Introduction
Prostate cancer (PC) is the most common cancer in
men and the second leading cause of cancer-associated mortality in
USA (1). PC is challenging to treat
due to its high metastasis rate (2);
despite considerable advances in the treatment of PC, metastatic
prostate cancer (MPC) remains incurable (3). The molecular mechanisms underlying the
metastasis of PC cells remain largely unknown and require further
study.
MicroRNAs (miRNAs/miRs) are a group of non-coding
RNAs of 17–27 nucleotides in length that regulate gene expression
by binding to the 3′untranslated regions of messenger RNAs (mRNAs)
(4). miRNAs have been demonstrated to
serve important roles in a number of cellular processes as
post-transcriptional regulators, in addition to roles in cancer
development and progression (5,6).
Dysregulation of miRNAs has been demonstrated to contribute to
tumorigenesis by stimulating proliferation, angiogenesis and
invasion (7,8). Previous studies have investigated miRNA
expression profiles in primary prostate cancer (PPC) or MPC and
several miRNAs have been suggested as diagnostic markers for PC
(9,10). However, the molecular mechanisms
underlying the roles of miRNAs and their target differentially
expressed genes (DEGs) in PC metastasis remain unclear.
Based on 218 prostate tumor samples (181 primaries
and 37 metastases), Taylor et al (11) conducted an integrated analysis
(including concordant assessment of DNA copy number, mRNA
expression and focused exon resequencing), and revealed that
nuclear receptor coactivator NCOA2 functions as an oncogene
in ~11% of PC tumors, and FOXP, RYBP and SHQ1
serve as potential cooperative tumor suppressors in human PC. Using
the same microarray datasets, the aim of the present study was to
identify miRNAs and DEGs that are associated with the metastasis of
PC cells by screening miRNAs and genes that are differentially
expressed between MPC and PPC samples, with the objective to
further understand the molecular mechanisms of MPC.
Materials and methods
Source of microarray data
The raw microarray datasets GSE21036 and GSE21034
were downloaded from the Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo/) database. The
miRNA expression dataset GSE21036 was collected from 141 patients
with PC treated by radical prostatectomy, including 14 metastatic
samples, 99 primary non-metastatic tumor samples and 28 normal
adjacent benign prostate samples. The annotation platform was the
Agilent-019118 Human miRNA Microarray 2.0 G4470B (miRNA ID version)
(Agilent Technologies, Inc., Santa Clara, CA, USA). The GSE21034
transcript dataset was collected form 179 samples, including 29
normal samples, 131 primary samples and 19 metastatic samples. The
annotation platform was the Affymetrix Human Exon 1.0 ST Array
(Affymetrix, Inc., Santa Clara, CA, USA). As illustrated in
Table I, 139 samples from datasets of
GSE21036 and GSE21034 were overlapped and used for the following
analyses.
 | Table I.Microarray datasets used from the
Gene Expression Omnibus database, and the proportions of
metastatic, primary and normal samples in each dataset. |
Table I.
Microarray datasets used from the
Gene Expression Omnibus database, and the proportions of
metastatic, primary and normal samples in each dataset.
| Microarray
dataset | Metastatic | Primary | Normal | Total |
|---|
| miRNA
(GSE21036) | 14 | 99 | 28 | 141 |
| Transcript
(GSE21034) | 19 | 131 | 29 | 179 |
| miRNA and
transcript (common to GSE21036 and GSE21034) | 13 | 98 | 28 | 139 |
Microarray data pre-processing
Using the AgiMicroRna Bioconductor library of R
(http://bioconductor.org/help/search/index.html?q=AgiMicroRna),
the GSE21036 raw data were subject to pre-processing as previously
described (12), including removing
the probes with a low detection rate (failure in >75% samples),
background adjustment and quantile normalization by the Robust
Multiarray Averaging (RMA) method (13). The GSE21034 gene transcript data had
been pre-processed (by background adjustment and quantile
normalization using the RMA method) prior to being downloaded, and
were subsequently subjected to log2-transformation.
Differential expression analysis of
miRNA and genes
The differential expression analyses of miRNAs and
gene transcripts between patients with MPC and patients with PPC
were performed using the Linear Models for Microarray and RNA-Seq
Data 4 package of R (14). miRNAs or
genes with |log2 fold change (FC)| >1 and a false
discovery rate (FDR) value <0.05 were selected as differentially
expressed miRNAs (DEMs) or DEGs. The FDR value was obtained by
adjusting the raw P-values with the Benjamini and Hochberg method
(15).
Construction of a DEM-DEG regulatory
network
First, miRNA-gene pairs predicted by the miRanda
method were downloaded from the miRNA.org database
(16). Then, miRNA-gene interaction
pairs of upregulated DEM to downregulated DEG, and downregulated
DEM to upregulated DEG were screened to construct a DEM-DEG
regulatory network. Cytoscape (version 3.2.0; http://www.cytoscape.org/release_notes_3_2_0.html) was
used to visualize the resulting network (17).
Functional annotation of DEM-DEG
modules
In the constructed DEM-DEG network, a miRNA and its
target genes were defined as a module. For the DEGs in each module,
functional annotation analysis was performed using the Database for
Annotation, Visualization and Integrated Discovery online tool
based on the Gene Ontology database (P<0.01) (18).
Results
Differentially expressed miRNA and
genes between patients with MPC and PPC
Based on the analytical threshold, DEMs/DEGs between
MPC and PPC were screened out. The numbers of DEMs/DEGs between
patients with MPC and those with PPC are presented in Table II, in addition to the numbers of up-
and downregulated DEMs/DEGs. The number of downregulated DEMs/DEGs
was markedly greater compared with the number of upregulated
DEM/DEGs identified between patients with MPC and PPC. DEGs with
|log2 FC| >2 are presented in Table III.
| Table II.Overview of differentially expressed
microRNAs and genes. |
Table II.
Overview of differentially expressed
microRNAs and genes.
| Differential
expression | Upregulated | Downregulated | Total |
|---|
| DEM | 25 | 48 | 73 |
| DEG | 22 | 191 | 213 |
| Table III.Differentially expressed genes with
|log2 FC| >2 between metastatic and primary prostate
cancer samples. |
Table III.
Differentially expressed genes with
|log2 FC| >2 between metastatic and primary prostate
cancer samples.
| Gene
transcript | log2 FC
value | Adjusted
P-value | Gene
transcript | log2 FC
value | Adjusted
P-value |
|---|
| TAGLN | −2.4964 |
1.1987×10−31 | SORBS1 | −2.24347 |
1.4898×10−23 |
| SLC22A3 | −2.0225 |
5.9268×10−10 | SLC14A1 | −2.0417 |
4.5910×10−12 |
| SERPINA3 | −2.1640 |
1.0787×10−08 | PI15 | −2.5879 |
1.0048×10−13 |
| PGM5 | −2.1181 |
5.0371×10−23 | PDE5A | −2.1878 |
5.7525×10−19 |
| PCP4 | −2.3182 |
1.0747×10−24 | MYLK | −2.5515 |
1.9944×10−29 |
| MYH11 | −3.7948 |
1.5295×10−42 | MYBPC1 | −2.3552 |
1.8570×10−12 |
| MSMB | −3.1696 |
1.6542×10−15 | LTF | −2.5535 |
4.2645×10−07 |
| IGJ | −2.0615 |
2.2689×10−07 | PAM3B | −2.1011 |
5.5298×10−11 |
| DDP4 | −2.4582 |
1.0125×10−12 | CSRP1 | −2.0686 |
7.1619×10−22 |
| CNN1 | −2.7962 |
1.4001×10−35 | CHRDL1 | −2.2994 |
1.2660×10−22 |
| AZGP1 | −2.1882 |
2.8966×10−18 | ACTG2 | −3.2887 |
3.1041×10−33 |
DEM-DEG regulatory network
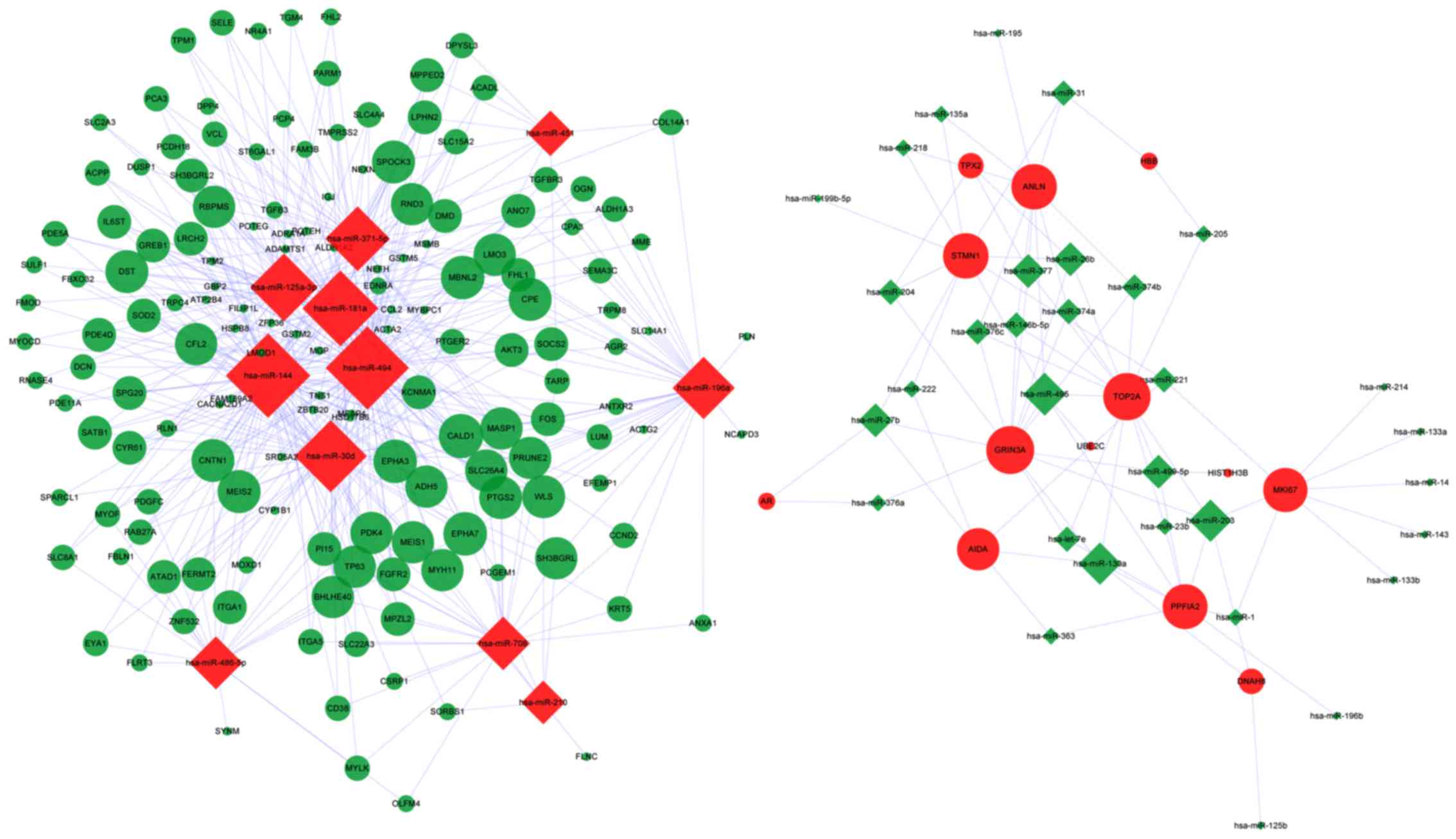
The resulting DEM-DEG regulatory network is
illustrated in Fig. 1. The average
connection degree of DEM was 12.4 (523/43), and that of DEG was 3.2
(523/166) (Table IV). Compared with
that of the downregulated miRNAs, upregulated miRNAs had a higher
degree of connection, that is, the latter regulate more
downregulated DEGs, including miR-144, miR-494 and miR-181a.
Certain DEGs were regulated by several miRNAs, including glutamate
ionotropic receptor NMDA type subunit 3A (GRIN3A), topoisomerase II
alpha (TOP2A) and caldesmon 1 (CALD1). GRIN3A and TOP2A were
upregulated by 14 and 13 downregulated miRNAs, respectively, while
CALD1 was downregulated by 8 upregulated DEMs.
| Table IV.Nodes and regulation pair statistics
of the DEM-DEG regulatory network. |
Table IV.
Nodes and regulation pair statistics
of the DEM-DEG regulatory network.
|
| Nodes |
|
|
|---|
|
|
|
|
|
|---|
| Regulation | DEM | DEG | Edges | Total |
|---|
| Up | 11 | 13 |
DEM.up-DEG.down | 442 |
| Down | 32 | 153 |
DEM.down-DEG.up | 81 |
Functional annotation of DEM-DEG
modules
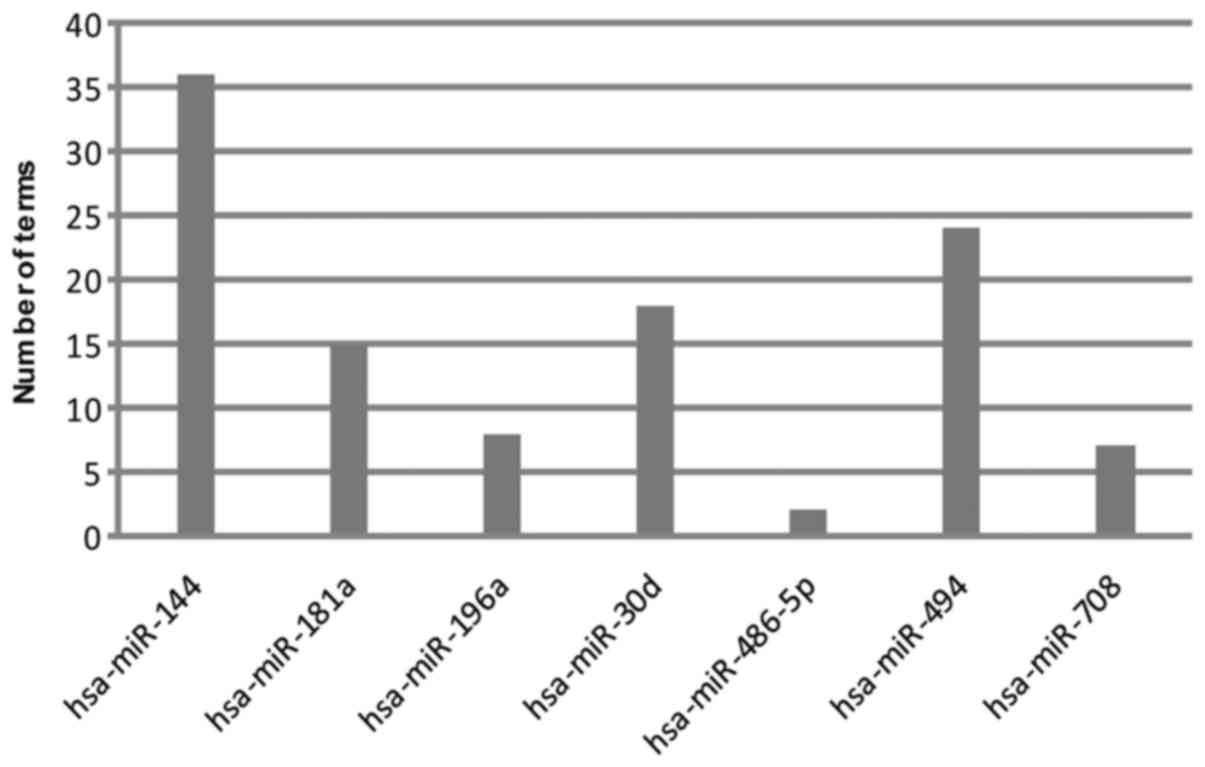
Seven modules were obtained (P<0.01), and their
enriched biological processes (BPs) are presented in Fig. 2. DEGs regulated by miR-144 enriched
the most BP terms (36), followed by
miR-494 (24), miR-30d (18), miR-181a (15), miR-196a (8), miR-708 (7)
and miR-486-5p (2). All these miRNAs
were upregulated.
Discussion
In the present study, DEMs and DEGs between patients
with PPC and MPC may function as biomarkers for the occurrence of
MPC. Furthermore, miRNAs and genes (particularly those with large
differential expression values) involved in the seven resulting
DEM-DEG regulatory modules may serve significant roles in the
metastasis of PC cells.
The target DEGs of miR-144 were enriched in the most
BP terms, implying that this integrated analysis of
multidimensional data miRNA and its targeted DEGs may perform
important roles in the occurrence of MPC. miR-144 has been reported
to improve the growth of HeLa cells (19), suggesting its role in tumor cell
proliferation. Zhang et al (20) have proposed that miR-144 promotes the
malignant progression of nasopharyngeal carcinoma cells by
targeting the tumor-suppressor gene phosphatase and tensin homolog;
however, its role in PC has not been reported thus far, to the best
of our knowledge. In the present study, numerous DEGs were observed
to be downregulated by miR-144. Among them, myosin heavy chain 11
(MYH11), solute carrier family 22 member 3 (SLC22A3)
and dipeptidyl peptidase 4 (DPP4) exhibited large
|log2 FC| values. MYH11 (also known as
SMMHC) encodes the smooth muscle myosin heavy chain, which
serves a key role in smooth muscle contraction (21). Its downregulation has been reported by
Lin et al (22) in
proliferating smooth muscle cells of human prostate tissue.
SLC22A3 is one of three similar cation transporter genes
located in a cluster on chromosome 6 and has previously been
suggested to be associated with PC pathogenesis (23). DPP4 encodes a serine
exopeptidase that has been implicated in cell-extracellular matrix
interactions and bioactive peptide/cytokine/growth factor
metabolism (24,25). DPP4 activity is elevated in PC
and adjacent benign hyperplastic glands (26). Taken together, miR-144 may serve roles
in the metastasis of PC cells by downregulating a number of target
genes, including MYH11, SLC22A3 and DPP4.
The regulator miR-494 may also serve an important
role in the metastasis of PC. MYH11 was the only target gene
with |log2 FC| >2 among the downregulated DEGs of
miR-494 in the present study. In a previous study, using multiple
experimental methods, Shen et al (27) demonstrated that miRNA-494-3p could
suppress the proliferation, invasion and migration of PC by
downregulating C-X-C motif chemokine receptor 4 (CXCR4),
which was overexpressed in PC. This is consistent with the
upregulation of this miRNA observed in the present study, although
differential expression of CXCR4 was not observed.
miR-30d downregulated a number of genes in the
present study. Kobayashi et al (28) observed significantly higher expression
levels of miR-30d in three PC cell lines compared with those in two
normal prostate cell lines using miRNA microarray and quantitative
polymerase chain reaction analysis (28). Furthermore, the authors suggested that
miR-30d mediated its effects in PC by downregulating suppressor of
cytokine signaling 1. In the present study, among the target DEGs
of miR-30d, sorbin and SH3 domain containing 1 (SORBS1),
phosphodiesterase-5 (PDE5) and myosin light chain kinase
(MYLK) exhibited large |log2 FC| values.
SORBS1 encodes a Cbl-associated protein involved in the
formation of actin stress fibers and focal adhesions (29). The observed downregulation of
SORBS1 in the present study was consistent with that
reported by Vanaja et al (30)
in PC tissues. PDE5 encodes an enzyme that hydrolyzes the
3′,5′-phosphodiester bond in the second messenger molecule cyclic
guanosine monophosphate (GMP) to form 5′-GMP (31). Its expression has been observed in the
smooth muscles of the prostate, and PDE5 inhibitors (such as
sildenafil) are able to relax the prostate (32). Therefore, the upregulation of
PDE5 is suggested to be disadvantageous for patients with
PC. MYLK has been demonstrated to be necessary for the
invasiveness of MPC cells (33).
However, PDE5 and MYLK were observed to be
downregulated in the present study; therefore, further studies are
required to elucidate the roles of PDE5 and MYLK in
PC metastasis.
In addition to their observed downregulation by
miR-144 MYH11 and SLC22A3 were downregulated by
miR-181a, which was upregulated in the present study. miR-181a has
been reported in previous studies to induce apoptosis in a number
of cancer types by downregulating the apoptosis regulator B-cell
lymphoma 2 (11,34), and it has been observed to mediate
bufalin-induced apoptosis in PC-3 PC cells by Zhai et al
(35). However, the implications of
upregulation of miR-181a in MPC samples remains unclear.
Additionally, Su et al (36)
reported that the downregulation of miR-30d and miR-181a in
prostate tumors cooperatively suppresses the expression of
glucose-regulated protein, 78 kDa (GRP78), a major
endoplasmic reticulum chaperone and signaling regulator that is
typically overexpressed in cancer (37). Differential expression of GRP78
was not observed in the present study; however, MYH11 was
revealed to be another common target of miR-30d and miR-181a,
indicating that miR-30d and miR-181a may cooperate to regulate the
metastasis of PC cells.
A previous study suggested that miR-196a regulates
homeobox (Hox) gene expression during vertebrate
embryogenesis (38). In addition, a
correlation between aberrant HoxC8 expression and a malignant
phenotype in human PC has been reported (39,40).
Therefore, it can be inferred that miR-196a may serve a role in the
occurrence of MPC. In the present study, one of the target genes of
miR-196a, actin, gamma 2 (ACTG2), exhibited |log2
FC| >2, in agreement with the significant downregulation in
metastatic and primary tumor samples observed by Chandran et
al (41). The findings from the
present study further suggest that miR-196a may serve a role in the
metastasis of PC cells by downregulating ACTG2.
Regarding miR-708 and miR-486-5p, no DEGs with a
|log2 FC| >2 were observed among their target genes
in the present study, implying that these two miRNAs do not perform
crucial roles in the occurrence of MPC. According to Watahiki et
al (10), miR-708 exhibited a
>5-fold decrease in an MPC cell line following comparative
analysis of miRNAs libraries between MPC and PPC cell lines. This
was not consistent with the upregulation of miR-708 observed in the
present study. Therefore, the expression changes and role of
miR-708 in MPC require further investigation. Additionally,
Watahiki et al (10) also
reported that elevated miR-486 level enhanced the invasiveness of
MPC cells, which is consistent with its upregulation in the present
study.
In conclusion, the significantly upregulated
miR-144, miR-494, miR-30d, miR-181a, miR-196a, miR-708 and
miR-486-5p screened in the present study may participate in the
metastasis of PC cells via the downregulation of their
corresponding target DEGs, particularly those with large
|log2 FC| values, including MYH11, SLC22A3, DPP4,
SORBS1, PDE5, MYLK and ACTG2. The effects
on these target DEGs require further experimental verification. A
number of these DEMs or DEGs have been associated with the
occurrence of PC; however, the molecular mechanisms underlying
their roles in the occurrence of MPC remain unclear and require
further investigation.
Acknowledgements
The present study was supported by a grant from the
Jilin Provincial Health Department (grant no. 3D5157343428).
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bonci D, Coppola V, Patrizii M, Addario A,
Cannistraci A, Francescangeli F, Pecci R, Muto G, Collura D, Bedini
R, et al: A microRNA code for prostate cancer metastasis. Oncogene.
35:1180–1192. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guinney J, Wang T, Laajala TD, Winner KK,
Bare JC, Neto EC, Khan SA, Peddinti G, Airola A, Pahikkala T, et
al: Prediction of overall survival for patients with metastatic
castration-resistant prostate cancer: Development of a prognostic
model through a crowdsourced challenge with open clinical trial
data. Lancet Oncol. 18:132–142. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Valencia-Sanchez MA, Liu J, Hannon GJ and
Parker R: Control of translation and mRNA degradation by miRNAs and
siRNAs. Genes Dev. 20:515–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fabris L, Ceder Y, Chinnaiyan AM, Jenster
GW, Sorensen KD, Tomlins S, Visakorpi T and Calin GA: The potential
of MicroRNAs as prostate cancer biomarkers. Eur Urol. 70:312–322.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Endzeliņš E, Melne V, Kalniņa Z,
Lietuvietis V, Riekstiņa U, Llorente A and Linē A: Diagnostic,
prognostic and predictive value of cell-free miRNAs in prostate
cancer: A systematic review. Mol Cancer. 15:412016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baranwal S and Alahari SK: miRNA control
of tumor cell invasion and metastasis. Int J Cancer. 126:1283–1290.
2010.PubMed/NCBI
|
|
9
|
Ambs S, Prueitt RL, Yi M, Hudson RS, Howe
TM, Petrocca F, Wallace TA, Liu CG, Volinia S, Calin GA, et al:
Genomic profiling of microRNA and messenger RNA reveals deregulated
microRNA expression in prostate cancer. Cancer Res. 68:6162–6170.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Watahiki A and Wang Y, Morris J, Dennis K,
O'Dwyer HM, Gleave M, Gout PW and Wang Y: MicroRNAs associated with
metastatic prostate cancer. PLoS One. 6:e249502011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Taylor BS, Schultz N, Hieronymus H,
Gopalan A, Xiao Y, Carver BS, Arora VK, Kaushik P, Cerami E, Reva
B, et al: Integrative genomic profiling of human prostate cancer.
Cancer Cell. 18:11–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
López-Romero P: Pre-processing and
differential expression analysis of Agilent microRNA arrays using
the AgiMicroRna Bioconductor library. BMC Genomics. 12:642011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Smyth GK: Limma: Linear models for
microarray dataBioinformatics and Computational Biology Solutions
Using R and Bioconductor. Springer; pp. 397–420. 2005, View Article : Google Scholar
|
|
15
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc Ser B (Methodological). 57:289–300.
1995.
|
|
16
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human microRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
da Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng AM, Byrom MW, Shelton J and Ford LP:
Antisense inhibition of human miRNAs and indications for an
involvement of miRNA in cell growth and apoptosis. Nucleic Acids
Res. 33:1290–1297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang LY, Lee Ho-Fun V, Wong AM, Kwong DL,
Zhu YH, Dong SS, Kong KL, Chen J, Tsao SW, Guan XY and Fu L:
MicroRNA-144 promotes cell proliferation, migration and invasion in
nasopharyngeal carcinoma through repression of PTEN.
Carcinogenesis. 34:454–463. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lopezcamacho C, van Wijnen AJ, Lian JB,
Stein JL and Stein GS: (CBFβ) and the leukemogenic fusion protein
CBFβ-SMMHC associate with mitotic chromosomes to epigenetically
regulate ribosomal gene. J Cell Biochem. 115:2155–2164. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin VK, Wang D, Lee I, Vasquez D, Fagelson
JE and McConnell JD: Myosin heavy chain gene expression in normal
and hyperplastic human prostate tissue. Prostate. 44:193–203. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grisanzio C, Werner L, Takeda D, Awoyemi
BC, Pomerantz MM, Yamada H, Sooriakumaran P, Robinson BD, Leung R,
Schinzel AC, et al: Genetic and functional analyses implicate the
NUDT11, HNF1B, and SLC22A3 genes in prostate cancer pathogenesis.
Proc Natl Acad Sci USA. 109:11252–11257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zilleßen P, Celner J, Kretschmann A,
Pfeifer A, Racké K and Mayer P: Metabolic role of dipeptidyl
peptidase 4 (DPP4) in primary human (pre)adipocytes. Sci Rep.
6:230742016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhong J, Kankanala S and Rajagopalan S:
Dipeptidyl peptidase-4 inhibition: Insights from the bench and
recent clinical studies. Curr Opin Lipidol. 27:484–492. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wilson MJ, Ruhland AR, Quast BJ, Reddy PK,
Ewing SL and Sinha AA: Dipeptidylpeptidase IV activities are
elevated in prostate cancers and adjacent benign hyperplastic
glands. J Androl. 21:220–226. 2000.PubMed/NCBI
|
|
27
|
Shen PF, Chen XQ, Liao YC, Chen N, Zhou Q,
Wei Q, Li X, Wang J and Zeng H: MicroRNA-494-3p targets CXCR4 to
suppress the proliferation, invasion, and migration of prostate
cancer. Prostate. 74:756–767. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kobayashi N, Uemura H, Nagahama K, Okudela
K, Furuya M, Ino Y, Ito Y, Hirano H, Inayama Y, Aoki I, et al:
Identification of miR-30d as a novel prognostic maker of prostate
cancer. Oncotarget. 3:1455–1471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ribon V, Herrera R, Kay BK and Saltiel AR:
A role for CAP, a novel, multifunctional Src homology 3
domain-containing protein in formation of actin stress fibers and
focal adhesions. J Biol Chem. 273:4073–4080. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vanaja DK, Ballman KV, Morlan BW, Cheville
JC, Neumann RM, Lieber MM, Tindall DJ and Young CY: PDLIM4
repression by hypermethylation as a potential biomarker for
prostate cancer. Clin Cancer Res. 12:1128–1136. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stoclet JC, Keravis T, Komas N and Lugnier
C: Section Review: Cardiovascular & Renal: Cyclic nucleotide
phosphodiesterases as therapeutic targets in cardiovascular
diseases. Expert Opinion Investigational Drugs. 4:19952008.
|
|
32
|
Uckert S, Küthe A, Jonas U and Stief CG:
Characterization and functional relevance of cyclic nucleotide
phosphodiesterase isoenzymes of the human prostate. J Urol.
166:2484–2490. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tohtong R, Phattarasakul K, Jiraviriyakul
A and Sutthiphongchai T: Dependence of metastatic cancer cell
invasion on MLCK-catalyzed phosphorylation of myosin regulatory
light chain. Prostate Cancer Prostatic Dis. 6:212–216. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu DX, Zhu W, Fang C, Fan L, Zou ZJ, Wang
YH, Liu P, Hong M, Miao KR, Liu P, et al: miR-181a/b significantly
enhances drug sensitivity in chronic lymphocytic leukemia cells via
targeting multiple anti-apoptosis genes. Carcinogenesis.
33:1294–1301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhai XF, Fang FF, Liu Q, Meng YB, Guo YY
and Chen Z: MiR-181a contributes to bufalin-induced apoptosis in
PC-3 prostate cancer cells. BMC Complement Altern Med. 13:3252013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Su SF, Chang YW, Andreu-Vieyra C, Fang JY,
Yang Z, Han B, Lee AS and Liang G: miR-30d, miR-181a and
miR-199a-5p cooperatively suppress the endoplasmic reticulum
chaperone and signaling regulator GRP78 in cancer. Oncogene.
32:4694–4701. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen WT, Zhu G, Pfaffenbach K, Kanel G,
Stiles B and Lee AS: GRP78 as a regulator of liver steatosis and
cancer progression mediated by loss of the tumor suppressor PTEN.
Oncogene. 33:4997–5005. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yekta S, Shih IH and Bartel DP:
MicroRNA-directed cleavage of HOXB8 mRNA. Science. 304:594–596.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Waltregny D, Alami Y, Clausse N, de Leval
J and Castronovo V: Overexpression of the homeobox gene HOXC8 in
human prostate cancer correlates with loss of tumor
differentiation. Prostate. 50:162–169. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miller GJ, Miller HL, van Bokhoven A,
Lambert JR, Werahera PN, Schirripa O, Lucia MS and Nordeen SK:
Aberrant HOXC expression accompanies the malignant phenotype in
human prostate. Cancer Res. 63:5879–5888. 2003.PubMed/NCBI
|
|
41
|
Chandran UR, Ma C, Dhir R, Bisceglia M,
Lyons-Weiler M, Liang W, Michalopoulos G, Becich M and Monzon FA:
Gene expression profiles of prostate cancer reveal involvement of
multiple molecular pathways in the metastatic process. BMC Cancer.
7:642007. View Article : Google Scholar : PubMed/NCBI
|