Introduction
Serine/arginine splicing factor 1 (SRSF1, also known
as SF2/ASF) is a member of the SR protein family of splicing
regulators. SRSF1 regulates alternative splicing through
recognizing and binding the exonic splicing enhancers (ESEs)
(1). In addition, SRSF1 is involved
in the regulation of mRNA stability (2), mRNA export (3), nonsense-mediated mRNA decay (4), translation (5–7) and
microRNA processing (8). It has been
demonstrated that SRSF1 is a well-characterized oncogenic factor
that promotes tumorigenesis by controlling a number of alternative
splicing (AS) events (9). A previous
study has identified that SRSF1 promotes the skipping of RON
proto-oncogene (RON) exon 11 to generate RONΔ11, which stimulates
cell migration and invasion (10).
The expression level of SRSF1 is upregulated in human breast
cancer, and SRSF1 overexpression promotes the transformation of
mammary cells by regulating a number of AS events (11). Additionally, the overexpression of
SRSF1 has been identified in colon, thyroid, small intestine,
kidney, lung, liver and pancreas tumors (12).
Colorectal cancer, as the third most common type of
cancer and the fourth leading cause of cancer-associated mortality,
affects >1,000,000 people and causes >500,000 mortalities
every year worldwide (13).
Histological types of colon cancer include adenocarcinoma,
scirrhous tumors and neuroendocrine (14).
Treatments for patients with colon cancer include
combinations of surgery, radiation therapy, chemotherapy and
targeted therapy (13). Cancer of the
colon may be cured with surgery when localized to the bowel;
however, cancer that has spread or cancer recurrence following
surgery are typically incurable (13). Thus, the development of effective
biomarkers and specific therapeutic targets is required.
SRSF1 serves a key function in the tumorigenesis of
lung and breast cancer (9,11,15,16), but
there is limited information demonstrating the association between
SRSF1 and colorectal cancer. SRSF1 may regulate alternative
splicing of tumor-related Ras-related C3 botulinum toxin substrate
1b (Rac1b) by promoting the inclusion of alternative exon 3b in
colorectal cells (17). Furthermore,
alternative splicing of solute carrier family member 39 member 14
in colorectal cancer was identified to be regulated in the Wnt
signaling pathway, and this was hypothesized to occur via the
regulation of SRSF protein kinase 1 and SRSF1 (18). However, there is a limited amount of
network analysis from a global aspect to study the effect of SRSF1
on colorectal cancer.
In the present study, the enriched functions and
signaling pathways of SRSF1 were investigated using Gene Ontology
(GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis of
available gene regulation data. Additionally, Oncomine analysis was
performed, which revealed that SRSF1 was upregulated in a number of
types of colon cancer. Furthermore, the Search Tool for the
Retrieval of Interacting Genes/Proteins (STRING) database and
Cytoscape analysis demonstrated that SRSF1-related colorectal
cancer genes have potential interaction networks in which these
genes were enriched in DNA metabolic processes, cell cycle
regulation and regulation of apoptosis.
Materials and methods
The expression levels of SRSF1 in a number of types
of colon cancer were obtained from the Oncomine dataset (https://www.oncomine.org/resource/login.html). The
immunohistochemical staining results of SRSF1 in colorectal cancer
and normal tissues were selected from the Human Protein Atlas
(http://www.proteinatlas.org). cBioPortal
(http://www.cbioportal.org/) was applied
to identify the SRSF1 co-expressed genes. Briefly, the threshold
selected was the absolute value of Pearson correlation coefficient
>0.4. Furthermore, Venny 2.1 (BioinfoGP, CNB-CSIC, http://bioinfogp.cnb.csic.es/tools/venny/index.html)
was utilized to overlap the identified gene set with a colon
cancer-related gene list, screened using GeneCards. The resulting
overlapped genes were defined as SRSF1-related genes in colon
cancer. The GO and KEGG analysis were conducted with all genes on
Database for Annotation, Visualization and Integrated Discovery
platform (http://david.abcc.ncifcrf.gov/summary.jsp). Fisher
exact P-values were calculated using SPSS 19.0 (IBM Corp., Armonk,
NY, USA) for each enriched functional category and pathway, and
enriched functional categories and pathways with P<0.05 were
presented. The online tool STRING (http://string-db.org) was used to identify the
potential interaction networks of protein products of these genes.
The Cytoscape 2.8.3 software (Institute of Systems Biology,
Seattle, WA, USA) was further utilized to construct interaction
networks and sub-networks among these SRSF1-associated gene
products in colon cancer. The gene regulation networks and protein
interactions were visualized by using Cytoscape 2.8.3 software.
Results
Network of SRSF1-related genes
The co-expression network of SRSF1-regulated genes,
from the colorectal cancer gene expression profiling database, was
established using cBioPortal by setting the Pearson correlation
efficient at >0.4. A total of 2,678 genes were identified to be
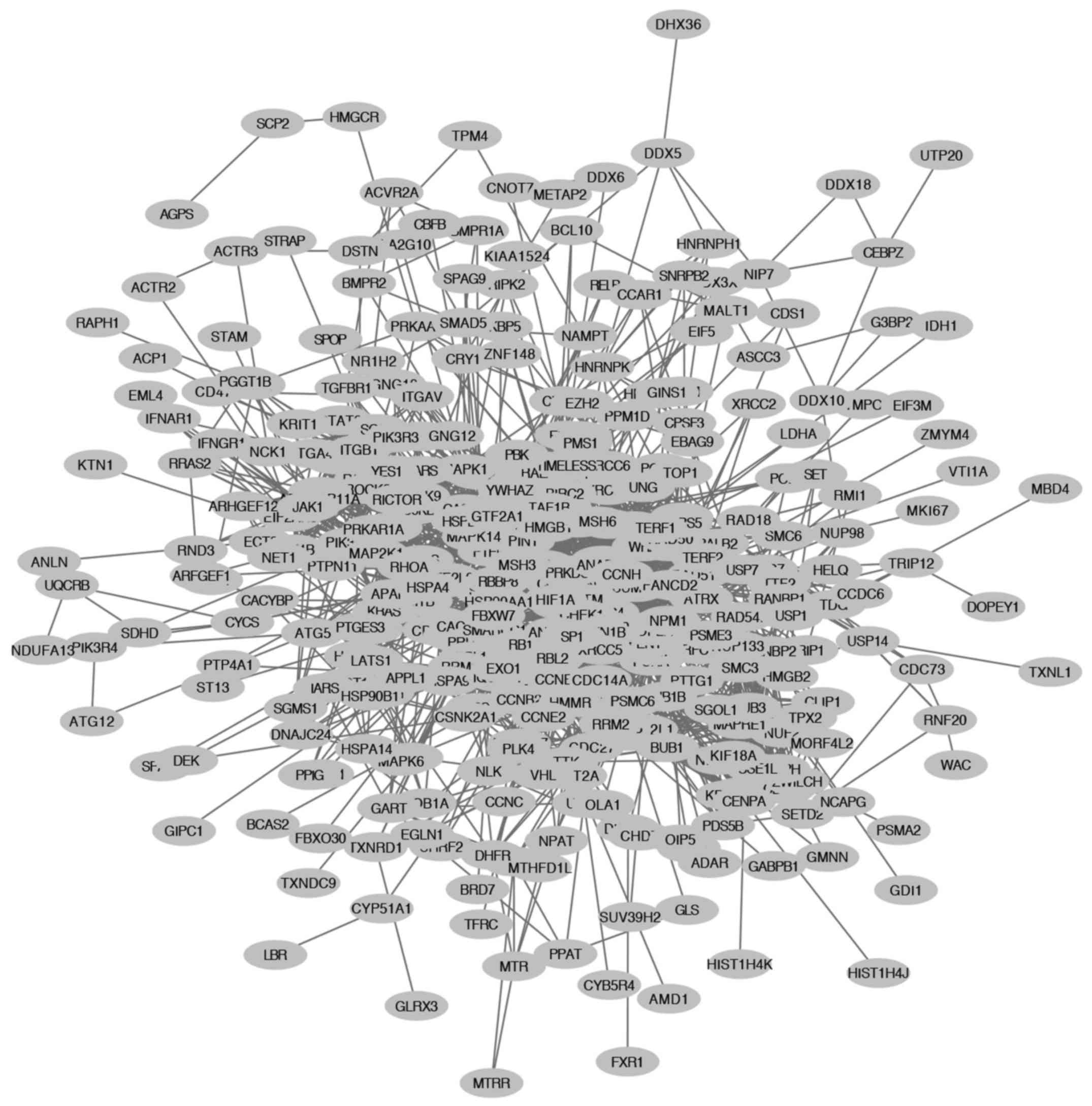
SRSF1-related. Protein-protein interaction networks of 913/2,678
genes were constructed using the STRING database analysis (Fig. 1).
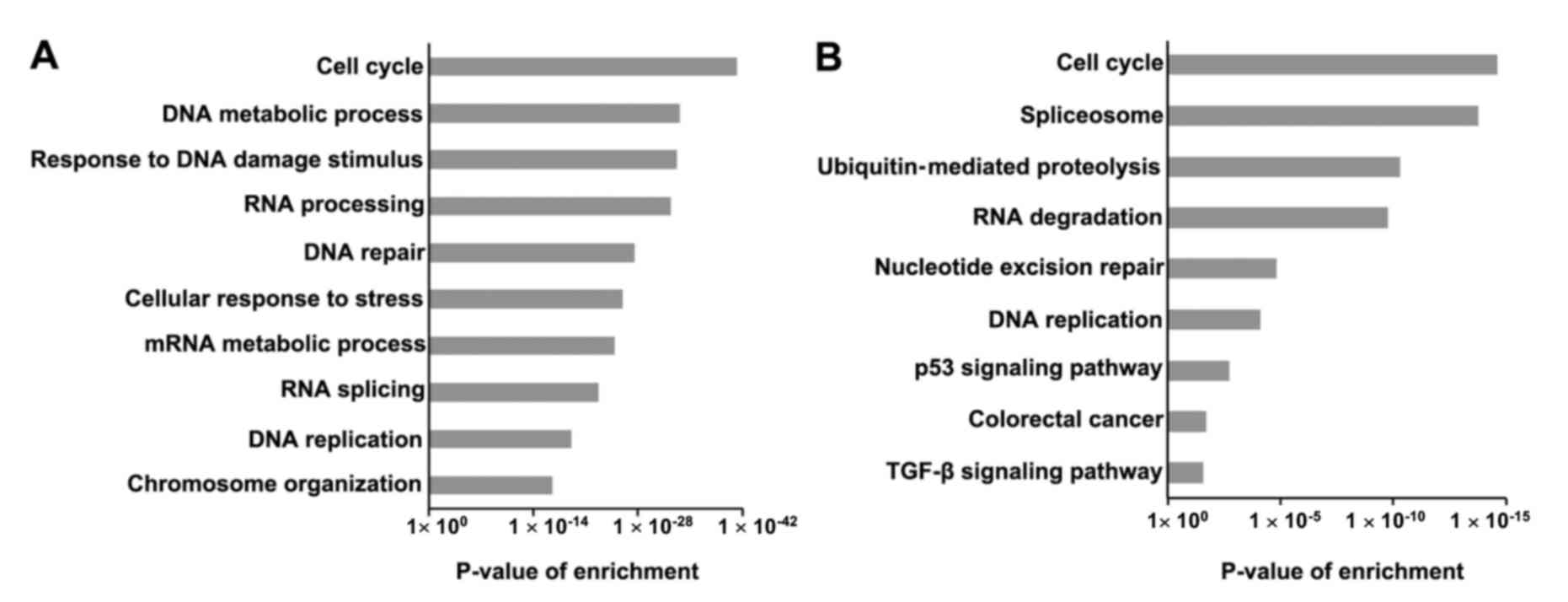
GO analysis for the SRSF1-related proteins revealed
that SRSF1-regulated cancers genes were markedly enriched in a
number of functions, including regulation of the cell cycle, DNA
metabolic processes, RNA processing and DNA replication (Fig. 2A). The results indicated that SRSF1 is
involved in cancer cell proliferation. In addition, KEGG pathway
analysis revealed that the SRSF1-related genes were markedly
enriched in cell cycle, spliceosome, ubiquitin-mediated
proteolysis, RNA degradation, nucleotide excision repair, DNA
replication and p53 signaling pathway (Fig. 2B). Notably, SRSF1-regulated genes
participated in the progression of colorectal cancer, suggesting
that SRSF1 may affect colorectal cancer development.
SRSF1 serves a key function in
colorectal cancer progression
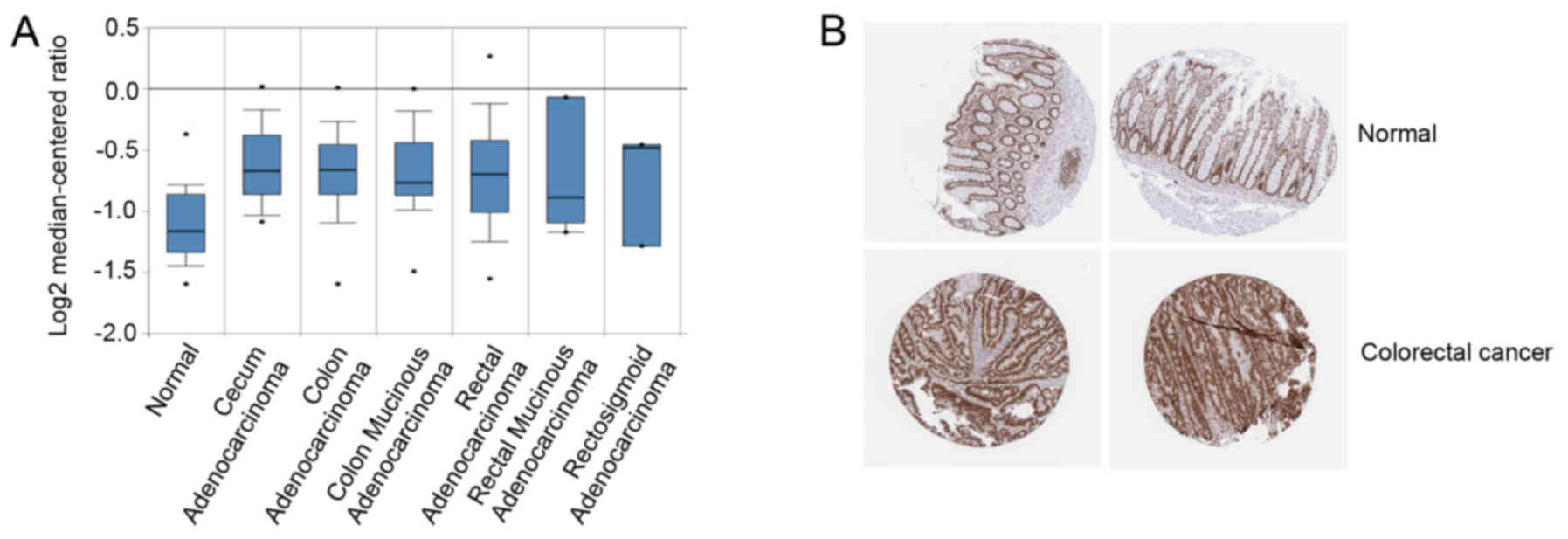
The association between SRSF1 and colorectal cancer
was further analyzed. Using Oncomine analysis, the expression level
of SRSF1 was determined in colon cancer. It was identified that
SRSF1 was upregulated in a number of types of colon cancer,
including cecum adenocarcinoma, colon adenocarcinoma and colon
mucinous adenocarcinoma (Fig. 3A). In
addition, the immunohistochemistry staining results of SRSF1,
obtained from the Human Protein Atlas database (http://www.proteinatlas.org) (19), were analyzed, which revealed that
SRSF1 was upregulated in colorectal cancer, compared with normal
tissues (Fig. 3B) (http://www.proteinatlas.org/ENSG00000136450-SRSF1/cancer/tissue/colorectal+cancer).
Taken together, SRSF1 may be involved in colorectal cancer
progression.
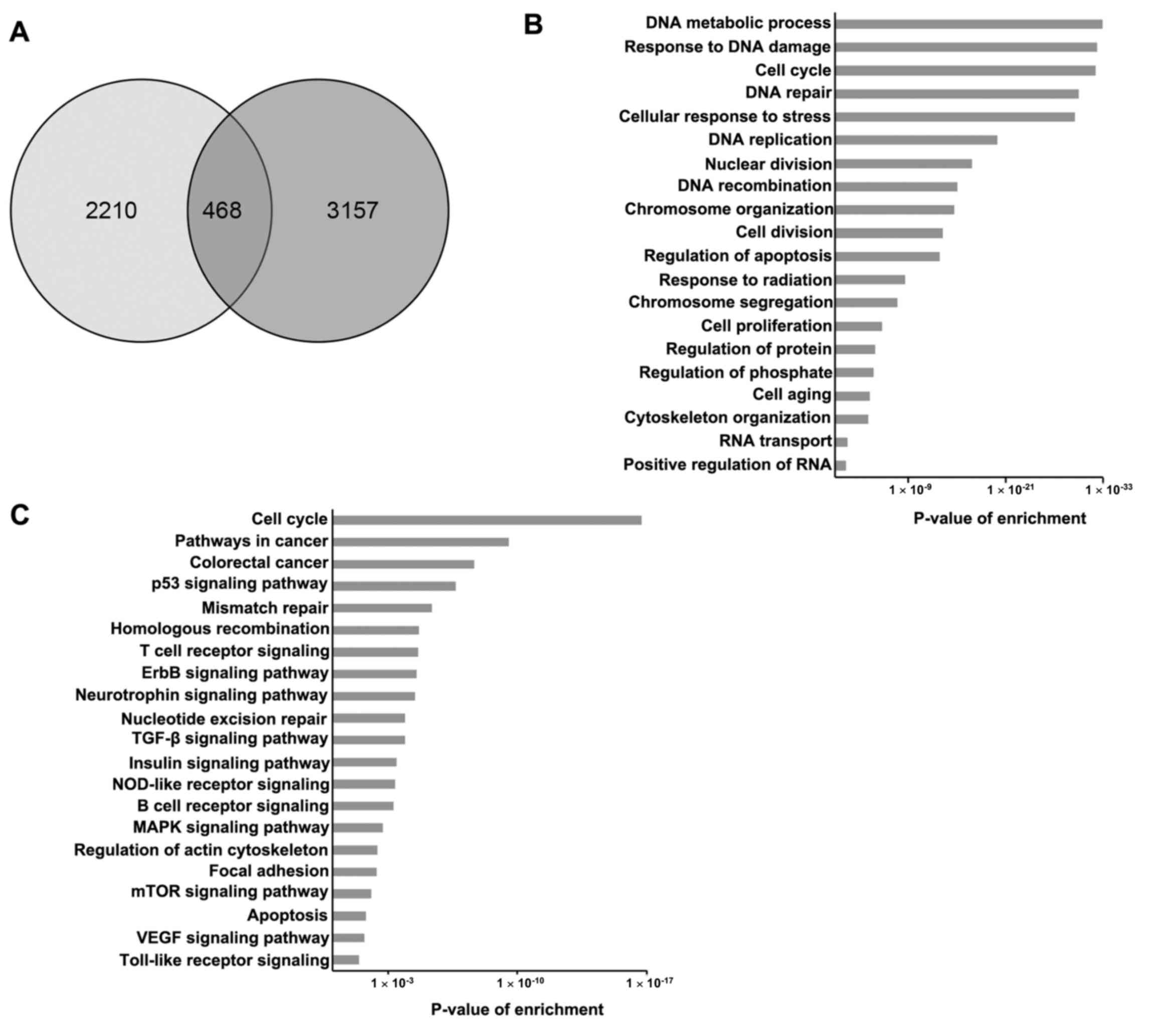
Overlapping analysis of 2,678 SRSF1-related genes
and 3,625 genes defined as colorectal cancer genes in GeneCards
identified 468 genes as SRSF1-related colorectal cancer genes
(Fig. 4A). Subsequently, GO and KEGG
pathway analysis was performed to classify the 468 overlapped
genes. The GO results demonstrated that the overlapped genes were
enriched in a number of biological processes, including metabolic
processes, response to DNA damage and cell cycle regulation
(Fig. 4B). Furthermore, KEGG pathway
analysis revealed that SRSF1-related colorectal cancer genes were
involved in the regulation of the cell cycle, deregulated signaling
pathways associated with cancer progression and the colorectal
cancer signaling pathway (Fig.
4C).
Networks of SRSF1-related colorectal
cancer genes
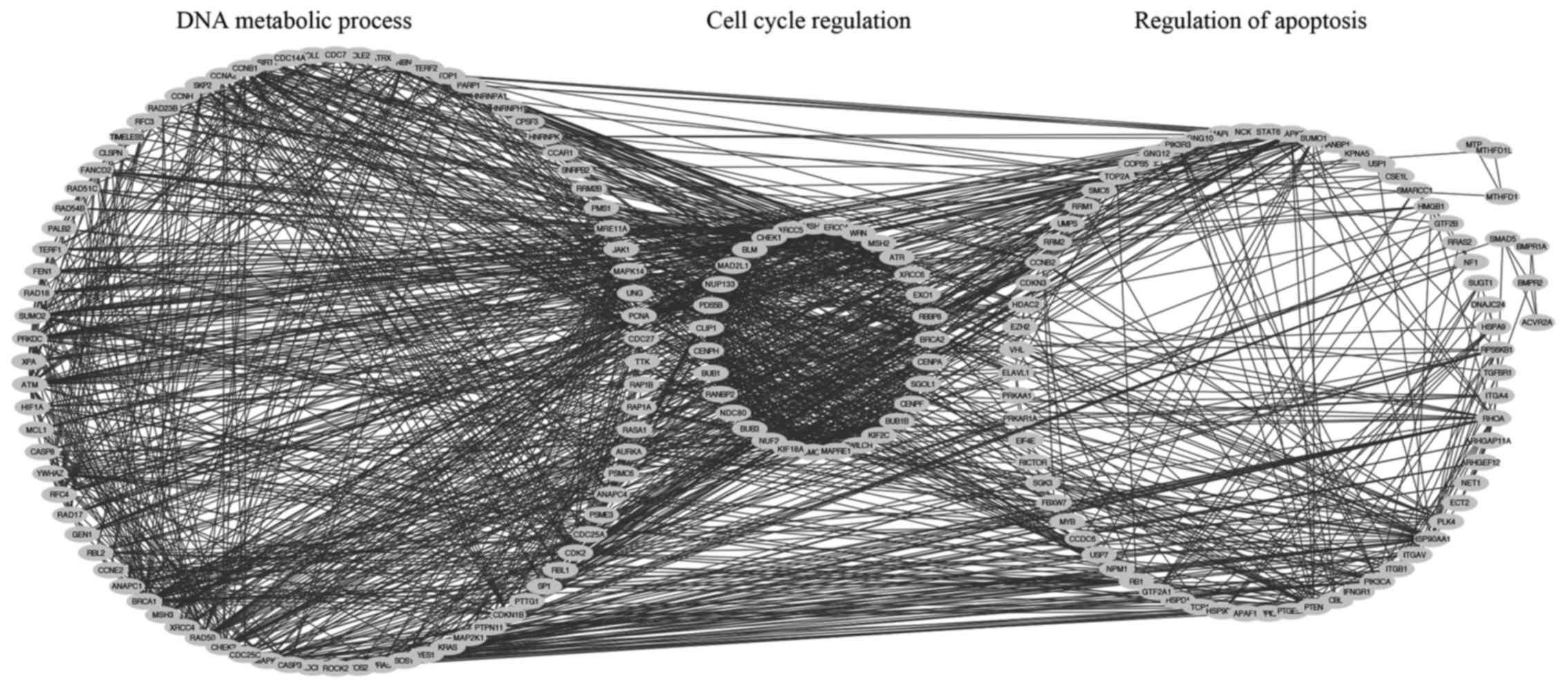
The STRING database and Cytoscape analysis was
conducted on SRSF1-related colorectal cancer genes to reveal
potential interaction networks. GO demonstrated that
SRSF1-associated colorectal cancer genes were enriched in the
functions of DNA metabolic processes, cell cycle regulation and
regulation of apoptosis (Fig. 5).
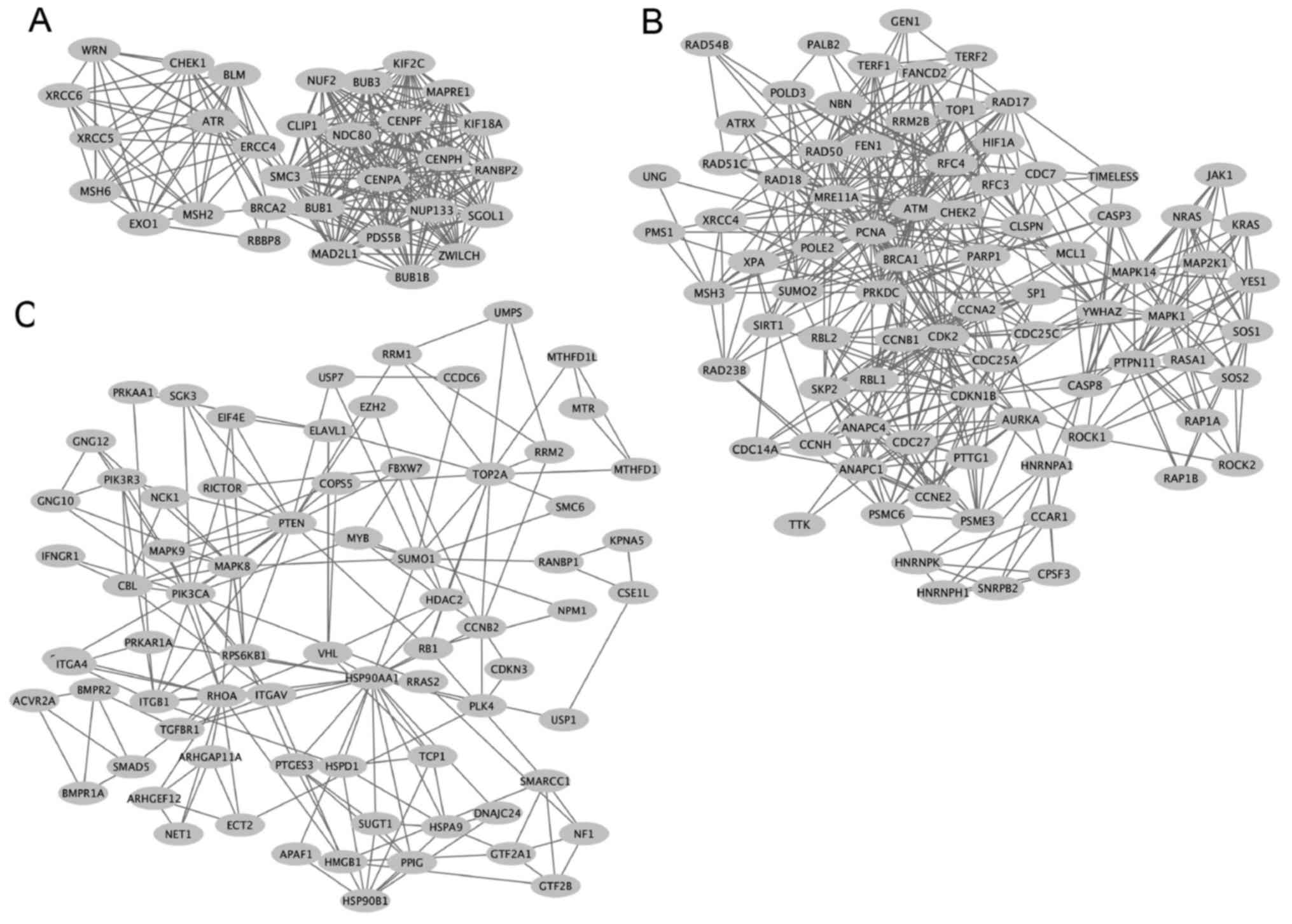
A sub-network of SRSF1-related colorectal cancer
genes was identified to be enriched in cell cycle regulation. NUF2
NDC80 kinetochore complex component (NUF2), kinesin family member
2c (KIF2C), structural maintenance of chromosome 3 (SMC3), MAD2
mitotic arrest deficient-like 1 (MAD2L1) and ZWILCH kinetochore
protein (ZWILCH) genes were identified to exhibit a high degree of
interaction in colorectal cancer (Fig.
6A). In the sub-network of SRSF1-related colorectal cancer
genes that were involved in DNA metabolic processes, ATM
serine/threonine kinase (ATM), BRCA1 DNA repair associated (BRCA1),
protein kinase DNA activated catalytic polypeptide (PRKDC),
proliferating cancer nuclear antigen (PCNA), topoisomerase (DNA) 1
(TOP1) and MRE11 homolog double strain break repair nuclease
(MRE11A) genes revealed a high connectivity value (Fig. 6B). Furthermore, heat shock protein 90
alpha family class A member 1 (HSP90AA1), Ras homolog family member
A (RHOA), phosphatase and tensin homolog (PTEN) and ribosomal
protein S6 kinase B1 (RPS6KB1) exhibited a high degree of
interaction in another sub-network that regulated apoptosis
(Fig. 6C).
Discussion
Serine/Arginine-rich (SR) proteins are a family of
RNA-binding proteins with a serine/arginine rich domain. SR
proteins regulate alternative splicing of a number of genes that
are involved in cell cycle regulation, cell proliferation,
apoptosis, epithelial-mesenchymal transition and drug resistance
(10,12,20–22). As a
member of SR proteins, SRSF1 (SF2/ASF) functions as a
proto-oncogene that is generally upregulated in various types of
cancer (12,23). In addition, SRSF1 may promote
tumorigenesis by controlling the alternative splicing of a number
of cancer-related genes, including RPS6KB1 the tumor suppressor
bridging integrator 1, the tyrosine kinase receptor for
macrophage-stimulating protein, cyclin D1, and vascular endothelial
growth factor (24,25).
In the present study, bioinformatics analysis was
performed systematically to determine the effect of SRSF1 on
colorectal cancer. Gene expression data were obtained from The
Cancer Genome Atlas database. Subsequently, potential
SRSF1-regulated genes in colorectal cancer were obtained using
cBioPortal online tools, followed by GO, KEGG and protein
interactome analyses. Oncomine analysis was performed which
revealed that SRSF1 is upregulated in a number of types of colon
cancer. A total of 468 genes were identified as SRSF1-related
colorectal cancer genes by the overlapping analysis of 2,678
SRSF1-related genes and 3,625 colorectal cancer genes (identified
using GeneCards). The GO results suggested that these
SRSF1-associated colorectal cancer genes were primarily enriched in
metabolic processes, response to DNA damage and regulation of the
cell cycle. In addition, KEGG pathway analysis revealed that
SRSF1-related colorectal cancer genes were closely associated with
cell cycle, deregulated signaling pathways associated with cancer
progression and colorectal cancer signaling pathways. STRING
database and Cytoscape analysis demonstrated that 468 SRSF1-related
colorectal cancer genes exhibited potential interaction networks
with functions of DNA metabolic process, cell cycle regulation and
regulation of apoptosis. Notably, a panel of genes (NUF2, KIF2C,
SMC3, MAD2L1 and ZWILCH) responsible for cell cycle regulation were
identified to be highly associated with the expression level of
SRSF1. NUF2 is a component of NDC80 kinetochore complex and is
known to be critical for stable spindle microtubule-kinetochore
attachment (26). The expression
level of NUF2 has been identified to be upregulated and associated
with poor prognosis in patients with colorectal cancer (27). Previous studies have revealed that
NUF2 serves a function in a number of types of human cancer
including lung, colorectal, gastric, prostate, urinary bladder,
renal carcinoma and ovarian cancer (28–33). KIF2C
(Kinesin family member 2C, also known as the mitotic
centromere-associated kinesin, MACK) is the best characterized
member of the kinesin-13 family, which is critical in the
regulation of microtubule dynamics. KIF2C has been identified to be
aberrantly regulated in cancer cells, which is associated with
increased malignance, invasiveness, metastasis, drug resistance,
chromosomal instability and remodeling of the microtubule
cytoskeleton (34,35).
ATM, BRCA1, PRKDC, PCNA, TOP1 and MRE11A genes,
involved in DNA metabolic processes, exhibit a high degree of
interaction with SRSF1. ATM is an important cell cycle checkpoint
kinase and a key regulator of the DNA double-strand-break response
(36). ATM may regulate a number of
downstream proteins, including p53, BRCA1, checkpoint kinase 2,
checkpoint clamp loader component RAD17, cell cycle checkpoint
control protein RAD9 and Nibrin. ATM and serine/threonine-protein
kinase ATR, which are hypothesized to be regulators of cell cycle
checkpoint signaling pathways (36).
Genes that regulate apoptosis, including HSP90AA1,
RHOA, PTEN and RPS6KB1, were highly associated with the expression
level of SRSF1. RHOA is a member of Rho GTPases, which are involved
in major aspects of cancer development, including cell
proliferation, apoptosis, cell polarity, adhesion, migration and
invasion (37). Overexpression of
RHOA is associated with invasion and poor prognosis in colorectal
cancer (38).
The results of the present study, which was
conducted using bioinformatics analysis, indicated that SRSF1 may
have key functions in the progression and development of colorectal
cancer by interaction networks. Additional studies may verify this
potential regulatory network by performing molecular biology
experiments.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 31400726) and the
Natural Science Foundation of Liaoning Province of China (grant no.
2015020301).
References
|
1
|
Sun Q, Mayeda A, Hampson RK, Krainer AR
and Rottman FM: General splicing factor SF2/ASF promotes
alternative splicing by binding to an exonic splicing enhancer.
Genes Dev. 7:2598–2608. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lemaire R, Prasad J, Kashima T, Gustafson
J, Manley JL and Lafyatis R: Stability of a PKCI-1-related mRNA is
controlled by the splicing factor ASF/SF2: A novel function for SR
proteins. Genes Dev. 16:594–607. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang Y, Gattoni R, Stevenin J and Steitz
JA: SR splicing factors serve as adapter proteins for TAP-dependent
mRNA export. Mol Cell. 11:837–843. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang Z and Krainer AR: Involvement of SR
proteins in mRNA surveillance. Mol Cell. 16:597–607. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Michlewski G, Sanford JR and Cáceres JF:
The splicing factor SF2/ASF regulates translation initiation by
enhancing phosphorylation of 4E-BP1. Mol Cell. 30:179–189. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun S, Zhang Z, Sinha R, Karni R and
Krainer AR: SF2/ASF autoregulation involves multiple layers of
post-transcriptional and translational control. Nat Struct Mol
Biol. 17:306–312. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sanford JR, Gray NK, Beckmann K and
Cáceres JF: A novel role for shuttling SR proteins in mRNA
translation. Genes Dev. 18:755–768. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu H, Sun S, Tu K, Gao Y, Xie B, Krainer
AR and Zhu J: A splicing-independent function of SF2/ASF in
microRNA processing. Mol Cell. 38:67–77. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Anczukow O, Rosenberg AZ, Akerman M, Das
S, Zhan L, Karni R, Muthuswamy SK and Krainer AR: The splicing
factor SRSF1 regulates apoptosis and proliferation to promote
mammary epithelial cell transformation. Nat Struct Mol Biol.
19:220–228. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ghigna C, Giordano S, Shen H, Benvenuto F,
Castiglioni F, Comoglio PM, Green MR, Riva S and Biamonti G: Cell
motility is controlled by SF2/ASF through alternative splicing of
the Ron protooncogene. Mol Cell. 20:881–890. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anczuków O, Akerman M, Cléry A, Wu J, Shen
C, Shirole NH, Raimer A, Sun S, Jensen MA, Hua Y, et al:
SRSF1-regulated alternative splicing in breast cancer. Mol Cell.
60:105–117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Karni R, de Stanchina E, Lowe SW, Sinha R,
Mu D and Krainer AR: The gene encoding the splicing factor SF2/ASF
is a proto-oncogene. Nat Struct Mol Biol. 14:185–193. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Colon Cancer Treatment (PDQ (R)): Health
Professional VersionPDQ Cancer Information Summaries. Bethesda
(MD): 2002
|
|
15
|
de Miguel FJ, Sharma RD, Pajares MJ,
Montuenga LM, Rubio A and Pio R: Identification of alternative
splicing events regulated by the oncogenic factor SRSF1 in lung
cancer. Cancer Res. 74:1105–1115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Das S, Anczukow O, Akerman M and Krainer
AR: Oncogenic splicing factor SRSF1 is a critical transcriptional
target of MYC. Cell Rep. 1:110–117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gonçalves V, Henriques AF, Pereira JF,
Costa Neves A, Moyer MP, Moita LF, Gama-Carvalho M, Matos P and
Jordan P: Phosphorylation of SRSF1 by SRPK1 regulates alternative
splicing of tumor-related Rac1b in colorectal cells. Rna.
20:474–482. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thorsen K, Mansilla F, Schepeler T, Øster
B, Rasmussen MH, Dyrskjøt L, Karni R, Akerman M, Krainer AR,
Laurberg S, et al: Alternative splicing of SLC39A14 in colorectal
cancer is regulated by the Wnt pathway. Mol Cell Proteomics.
10:M110.0029982011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Uhlén M, Fagerberg L, Hallström BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteom. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He X, Ee PL, Coon JS and Beck WT:
Alternative splicing of the multidrug resistance protein 1/ATP
binding cassette transporter subfamily gene in ovarian cancer
creates functional splice variants and is associated with increased
expression of the splicing factors PTB and SRp20. Clin Cancer Res.
10:4652–4660. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He X, Arslan AD, Pool MD, Ho TT, Darcy KM,
Coon JS and Beck WT: Knockdown of splicing factor SRp20 causes
apoptosis in ovarian cancer cells and its expression is associated
with malignancy of epithelial ovarian cancer. Oncogene. 30:356–365.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Amin EM, Oltean S, Hua J, Gammons MV,
Hamdollah-Zadeh M, Welsh GI, Cheung MK, Ni L, Kase S, Rennel ES, et
al: WT1 mutants reveal SRPK1 to be a downstream angiogenesis target
by altering VEGF splicing. Cancer Cell. 20:768–780. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ezponda T, Pajares MJ, Agorreta J,
Echeveste JI, López-Picazo JM, Torre W, Pio R and Montuenga LM: The
oncoprotein SF2/ASF promotes non-small cell lung cancer survival by
enhancing survivin expression. Clin Cancer Res. 16:4113–4125. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oltean S and Bates DO: Hallmarks of
alternative splicing in cancer. Oncogene. 33:5311–5318. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Silipo M, Gautrey H and Tyson-Capper A:
Deregulation of splicing factors and breast cancer development. J
Mol Cell Biol. 7:388–401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McCleland ML, Gardner RD, Kallio MJ, Daum
JR, Gorbsky GJ, Burke DJ and Stukenberg PT: The highly conserved
Ndc80 complex is required for kinetochore assembly, chromosome
congression and spindle checkpoint activity. Genes Dev. 17:101–114.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kobayashi Y, Takano A, Miyagi Y, Tsuchiya
E, Sonoda H, Shimizu T, Okabe H, Tani T, Fujiyama Y and Daigo Y:
Cell division cycle-associated protein 1 overexpression is
essential for the malignant potential of colorectal cancers. Int J
Oncol. 44:69–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hayama S, Daigo Y, Kato T, Ishikawa N,
Yamabuki T, Miyamoto M, Ito T, Tsuchiya E, Kondo S and Nakamura Y:
Activation of CDCA1-KNTC2, members of centromere protein complex,
involved in pulmonary carcinogenesis. Cancer Res. 66:10339–10348.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Numnum TM, Makhija S, Lu B, Wang M, Rivera
A, Stoff-Khalili M, Alvarez RD, Zhu ZB and Curiel DT: Improved
anti-tumor therapy based upon infectivity-enhanced adenoviral
delivery of RNA interference in ovarian carcinoma cell lines.
Gynecol Oncol. 108:34–41. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kaneko N, Miura K, Gu Z, Karasawa H,
Ohnuma S, Sasaki H, Tsukamoto N, Yokoyama S, Yamamura A, Nagase H,
et al: siRNA-mediated knockdown against CDCA1 and KNTC2, both
frequently overexpressed in colorectal and gastric cancers,
suppresses cell proliferation and induces apoptosis. Biochem
Biophys Res Commun. 390:1235–1240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sethi G, Pathak HB, Zhang H, Zhou Y,
Einarson MB, Vathipadiekal V, Gunewardena S, Birrer MJ and Godwin
AK: An RNA interference lethality screen of the human druggable
genome to identify molecular vulnerabilities in epithelial ovarian
cancer. PLoS One. 7:e470862012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shiraishi T, Terada N, Zeng Y, Suyama T,
Luo J, Trock B, Kulkarni P and Getzenberg RH: Cancer/Testis
Antigens as potential predictors of biochemical recurrence of
prostate cancer following radical prostatectomy. J Transl Med.
9:1532011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu P, Shangguan J and Zhang L:
Downregulation of NUF2 inhibits tumor growth and induces apoptosis
by regulating lncRNA AF339813. Int J Clin Exp Pathol. 8:2638–2648.
2015.PubMed/NCBI
|
|
34
|
Sanhaji M, Friel CT, Wordeman L, Louwen F
and Yuan J: Mitotic centromere-associated kinesin (MCAK): A
potential cancer drug target. Oncotarget. 2:935–947. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ishikawa K, Kamohara Y, Tanaka F,
Haraguchi N, Mimori K, Inoue H and Mori M: Mitotic
centromere-associated kinesin is a novel marker for prognosis and
lymph node metastasis in colorectal cancer. Br J Cancer.
98:1824–1829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Di Domenico EG, Romano E, Del Porto P and
Ascenzioni F: Multifunctional role of ATM/Tel1 kinase in genome
stability: from the DNA damage response to telomere maintenance.
Biomed Res Int. 2014:7874042014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang J, Wu Q, Zhang LH, Zhao YX and Wu X:
The role of RhoA in vulvar squamous cell carcinoma: A
carcinogenesis, progression, and target therapy marker. Tumour
Biol. 37:2879–2890. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jeong D, Park S, Kim H, Kim CJ, Ahn TS,
Bae SB, Kim HJ, Kim TH, Im J, Lee MS, et al: RhoA is associated
with invasion and poor prognosis in colorectal cancer. Int J Oncol.
48:714–722. 2016. View Article : Google Scholar : PubMed/NCBI
|