model,
immunohistochemistry demonstrated that the E-cadherin expression
levels of the tumors in the stathmin group were higher than those
in the control group. This suggested the metastatic ability of the
tumors would be poor following stathmin-knockdown. The result was
verified in vitro. E-cadherin is anchored to the
cytoskeleton by catenin, to form stable connections with
neighboring cells. The knockdown of stathmin in MG63/dox cells may
have prompted β-catenin phosphorylation and proteasomal
degradation. The E-cadherin/β-catenin complex then disaggregated,
affecting the stability of cell junctions. The E-cadherin/β-catenin
pathway may act downstream of stathmin to regulate migration and
invasion (18). In a further study, a
mouse model will be constructed by intravenous injection into the
tail to allow the effect of stathmin on in vivo OS cell
invasion to be investigated in detail.
In conclusion, the present study data demonstrated
that stathmin function is associated with the migration and
invasion, and therefore, the metastasis of OS cells.
As2O3treatment decreased stathmin expression
and may have inhibited the growth and invasion of OS cells via the
ERK/JNKMAPK pathways. This observation provides an insight into the
function of stathmin in cancer progression and indicates that a
decrease in stathmin may contribute to the metastasis of OS.
Stathmin is therefore a promising biomarker and therapeutic target
for metastatic OS.
References
|
1
|
Man TK, Chintagumpala M, Visvanathan J,
Shen J, Perlaky L, Hicks J, Johnson M, Davino N, Murray J, Helman
L, et al: Expression profiles of osteosarcoma that can predict
response to chemotherapy. Cancer Res. 65:8142–8150. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim HJ, Chalmers PN and Morris CD:
Pediatric osteogenic sarcoma. Curr Opin Pediatr. 22:61–66. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hongtao L, Hui Z, Bingshun W, Xiaojin W,
Zhiyu W, Shuier Z, Aina H, Yuanjue S, Daliu M, Zan S and Yang Y:
18F-FDG positron emission tomography for the assessment of
histological response to neoadjuvant chemotherapy in osteosarcomas:
A meta-analysis. Surg Oncol. 21:e165–e170. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bachleitner-Hofmann T, Kees M and
Gisslinger H: Arsenic trioxide: Acute promyelocytic leukemia and
beyond. Leuk Lymphoma. 43:1535–1540. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miller WH Jr, Schipper HM, Lee JS, Singer
J and Waxman S: Mechanisms of action of arsenic trioxide. Cancer
Res. 62:3893–3903. 2002.PubMed/NCBI
|
|
6
|
Feng T, Qiao G, Feng L, Qi W, Huang Y, Yao
Y and Shen Z: Stathmin is key in reversion of doxorubicin
resistance by arsenic trioxide in osteosarcoma cells. Mol Med Rep.
10:2985–2992. 2014.PubMed/NCBI
|
|
7
|
Niethammer P, Bastiaens P and Karsenti E:
Stathmin-tubulin interaction gradients in motile and mitotic cells.
Science. 303:1862–1866. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu JY, Chu ZG, Han J, Dang YM, Yan H,
Zhang Q, Liang GP and Huang YS: The p38/MAPK pathway regulates
microtubule polymerization through phosphorylation of MAP4 and Op18
in hypoxic cells. Cell Mol Life Sci. 67:321–333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schmalhofer O, Brabletz S and Brabletz T:
E-cadherin, beta-catenin, and ZEB1 in malignant progression of
cancer. Cancer Metastasis Rev. 28:151–166. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen HN, Yuan K, Xie N, Wang K, Huang Z,
Chen Y, Dou Q, Wu M, Nice EC, Zhou ZG and Huang C: PDLIM1
stabilizes the E-cadherin/β-catenin complex to prevent
epithelial-mesenchymal transition and metastatic potential of
colorectal cancer cells. Cancer Res. 76:1122–1134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baldassarre G, Belletti B, Nicoloso MS,
Schiappacassi M, Vecchione A, Spessotto P, Morrione A, Canzonieri V
and Colombatti A: p27 (Kip1)-stathmin interaction influences
sarcoma cell migration and invasion. Cancer Cell. 7:51–63. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li N, Jiang P, Du W, Wu Z, Li C, Qiao M,
Yang X and Wu M: Siva1 suppresses epithelial-mesenchymal transition
and metastasis of tumor cells by inhibiting stathmin and
stabilizing microtubules. Proc Natl Acad Sci USA. 108:pp.
12851–12856. 2011; View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu F, Sun YL, Xu Y, Liu F, Wang LS and
Zhao XH: Expression and phosphorylation of stathmin correlate with
cell migration in esophageal squamous cell carcinoma. Oncol Rep.
29:419–424. 2013.PubMed/NCBI
|
|
14
|
Chandhanayingyong C, Kim Y, Staples JR,
Hahn C and Lee FY: MAPK/ERK signaling in osteosarcomas, Ewing
sarcomas and chondrosarcomas: Therapeutic implications and future
directions. Sarcoma. 2012:4048102012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Miao JH, Wang SQ, Zhang MH, Yu FB, Zhang
L, Yu ZX and Kuang Y: Knockdown of galectin-1 suppresses the growth
and invasion of osteosarcoma cells through inhibition of the
MAPK/ERK pathway. Oncol Rep. 32:1497–1504. 2014.PubMed/NCBI
|
|
16
|
Howard S, Deroo T, Fujita Y and Itasaki N:
A positive role of cadherin in Wnt/β-catenin signalling during
epithelial-mesenchymal transition. PLoS One. 6:e238992011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Faux MC, Coates JL, Kershaw NJ, Layton MJ
and Burgess AW: Independent interactions of phosphorylated
β-catenin with E-cadherin at cell-cell contacts and APC at cell
protrusions. PLoS One. 5:e141272010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Howard S, Deroo T, Fujita Y and Itasaki N:
A positive role of cadherin in Wnt/β-catenin signalling during
epithelial-mesenchymal transition. PLoS One. 6:e238992011.
View Article : Google Scholar : PubMed/NCBI
|