Introduction
When tumors emerge in the human body, the host
immune surveillance should recognize tumor-specific antigens and
eliminate the tumor through the tumor-specific T cell response
(1). However, the response is
ineffective due to an imbalance of co-stimulatory molecules in the
tumor microenvironment, which could determine the functional
maturation of T cells (2,3). Co-stimulatory members of the B7 family
has seven known members: B7.1 (CD80), B7.2 (CD86), B7-H3, B7-H4,
inducible costimulator ligand (ICOS-L), programmed death-1 ligand
and programmed death-2 ligand, which bind to receptors on
lymphocytes that to exert both inhibitory and stimulatory effects
on T cell activation (4). These
members were found to be abnormally expressed in various human
malignancies in previous years, and were considered crucial targets
in cancer immunotherapy (5). B7
homolog 3 protein (B7-H3), also termed cluster of differentiation
(CD)276, is one of B7 family members (6). B7-H3 expression is found on activated T,
natural killer and antigen-presenting cells (6,7), but
hardly found in normal organ tissues. The function of B7-H3 remains
a paradox. Both stimulatory and inhibitory effects were observed in
T cell activation and antitumor immune response by different
studies (8). Diverse associations
between the expression of B7-H3 in tumor tissues and clinical
parameters have also been reported (8). The contrasting results revealed that
there may be two or more receptors for B7-H3 with opposing
functions, which remain unknown (9).
The mechanism of B7-H3 in the tumor microenvironment remains
unclear, and the regulation of T cell activation, resistance of
drug-induced apoptosis and enhancement of metastasis were all
considered as probable mechanisms (10). However, the exact role of B7-H3 in the
tumor microenvironment remains unclear.
Tumor-associated macrophages (TAMs) are a major
leukocyte population in tumor tissues, and are involved in tumor
growth, invasion and metastasis (11–21). TAM
infiltration has been demonstrated to be associated with a poor
prognosis and outcome in numerous human malignancies (13–15).
Therefore, the manipulation of TAMs may be extremely valuable in
antitumor therapy. Macrophages are highly heterogeneous cells that
are induced into distinct functional states under various
circumstances. The type 1 macrophage (M1) phenotype is induced by
lipopolysaccharide (LPS) and interferon-γ, and exhibits the ability
to kill pathogens and tumor cells. The M2 phenotype is induced by
interleukin (IL)-4, IL-13 and IL-10, and is capable of suppressing
inflammation and promoting tissue repair, tumor growth and
angiogenesis (16–18). TAMs generally show polarized M2
phenotypes and elicit similar function in the tumor
microenvironment, including promotion of angiogenesis, matrix
remodeling and suppression of antitumor immunity (19–21). The
tumor microenvironment is hypothesized to educate TAMs towards an
M2 polarization, but the mechanisms underlying this are not fully
understood.
Colorectal carcinoma (CRC) is one of the most
frequent malignancies occurring in human beings, and ranks as the
third most common cancer and the fourth leading cause of
cancer-associated mortality worldwide (22). In China, CRC has been reported to have
an increasing incidence due to changes of diets and lifestyles
(23). Various therapeutic
strategies, including surgery, chemotherapy, radiotherapy and
immunotherapy, are used to treat patients with CRC, but the
therapies may lead to different outcomes. Therefore, key factors
that regulate tumor progression and antitumor immunity are required
in the individualized therapy of human CRC. A previous study has
demonstrated the clinical significance and regulation of B7-H3
expression in CRC patients (10). In
the present study, we investigated the immune cells that
infiltrated in CRC, and revealed the mechanism of B7-H3 signaling
in the regulation of immune cells in the tumor
microenvironment.
Materials and methods
Patients
In total, 117 CRC patients who underwent surgery
between January 2003 and December 2003 at the Department of
Gastrointestinal Surgery (the Fourth Affiliated Hospital, Suzhou
University, Wuxi, China) were enrolled in the present study. No
patient received pre-operative chemotherapy or radiotherapy, and
all specimens were identified as CRC under hematoxylin and eosin
staining. The paraffin-embedded blocks of tumor tissues were
obtained from the archival collections at the Department of
Pathology (the Fourth Affiliated Hospital, Suzhou University), and
the survival data of patients were collected by the end of December
2008. Fresh resected CRC tissues were obtained from the First
Affiliated Hospital, Suzhou University (Suzhou, China), and were
used for the purification of TAMs immediately subsequent to
surgery. The present study was approved by the Medical Ethics
Committee (the Fourth Affiliated Hospital, Soochow University) and
all patients provided informed consent.
Immunohistochemical staining and
evaluation
Immunohistochemistry was performed using the Dako
Envision™ (Agilent Technologies, Inc., Santa Clara, CA, USA),
according to the manufacturer's instructions and as previously
described (24). Mouse anti-human
B7-H3 [clone no. 21D4; established and characterized in our
institute (Soochow University) (24)], and CD8 and CD68 (cat. nos. MAB-0021
and Kit-0026 ready for use; Fuzhou Maxim Biotech Co., Ltd., Fuzhou,
China) monoclonal antibodies were used to stain for B7-H3
expression, CD8+ T cell infiltration and
CD68+ macrophage infiltration, respectively. The
staining was evaluated as previously described (24). The B7-H3 immunostaining densities were
assessed according to the H-score method described by Hammes et
al (25). Assessment of the
infiltration densities of CD8+ T cells and
CD68+ macrophages was performed in both the tumor stroma
and tumor nest. Tumor-infiltrating lymphocytes (TILs) in the tumor
stroma were examined and categorized according to the density as
follows: Grade 0, sparse; grade 1, moderate infiltration; grade 2,
abundant infiltration; and grade 3, massive infiltration. The group
consisting of grades 0 and 1 infiltration was defined as the low
infiltration group, and the group consisting of grades 2 and 3
infiltration was defined as the high infiltration group. TILs in
the tumor nest were counted and recorded using Image-Pro Plus 6.0
(Olympus, Tokyo, Japan) under high power field (×200
magnification). The results from the five areas were averaged and
used in the statistical analysis. The median value of all sections
was set as the cut-off value, as described in our previous study
(26), to categorize the low and high
infiltration densities in the tumor nest. The total density of
lymphocyte infiltration was determined by the summary evaluation of
both the tumor nest and stroma.
Purification of monocytes from
peripheral blood mononuclear cells (PBMCs) and TAMs obtained from
CRC tissue
PBMCs were isolated by Ficoll-Hypaque gradient
centrifugation from the peripheral blood of healthy donors (Suzhou
Central Blood Bank, Suzhou, China). Monocytes were purified with a
CD14-positive selection kit (Stemcell Technologies, Inc.,
Vancouver, BC, Canada). The purity of the monocyte preparation was
95%. The isolated monocytes were then incubated with LPS (1 mg/ml)
for up to 48 h for putative B7-H3 receptor detection.
TAMs were isolated from fresh resected CRC tissues.
Tumor specimens were gently minced over a wire mesh screen, and
then digested with collagenase IV (1 mg/ml; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) at 37°C for 1 h on a shaking platform to
obtain a cell suspension. TAMs were then insolated with the
CD14-positive selection kit.
Putative B7-H3 receptor detection
Stimulated monocytes, induced human monocyte THP-1
cells (American Type Culture Collection, Manassas, VA, USA)
stimulated with 1 mg/ml phorbol 12-myristate 13-acetate (PMA;
Sigma-Aldrich; Merck KGaA) for 48 h, and purified CRC TAMs were
firstly incubated with human AB serum (10 ml/105 cells)
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at
4°C for 30 min to block the Fc receptor (FcR). The cells were then
stained with biotinylated hB7-H3 Ig (100 ng/ml) or biotinylated
human IgG (100 ng/ml) (both from R&D Systems, Inc.,
Minneapolis, MN, USA) as the control, then the putative B7-H3
receptor was analyzed using flow cytometer and Diva software
(version 6.1.2; BD Biosciences, San Jose, CA, USA).
Tumor cell culture supernatants
For tumor supernatant (TSN) collection,
5×106 human CRC SW480 cells (Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences, Shanghai, China)
were seeded to the flask for 1–2 day growth at 37°C. When cells
reached 70–80% confluence, fresh medium was added for another 24 h
growth. The culture medium was harvested by centrifugation (1,500 ×
g for 10 min at 4°C), and filtration, and used as a stimulus.
Differentiation and polarization of
macrophage cells with B7-H3
Purified human peripheral blood monocytes were used
for the induction of macrophages. In total, 5×106 cells
were cultured in the conditioned medium containing 10 ng/ml PMA for
macrophage differentiation. After 1 day, the cells were thoroughly
washed with PBS 3 times to remove the remaining PMA, and then
re-seeded to the plates overnight. To polarize macrophages, 30% TSN
was added to complete RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) for macrophage culture for 7 days. hB7-H3 Ig or
control human Ig (5 ìg/ml; R&D Systems, Inc.) was added during
the polarization of cells. The cells and supernatant were then
collected for further study.
Macrophage phenotype and cytokine
secretion analysis
The induced macrophages from different culture
conditions were collected and firstly incubated with human AB serum
to block the FcR, and then stained with phycoerythrin-labeled
anti-CD206 antibody (cat. no. 321105; BioLegend, Inc., San Diego,
CA, USA) and fluorescein isothiocyanate-labeled anti-human
leukocyte antigen-antigen D related antibody (cat. no. IM0463U,
HLA-DR; Beckman Coulter, Inc., Brea, CA, USA) (1 µg/105
cells), the cells was ultimately analyzed by FACScan to determine
the macrophage phenotype. The culture supernatant of different
groups was collected by centrifugation (1,500 × g for 10 min at
4°C). ELISA was performed to detect the concentration of cytokine
IL-10, IL-12p70 and TNF-α using the Human IL-10 Quantikine ELISA
kit (cat. no. D1000B), Human IL-12p70 Quantikine ELISA kit (cat.
no. D1200), Human TNF-α Quantikine ELISA kit (cat. no. DTA00C;
R&D Systems, Inc.). Reverse transcription-polymerase chain
reaction (RT-PCR) was performed to analyze inducible nitric oxide
synthase (iNOS) RNA expression. RNA was extracted from the induced
macrophages using TRIzol reagent (Takara Bio, Inc., Shiga, Japan)
and converted into cDNA with an oligo(dT) primer using the
PrimeScript First Strand cDNA Synthesis kit at 42°C for 1 h (Takara
Bio, Inc.). The sequences of the primers for iNOS were as follows:
Forward, 5′-TCCGAGGCAAACAGCACATTCA-3′ and reverse,
5′-GGGTTGGGGGTGTGGTGATGT-3′. The primers were synthesized by
Invitrogen (Thermo Fisher Scientific, Inc.) and the PCR reaction
conditions for amplification of DNA were as follows: Initial
denaturation at 98°C for 2 min, followed by 30 cycles of annealing
commencing at 65°C and ending at 55°C for 15 sec and extension at
68°C for 30 sec.
Statistical analysis
B7-H3 expression and the densities of immune cells
in association with the postoperative prognosis of CRC patients
were examined by log-rank survival analysis. The association
between B7-H3 expression and densities of immune cell infiltration
were analyzed by χ2. The comparison of cytokine
secretion from induced macrophages was analyzed using Student's
t-test. All statistical tests were two tailed. All statistical
analyses were performed using GraphPad Prism 5.0 software (GraphPad
Software, Inc., San Diego, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
B7-H3 expression and infiltrated
macrophage density were associated with the survival of CRC
patients
Immunohistochemical staining showed that B7-H3
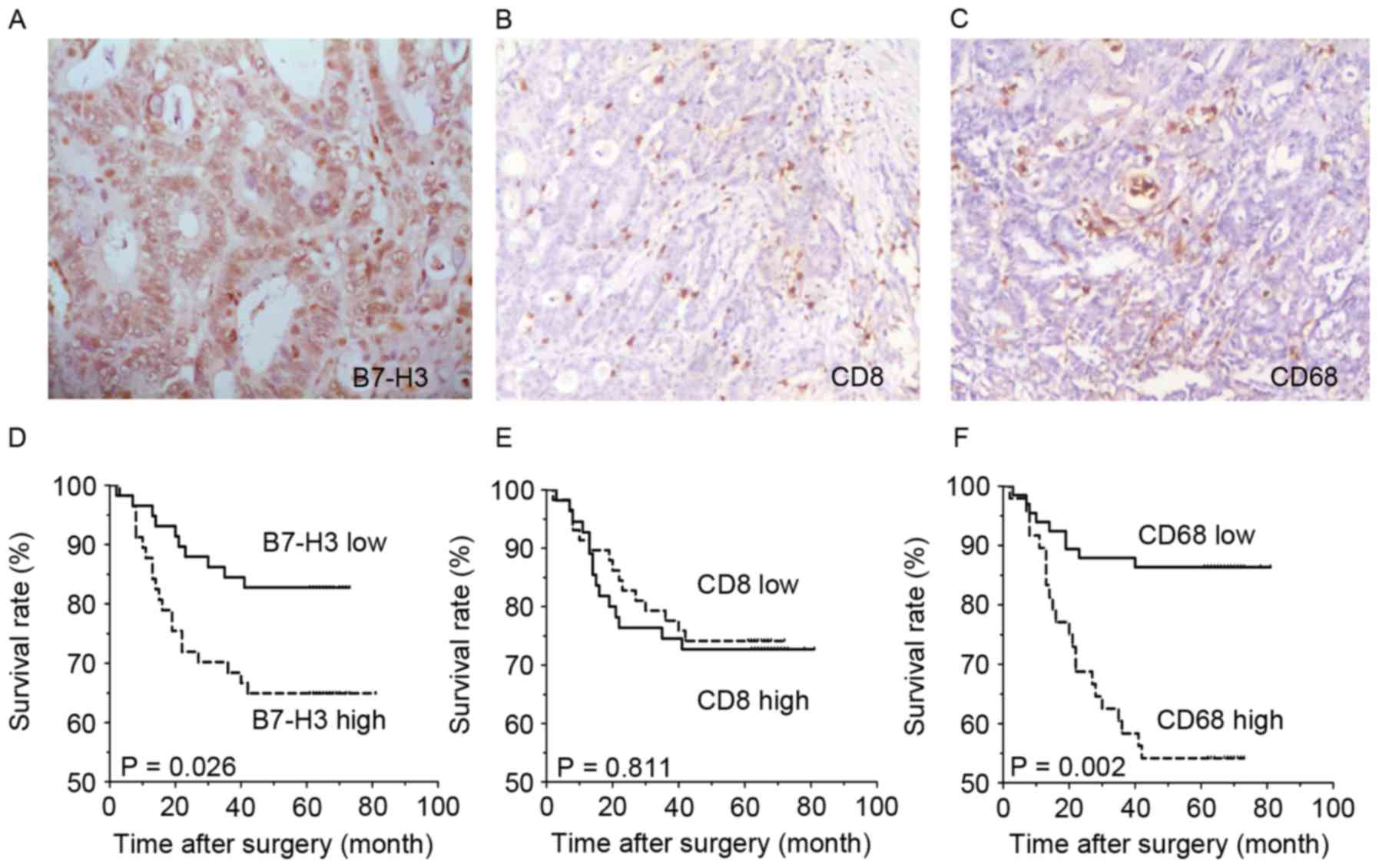
expression was present in tumor cells of CRC tissues (Fig. 1A), and was located in the cell
membrane, cytoplasm and nucleus. B7-H3 expression of 117 patients
was categorized into two major subgroups, according to the H-score
of immunohistochemical staining, as described in previous studies
(24,26). In total, 113 out of 117 (96.6%) CRC
tissues showed positive B7-H3 immunohistochemical staining, and 59
cases with high expression and 58 cases with low expression.
CD8-labeled effective T cells and CD68-labeled macrophages were
also found to have infiltrated all CRC tissues (Fig. 1B and C), both in the tumor nest and
stroma. The infiltration density was also categorized into two
major subgroups (Table I), according
to the criterion in previous studies (24,26).
Survival analysis demonstrated that the overall survival rate of
the subgroup with low B7-H3 expression was significantly improved
compared with the high B7-H3 expression subgroup (P=0.0262;
Fig. 1D). Additionally, the density
of infiltrating CD68+ macrophages was negatively
associated with CRC survival. The survival rate of patients with a
low infiltrating density of was improved compared with the patients
with a high infiltration density of macrophages (P=0.002: Fig. 1F), while the infiltration density of
CD8+ T cells was not significantly associated with the
overall survival rate of CRC patients.
 | Table I.Association between B7-H3 expression
and lymphocytes infiltration in colorectal cancer tissue. |
Table I.
Association between B7-H3 expression
and lymphocytes infiltration in colorectal cancer tissue.
|
|
| B7-H3
expression |
|
|---|
|
|
|
|
|
|---|
| Infiltration | Cases, n | Low, n (%) | High, n (%) | P-value |
|---|
| CD8+ T
cells |
|
|
|
|
| Tumor
stroma |
| 58 (49.6) | 59 (50.4) | 0.268 |
|
Low density | 55 | 24 (43.6) | 31 (56.4) |
|
|
High density | 62 | 34 (54.8) | 28 (45.2) |
|
| Tumor
nest |
|
|
| 0.095 |
|
Low density | 52 | 21 (40.4) | 31 (59.6) |
|
|
High density | 65 | 37 (56.9) | 28 (43.1) |
|
| CD68+
TAMs |
|
|
|
|
| Tumor
stroma |
| 58 (49.6) | 59 (50.4) | 0.039 |
|
Low density | 69 | 40 (58.0) | 29 (42.0) |
|
|
High density | 48 | 18 (37.5) | 30 (62.5) |
|
| Tumor
nest |
|
|
| 0.005 |
|
Low density | 61 | 38 (62.3) | 23 (37.7) |
|
|
High density | 56 | 20 (35.7) | 36 (64.3) |
|
B7-H3 expression was associated with
the density of infiltrating CD68+ macrophages, but not
infiltrating CD8+ T cells
The mechanism of how the B7-H3 pathway regulates the
immune response remains unknown; therefore, the association between
B7-H3 expression and density of infiltrated lymphocytes was
analyzed. As the statistical analysis shows in Table I, the level of cancer cell expression
of B7-H3 was found to be positively associated with the density of
infiltrated macrophages in both the tumor nest and tumor stroma
(P=0.039 and P=0.005, respectively; Table
I). The subgroups with increased B7-H3 expression contained
more cases with increased density of macrophage infiltration. No
significant association was found between B7-H3 expression and the
infiltrating density of CD8+ T cells (Table I).
B7-H3 putative receptor could be
detected on activated monocytes and TAMs of CRC
PBMC-derived monocytes were isolated from the
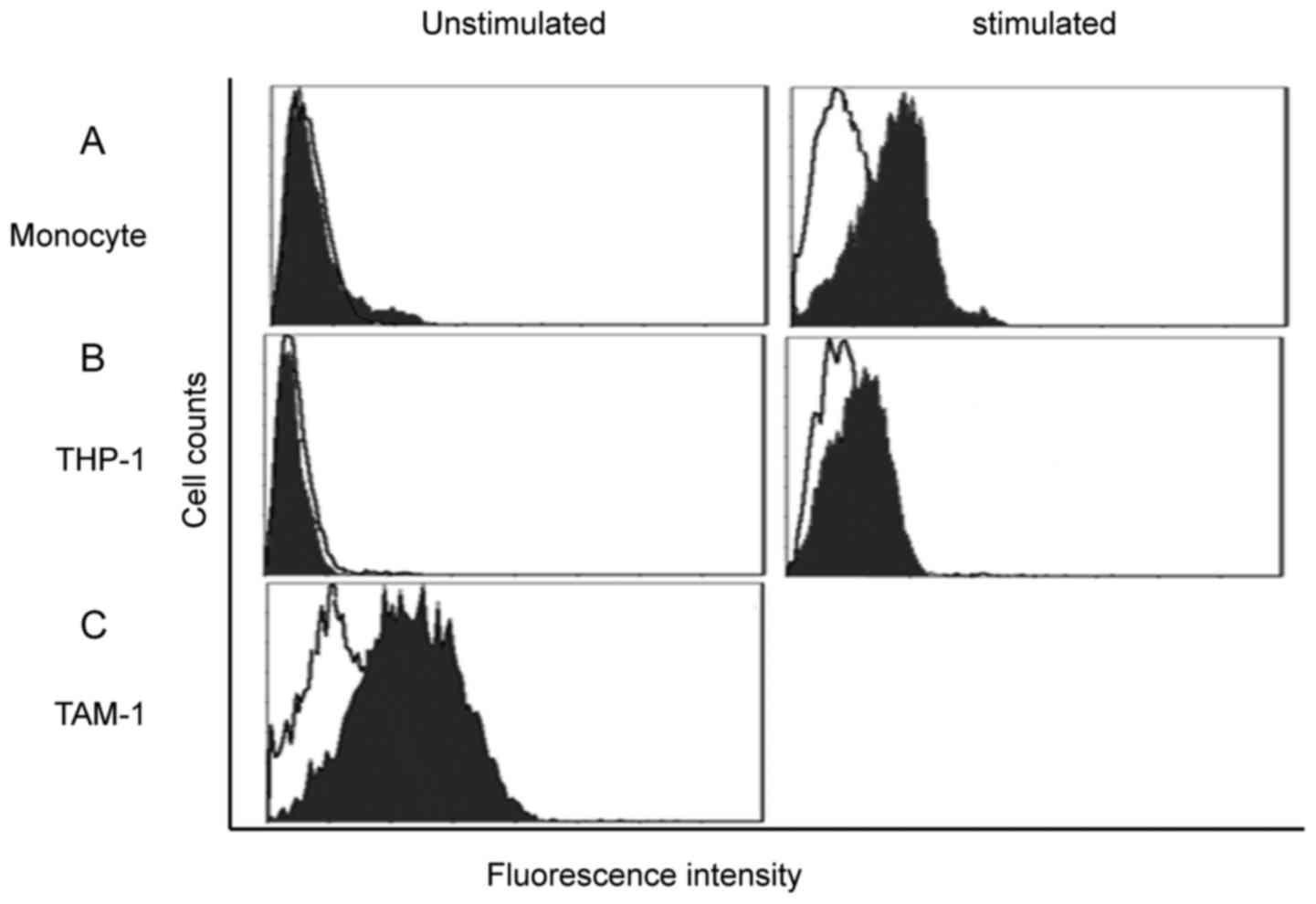
peripheral blood of healthy donors; the purification reached 95%.
FACScan showed that unstimulated monocytes had extremely low
expression of the B7-H3 receptor. Subsequent to 48 h of LPS
stimulation, the B7-H3 receptor was detected on the monocytes
(Fig. 2A). THP-1 cells were also
investigated; PMA was used for macrophage induction. Similarly, the
putative B7-H3 receptor was not detected on THP-1 cells, but the
expression was increased subsequent to PMA induction, which
indicates that the putative B7-H3 receptor was expressed by
macrophages (Fig. 2B). TAMs were
obtained directly from the CRC tissue, and FACScan showed that
B7-H3 receptor was highly expressed by TAMs (Fig. 2C).
B7-H3 signal could promote the M2-like
macrophage polarization induced by TSN
Based on the results of B7-H3 receptor detection,
monocytes purified from the PBMCs of healthy donors were assessed.
Monocytes were induced into macrophages by PMA stimulation and then
polarized by TSN with or without B7-H3. Surface molecules and
cytokine secretion were analyzed to determine the phenotype of
polarized macrophages. The results show that compared to TSN
polarization only, the combination of B7-H3 and TSN promoted CD206
expression and inhibited the HLA-DA expression on macrophages
(Fig. 3A). This indicated a M2
macrophage phenotype. ELISA showed that the IL-10 concentration was
increased when adding B7-H3 into the TSN polarization (P=0.0087),
but TNF-α concentration was significantly decreased meanwhile
(Fig. 3B) (P=0.0016). RT-PCR results
indicated that iNOS RNA expression was evidently inhibited by B7-H3
combined with TSN polarization compared with the TSN polarization
alone (Fig. 3C). All these results
suggest that B7-H3 signaling may promote the polarization of
M2-like macrophages induced by TSN.
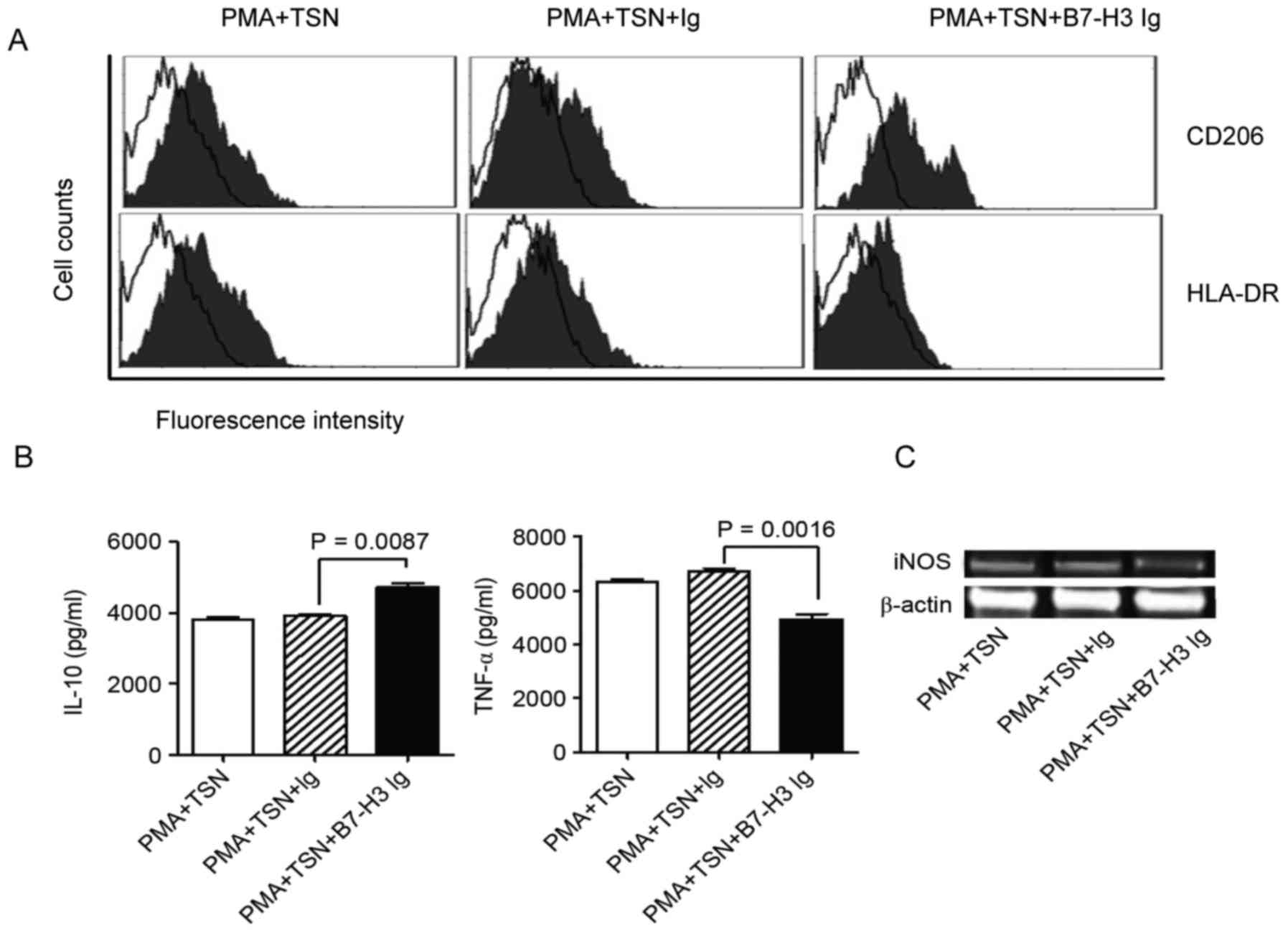 | Figure 3.The phenotype and cytokine secretion
of polarized macrophages. PBMC-derived monocytes were used for
macrophage induction and differentiation. PMA was used for the
induction of macrophages, and TSN and B7-H3 were used for
macrophage polarization. (A) Surface molecules (CD206 and HLA-DR)
were examined by flow cytometry to identify the phenotype of
macrophages. The figure presents overlaps of CD206/HLA-DR staining
and control Ig staining. (B) Cytokines (IL-10 and TNF-α) were
detected by ELISA, and (C) iNOS RNA expression was detected by
reverse transcription-polymerase chain reaction to analyze the
function of macrophages. All results together indicated that B7-H3
could promote M2 macrophage differentiation. CRC, colorectal
cancer; B7-H3, B7 homolog 3 protein; PBMCs, peripheral blood
mononuclear cells; TAMs, tumor-associated macrophages; PMA, phorbol
12-myristate 13-acetate; TSN, tumor supernatant; CD, cluster of
differentiation; HLA-DR, human leukocyte antigen-antigen D related;
IL-10, interleukin; TNF-α, tumor necrosis factor-α; iNOS, inducible
nitric oxide synthase. |
Discussion
In our previous study, B7-H3 expression in CRC
tissues and its clinical significance were investigated (24). In the present study, the prognostic
value of the B7-H3 expression in CRC was further examined, and it
was found that the overall survival rate of patients with lower
B7-H3 expression was improved compared with those with increased
B7-H3 expression. This finding confirmed that B7-H3 may be a useful
indicator for the prognostic prediction of human CRC. The mechanism
of B7-H3 signal involved in tumor progression remains unclear. B7
family members have important roles in the regulation of the tumor
immune response and tumor suppressor microenvironment. The
overexpression of B7-H3 and other B7 family molecules, and certain
immune cells, contribute to the shaping of the tumor
microenvironment (27); therefore,
the present study investigated the association between B7-H3
expression and tumor-associated immune cells.
Effective immune cells, particularly T cells and
macrophages, are immune effector cells of the antitumor immune
response. CD8+ T cells and CD68+ macrophages
were hypothesized to play important roles in the host immune
response against tumors. While previous studies have shown that the
characteristics of these cells, particularly the cytolytic and
regulatory nature of effective immune cells were transformed when
existing in different microenvironments (1,27). To
determine the roles of immune cells in CRC, the present study
detected the infiltration density of infiltrated CD8+ T
cells and CD68+ macrophages and examined the prognostic
value of as well. As showed in Fig.
1, we found that the infiltrating densities of CD8+
T cell had no association with the survival rate of CRC patients,
while the infiltrating densities of CD68+ macrophage was
significantly correlated to the patients survival rate. Patients
with a low macrophage infiltration density had a significantly
improved survival rate compared with patients with a high
macrophage infiltration density. The immune function of
CD4+ and CD8+ T cells were inhibited in
certain diseases, including tumors and infections (28). The tumor-driven exhaustion of
CD8+ T cells was associated with certain B7 family
members, including PD-1 and BTLA (29). Dysfunction of CD8+ T cells
leads to a failed antitumor immune response, and a high density of
CD8+ T cells in esophageal tumor tissue does not result
in improved outcomes for patients (26). TAMs are heterogeneous in response to
environmental signals and generally exhibit similarities with
polarized M2 macrophages (11,12).
Increased TAM infiltration has been demonstrated to be associated
with poorer prognosis in breast and ovarian cancer (30,31). The
present results identified in patients with CRC were consistent
with those of previous studies (10,24), and
identified that the macrophages that infiltrate CRC tissues may
have a harmful role in tumor immune response. The association
between B7-H3 expression and expression of CD8+ T cells
and CD68+ macrophages was then investigated. As shown in
Table I, the B7-H3 expression level
was not associated with CD8+ T cell infiltration
density, but was positively associated with the CD68+
macrophage infiltration density, indicating that the B7-H3 signal
was involved in the regulation of TAMs.
As a tumor-associated antigen, B7-H3 has dual
function in tumor progression, which requires additional
investigation to clarify the mechanism of B7-H3 signaling in the
tumor microenvironment. The unknown counter-receptor is the key to
illustrate the regulation mechanism of the B7-H3 function (9,10). Thus,
the identification of the B7-H3 receptor may be attempted on
tumor-associated cells. According to the aforementioned results,
B7-H3 expression was associated with macrophage infiltration.
Subsequently, the expression of the putative receptor of B7-H3 on
the monocytes and macrophages was then investigated. The B7-H3
receptor was detected on monocytes isolated from PBMCs and THP-1
cells. LPS was then used to activate monocytes, and PMA was used to
induce THP-1 cells into macrophages. The B7-H3 receptor was
detected on the activated monocytes and induced THP-1 cells, which
demonstrated that the expression of the putative B7-H3 receptor
could be induced by activated monocytes and macrophages. TAMs in
tumor tissues are a group of activated macrophages. TAMs were
isolated from CRC tissue, and the B7-H3 receptor was highly
expressed by TAMs. The expression of the putative B7-H3 receptor on
macrophages indicated that the B7-H3 signal may participate in the
regulation of TAMs. TAMs are the major component of tumor
inflammatory infiltration. It has been identified that TAMs are
heterogeneous, plastic cells with different functions and cytokine
production in response to various signals in the tumor
microenvironment (16,18).
Studies of numerous human tumors showed that TAMs
generally exhibit M2 phenotypes, and also function as the
immunosuppressive cell subset that promotes tumor growth, migration
and metastasis. Thus, identifying the key factors that modulate TAM
differentiation is crucial for inhibiting TAM-mediated promotion of
tumor growth. To assess the role of B7-H3 in macrophage
differentiation in the tumor microenvironment, TSN of CRC cells was
prepared and used for the induction and polarization of
macrophages. Monocytes were induced by PMA and polarized by TSN
with or without human B7-H3 Ig. As shown in Fig. 3, when induced and polarized with the
TSN and B7-H3 combination, the surface marker CD206 was increased
and HLA-DR was decreased compared to TSN use only, exhibiting a M2
phenotype. Meanwhile, IL-10 secretion was increase and TNF-α
secretion was decreased, also suggesting a M2 character. NO produce
is important feature of M1, which could be reflected by iNOS RNA
expression (16,18). The iNOS RNA expression was evidently
inhibited by co-culture with B7-H3 Ig and TSN compared with TSN use
only, which indicated the switch between M1 and M2. Overall, the
results from the present study support an important role of B7-H3
in M2 macrophage polarization in the tumor microenvironment. How to
manipulate the differentiation and phenotypical switch of TAMs
through the B7-H3 signal requires additional investigation.
In conclusion, the present study indicates that
B7-H3 expression and TAM density in tumor tissue are valuable
prognostic indicators in patients with CRC. The B7-H3 signal could
promote M2 macrophage differentiation via the putative receptor on
activated monocytes and macrophages. Thus, targeting the
manipulation of TAMs through the B7-H3 pathway may be valuable for
the development of a new strategy for antitumor therapy in CRC.
Acknowledgements
The authors thank Dr Yuyu Wu (Department of
Pathology, The Fourth People's Hospital of Wuxi, Wuxi, China) for
her suggestions and technical assistance. This study was supported
by the National Natural Science Foundation of China (grant nos.
31100634, 81201600, 81301960, 81372375 and 81502042), National
Natural Science Foundation of Jiangsu (grant nos. BK2012538 and
BK20140171) and Science and Technology plan of Suzhou (grant no.
SYS201523).
References
|
1
|
Finn OJ: Immuno-oncology: Understanding
the function and dysfunction of the immune system in cancer. Ann
Oncol. 23 Suppl 8:viii6–viii9. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marabelle A, Kohrt H, Caux C and Levy R:
Intratumoral immunization: A new paradigm for cancer therapy. Clin
Cancer Res. 20:1747–1756. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Turtle CJ, Hudecek M, Jensen MC and
Riddell SR: Engineered T cells for anti cancer therapy. Curr Opin
Immunol. 24:633–639. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Poirier N, Blancho G and Vanhove B: A more
selective costimulatory blockade of the CD28-B7 pathway. Transpl
Int. 24:2–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maj T, Wei S, Welling T and Zou W: T cells
and costimulation in cancer. Cancer J. 19:473–482. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chapoval AI, Ni J, Lau JS, Wilcox RA,
Flies DB, Liu D, Dong H, Sica GL, Zhu G, Tamada K and Chen L:
B7-H3: A costimulatory molecule for T cell activation and IFN-gamma
production. Nat Immunol. 2:269–274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suh WK, Gajewska BU, Okada H, Gronski MA,
Bertram EM, Dawicki W, Duncan GS, Bukczynski J, Plyte S, Elia A, et
al: The B7 family member B7-H3 preferentially down regulates T
helper type 1-mediated immune responses. Nat Immunol. 4:899–906.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Kang FB and Shan BE:
B7-H3-mediated tumor immunology: Friend or foe? Int J Cancer.
134:2764–2771. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hashiguchi M, Kobori H, Ritprajak P,
Kamimura Y, Kozono H and Azuma M: Triggering receptor expressed on
myeloid cell-like transcript 2 (TLT-2) is a counter-receptor for
B7-H3 and enhances T cell responses. Proc Natl Acad Sci USA.
105:10495–10500. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Loos M, Hedderich DM, Friess H and Kleeff
J: B7-h3 and its role in antitumor immunity. Clin Dev Immunol.
2010:6838752010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Riabov V, Gudima A, Wang N, Mickley A,
Orekhov A and Kzhyshkowska J: Role of tumor associated macrophages
in tumor angiogenesis and lymphangiogenesis. Front Physiol.
5:752014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lewis CE and Pollard JW: Distinct role of
macrophages in different tumor microenvironments. Cancer Res.
66:605–612. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bingle L, Brown NJ and Lewis CE: The role
of tumor-associated macrophages in tumor progression: Implications
for new anticancer therapies. J Pathol. 196:254–265. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karnevi E, Andersson R and Rosendahl AH:
Tumour-educated macrophages display a mixed polarisation and
enhance pancreatic cancer cell invasion. Immunol Cell Biol.
92:543–552. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou W and Bao S: Reciprocal supportive
interplay between glioblastoma and tumor-associated macrophages.
Cancers (Basel). 6:723–740. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gordon S and Taylor PR: Monocyte and
macrophage heterogeneity. Nat Rev Immunol. 5:953–964. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mantovani A, Sica A, Sozzani S, Allavena
P, Vecchi A and Locati M: The chemokine system in diverse forms of
macrophage activation and polarization. Trends Immunol. 25:677–686.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mosser DM and Edwards JP: Exploring the
full spectrum of macrophage activation. Nat Rev Immunol. 8:958–969.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hagemann T, Wilson J, Burke F, Kulbe H, Li
NF, Plüddemann A, Charles K, Gordon S and Balkwill FR: Ovarian
cancer cells polarize macrophages toward a tumor-associated
phenotype. J Immunol. 176:5023–5032. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sica A, Schioppa T, Mantovani A and
Allavena P: Tumor-associated macrophages are a distinct M2
polarised population promoting tumor progression: Potential targets
of anti-cancer therapy. Eur J Cancer. 42:717–727. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mantovani A, Allavena P and Sica A:
Tumor-associated macrophages as a prototypic type II polarised
phagocyte population: Role in tumor progression. Eur J Cancer.
40:1660–1667. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weitz J, Koch M, Debus J, Höhler T, Galle
PR and Büchler MW: Colorectal cancer. Lancet. 365:153–165. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong ZW QY, Li LD, Chen YD, Wang RT and
Lei TH: Report of Chinese cancer control strategy. Chin Cancer.
11:250–60. 2002.
|
|
24
|
Sun J, Chen LJ, Zhang GB, Jiang JT, Zhu M,
Tan Y, Wang HT, Lu BF and Zhang XG: Clinical significance and
regulation of the costimulatory molecule B7-H3 in human colorectal
carcinoma. Cancer Immunol Immunother. 59:1163–1171. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hammes LS, Tekmal RR, Naud P, Edelweiss
MI, Kirma N, Valente PT, Syrjänen KJ and Cunha-Filho JS:
Up-regulation of VEGF, c-fms and COX-2 expression correlates with
severity of cervical cancer precursor (CIN) lesions and invasive
disease. Gynecol Oncol. 110:445–451. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen LJ, Sun J, Wu HY, Zhou SM, Tan Y, Tan
M, Shan BE, Lu BF and Zhang XG: B7-H4 expression associates with
cancer progression and predicts patient's survival in human
esophageal squamous cell carcinoma. Cancer Immunol Immunother.
60:1047–1055. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Flies DB and Chen L: Modulation of immune
response by B7 family molecules in tumor microenvironments. Immunol
Invest. 35:395–418. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schietinger A and Greenberg PD: Tolerance
and exhaustion: Defining mechanisms of T cell dysfunction. Trends
Immunol. 35:51–60. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fourcade J, Sun Z, Pagliano O, Guillaume
P, Luescher IF, Sander C, Kirkwood JM, Olive D, Kuchroo V and
Zarour HM: CD8(+) T cells specific for tumor antigens can be
rendered dysfunctional by the tumor microenvironment through
up-regulation of the inhibitory receptors BTLA and PD-1. Cancer
Res. 72:887–896. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Leek RD and Harris AL: Tumor-associated
macrophages in breast cancer. J Mammary Gland Biol Neoplasia.
7:177–189. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hagemann T, Wilson J, Burke F, Kulbe H, Li
NF, Plüddemann A, Charles K, Gordon S and Balkwill FR: Ovarian
cancer cells polarize macrophages toward a tumor-associated
phenotype. J Immunol. 176:5023–5032. 2006. View Article : Google Scholar : PubMed/NCBI
|