Introduction
Acute lymphoblastic leukemia (ALL) is the most
common cancer malignancy and the leading cause of cancer-associated
mortality in children (1). Assessment
of the risk of relapse is crucial in modern treatment protocols, as
relapses are the main cause of therapeutic failure in childhood
ALL. Despite investigation of the clinical and cytogenetic
predictors, there remains no sufficient marker enabling early
identification of patients at high risk of relapse. Therefore, the
molecular markers for risk assessment at the time of diagnosis and
during the treatment course are urgently needed.
A particularly poor treatment outcome is observed in
patients with B-cell precursor ALL (BCP-ALL) with t(9;22)(q34;q11)
translocation, and in patients with breakpoint cluster region/ABL
proto-oncogene 1, non-receptor tyrosine kinase
(BCR/ABL1)-like ALL subtypes (2). BCR/ABL1-like ALL exhibits a gene
expression profile similar to BCR/ABL1-positive ALL, but
without t(9;22) rearrangement (3).
These leukemia subtypes are most commonly characterized by genetic
abnormalities affecting the following pathways: Lymphoid
development [early B-cell factor 1 (EBF1), ETS variant
6(ETV6), IKAROS family zinc finger (IKZF)1, LIM
domain only 2 and paired box 5 (PAX5)]; signal transduction
(ABL1, ABL2, cytokine receptor-like factor 2, colony
stimulating factor 1 receptor, erythropoietin receptor, interleukin
7 receptor, janus kinase 2, platelet derived growth factor receptor
β); and tumor suppression and cell cycle regulation [cyclin
dependent kinase inhibitor (CDKN)2A/CDKN2B,
retinoblastoma 1 (RB1) and tumor protein 53] (4).
Microdeletions and point mutations of the
aforementioned genes are considered important factors in
lymphoblast differentiation and leukemia progression, but the final
pathogenetic effect is controlled by post-transcriptional
regulators (5–7). Small noncoding RNA molecules (~22 base
pairs), referred to as microRNA (miRNA/miR), are types of these
regulators. miRNAs recognize a seed region in an mRNA sequence and
destabilize mRNA by acting in a RNA-induced silencing complex,
thereby inhibiting protein production (8). Evidence from previous studies indicates
that miRNAs promote neoplastic transformation. In lymphoid cells,
miRNA may serve a role in hematopoiesis, and in the development of
leukemia by the suppression of tumor suppressor genes or
stimulation of oncogenes (9,10). Dysregulation of miRNA expression is
observed in leukemic cells and is potentially linked to drug
resistance and poor outcome (11,12).
As aforementioned, miRNAs are involved in numerous
processes at the transcription level, therefore their signature may
reflect normal homeostatic processes as well as pathological
changes and leukemogenic disturbance. This suggests that miRNAs are
potential biomarkers for early leukemia detection.
The present study aimed to identify miRNAs whose
expression level variations may be associated with known molecular
defects in childhood ALL. To preselect the miRNAs, bioinformatics
tools were used for in silico miRNA analysis, such as
Mircancer.ecu.edu, miRWalk data base, miRTarBase,
and microRNA.org (13).
Based on target prediction, a set of potential miRNA markers:
miR-24, miR-31, miR-128, miR-542 and miR-708were identified, and
then their expression levels were evaluated in a cohort of children
with ALL and their association with known prognostic genetic
lesions and clinical features of ALL.
Materials and methods
miRNA profiles
For the analysis of the miRNA profiles, 5 miRNAs
(miR-128, miR-542, miR-708, miR-24 and miR-31) were selected using
bioinformatics tools and databases [miRCancer (14), miRWalk database (15), miRTarBase (16), microRNA.org
(17)], and were selected using
literature reviews (9–12). The mirSVR score was applied for
ranking microRNA target sites by a downregulation score. The mirSVR
is machine learning method based on regression modelling and was
calculated based on seed-site pairing, site context, conservation
and free-energy of the selected miRNAs (18).
Recruitment of patients
Children (<18 years old) with B-cell lineage ALL
(B-ALL) and T-cell lineage (T-ALL) were recruited from the
Caucasian Polish population at the Department of Pediatrics,
Oncology, Hematology and Diabetology, Medical University of Łódź
(Łódź, Poland), and cooperating Centres of Pediatric Oncohematology
in the cities of Bialystok, Bydgoszcz, Gdansk, Katowice, Kielce,
Olsztyn, Poznan, Szczecin, and Zabrze (Poland). Ethics committee
approval was obtained from the Institutional Review Board of the
Medical University of Lodz (number, RNN/226/11/KE//KE). All
patients were diagnosed between May 2004 and March 2014 and treated
according to ALL-IC 2002 or 2009 protocols (19). Patients with a set of microdeletions,
particularly IKZF1 defects, were preferentially enrolled
into the study.
Minimal residual disease (MRD)
monitoring
The bone marrow samples from patients with BCP-ALL
were collected at three time points: At diagnosis and at 15 and 33
days of treatment protocol. The bone marrow samples from patients
with BCP-ALL were analyzed with 8-color flow cytometry according to
the protocols of the EuroFlow Consortium (20). These samples were processed according
to the manufacturer's lyse and wash protocol (BD Biosciences, San
Jose, CA, USA), with 1X FACS Lyse used for erythrocyte lysis (BD
Biosciences) and analyzed with FACSCanto II flow cytometer (BD
Biosciences). To ensure a sensitivity level of ≥104,
low-cellular samples were first subjected to erythrocyte lysis in
1X ammonium chloride solution (Pharm Lyse; BD Biosciences) in a
bulk lysis protocol. Subsequently, the suspension of leukocytes was
stained in a single 8-color tube with antibodies adequate for the
blasts'phenotype at diagnosis. The reproducibility of the obtained
results was ensured by complying with the standard operating
procedures developed by EuroFlow based on daily quality assessment
with fluorescent beads (Sphero Rainbow Calibration Particles;
Spherotech, Lake Forest, IL, USA). For data analysis, FACSDiva 6.1
software (BD Biosciences) was used. Patients with number of blasts
in peripheral blood >1,000 on the 8th day following steroid
administration were identified as steroid resistant.
DNA and RNA extraction
A total of 90 bone marrow samples were collected at
the time of diagnosis (B-ALL, n=66; T-ALL, n=24). For DNA and RNA
isolation, 300 µl bone marrow stored in TRIzol® reagent
was used (Ambion; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The isolation procedure was performed according to the
manufacturer's protocol.
Reverse transcription and
expression
The RNA samples, isolated from bone marrow, were
used for miRNA expression analysis, namely reverse transcription
(RT) of miRNA to complementary (c)DNA, amplification of cDNA and
detection by TaqMan probes in quantitative polymerase chain
reaction (qPCR). Each RT reaction was performed according to the
manufacturer's protocol (TaqMan® MicroRNA Reverse
Transcription kit; Thermo Fisher Scientific, Inc.) in 15 µl [7 µl
mix, 3 µl primer (5X RT), 5 µl RNA sample]. The sequences of
Stem-Loop RT primer target sequences and miRNA assay IDs are
described in Table I. The amount of
RNA used for each reaction was 40 ng/well (RNA concentration, 8
ng/µl). RT-PCR was conducted under the following conditions: 30 min
at 16°C, 30 min at 42°C, 5 min 85°C and hold at 4°C.
 | Table I.miRNA assay identities and target
sequences. |
Table I.
miRNA assay identities and target
sequences.
| miRNA | Assay identity | Target
sequence |
|---|
| hsa-miR-31 | TM:002279 |
5′-AGGCAAGAUGCUGGCAUAGCU-3′ |
| hsa-miR-24–2 | TM:002441 |
5′-UGCCUACUGAGCUGAAACACAG-3′ |
| hsa-miR-542-5p | TM:002240 |
5′-UCGGGGAUCAUCAUGUCACGAGA-3′ |
| hsa-miR-708 | TM:002341 |
5′-AAGGAGCUUACAAUCUAGCUGGG-3′ |
| hsa-miR-128A | TM:002216 |
5′-UCACAGUGAACCGGUCUCUUU-3′ |
| U6 snRNA | TM:001973 |
5′-GTGCTCGCTTCGGCAGCACATATACTAAAATTGGAACGATACAGAGAAGATTAGCATGGCCCCTGCGCAAGGATGACACGCAAATTCGTGAAGCGTTCCATATTTT-3′ |
Each qPCR reaction was performed in duplicate, in 18
µl volumes (TaqMan® Gene Expression Master Mix 10 µl;
20X assay 1 µl; H2O 4 µl) with 3 µl of cDNA sample (8
ng). TaqMan® microRNA assays (Thermo Fisher Scientific,
Inc.) were used to analyze specific miRNAs (hsa-miR-31, TM:002279;
hsa-miR-24-2, TM:002441; hsa-miR-542-5p, TM:002240; hsa-miR-708,
TM:002341; hsa-miR-128a, TM:002216). For normalization of PCR, as
an internal control, U6 snRNA (TM:001973) was used. PCRs were
conducted in duplicate under the following protocol: 2 min at 50°C,
10 min at 95°C, then 40 cycles of: 95°C for 30 sec and 60°C for 1
min.
The expression levels are presented as
2−∆∆Cq values, where ∆Cq = Cq (reference) - Cq (miRNA of
interest), which produces a higher value for higher miRNA
expression, facilitating its use and interpretation as a biomarker
(21).
Multiplex ligation-dependent probe
amplification (MLPA)
Copy number variations (CNVs) were identified by
MLPA. Samples were screened for selected CNVs using P202-B1 and
P335-B1 SALSA MLPA kits (MRC-Holland, Amsterdam, The Netherlands).
These assays enable analysis of all exons of CDKN2A/B and
IKZF1 genes, and selected exons of PAX5, BTG
anti-proliferation factor 1 (BTG1), EBF1,
ETV6, RB1, immunoglobulin heavy locus, IKZF2,
IKZF3, metastasis associated 1 genes and the pseudoautosomal
region 1 (PAR1) region. The MLPA procedure was performed
according to the manufacturer's protocols.
Statistical analysis
Statistical analysis was performed using Statistical
12.5PL software (StatSoft, Inc., Tulsa, OK, USA). The distribution
of variables was tested with the Shapiro-Wilk test and
Kolmogorov-Smirnov test with Lilliefors correction. Due to the
non-normal distribution of all analyzed variables, non-parametric
tests were used and the results are presented as medians followed
by interquartile ranges (IQRs). P<0.05 was considered to
indicate a statistically significant difference. Categorical
variables were compared using the χ2 test or Fisher's
exact test. Continuous variables were compared using the U
Mann-Whitney test to analyze differences between two groups and the
Kruskal-Wallis along with post hoc (Dunn's) test to analyze
differences between more than two groups. The R Spearman's
correlation was used to analyze correlations between continuous
variables.
Results
Study group characteristics
A total of 90 children were recruited, of which 24
(26.67%) were diagnosed with T-ALL and 66 (73.33%) were diagnosed
with B-ALL (Table II). In the B-ALL
group, 2 patients (3.23%) exhibited a BCR/ABL fusion, 1
(1.67%) exhibited a mixed lineage leukemia (MLL)/AF4
fusion and 11 (19.64%) exhibited ETV6/runt related
transcription factor 1 (RUNX1) fusions. BCR/ABL,
MLL/AF4 and ETV6/RUNX1 fusions were not identified
among the patients with T-ALL.
| Table II.Clinical characteristics of the study
population. |
Table II.
Clinical characteristics of the study
population.
| Variable | Total | T-ALL | B-ALL | P-value |
|---|
| Total | 90 | 24 | 66 |
|
| Sex |
|
|
| 0.16519 |
| Male, n
(%) | 59 (65.56) | 19 (79.17) | 40 (60.61) |
|
| Female,
n (%) | 31 (34.44) | 5 (20.83) | 26 (39.39) |
|
| Age at diagnosis,
years(IQR) | 8.36
(3.62–12.88) | 8.87
(3.22–13.05) | 8.05
(3.79–12.73) | 0.98908 |
| WBC,
×103/µl (IQR) | 26.65
(6.36–88.10) | 108.85
(29.74–279.01) | 14.13
(4.45–53.00) | 0.00008 |
| MRD at day 15, %
(IQR) | 1.06
(0.13–9.50) | 4.2
(0.83–29.30) | 0.61
(0.07–5.25) | 0.00740 |
| Resistance to
steroids, % |
|
|
| 0.01456 |
|
Positive (IQR) | 14 (15.55) | 8 (33.33) | 6 (9.09) |
|
|
Negative (IQR) | 76 (84.44) | 16 (66.66) | 60 (90.91) |
|
Microdeletions were determined in 83 (92.22%)
patients (Table III). IKZF1
deletions and ETV6 deletions were more common in patients
with B-ALL compared with patients with T-ALL (40.63% vs. 10.53%,
P=0.0147 and 24.28% vs. 0%, P=0.029). No other significant
differences in the microdeletion profiles between T-ALL and B-ALL
were observed.
| Table III.Microdeletion profiles among included
patients. |
Table III.
Microdeletion profiles among included
patients.
| Microdeletion
locus | All | T-ALL | B-ALL | P-value |
|---|
| IKZF1 | 28 (33.73) | 2 (10.52) | 26 (40.62) | 0.01468 |
|
CDKN2A/B | 51 (61.44) | 13 (68.42) | 38 (59.37) | 0.65777 |
| MIR31 | 18 (23.07) | 3 (18.75) | 15 (24.19) | 0.75102 |
| PAX5 | 16 (20.00) | 2 (11.76) | 14 (22.22) | 0.50036 |
| PAR1 | 19 (26.76) | 3 (23.07) | 16 (27.58) | 1.00000 |
| ETV6 | 17 (24.28) | 0 (0) | 17 (29.82) | 0.02876 |
| BTG1 | 5 (7.14) | 0 (0) | 5 (8.77) | 0.57571 |
| EBF1 | 5 (7.14) | 0 (0) | 5 (8.77) | 0.57571 |
| RB1 | 10 (14.08) | 2 (15.38) | 8 (13.79) | 1.00000 |
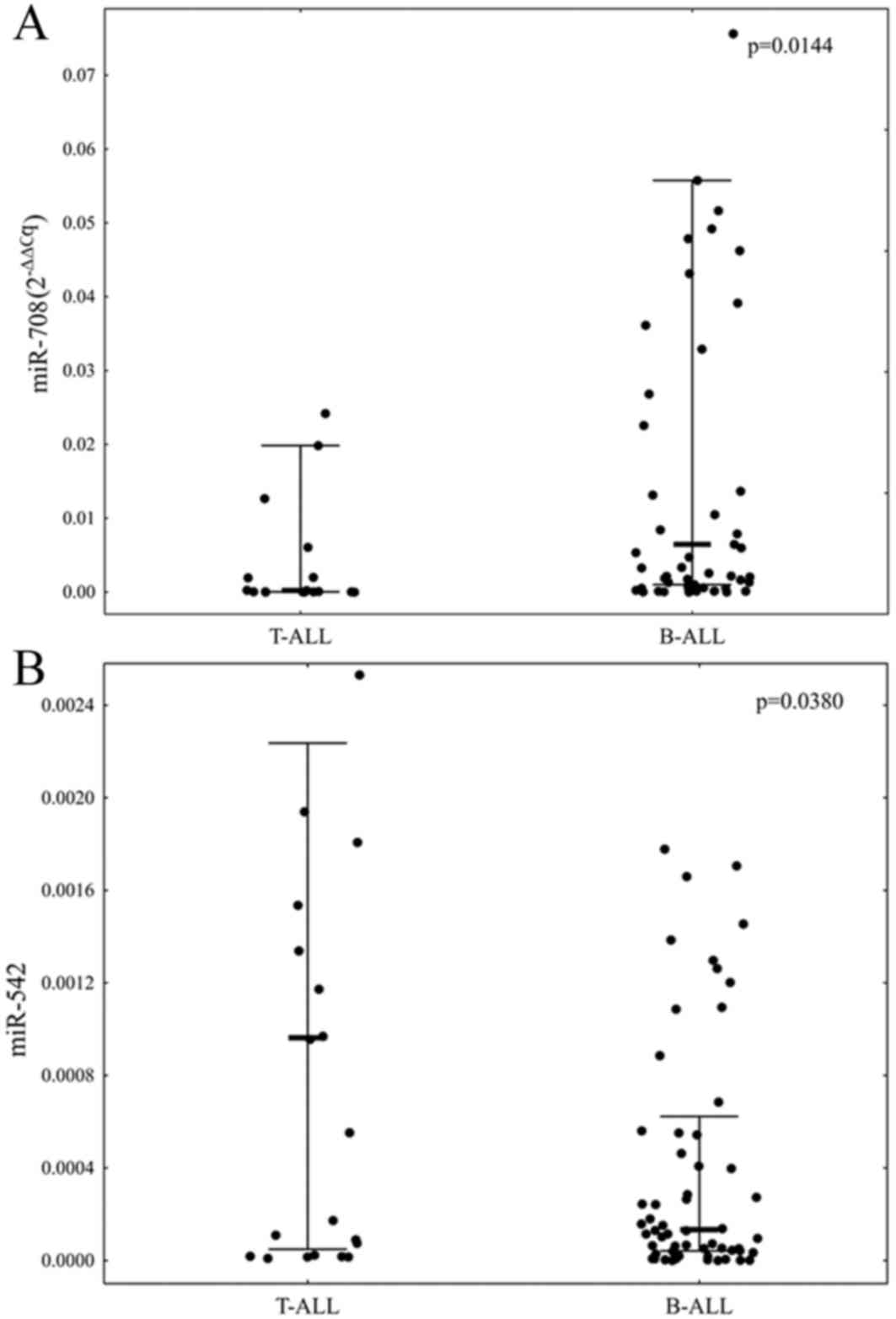
The expression levels of all selected miRNAs were
measurable in 76 (84.44%) of selected samples. A comparison of the
expression levels of the selected miRNAs between T-ALL and B-ALL
patients revealed that patients with B-ALL were characterized by
significantly elevated level of miR-708 and decreased level of
miR-542 (P=0.0144 and P=0.0380, respectively; Fig. 1).
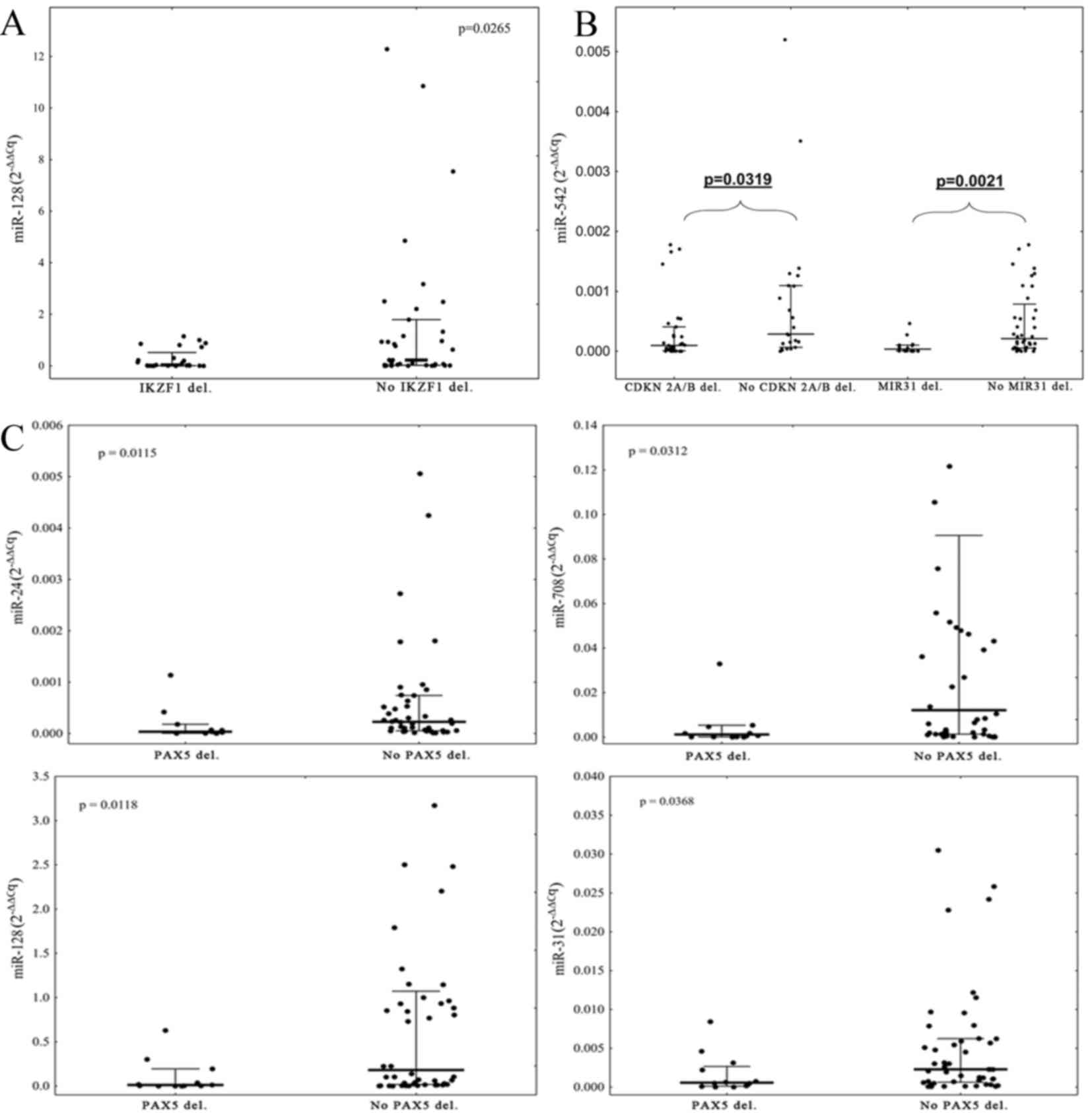
Due to the small number of patients with T-ALL with
microdeletions, the expression levels of miRNAs with the
microdeletion status were compared only within the group of
patients with B-ALL. It was identified that the expression level of
miR-128 was lower in patients with IKZF1 deletions compared
with patients without IKZF1 deletions (P=0.0265, Fig. 2A). Deletions of CDKN2A/B and
MIR31 were associated with low expression of miR-542 in
comparison with patients without deletions (P=0.0319 and P=0.0021,
respectively; Fig. 2B). Patients with
PAX5 deletions exhibited significantly lower expression
levels of miR-31, miR-24, miR-708 and miR-128 in comparison with
patients without deletions of PAX5 (P=0.0368, 0.0115, 0.0312
and 0.0118, respectively; Fig. 2C).
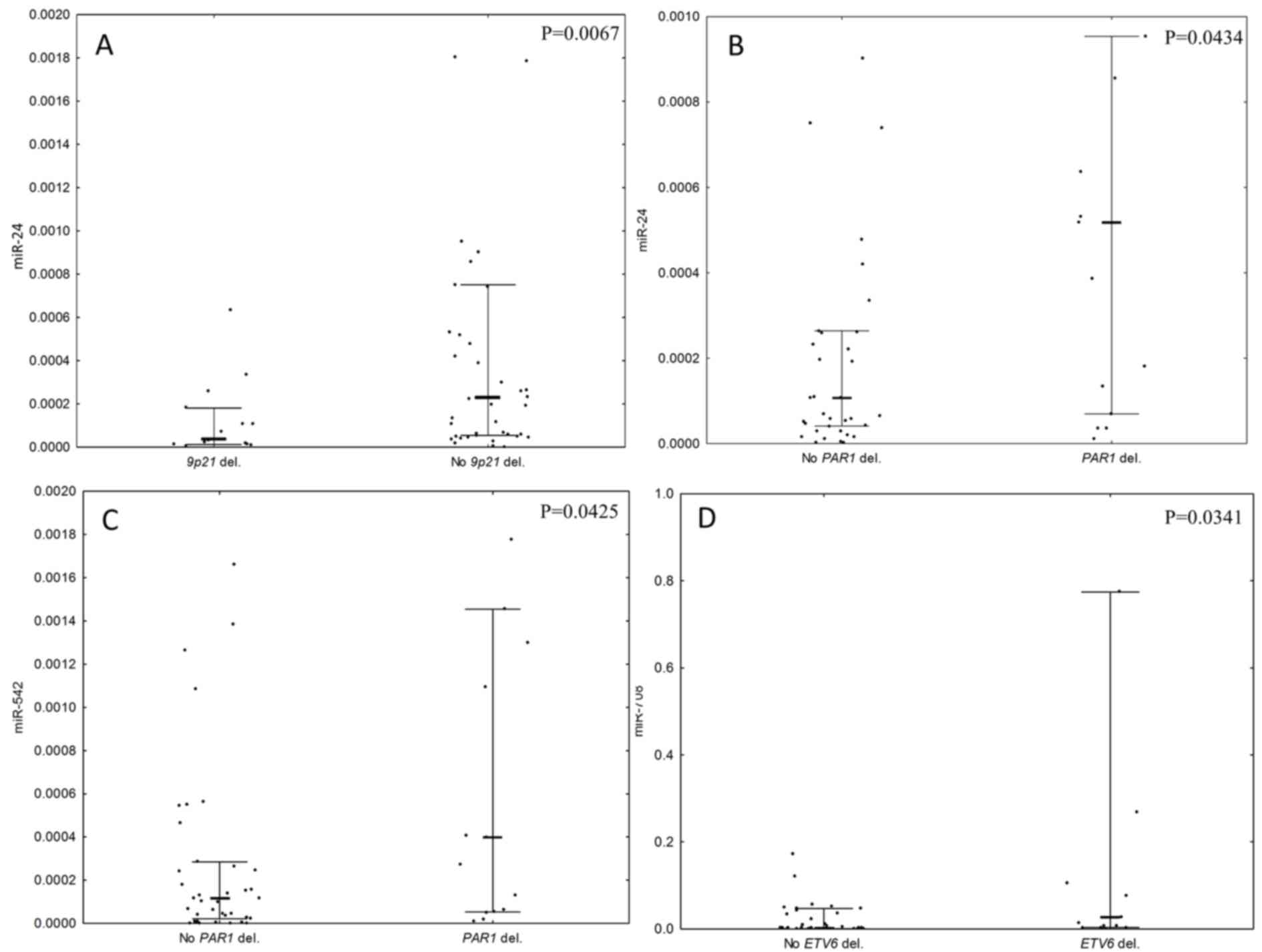
Large deletions in the 9p21 chromosomal region, encompassing
CDKN2A, CDKN2B and MIR31 deletions, were also
associated with low expression of miR-24 (P=0.0067; Fig. 3A). Deletions in the PAR1 region
were associated with high expression levels of miR-24 and miR-542
(P=0.0434 and 0.0425; Fig. 3B and C,
respectively). Deletions of ETV6 were associated with a high
expression of miR-708 (P=0.0341, Fig.
3D). There was no association between the expression levels of
selected miRNAs and deletions of BTG1, EBF1, or
RB1 observed.
Correlation analyses revealed a negative correlation
between age at diagnosis and miR-542 (R2=−0.26,
P=0.043), and between MRD at day 15 and miR-31 (−0.32,
P=0.017) and miR-708 (−0.32, P=0.016) in patients with B-ALL
(Table IV). There were no
statistically significant associations with sex, WBC or resistance
to steroid therapy observed.
| Table IV.Correlation analyses. |
Table IV.
Correlation analyses.
| Variable | miR-31 | miR-24 | miR-708 | miR-542 | miR-128 |
|---|
| Age at
diagnosis | −0.11 | −0.19 | −0.16 |
-0.26a | −0.16 |
| WBC at
diagnosis | −0.08 | 0.09 | −0.05 | −0.03 | 0.09 |
| MRD_15 |
-0.32a | −0.11 |
-0.32a | −0.11 | −0.19 |
| MRD_33 | −0.05 | 0.11 | 0.02 | 0.16 | −0.01 |
Discussion
The data of the present study indicate that selected
miRNAs: miR-128, miR-542, miR-708, miR-24 and miR-31, are
associated with the ALL immunophenotype and microdeletion status of
crucial genes affecting lymphoid development and cell cycle
regulation.
Firstly, it was identified that the expression of
miR-128 was lower in patients with IKZF1 deletions in
comparison with patients without these deletions. IKZF1
encodes Ikaros protein, which has a key role as a transcription
factor in lymphocyte differentiation (22). Ikaros influences more than half of the
known genes associated with hematopoiesis and early B-cell
development (23). IKZF1
deletions are associated with a poor outcome in high-risk groups of
pediatric patients (24,25). In a comparison between normal controls
and patients with AML, an increased expression of miR-128 in
patients with ALL has been suggested (26). Altered miR-128 expression may
discriminate ALL from adult myeloid leukemia and normal bone marrow
samples (27–30). Notably, Mi et al (27) revealed that the differential
expression of miR-128 in acute leukemia is not a consequence of an
altered genomic DNA copy number, and suggested that this miRNA is
also under other types of DNA epigenetic regulation. The
association between the lower expression of miR-128 and steroid
resistance was not evaluated in the present study. In these and
other previous studies, miR-128 expression did not correlate with
other biological and clinical features (28,31). The
role of miR-128 insufficiency in the pathogenesis and response to
treatment in patients with IKZF1-deletion ALL should be
additionally examined with large-scale pediatric patients with
ALL.
Lower miR-128 expression was detected in patients
with deletion of another transcriptional factor gene, PAX5.
Patients with PAX5 deletions exhibited significantly lower
expression levels of miR-31, miR-24 and miR-708 in comparison with
patients without PAX5 deletions.
Compared to patients with T-ALL, a high expression
of miR-708 and a low expression of miR-542 was observed in patients
with B-ALL. miR-708 was overexpressed in patients with ETV6
deletions, and this gene is involved in lymphoid development
(32). These miRNAs were negatively
correlated with MRD status at day 15 of the therapeutic protocol.
Higher MRD was associated with lower miR-708. The investigation of
miR-708 performed by de Oliveira et al (33) revealed lower miR-708 expression in
normal bone marrow samples compared with ALL samples. In their
study, miR-708 expression was not associated with clinical
features, such as white blood cell level, bone marrow blast
infiltration, steroid resistance, or central nervous system or
testis involvement. Han et al (34) revealed a correlation between miR-708
and the response to glucocorticoid therapy, and suggested that low
miR-708 expression was associated with higher risk of
leukemogenesis or relapse. Higher miR-708 expression has been
identified in patients with transcription factor ETV6-acute
myeloid leukemia 1 protein, breakpoint cluster region-abelson
tyrosine-protein kinase 1 (BCR-ABL1), 3-pre-B-cell leukemia
transcription factor 1, hyperdiploid and ‘B-other’ ALL (B-cell ALL
patients without aforementioned defects) compared with
MLL-rearranged B-ALL and T-ALL cases (12,35). In
another study, Li et al (36)
demonstrated that miR-708 was overexpressed in small groups of
Chinese common precursor patients with B-ALL: Expression of miR-708
was higher in the high-risk group compared with the
intermediate-risk group.
In the present study, deletions in the 9p21
chromosomal region (CDKN2A and CDKN2B, MIR31) were
associated with a lower expression of miR-542 in comparison with
patients without deletions. miR-542 regulates the DNA repair/notch
pathway in epithelial prostate cancer, and functions as a tumor
suppressor in neuroblastoma (37,38). This
miRNA may positively regulate tumor protein 53 (p53) activity
(39). The homozygous loss of the
miR-31 gene is described in 9p instability in ALL (40). The 9p instability is detected in 19%
of patients with ALL, and always includes the homozygous loss of
CDKN2A along with loss of CDKN2B, which are
associated with BCR/ABL1 and IKZF1 dysfunction
(40–43). A lower expression of miR-542 is
associated with a deletion of the 9p region. Schotte et al
(12) demonstrated that miR-542 was 1
of 6 significantly upregulated miRNAs in T-ALL compared with normal
thymocytes. Reduced expression of miR-542 and its role in B-ALL
biology requires additional examination, as the current literature
regarding miR-542 expression in ALL and lymphoproliferative
diseases is limited. The present study also identified that
deletions of MIR31were associated with a low miR-24
expression. miR-24 serves a role in the response to DNA damage, and
may enhance apoptosis by targeting B-cell lymphoma 2 (44,45).
miR-24 potentially reduces cellular viability and induces apoptosis
in combination with docetaxel (46).
High expression of miR-24 also correlates with the response to DNA
damage and apoptosis in cells treated with etoposide (47).
The present study observed altered miR-24 expression
in patients with ALL with deletions in PAR1. PAR1 deletions are
frequently observed in patients who are positive for IKZF1
deletion (40). High expressions of
miR-24 and miR-542 in patients who are positive for PAR1 deletion
were detected. The expression level of miR-24 in patients with
acute leukemia is higher compared with the healthy individuals, and
is associated with shorter overall survival, high risk of relapse
and poor survival (48).
Notably, the present study demonstrated that the
lower expression of miR-31 and miR-708 was associated with a high
MRD level and positive PAX5 mutation status. It should be
emphasized that those two investigated miRNAs (miR-31 and miR-708)
negatively influence the NF-κB signaling pathway, which has a role
in B-cell receptor activation (49).
Aberrant B-cell receptor activation leads to a number of lymphoid
malignancies (50). At present,
targeted therapies are focused on the inhibition of BCR signaling
(51,52). miR-708 strongly represses NF-κB
signaling in chronic lymphocytic leukemia (53). miR-31 in adult T-ALL negatively
regulates the NF-κB pathway by targeting NF-κB inducing kinase
(54). Based on this, it is
hypothesized that the loss of activity of these miRNAs may serve a
role in the pathogenesis of ALL, and in treatment failure.
Certain issues must be taken into consideration when
interpreting the present data. One of the most conspicuous points
is the mutation profile of the patients. The mutation rate in the
study population is inconsistent with that of the general
population, which is a consequence of the enrollment criteria.
Patients with IKZF1 deletions were enrolled with priority as
the present study was designed to focus on high-risk patients such
as patients with BCR/ABL1-like ALL. Another important issue
is the underrepresentation of patients with T-ALL, and these
patients were excluded from the majority of additional analyses.
However, differences in the biological features between B-ALL and
T-ALL were demonstrated. Data from another study suggesting that
patients with B-ALL, in comparison to T-ALL, were characterized by
lower white blood cell counts at diagnosis, lower MRD at day 15 of
the therapeutic protocol and greater sensitivity to steroid
treatment were also confirmed by the present study. Finally, it was
not possible to elaborate the activity of all selected miRNAs in
all the investigated samples. Despite this, the expression levels
of at least 4 selected miRNAs were successfully measured in 84
patients (93.33%). To the best of our knowledge, this is the first
study estimating selected miRNA expression, genetic defects and
clinical features in pediatric ALL subtypes. These data indicate
that an aberrant miRNA pattern is associated with ALL types, and
that microdeletions are commonly observed in high-risk patients
with B-ALL.
As demonstrated in the present study, identifying
the miRNA expression status in patients with ALL may be useful for
elucidating the mechanisms of the disease. miRNAs may be used as
molecular markers to assess risk of leukemia relapse, at the time
of diagnosis, and to tailor specific therapies to the individual
patient. Additional studies with larger cohorts are needed to
examine the potential use of these miRNAs (miR-128, miR-542,
miR-708, miR-24 and miR-31) as markers of genetic lesions.
Acknowledgements
The present study was supported by the National
Science Centre, Poland (grant no. 2011/01/B/NZ4/03345). The study
was performed within the project Centre for Innovative Research in
Medical and Natural Sciences realized by the University of Rzeszow,
and co-financed by Regional Operational Programme for the
Podkarpackie Province for the years 2007–2013 (contract no.
UDA-RPPK.01.03.00–18-004/12-00). Ms J. Madzio was supported by The
National Science Centre (grant no. PRELUDIUM 2015/17/N/NZ5/00669).
Dr M. Braun was supported by Polish Ministry of Science and Higher
Education under the Diamond Grant Program (grant no. DI2012017042).
Ms J. Madzio, Dr M. Braun and A.P. were also supported by National
Centre of Research and Development (grant no. LIDER
031/635/l-5/13/NCBR/2014).
References
|
1
|
Ward E, DeSantis C, Robbins A, Kohler B
and Jemal A: Childhood and adolescent cancer statistics, 2014. CA
Cancer J Clin. 64:83–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schwab CJ, Chilton L, Morrison H, Jones L,
Al-Shehhi H, Erhorn A, Russell LJ, Moorman AV and Harrison CJ:
Genes commonly deleted in childhood B-cell precursor acute
lymphoblastic leukemia: Association with cytogenetics and clinical
features. Haematologica. 98:1081–1088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Den Boer ML, van Slegtenhorst M, De
Menezes RX, Cheok MH, Buijs-Gladdines JG, Peters ST, Van Zutven LJ,
Beverloo HB, Van der Spek PJ, Escherich G, et al: A subtype of
childhood acute lymphoblastic leukaemia with poor treatment
outcome: A genome-wide classification study. Lancet Oncol.
10:125–134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mullighan CG: The genomic landscape of
acute lymphoblastic leukemia in children and young adults.
Hematology Am Soc Hematol Educ Program. 2014:174–180.
2014.PubMed/NCBI
|
|
5
|
Figueroa ME, Chen SC, Andersson AK,
Phillips LA, Li Y, Sotzen J, Kundu M, Downing JR, Melnick A and
Mullighan CG: Integrated genetic and epigenetic analysis of
childhood acute lymphoblastic leukemia. J Clin Invest.
123:3099–3111. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yoshida T, Landhuis E, Dose M, Hazan I,
Zhang J, Naito T, Jackson AF, Wu J, Perotti EA, Kaufmann C, et al:
Transcriptional regulation of the Ikzf1 locus. Blood.
122:3149–3159. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mavrakis KJ, Van Der Meulen J, Wolfe AL,
Liu X, Mets E, Taghon T, Khan AA, Setty M, Rondou P, Vandenberghe
P, et al: A cooperative microRNA-tumor suppressor gene network in
acute T-cell lymphoblastic leukemia (T-ALL). Nat Genet. 43:673–678.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Calin GA and Croce CM: Investigation of
microRNA alterations in leukemias and lymphomas. Methods Enzymol.
427:193–213. 2007.PubMed/NCBI
|
|
12
|
Schotte D, De Menezes RX, Akbari Moqadam
F, Khankahdani LM, Lange-Turenhout E, Chen C, Pieters R and Den
Boer ML: MicroRNA characterize genetic diversity and drug
resistance in pediatric acute lymphoblastic leukemia.
Haematologica. 96:703–711. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brennecke J, Stark A, Russell RB and Cohen
SM: Principles of microRNA-target recognition. PLoS Biol.
3:e852005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xie B, Ding Q, Han H and Wu D: miRCancer:
A microRNA-cancer association database constructed by text mining
on literature. Bioinformatics. 29:638–644. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dweep H, Sticht C, Pandey P and Gretz N:
miRWalk-database: Prediction of possible miRNA binding sites by
‘walking’ the genes of three genomes. J Biomed Inform. 44:839–847.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chou CH, Chang NW, Shrestha S, Hsu SD, Lin
YL, Lee WH, Yang CD, Hong HC, Wei TY, Tu SJ, et al: miRTarBase
2016: Updates to the experimentally validated miRNA-target
interactions database. Nucleic Acids Res. 44:D239–D247. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36:D149–D153. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Betel D, Koppal A, Agius P, Sander C and
Leslie C: Comprehensive modeling of microRNA targets predicts
functional non-conserved and non-canonical sites. Genome Biol.
11:R902010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Campbell M, Castillo L, Janic D, Jazbec J,
Kaiserova E, Konja J, Kovas G, Kowalczyk J and Soycan LY: ALL
IC-BFM 2009 - A Randomized Trial of the I-BFM-SG for the Management
of Childhood non-B Acute Lymphoblastic Leukemia. Final Version of
Therapy Protocol from August-14-2009. http://tphd.org.tr/5th_hematoloji_sempozyumu/Lebriz_Yuksel_ALLIC_BFM_2009
|
|
20
|
van Dongen JJ, Lhermitte L, Böttcher S,
Almeida J, van der Velden VH, Flores-Montero J, Rawstron A, Asnafi
V, Lécrevisse Q, Lucio P, et al: EuroFlow antibody panels for
standardized n-dimensional flow cytometric immunophenotyping of
normal, reactive and malignant leukocytes. Leukemia. 26:1908–1975.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-(Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schwickert TA, Tagoh H, Gültekin S, Dakic
A, Axelsson E, Minnich M, Ebert A, Werner B, Roth M, Cimmino L, et
al: Stage-specific control of early B cell development by the
transcription factor Ikaros. Nat Immunol. 15:283–293. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ferreirós-Vidal I, Carroll T, Taylor B,
Terry A, Liang Z, Bruno L, Dharmalingam G, Khadayate S, Cobb BS,
Smale ST, et al: Genome-wide identification of Ikaros targets
elucidates its contribution to mouse B-cell lineage specification
and pre-B-cell differentiation. Blood. 121:1769–1782. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mullighan CG, Su X, Zhang J, Radtke I,
Phillips LA, Miller CB, Ma J, Liu W, Cheng C, Schulman BA, et al:
Deletion of IKZF1 and prognosis in acute lymphoblastic leukemia. N
Engl J Med. 360:470–480. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dörge P, Meissner B, Zimmermann M, Möricke
A, Schrauder A, Bouquin JP, Schewe D, Harbott J, Teigler-Schlegel
A, Ratei R, et al: IKZF1 deletion is an independent predictor of
outcome in pediatric acute lymphoblastic leukemia treated according
to the ALL-BFM 2000 protocol. Haematologica. 98:428–432. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu YD, Wang L, Sun C, Fan L, Zhu DX, Fang
C, Wang YH, Zou ZJ, Zhang SJ, Li JY and Xu W: Distinctive microRNA
signature is associated with the diagnosis and prognosis of acute
leukemia. Med Oncol. 29:2323–2331. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mi S, Lu J, Sun M, Li Z, Zhang H, Neilly
MB, Wang Y, Qian Z, Jin J, Zhang Y, et al: MicroRNA expression
signatures accurately discriminate acute lymphoblastic leukemia
from acute myeloid leukemia. Proc Natl Acad Sci USA.
104:19971–19976. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
de Oliveira JC, Scrideli CA, Brassesco MS,
Morales AG, Pezuk JA, Queiroz Rde P, Yunes JA, Brandalise SR and
Tone LG: Differential miRNA expression in childhood acute
lymphoblastic leukemia and association with clinical and biological
features. Leuk Res. 36:293–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang H, Luo XQ, Zhang P, Huang LB, Zheng
YS, Wu J, Zhou H, Qu LH, Xu L and Chen YQ: MicroRNA patterns
associated with clinical prognostic parameters and CNS relapse
prediction in pediatric acute leukemia. PLoS One. 4:e78262009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Li Z, He C, Wang D, Yuan X, Chen J
and Jin J: MicroRNAs expression signatures are associated with
lineage and survival in acute leukemias. Blood Cells Mol Dis.
44:191–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang LC, Swat W, Fujiwara Y, Davidson L,
Visvader J, Kuo F, Alt FW, Gilliland DG, Golub TR and Orkin SH: The
TEL/ETV6 gene is required specifically for hematopoiesis in the
bone marrow. Genes Dev. 12:2392–2402. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
de Oliveira JC, Scrideli CA, Brassesco MS,
Yunes JA, Brandalise SR and Tone LG: MiR-708-5p is differentially
expressed in childhood acute lymphoblastic leukemia but not
strongly associated to clinical features. Pediatr Blood Cancer.
62:177–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han BW, Feng DD, Li ZG, Luo XQ, Zhang H,
Li XJ, Zhang XJ, Zheng LL, Zeng CW, Lin KY, et al: A set of miRNAs
that involve in the pathways of drug resistance and leukemic
stem-cell differentiation is associated with the risk of relapse
and glucocorticoid response in childhood ALL. Hum Mol Genet.
20:4903–4915. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schotte D, Chau JC, Sylvester G, Liu G,
Chen C, van der Velden VH, Broekhuis MJ, Peters TC, Pieters R and
den Boer ML: Identification of new microRNA genes and aberrant
microRNA profiles in childhood acute lymphoblastic leukemia.
Leukemia. 23:313–322. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li X, Li D, Zhuang Y, Shi Q, Wei W and Ju
X: Overexpression of miR-708 and its targets in the childhood
common precursor B-cell ALL. Pediatr Blood Cancer. 60:2060–2067.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rane JK, Ylipää A, Adamson R, Mann VM,
Simms MS, Collins AT, Visakorpi T, Nykter M and Maitland NJ:
Construction of therapeutically relevant human prostate epithelial
fate map by utilising miRNA and mRNA microarray expression data. Br
J Cancer. 113:611–615. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bray I, Tivnan A, Bryan K, Foley NH,
Watters KM, Tracey L, Davidoff AM and Stallings RL: MicroRNA-542-5p
as a novel tumor suppressor in neuroblastoma. Cancer Lett.
303:56–64. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Y, Huang JW, Castella M, Huntsman DG
and Taniguchi T: p53 is positively regulated by miR-542-3p. Cancer
Res. 74:3218–3227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Usvasalo A, Ninomiya S, Räty R, Hollmén J,
Saarinen-Pihkala UM, Elonen E and Knuutila S: Focal 9p instability
in hematologic neoplasias revealed by comparative genomic
hybridization and single-nucleotide polymorphism microarray
analyses. Genes. Chromosomes Cancer. 49:309–318. 2010.
|
|
41
|
Sherborne AL, Hosking FJ, Prasad RB, Kumar
R, Koehler R, Vijayakrishnan J, Papaemmanuil E, Bartram CR,
Stanulla M, Schrappe M, et al: Variation in CDKN2A at 9p21.3
influences childhood acute lymphoblastic leukemia risk. Nat Genet.
42:492–494. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
42
|
Olsson L, Albitar F, Castor A, Behrendtz
M, Biloglav A, Paulsson K and Johansson B: Cooperative genetic
changes in pediatric B-cell precursor acute lymphoblastic leukemia
with deletions or mutations of IKZF1. Genes Chromosomes Cancer.
54:315–325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Braun M, Pastorczak A, Fendler W, Madzio
J, Tomasik B, Taha J, Bielska M, Sedek L, Szczepanski T, Matysiak
M, et al: Biallelic loss of CDKN2A is associated with poor response
to treatment in pediatric acute lymphoblastic leukemia. Leuk
Lymphoma. 18:1162–1171. 2016.
|
|
44
|
Lal A, Pan Y, Navarro F, Dykxhoorn DM,
Moreau L, Meire E, Bentwich Z, Lieberman J and Chowdhury D:
miR-24-mediated downregulation of H2AX suppresses DNA repair in
terminally differentiated blood cells. Nat Struct Mol Biol.
16:492–498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Srivastava N, Manvati S, Srivastava A, Pal
R, Kalaiarasan P, Chattopadhyay S, Gochhait S, Dua R and Bamezai
RN: miR-24-2 controls H2AFX expression regardless of gene copy
number alteration and induces apoptosis by targeting antiapoptotic
gene BCL-2: A potential for therapeutic intervention. Breast Cancer
Res. 13:R392011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Manvati S, Mangalhara KC, Kalaiarasan P,
Srivastava N and Bamezai RN: miR-24-2 regulates genes in survival
pathway and demonstrates potential in reducing cellular viability
in combination with docetaxel. Gene. 567:217–224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Brunner S, Herndler-Brandstetter D, Arnold
CR, Wiegers GJ, Villunger A, Hackl M, Grillari J, Moreno-Villanueva
M, Bürkle A and Grubeck-Loebenstein B: Upregulation of miR-24 is
associated with a decreased DNA damage response upon etoposide
treatment in highly differentiated CD8(+) T cells sensitizing them
to apoptotic cell death. Aging Cell. 11:579–587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Organista-Nava J, Gómez-Gómez Y,
Illades-Aguiar B, Del Carmen Alarcón-Romero L, Saavedra-Herrera MV,
Rivera-Ramírez AB, Garzón-Barrientos VH and Leyva-Vázquez MA: High
miR-24 expression is associated with risk of relapse and poor
survival in acute leukemia. Oncol Rep. 33:1639–1649.
2015.PubMed/NCBI
|
|
49
|
Talab F, Thompson V, Allen JC, Lin K and
Slupsky JR: Characterisation of B cell receptor-induced NF-κB
activation in chronic lymphocytic leukaemia cells. Blood.
116:37652015.
|
|
50
|
Niemann CU and Wiestner A: B-cell receptor
signaling as a driver of lymphoma development and evolution. Semin
Cancer Biol. 23:410–421. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lannutti BJ, Meadows SA, Herman SE,
Kashishian A, Steiner B, Johnson AJ, Byrd JC, Tyner JW, Loriaux MM,
Deininger M, et al: CAL-101, a p110delta selective
phosphatidylinositol-3-kinase inhibitor for the treatment of B-cell
malignancies, inhibits PI3K signaling and cellular viability.
Blood. 117:591–594. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Byrd JC, Furman RR, Coutre SE, Flinn IW,
Burger JA, Blum KA, Grant B, Sharman JP, Coleman M, Wierda WG, et
al: Targeting BTK with ibrutinib in relapsed chronic lymphocytic
leukemia. N Engl J Med. 369:32–42. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Baer C, Oakes CC, Ruppert AS, Claus R,
Kim-Wanner SZ, Mertens D, Zenz T, Stilgenbauer S, Byrd JC and Plass
C: Epigenetic silencing of miR-708 enhances NF-κB signaling in
chronic lymphocytic leukemia. Int J Cancer. 137:1352–1361. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yamagishi M, Nakano K, Miyake A, Yamochi
T, Kagami Y, Tsutsumi A, Matsuda Y, Sato-Otsubo A, Muto S,
Utsunomiya A, et al: Polycomb-mediated loss of miR-31 activates
NIK-dependent NF-κB pathway in adult T cell leukemia and other
cancers. Cancer Cell. 21:121–135. 2012. View Article : Google Scholar : PubMed/NCBI
|