Introduction
Colorectal cancer (CRC) is the third most prevalent
cancer worldwide and the total number of cases in the world has
been estimated to increase from 1.36 million in 2012 to 2.4 million
in 2035 (1). The effects of
environment and lifestyle (2) have
been shown to largely affect the incidence of CRC. However, the
effects of inherited as well as acquired genetic and epigenetic
dysregulations (3–5) on the onset and development of CRC remain
controversial. Early detection and treatment may significantly
improve the survival of patients with CRC (6). Therefore, identification of new
molecular mechanisms underlying CRC tumorigenesis and progression
is pivotal for developing promising therapies.
Since at least two-thirds of all CRCs develop from
precancerous lesions with adenomatous features (7), colorectal adenomas (AD) are considered
to be precursor lesions of CRC. Fearon and Vogelstein (8) proposed that the process of
histopathological transition between AD and carcinoma (CA) in
patients with CRC was associated with a series of events that
significantly promote growth of a clonal population of CRC cells.
This multistep genetic alteration model has identified a series of
key regulatory oncogenes and tumor suppressive genes that harbor
either activation or loss of functional mutations, driving the
progression between normal colon epithelia and CRC cells (3–10). Despite
Vogelstein's model (11) and the high
risk or advanced AD concepts (12,13), there
are no absolute criteria to describe the AD-CA sequence. Classical
morphological characteristics have failed to accurately distinguish
between ADs that are potentially high risk to become malignant
disease and those that are not. Therefore, improved understanding
of cancer development may aid the characterization of ADs at high
risk for malignant progression.
Substance P (SP) was the first identified member of
the tachykinin family. It is a pro-inflammatory neuropeptide
reported as a component in extracts of horse brain and intestinal
tissues by von Euler and Gaddum (14), with functions on intestinal
contractility and blood pressure regulation. SP is an undecapeptide
synthesized by various cell types, most frequently in neurons and
inflammatory cells, including human monocytes and macrophages
(15). Neurokinin-1 receptor (NK-1R),
which is widely expressed in the human body, is the primary
receptor of SP. SP can bind to NK-1R and subsequently activate the
receptor, which results in phosphoinositide hydrolysis (16), calcium stabilization (17) and mitogen-activated protein kinase
(MAPK) activation (18,19). Numerous studies have hypothesized that
the SP/NK-1R system is involved in various cancers (20), including brain, thyroid, skin,
laryngeal, breast, gastrointestinal, pancreatic and ovarian cancers
(21–25). SP and NK-1R have been identified in
tumor cells and intra- and peri-tumoral blood vessels (24,26,27). The
activation of NK-1R was revealed to be involved in several stages
of oncological progression, including proliferation, angiogenesis
and cell metastasis (28). Kage et
al (27) revealed that NK-1R is a
G protein-coupled receptor (GPCR), and that it has two isoforms
transcribed: A full-length and truncated form, containing 407 and
311 amino acids, respectively. The truncated transcript arises from
a splice variant and does not possess the cytoplasmic C-terminal
tail (29). Additional studies were
performed to demonstrate the functional distinctions between the
two isoforms. Patel et al reported that the full-length
NK-1R (fl-NK-1R) mediated a slower tumor cell growth, while the
truncated NK-1R (tr-NK-1R) enhanced tumor cell growth and induced
the production of cytokines with growth-promoting functions
(30). A previous study by Zhou et
al (31) demonstrated that
tr-NK-1R may promote tumor progression and distant metastasis in
breast cancer.
In light of the present findings, the expression of
two isoforms of NK-1R was evaluated in the archival formalin-fixed
paraffin-embedded (FFPE) tissue of AD and CA from the same patients
that underwent radical CRC resections. The present study attempted
to differentiate between the two isoforms of NK-1R and explore
their functions in the progression of AD-CA.
Materials and methods
Patients
A total of 15 patients (9 males and 6 females; aged
69.5±12.6 years; range 45–86 years), attending the General Surgery
Department of Beijing Tong Ren Hospital (Beijing, China) for
radical colectomy or rectectomy between September 2013 and August
2014, were involved in the present study. Colorectal AD samples,
including AD, CA and PC tissue were collected from each patient.
The patients were included in the present study if the diagnoses of
the patients were reconfirmed histologically by two
gastrointestinal pathologists independently and each resected
sample contained AD, CA and PC tissue. The advantage of obtaining
all histopathological types from the same case was that variances
in patient characteristics were avoided, including genetic
background, environmental effects, lifestyle, dietary and bowel
habits, disease duration, and preoperative therapeutic
interventions.
The present study was approved by the ethics
committee of Beijing Chao-Yang Hospital, Capital Medical
University. All patients provided informed consent prior to their
inclusion in the present study. This article contains no personal
information of any patients enrolled.
Tissue preparation and analysis
RNA extraction from FFPE samples was demonstrated to
be feasible (32) and capable of
yielding similar results as frozen fixed tissue, although RNA
quantity and quality in those samples are partially decreased
(33). A total of 15 patients with
FFPE blocks from each of the three tissue areas (AD, CA and PC)
were selected. Samples were cut into 10-µm thick sections and
mounted on glass slides. RNA was extracted from the slides using
the miRNeasy FFPE kit (catalog no. 217,504; Qiagen China Co., Ltd.,
Shanghai, China), according to the manufacturer's protocol.
Purified RNA was then reverse transcribed using GoScript™ Reverse
Transcription System (catalog no. A5000; Promega Biotech Co., Ltd.,
Beijing, China).
Custom polymerase chain reaction arrays (Qiagen
China Co., Ltd.) containing primers for the full-length and
truncated TACR1 and β-actin as the control (Table I). The amplification threshold for the
samples was set at (Cq) value of ≤40. Results were calculated using
the 2−ΔΔCq method (34)
and SPSS 19.0 software (IBM SPSS, Armonk, NY, USA) was used for
statistical analysis.
 | Table I.The forward and reverse sequences for
the primers. |
Table I.
The forward and reverse sequences for
the primers.
| Primer | Primer sequence (5′
to 3′) | Size of the PCR
products (base pair) |
|---|
| NK-1R forward
(NK-1R-Fa) |
CAAGCGCAAGGTGGTCAAA | 227 |
| NK-1R-Long reverse
(NK-1R-R-Longa) |
TGCTTGAAGCCCAGACGG |
|
| NK-1R
(NK-1R-Fb) forward |
CAAGCGCAAGGTGGTCAAA | 245 |
| NK-1R-Short reverse
(NK-1R-R-Shortb) |
TGTGGCCCCTGGAGAGCT |
|
| Actin forward |
ACTTAGTTGCGTTACACCCTT | 156 |
| Actin reverse |
GTCACCTTCACCGTTCCA |
|
Immunohistochemistry
The FFPE samples from the 15 aforementioned patients
were cut into 5-µm thick sections, mounted on glass slides,
deparaffinized and rehydrated. Slides were incubated in PBS and
then probed with two primary antibodies for NK-1R: NK-1R T-20
antibody that binds to the epitope on the second extracellular loop
(dilution, 1:50; catalog no. sc-5220; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA) and NK-1R K-18 antibody (dilution, 1:50;
catalog no. sc-14116; Santa Cruz Biotechnology, Inc.) that binds to
the epitope on the C terminus. The specificity of the two
antibodies was tested by probing samples with or without blocking
peptides. Samples were then incubated with the corresponding
fluorescein-labeled secondary antibody (dilution, 1:100; catalog
no. 31509; Thermo Fisher Scientific, Inc.), mounted (VECTASHIELD
mounting medium; Vector Laboratories, Inc., Burlingame, CA, USA),
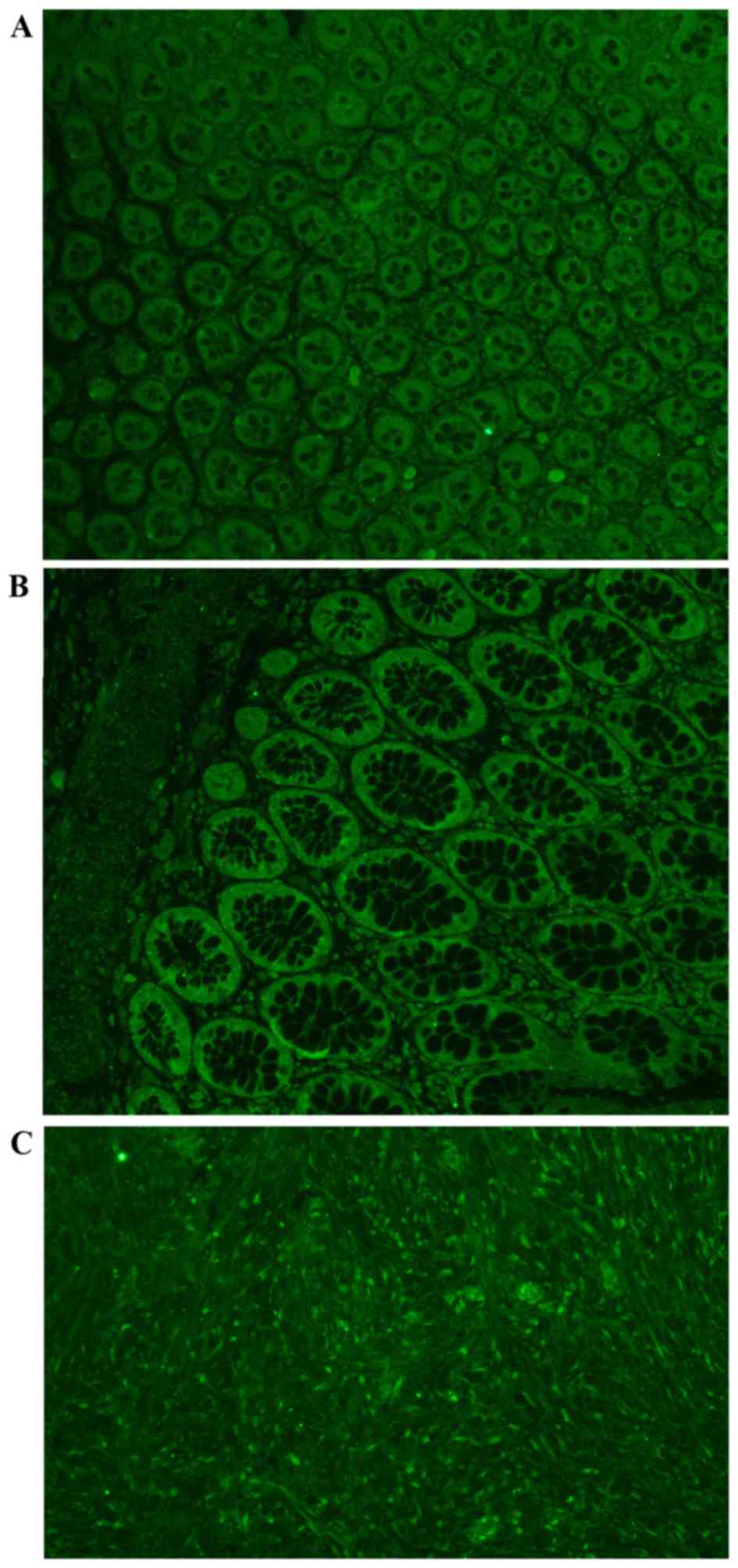
and images were captured and photographed at ×400 magnifications
(Leica AF6000 Fluorescence Microscope; Leica Microsystems, Inc.,
Buffalo Grove, IL, USA; Fig. 1). A
total of four representative fields from each slide were randomly
selected for calculation, and their mean fluorescent intensities
were quantified.
Statistical analysis
Student's t-test was used to compare TACR1
expression among different groups and normality and variance were
examined prior to the comparisons. The fluorescent intensities were
then analyzed using one-way analysis of variance and the normality
and variance were examined prior to the comparisons. Post-hoc
pairwise comparison of the means was performed using the Student
Newman Keuls test (SPSS 19.0 software; IMB SPSS).
Results
mRNA levels of tr-TACR1 were increased
in the CA groups
The mRNA expression of the tr-TACR1 transcript had a
1.7-fold increase in CA, compared with AD (P=0.026). However, the
mRNA expression of the fl-TACR1 transcript did not show a
significant difference between the AD and CA groups (P=0.438).
Protein level of truncated NK-1R was
upregulated in the AD and CA groups
Two types of antibodies that probe NK-1R protein
were applied in the analysis. NK-1R T-20 antibody binds to the
second extracellular loop so that it can probe the two isoforms.
However, the NK-1R K-18 antibody binds to the cytoplasmic
C-terminal tail and therefore only binds with fl-NK-1R. NK-1R T-20
antibody staining demonstrated an increase in fluorescence
intensity in the two groups (P=0.016). Multiple mean protein level
comparisons revealed that total NK-1R was significantly increased
in the AD and CA group compared with the PC group (P=0.026 and
P=0.007, respectively). The K-18 antibody showed no change in
fluorescence intensity among the groups (P=0.244), indicating that
the protein levels of fl-NK-1R did not change. Therefore, the
increase in total NK-1R protein in AD and CA was the result of an
increase in tr-NK-1R, rather than fl-NK-1R. The present finding
indicated that the tr-NK-1R may perform an important role in the
epithelial transition towards malignancy and provides new evidence
of AD-CA in the development and progression of colorectal
cancer.
Discussion
Colorectal ADs are commonly acknowledged as
precursor lesions of CRC, as are the ADs developed in other organs.
In 1990, Fearon and Vogelstein (8)
proposed a theory for the development and progression of AD-CA, but
the accurate progression and mechanism remains unknown. In the
present study, the expression of tr-NK-1R and fl-NK-1R was examined
in human colorectal cancer. We used the colorectal specimen from a
group of patients who underwent radical surgery for CRC in the
General Surgery Department of Beijing Tong Ren Hospital between
September 2013 and August 2014 (Beijing, China). Since each
resected specimen contained tissues in various stages of transition
between AD and CA, the two types of NK-1R mRNA and protein levels
were compared in areas of the AD with precancerous dysplasia, CA,
as well as the normal tissue adjacent to the CA. It was
demonstrated that the truncated (but not the full-length) isoform
of NK-1R was significantly increased in AD and adenocarcinoma
samples compared with the PC tissue (P=0.016). Additionally, using
RNA extraction and reverse transcription-polymerase chain reaction
(RT-PCR), a 1.7-fold increase in tr-NK-1R mRNA in CA was identified
compared with AD. Using immunohistochemistry, an increase of 30 and
40% in tr-NK-1R protein was revealed in AD and CA tissue,
respectively, compared with PC tissue (P=0.026 and P=0.007,
respectively). These outcomes suggest that protein upregulation was
not simply due to the increased transcription of tr-TACR1. Thus,
the mechanisms underlying the tr-NK-1R increase need to be further
studied. The present results indicated that tr-NK-1R may be
significant in the progression of colorectal epithelium transition
between hyperplasia and malignancy, which may provide a new basis
for understanding the mechanism of AD-CA progression.
The NK-1R was initially cloned from rat brain in an
electrophysiological study of receptor expression in Xenopus
oocytes and cross-hybridization with a known bovine NK-2R (35,36), a
clone of 3,408 nucleotides encoding the 407 residues of GPCR.
Although the degree of similarity of NK-1Rs between different
species is relatively high (94.5% identity between rat and human),
there are different key residues that are sufficient to affect the
overall interaction with antagonists (35). A splice variant of human NK-1R has an
exon 5 deletion, which yields a truncated NK-1R with 311 residues
that lacks the majority of the intracellular C-tail (37). The truncated NK-1R was identified in
human monocytes and macrophages (38), brain regions (cortex, cerebellum)
(39) and colonic epithelial cells of
colitis-associated cancer (40).
Neurokinin receptor signaling has been well studied
using rodent animal cells (41–45).
Activation of the NK-1R at the membrane triggers G protein-mediated
signaling events: Activation of phospholipase C, leading to
inositol triphosphate formation, which stabilizes intracellular
Ca2+ and eventually activates protein kinase C;
activation of adenylate cyclase, leading to stimulation of protein
kinase A; and activation of phospholipase A2 and production of
arachidonic acid. Activation of NK-1R in HEK293 cells causes a
sharp change in cell morphology, including the generation of blebs
in the plasma membrane, which includes changes in the
Rho-associated protein kinase system and phosphorylation of the
myosin regulatory light chain (46).
Signal transduction was studied in NK-1R-transfected
NCM460 human colonocytes (47).
Previous studies revealed that NK-1R activates the epidermal growth
factor receptor (EGFR) by a mechanism involving G protein-dependent
activation of members of the disintegrin and metalloproteinase
domain-containing proteases (47,48). EGFR
dimerizes, phosphorylates and assembles a SHC/Grb2 complex, which
leads to the activation of MAPK signaling (47,48). Due
to the numerous interactions among pathways, the details of
activation vary between cell types. Once activated, ERK1/2
translocates to the nucleus, inducing mitosis and preventing cell
apoptosis (19). Therefore, the
mechanism may partially mediate the ability of the NK-1R to promote
healing of the inflamed colonic epithelium (47,49).
However, it may also explain the chronic inflammation in colonic
epithelium and the development of colorectal AD initiating
carcinogenesis once the cellular balance between proliferation and
apoptosis is broken.
GPCR signaling is terminated by removing agonists
from the extracellular fluid, which restricts the capacity of the
receptor to couple to the signaling machinery. Following
stimulation with SP, subsequent responses usually fade and then
recover. G protein coupled receptor kinases (GRKs) and β-arrestins
mediate desensitization of NK-1R. GRK2, GRK3 and GRK5 can bind and
phosphorylate NK-1R (50–52). NK-1R is phosphorylated, which
subsequently promotes high-affinity interactions with β-arrestins
at the plasma membrane and in endosomes. Conversely, β-arrestins
release NK-1R from G proteins and desensitize G protein-mediated
signaling. However, the truncated NK-1R is resistant to
phosphorylation and does not interact with β-arrestins (53). This mechanism may suggest another
important reason for the termination of NK-1R signaling that the
increase of tr-NK-1R in AD leads to the transition to malignancy
and the appearance of adenocarcinoma in the present study.
Multiple observations support the involvement of SP
and NK-1R in the proliferation and tumorigenesis of colonic
epithelium. In NCM460 colonic epithelial cells, SP activates
multiple pathways that are associated with cancer cell
proliferation (47). NK-1R is
detected in SW-403 colorectal cancer cells, and NK-1R antagonist
L-733,060 impedes proliferation with or without exogenous SP,
indicating the possibility of an autocrine mechanism (54). tr-NK-1R is preferentially upregulated
in patients with colorectal AD who develop colorectal
adenocarcinoma, suggesting the functional role of the tr-NK-1R in
the histopathological transformation. The diminished
desensitization and endocytosis of tr-NK-1R may amplify its
tumorigenic potential.
Previously, the tumor microenvironment was
identified as an integral part of tumor growth and survival
(28). The microenvironment of tumors
includes any given interaction of the tumor cell with its
surroundings, including molecular and cellular structures. Thus,
the interaction of tr-NK-1R with its ligand may perform an
important role in the tumor microenvironment. tr-NK-1R, or
molecules associated with its downstream signaling pathways, may
prove to be useful as diagnostic markers in the identification of
patients with colorectal CA. Diminishing the expression or activity
of tr-NK-1R may be a potential therapeutic strategy to prevent
dysplasia of the AD from progressing to CA, or even for treating
CRC directly. Additional studies to differentiate between the
function of tr-NK-1R and fl-NK-1R in the occurrence and development
of colorectal cancer may provide new effective antitumor
therapeutic strategies in the clinic.
References
|
1
|
WCRF (2014) World Cancer Research Fund
International. http://www.wcrf.org/
|
|
2
|
Tomeo CA, Colditz GA, Willett WC,
Giovannucci E, Platz E, Rockhill B, Dart H and Hunter D: Harvard
report on cancer prevention. Volume 3: Prevention of colon cancer
in the United States. Cancer Causes Control. 10:167–180. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fearon ER: Molecular genetics of
colorectal cancer. Annu Rev Pathol. 6:479–507. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saif MW and Chu E: Biology of colorectal
cancer. Cancer J. 16:196–201. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Issa JP: Colon cancer: It's CIN or CIMP.
Clin Cancer Res. 14:5939–5940. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
UEG (2014) Colorectal cancer in Europe.
http://www.ueg.eu/press/crceurope/
|
|
7
|
Peipins LA and Sandler RS: Epidemiology of
colorectal adenomas. Epidemiol Rev. 16:273–297. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fearon ER and Vogelstein B: A genetic
model for colorectal tumorigenesis. Cell. 61:759–767. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vogelstein B and Kinzler KW: Cancer genes
and the pathways they control. Nat Med. 10:789–799. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wood LD, Parsons DW, Jones S, Lin J,
Sjöblom T, Leary RJ, Shen D, Boca SM, Barber T, Ptak J, et al: The
gnomic landscapes of human breast and colorectal cancers. Science.
318:1108–1113. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bhalla A, Zulfiqar M, Weindel M and
Shidham VB: Molecular diagnostics in colorectal carcinoma. Clin Lab
Med. 33:835–859. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chung SJ, Kim YS, Yang SY, Song JH, Kim D,
Park MJ, Kim SG, Song IS and Kim JS: Five-year risk for advanced
colorectal neoplasia after initial colonoscopy according to the
baseline risk stratification: A prospective study in 2452
asymptomatic Koreans. Gut. 60:1537–1543. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nusko G, Hahn EG and Mansmann U: Risk of
advanced metachronous colorectal adenoma during long-term
follow-up. Int J Colorectal Dis. 23:1065–1071. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Euler VUS and Gaddum JH: An unidentified
depressor substance in certain tissue extracts. J Physiol.
72:74–87. 1931. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ho WZ, Lai JP, Zhu XH, Uvaydova M and
Douglas SD: Human monocytes and macrophages express substance P and
neurokinin-1 receptor. J Immunol. 159:5654–5660. 1997.PubMed/NCBI
|
|
16
|
Rollandy I, Dreux C, Imhoff V and
Rossignol B: Importance of the presence of the N-terminal
tripeptide of substance P for the stimulation of
phosphatidylinositol metabolism in rat parotid gland: A possible
activation of phospholipases C and D. Neuropeptides. 13:175–185.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pradier L, Heuillet E, Hubert JP, Laville
M, Le Guern S and Doble A: Substance P-evoked calcium mobilization
and ionic current activation in the human astrocytoma cell line U
373 MG: Pharmacological characterization. J Neurochem.
61:1850–1858. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luo W, Sharif TR and Sharif M: Substance
P-induced mitogenesis in human astrocytoma cells correlates with
activation of the mitogen-activated protein kinase signaling
pathway. Cancer Res. 56:4983–4991. 1996.PubMed/NCBI
|
|
19
|
DeFea KA, Zalevsky J, Thoma MS, Déry O,
Mullins RD and Bunnett NW: beta-arrestin-dependent endocytosis of
proteinase-activated receptor 2 is required for intracellular
targeting of activated ERK1/2. J Cell Biol. 148:1267–1281. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Muñoz M, Rosso M and Coveñas R: The NK-1
receptor: A new target in cancer therapy. Curr Drug Targets.
12:909–921. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Muñoz M, Rosso M, Robles-Frias MJ,
Salinas-Martín MV, Rosso R, González-Ortega A and Coveñas R: The
NK-1 receptor is expressed in human melanoma and is involved in the
antitumor action of the NK-1 receptor antagonist aprepitant on
melanoma cell lines. Lab Invest. 90:1259–1269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Esteban F, Gonzalez-Moles MA, Castro D,
Martin-Jaen Mdel M, Redondo M, Ruiz-Avila I and Muñoz M: Expression
of substance P and neurokinin-1-receptor in laryngeal cancer:
Linking chronic inflammation to cancer promotion and progression.
Histopathology. 54:258–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rosso M, Robles-Frías MJ, Coveñas R,
Salinas-Martín MV and Muñoz M: The NK-1 receptor is expressed in
human primary gastric and colon adenocarcinomas and is involved in
the antitumor action of L-733,060 and the mitogenic action of
substance P on human gastrointestinal cancer cell lines. Tumour
Biol. 29:245–254. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hennig IM, Laissue JA, Horisberger U and
Reubi JC: Substance-P receptors in human primary neoplasms: Tumoral
and vascular localization. Int J Cancer. 61:786–792. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schulz S, Stumm R, Röcken C, Mawrin C and
Schulz S: Immunolocalization of full-length NK1 tachykinin
receptors in human tumors. J Histochem Cytochem. 54:1015–1020.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Muñoz M, Rosso M and Coveñas R: A new
frontier in the treatment of cancer: NK-1 receptor antagonists.
Curr Med Chem. 17:504–516. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kage R, Leeman SE and Boyd ND: Biochemical
characterization of two different forms of the substance P receptor
in rat submaxillary gland. J Neurochem. 60:347–351. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rosso M, Muñoz M and Berger M: The role of
neurokinin-1 receptor in the microenvironment of inflammation and
cancer. ScientificWorldJournal. 2012:3814342012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fong TM, Anderson SA, Yu H, Huang RR and
Strader CD: Differential activation of intracellular effector by
two isoforms of human neurokinin-1 receptor. Mol Pharmacol.
41:24–30. 1992.PubMed/NCBI
|
|
30
|
Patel HJ, Ramkissoon SH, Patel PS and
Rameshwar P: Transformation of breast cells by truncated
neurokinin-1 receptor is secondary to activation by
preprotachykinin-A peptides. Proc Natl Acad Sci USA.
102:17436–17441. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou Y, Zhao L, Xiong T, Chen X, Zhang Y,
Yu M, Yang J and Yao Z: Roles of full-length and truncated
neurokinin-1 receptors on tumor progression and distant metastasis
in human breast cancer. Breast Cancer Res Treat. 140:49–61. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pagedar NA, Wang W, Chen DH, Davis RR,
Lopez I, Wright CG and Alagramam KN: Gene expression analysis of
distinct populations of cells isolated from mouse and human inner
ear FFPE tissue using laser capture microdissection-a technical
report based on preliminary findings. Brain Res. 1091:289–299.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nonn L, Vaishnav A, Gallagher L and Gann
PH: mRNA and micro-RNA expression analysis in laser-capture
microdissected prostate biopsies: Valuable tool for risk assessment
and prevention trials. Exp Mol Pathol. 88:45–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yokota Y, Sasai Y, Tanaka K, Fujiwara T,
Tsuchida K, Shigemoto R, Kakizuka A, Ohkubo H and Nakanishi S:
Molecular characterization of a functional cDNA for rat substance P
receptor. J Biol Chem. 264:17649–17652. 1989.PubMed/NCBI
|
|
36
|
Masu Y, Nakayama K, Tamaki H, Harada Y,
Kuno M and Nakanishi S: cDNA cloning of bovine substance-K receptor
through oocyte expression system. Nature. 329:836–838. 1987.
View Article : Google Scholar
|
|
37
|
Baker SJ, Morris JL and Gibbins IL:
Cloning of a C-terminally truncated NK-1 receptor from guinea-pig
nervous system. Brain Res. 111:136–147. 2003. View Article : Google Scholar
|
|
38
|
Lai JP, Ho WZ, Kilpatrick LE, Wang X,
Tuluc F, Korchak HM and Douglas SD: Full-length and truncated
neurokinin-1 receptor expression and function during
monocyte/macrophage differentiation. Proc Natl Acad Sci USA.
103:7771–7776. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lai JP, Cnaan A, Zhao H and Douglas SD:
Detection of full-length and truncated neurokinin-1 receptor mRNA
expression in human brain regions. J Neurosci Methods. 168:127–133.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gillespie E, Leeman SE, Watts LA, Coukos
JA, O'Brien MJ, Cerda SR, Farraye FA, Stucchi AF and Becker JM:
Truncated neurokinin-1 receptor is increased in colonic epithelial
cells from patients with colitis-associated cancer. Proc Natl Acad
Sci USA. 108:17420–17425. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Krause JE, Chirgwin JM, Carter MS, Xu ZS
and Hershey AD: Three rat preprotachykinin mRNAs encode the
neuropeptides substance P and neurokinin A. Proc Natl Acad Sci USA.
84:881–885. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shigemoto R, Yokota Y, Tsuchida K and
Nakanishi S: Cloning and expression of a rat neuromedin K receptor
cDNA. J Biol Chem. 265:623–628. 1990.PubMed/NCBI
|
|
43
|
Bowden JJ, Garland AM, Baluk P, Lefevre P,
Grady EF, Vigna SR, Bunnett NW and McDonald DM: Direct observation
of substance P-induced internalization of neurokinin 1 (NK1)
receptors at sites of inflammation. Proc Natl Acad Sci USA.
91:8964–8968. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bradesi S, Svensson CI, Steinauer J,
Pothoulakis C, Yaksh TL and Mayer EA: Role of spinal microglia in
visceral hyperalgesia and NK1R up-regulation in a rat model of
chronic stress. Gastroenterology. 136(1339–1348): e1–e2. 2009.
|
|
45
|
Steinhoff MS, von Mentzer B, Geppetti P,
Pothoulakis C and Bunnett NW: Tachykinins and their receptors:
Contributions to physiological control and the mechanisms of
disease. Physiol Rev. 94:265–301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Meshki J, Douglas SD, Lai JP, Schwartz L,
Kilpatrick LE and Tuluc F: Neurokinin 1 receptor mediates membrane
blebbing in HEK293 cells through a Rho/Rho-associated coiled-coil
kinase-dependent mechanism. J Biol Chem. 284:9280–9289. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Koon HW, Zhao D, Na X, Moyer MP and
Pothoulakis C: Metalloproteinases and transforming growth
factor-alpha mediate substance P-induced mitogen-activated protein
kinase activation and proliferation in human colonocytes. J Biol
Chem. 279:45519–45527. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Castagliuolo I, Valenick L, Liu J and
Pothoulakis C: Epidermal growth factor receptor transactivation
mediates substance P-induced mitogenic responses in U-373 MG cells.
J Biol Chem. 275:26545–26550. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Castagliuolo I, Morteau O, Keates AC,
Valenick L, Wang CC, Zacks J, Lu B, Gerard NP and Pothoulakis C:
Protective effects of neurokinin-1 receptor during colitis in mice:
Role of the epidermal growth factor receptor. Br J Pharmacol.
136:271–279. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Barak LS, Warabi K, Feng X, Caron MG and
Kwatra MM: Real-time visualization of the cellular redistribution
of G protein-coupled receptor kinase 2 and beta-arrestin 2 during
homologous desensitization of the substance P receptor. J Biol
Chem. 274:7565–7569. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jorgensen R, Holliday ND, Hansen JL, Vrecl
M, Heding A, Schwartz TW and Elling CE: Characterization of
G-protein coupled receptor kinase interaction with the neurokinin-1
receptor using bioluminescence resonance energy transfer. Mol
Pharmacol. 73:349–358. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kwatra MM, Schwinn DA, Schreurs J, Blank
JL, Kim CM, Benovic JL, Krause JE, Caron MG and Lefkowitz RJ: The
substance P receptor, which couples to Gq/11, is a substrate of
beta-adrenergic receptor kinase 1 and 2. J Biol Chem.
268:9161–9164. 1993.PubMed/NCBI
|
|
53
|
Li H, Leeman SE, Slack BE, Hauser G,
Saltsman WS, Krause JE, Blusztajn JK and Boyd ND: A substance P
(neurokinin-1) receptor mutant carboxyl-terminally truncated to
resemblea naturally occurring receptor isoform displays enhanced
responsiveness and resistance to desensitization. Proc Natl Acad
Sci USA. 94:9475–9480. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Rosso M, Robles-Frias MJ, Coveñas R,
Salinas-Martin MV and Muñoz M: The NK-1 receptor is expressed in
human primary gastric and colon adenocarcinomas and is involved in
the antitumor action of L-733,060 and the mitogenic action of
substance P on human gastrointestinal cancer cell lines. Tumour
Biol. 29:245–254. 2008. View Article : Google Scholar : PubMed/NCBI
|