Introduction
The survivin gene is located at the 17q15 region
chromosomal region and contains four exons and three introns
(1). Survivin was firstly separated
from a human genomic library using the complementary DNA (cDNA) of
effector cell protease receptor-1 (EPR-1) through hybridization
screening (2). Survivin, containing
only one baculovirus inhibitor of apoptosis protein repeat (BIR)
domain, is the smallest member of the mammalian inhibitors of the
apoptosis protein (IAP) family (2,3). Survivin
is detectable during fetal development and is highly expressed in
various malignant tumors, but is undetectable in adult terminal
differentiation tissues (2,4–6). Survivin
serves multiple functions in cell division, apoptosis, metastasis
and angiogenesis (7). It has been
reported that survivin acts as an intermediary between cell
apoptosis and cell cycle checkpoints, serving an important role in
reducing cellular apoptosis and regulating mitosis (8). Survivin suppresses apoptosis through
inhibition of the terminal effectors caspase-3 and caspase-7,
promoting the differentiation of tumor cells (6,9). Survivin
expression peaks at the G2/M phase of the cell cycle,
where it associates with mitotic spindle microtubules; its
overexpression is required to protect cells from apoptosis during
mitosis (10). It has previously been
reported that the overall survival rate of patients that express
high levels of survivin is decreased, and that survivin is an
indicator of poor prognosis (11,12); the
recurrence rate of these patients was increased and patients were
not sensitive to radiotherapy and chemotherapy (13). Therefore, survivin may be a promising
target for anticancer therapy (14).
RNA interference (RNAi) is the sequence-specific
silencing of genes induced by endogenous or exogenous 21–23 nt
double-stranded RNA in cells (15).
RNAi is a feasible and effective method of inhibiting the
endogenous expression of target genes, and has been widely employed
in the functional studies of genes in vivo and in
vitro (16,17). Two modified strategies of RNAi
technology have been used to block gene expression in mammalian
cells, including small interfering RNA (siRNA) and vector-mediated
short hairpin RNA (shRNA) (18).
Compared with shRNA vectors, siRNAs are more readily synthesized
and delivered into cells; however, siRNA-mediated gene silencing is
transient and siRNAs are susceptible to RNase degradation (19–21).
shRNAs, on the other hand, can be used for long-term gene silencing
and can also be propagated indefinitely (22,23). Once
delivered into cells, shRNAs are cleaved into active siRNAs by
Dicer, which then induce the homology-dependent degradation of
cognate mRNA (24).
In the present study, the vector-derived shRNA
technique was utilized and the recombinant plasmid
pGP-U6-GFP-Survivin-shRNA was constructed, using a range of
different shRNAs with a U6 small nuclear RNA promoter. The shRNAs
used were targeted against different sites of the survivin gene,
including exonic and intronic sequences, and were transfected into
HeLa cells. The influence of survivin expression and cell apoptosis
were both investigated. The present study may aid the development
of a theoretical foundation for gene therapy in cancer.
Materials and methods
Cells line and culture
The human cervical carcinoma HeLa cell line was
provided by the Institute of Biochemistry and Molecular Biology of
Guangdong Medical University (Zhanjiang, China). The HeLa cells
were cultured in Dulbecco's Modified Eagles Medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Zhejiang Tianhang Biotechnology Co.,
Ltd., Zhejiang, China), 100 U/ml penicillin, and 0.1 g/ml
streptomycin (Beyotime Institute of Biotechnology, Shanghai, China)
at 37°C in a humidified atmosphere containing 5%
CO2.
Recombinant plasmid-based
survivin-targeting shRNA design
A total of 6 shRNAs targeting the survivin gene (and
1 negative control) were designed to be homologous to the survivin
mRNA sequence (GeneBank no. NM001168). shRNA1 was targeted at
intron 1; shRNA2 was targeted at intron 2; shRNA3 and shRNA4 were
targeted at intron 3; shRNA5 was targeted at exon 1 and shRNA6 was
targeted at exon 4 (Table I).
According to BLAST analysis, the nucleotide sequences of
survivin-targeted did not exhibit any non-specific interactions
with other mRNA transcripts. All nucleotide sequences were
synthesized and inserted into the recombinant plasmid vectors
pGP-U6-GFP-Survivin by Shanghai GenePharma Co., Ltd. (Shanghai,
China). The company presented a negative control shRNA (shNC),
which did not possess any complementary region with survivin
mRNA.
 | Table I.Sequences of survivin-shRNAs and
negative control. |
Table I.
Sequences of survivin-shRNAs and
negative control.
| shRNA | Sequence
length | Number of
nucleotides |
|---|
| shRNA1 |
GGTGATGCTTACAACCTAA | 19 |
| shRNA2 |
GGGAGAGAGAAGGTGCTAA | 19 |
| shRNA3 |
GCTCATGCTTTCCTTGCTA | 19 |
| shRNA4 |
GCATTGGGCGCTGATTCTT | 19 |
| shRNA5 |
CCGCATCTCTACATTCAAGAA | 21 |
| shRNA6 |
GCACCACTTCCAGGGTTTATT | 21 |
| shNC |
GTTCTCCGAACGTGTCACGT | 20 |
Transfection of the shRNAs into HeLa
cells
Cells were seeded into 6-well plates in DMEM without
antibiotics prior to transfection. When cells were at 60–70%
confluence in monolayer, transfection of shRNA was performed using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol, with a
non-transfected group acting as the blank control. The shRNA:lipid
reagent was used at a ratio of 1:2; each shRNA was transfected into
three marked wells, cultured in humidified incubator and the medium
was changed with fresh medium each day.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
After a 48-h transfection, the total RNA of all
groups was extracted using TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocol.
cDNA was synthesized from total RNA (1 µg) using the FastQuant RT
kit (With gDNase) (Tiangen Biotech Co., Ltd., Beijing, China). qPCR
was performed using the SYBR®Premix Ex TaqII kit (Takara
Bio, Inc., Otsu, Japan) on LightCycler®480 (Roche,
Switzerland). The human GAPDH gene served as an internal control.
The primers used were: Survivin forward,
5′-TGACGACCCCATAGAGGAACA-3′ and reverse,
5′-CGCACTTTCTCCGCAGTTTC-3′; and GAPDH forward,
5′-GGGTGTGAACCATGAGAAGT-3′ and reverse, 5′-CAGTGATGGCATGGACTGTG-3′.
The final qPCR volume was 20 µl, with 2 µl cDNA template and 0.4 µM
of each primer. The qPCR parameters were as follows: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles of
denaturation at 95°C for 5 sec and annealing and extension at 60°C
for 20 sec. Results were analyzed in relation to GAPDH levels using
the 2−ΔΔCq method (25).
Each experiment was performed in triplicate.
Western blot analysis
The total protein was obtained from each group using
cold radio immune precipitation lysis buffer (cat. no. P0013B,
Beyotime Institute of Biotechnology) containing
phenylmethanesulfonyl fluoride (PMSF, catalog no. ST506-2, Beyotime
Institute of Biotechnology) and protease inhibitors after a 72-h
transfection. Protein concentration was quantified using the
bicinchoninic acid (BCA) method using the Enhanced BCA Protein
Assay kit (Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. The proteins (60 µg/lane) were
concentrated by 5% SDS-PAGE, separated by 12% SDS-PAGE and then
transferred to a polyvinylidene fluoride membrane (cat. no.
ISEQ00010, EMD Millipore, Billerica, MA, USA). The membrane was
blocked using 5% skimmed milk in Tris-HCl Buffered Saline Tween-20
(TBST, 25 mM Tris-HCl, 125 mM NaCl, 0.1% Tween-20) at room
temperature for 2 h with gentle agitation, and incubated overnight
at 4°C with monoclonal anti-survivin (1:1,000; cat. no. 2808T; Cell
Signaling Technology, Inc., Danvers, MA, USA) and anti-GAPDH
(1:1,000; cat. no. 2118S, Cell Signaling Technology, Inc.)
antibodies. Following a wash with TBST, the membrane was incubated
with mice anti-rabbit IgG-horseradish-peroxidase monoclonal
antibodies (1:2,000; cat. no. 5127S, Cell Signaling Technology,
Inc.) for 1 h at room temperature. The antibodies were diluted with
5% skimmed milk in TBST buffer. The protein bands were then
visualized using ECL Chemiluminescence Detection kit (Beyotime
Institute of Biotechnology), using GAPDH protein as reference. The
density of the brands on the membrane was scanned with Canon
Solution Menu EX (Canon, Zhanjiang, China) and analyzed with ImageJ
1.46 software (National Institutes of Health, Bethesda, MD,
USA).
FCM analysis
A total of 4×105 HeLa cells were seeded
in 6-well plants. All groups of cells were treated with EDTA-free
trypsin (Beyotime Institute of Biotechnology) for 3 min after a
48-h transient transfection. Cells were then centrifuged at 425 × g
at 4°C for 5 min and the supernatant was discarded. Cells were then
washed with cold PBS and centrifuged at 425 × g at 4°C for 5 min,
removed and supernatant discarded carefully, twice. Cells were
re-suspended in 100 µl of 1X Binding Buffer, and 5 µl Annexin
V-fluorescein isothiocyanate (FITC) and 5 µl propidium iodide (PI)
staining solution (cat. no. A221-01/02, all Vazyme Biotech Co.,
Ltd., Nanjing, China) were added, the solution was mixed gently and
incubated at room temperature for 10 min in darkness. Next, 400 µl
of 1X Binding Buffer was added. The ratios of apoptotic cells were
assessed using a Coulter EPICS XL Flow Cytometer using Expo32-ADC
v. 1.2B software (Beckman Coulter, Inc., Brea, CA, USA).
Hoechst 33258 stain analysis
In order to access nuclear condensation by Hoechst
33258 staining, 2×105 HeLa cells per well were washed
twice with 2 ml PBS 48 h after transfection. A total of 1 ml
Hoechst 33258 reagents (Beyotime Institute of Biotechnology) were
added to each well and cells were incubated at 37°C for 30 min in
the dark. Hoechst 33258 reagents were then removed and the cells
were washed with PBS three times, 5 min each time. Morphological
changes of apoptotic cells were observed under an inverted
fluorescence microscope and images were captured.
Statistical analysis
SPSS 17.0 (SPSS, Inc., Chicago, IL, USA) software
was used for statistical analysis. The results were presented as
the mean ± standard deviation (SD) of three independent
experiments. One-way analysis of variance was used to perform
statistical comparisons of the data. P<0.05 was considered to
indicate a statistically significant difference.
Results
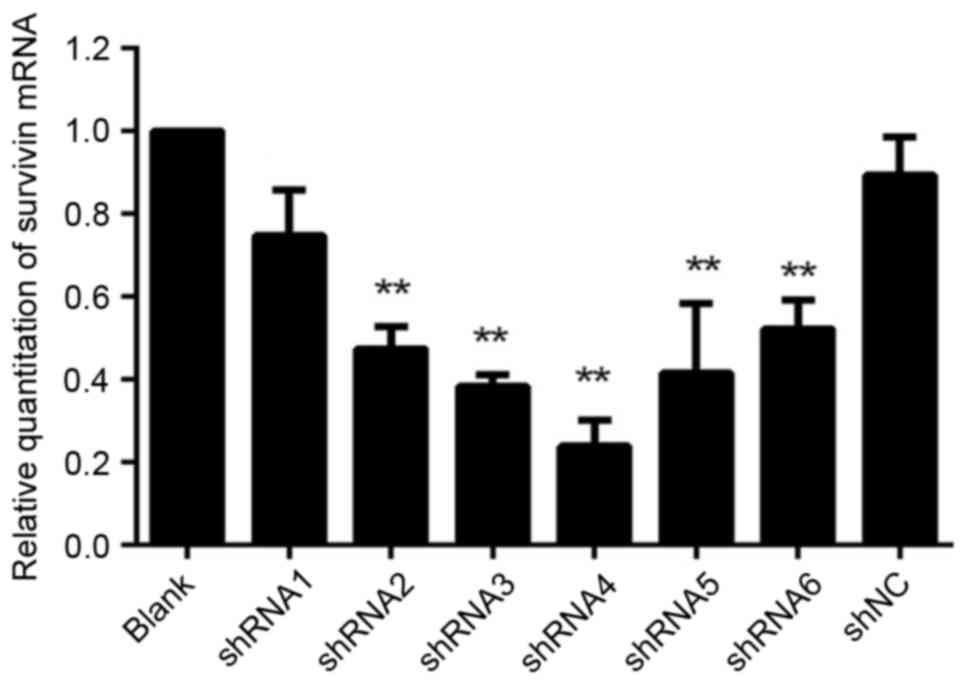
Expression of survivin mRNA following
transfection of HeLa cells with shRNAs
Survivin mRNA expression was evaluated in HeLa cells
by RT-qPCR following transfection with recombinant plasmid
vector-mediated survivin shRNAs (Fig.
1). For all groups treated with survivin-shRNA, survivin mRNA
expression was significantly reduced compared with the blank
control and shNC (P<0.05). No significant differences were
observed in survivin mRNA expression between the shNC and the blank
control groups (P=0.09). However, the effect of the suppression for
the group transfected with shRNA1 was not well enough compared with
other groups, and therefore it was not used in the following
experiments.
Expression of survivin protein
following shRNAs transfection in HeLa cells
Survivin protein expression was examined by western
blot analysis (Fig. 2). Survivin
protein levels in all shRNA-treated groups were significantly
reduced (P<0.05; Fig. 2B);
however, no significant differences were observed between the
shNC-treated and blank-control groups (P=0.40). The expression of
survivin protein can not only be reduced by RNA1 extron-specific
shRNAs but also inhibited by reduced intron-specific shRNAs in HeLa
cells.
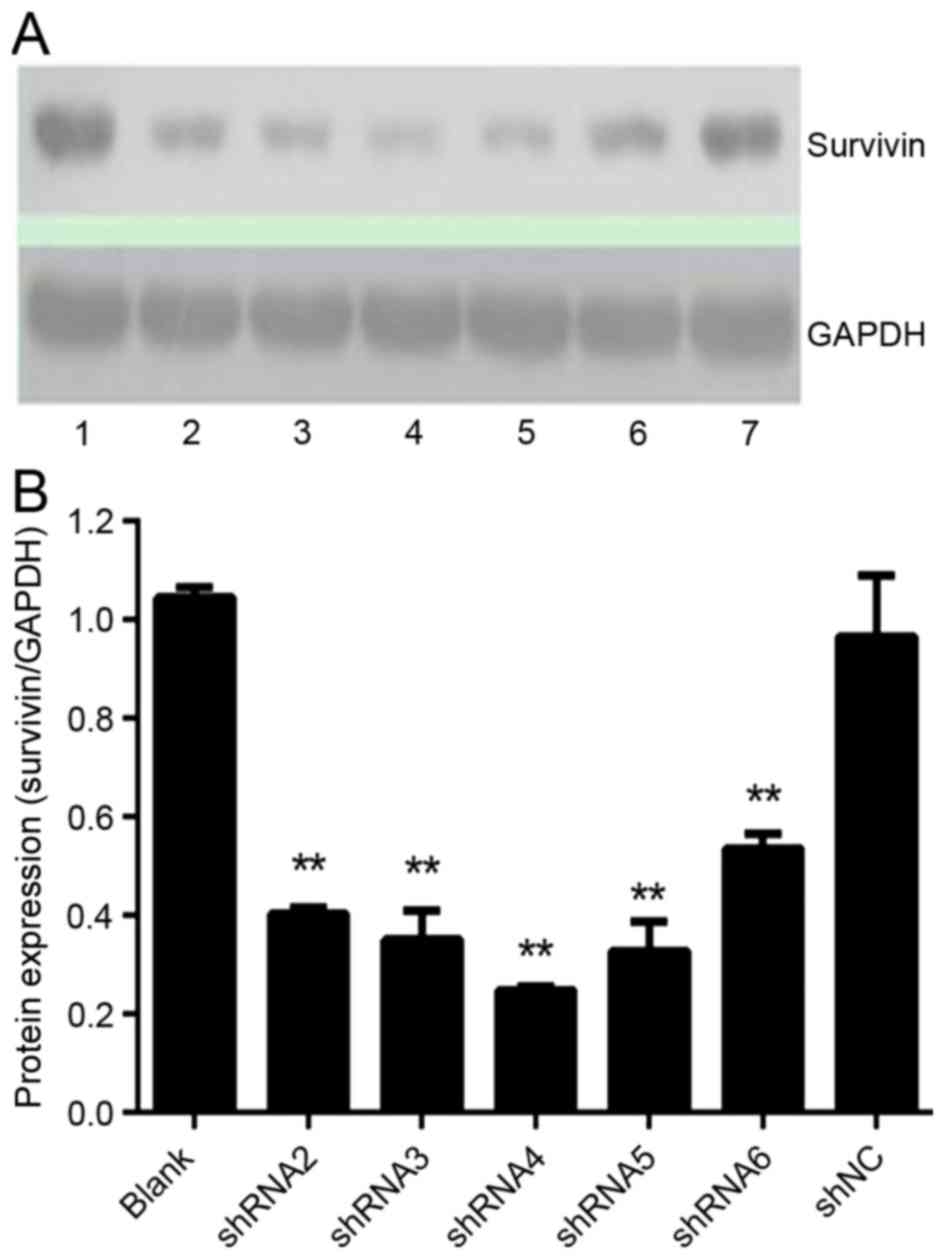 | Figure 2.Effect of shRNAs on survivin protein
expression. Protein levels were analyzed by western blot analysis
after a 72-h transfection. (A) Bands of survivin and GAPDH were
scanned. Lane 1, blank control; lane 2, shRNA2; lane 3, shRNA3;
lane 4, shRNA4; lane 5, shRNA5; lane 6, shRNA6; lane 7, shNC. (B)
The quantitative representation of survivin protein levels were
determined by the density of the bands and one representation of
three independent experiments was demonstrated, **P<0.01 vs.
blank control. shRNA, short hairpin RNA; shNC, negative control
shRNA. |
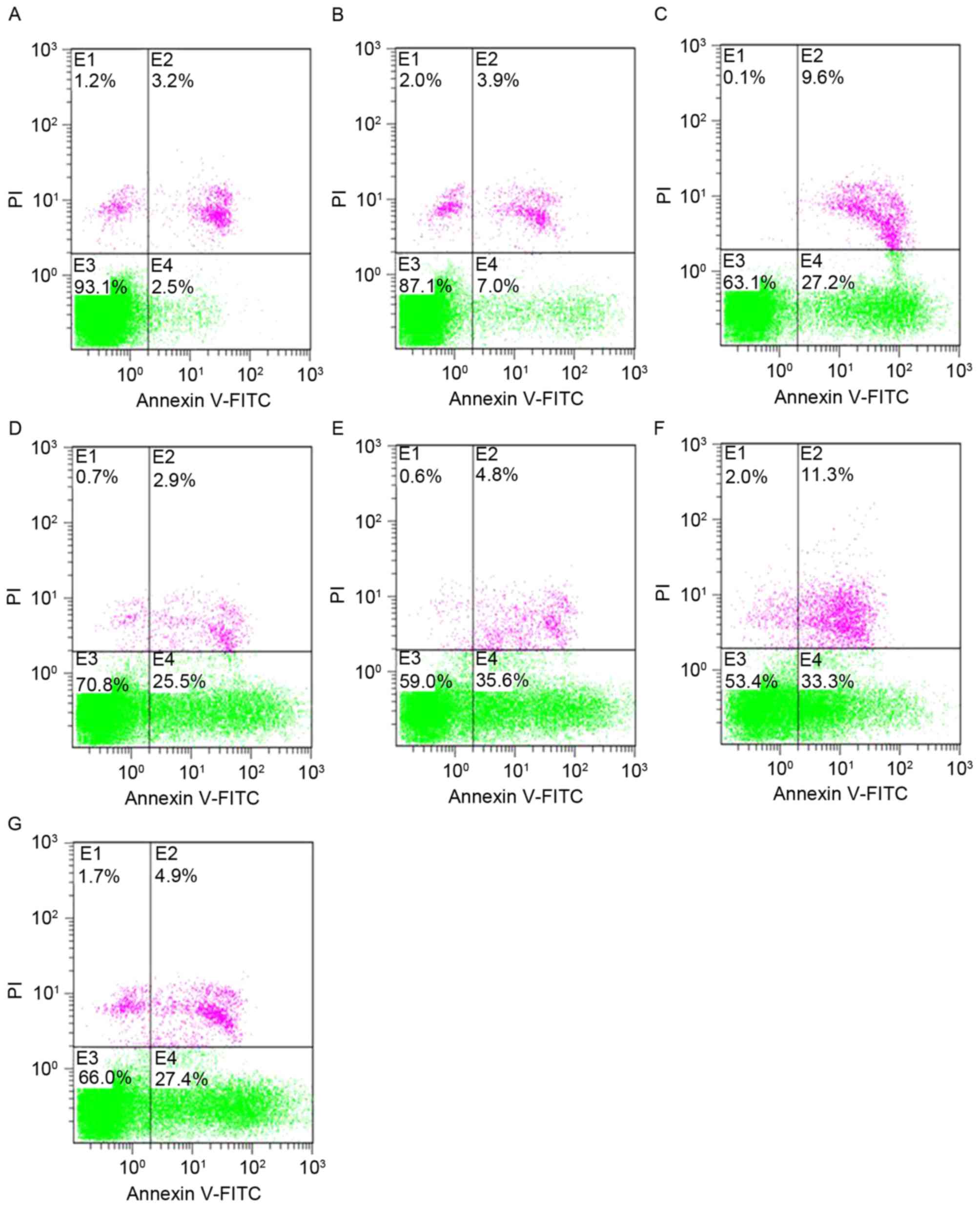
Effect of shRNA treatment on apoptotic
rates, assessed by flow cytometry (FCM)
Cells transfected with shRNAs exhibited a
significant increase in the level of apoptosis, as detected by FCM
analysis via Annexin V-FITC/PI staining (Fig. 3; Table
II). All data were presented as the mean ± SD of three
independent experiments. The apoptosis rates were significantly
increased in the shRNA-treated groups compared with shNC-treated
and blank-control groups (P<0.05). No significant differences
were observed between the shNC-treated and blank-control
groups.
| Table II.Cell apoptosis rates after a 48-h
transfection. |
Table II.
Cell apoptosis rates after a 48-h
transfection.
| Groups | Apoptosis rate,
% |
|---|
| Blank control |
3.5±0.87 |
| shNC |
5.67±1.15 |
| shRNA-2 |
27.63±1.59a |
| shRNA-3 |
25.77±0.83a |
| shRNA-4 |
37.87±2.25a |
| shRNA-5 |
35.27±1.76a |
| shRNA-6 |
28.5±2.91a |
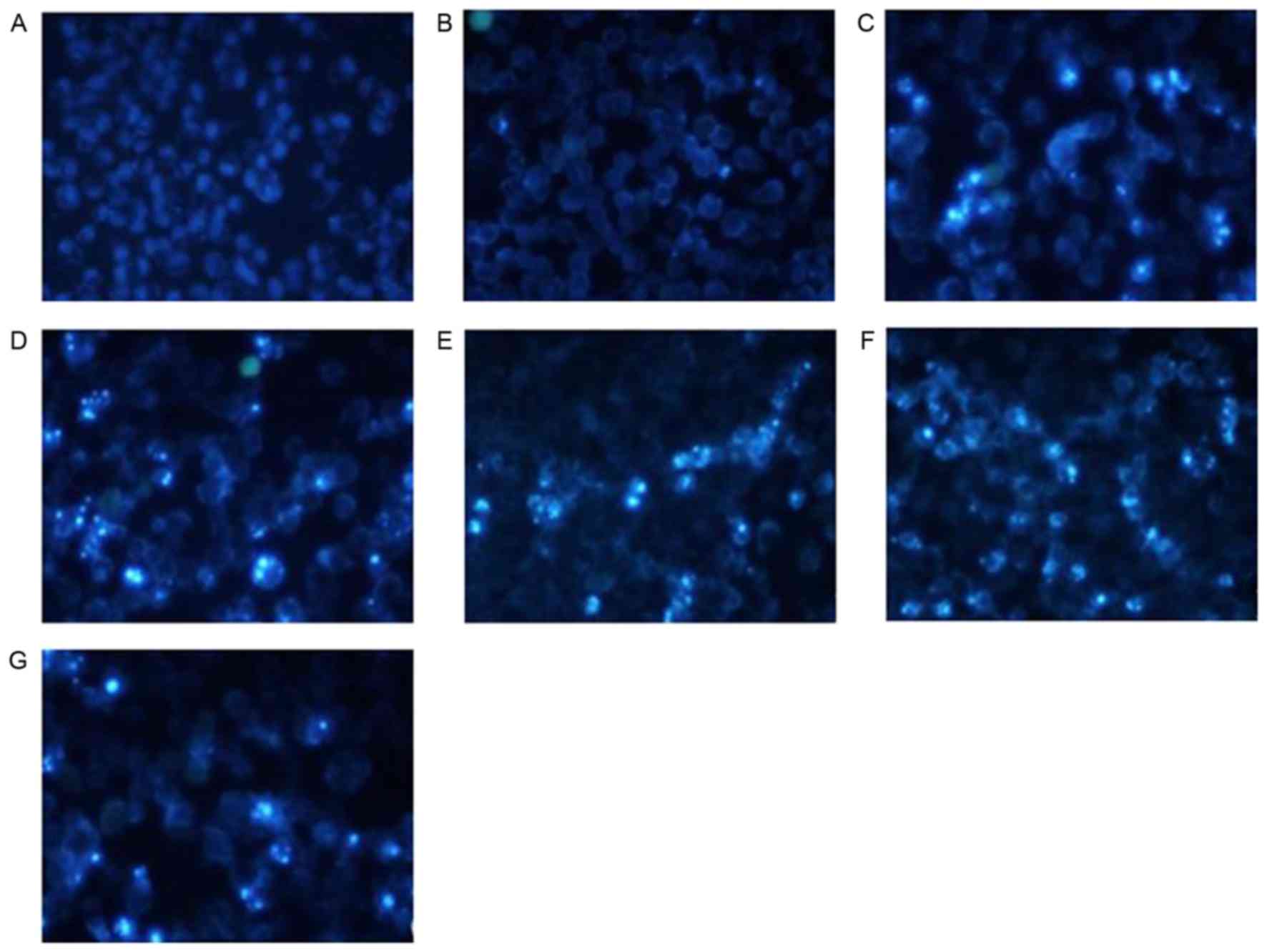
Effect of shRNA treatment on
morphological changes of apoptotic cells by Hochest 33258 stain
analysis
HeLa cells treated with shRNAs exhibited the
following typical apoptotic changes following transfection
(Fig. 4): Cell shrinkage, nuclear
condensation and dense staining with some white color were observed
under an inverted fluorescence microscope. Fluorescence staining in
the group treated with shNC and the blank control group was light
and uniform, and the number of apoptotic cells was markedly
decreased.
Discussion
The capability of cells to escape apoptosis is one
of the defined hallmarks of cancer (26,27).
Multiple previous reports indicated that survivin serves a critical
function during tumorigenesis by inhibiting apoptosis, accelerating
the progress of mitosis and promoting the growth of tumor cells
(28,29). High survivin expression is observed in
the majority malignant tissues yet is absent in mature, healthy
tissues (3), indicating it may
represent a promising therapeutic target. Various strategies have
been taken to inhibit the expression of survivin in cancer cells,
including the use of antisense oligonucleotides (30), dominant-negative mutants (31), ribozymes (32,33) and
anticancer vaccines (34). RNAi may
be a powerful tool for cancer therapy (35). RNAi is efficient at lower
concentrations for anticancer treatment and results in fewer side
effects compared with other techniques (35,36).
RNAi-mediated inhibition of survivin expression has been used to
delay mitosis by causing chromosome misalignment and prometaphase
accumulation in HeLa cells (19,37), and
has successfully reduced survivin expression (26,38–40).
The vast majority of human genes are made up of
introns. They have been considered to be ‘junk DNA’ because they
are degraded following splicing (41–43).
However, a previous study demonstrated that introns serve a
critical role in transcriptional regulation (44). Introns are not only involved in the
formation of microRNAs and small nucleolar RNAs, but also regulate
alternative splicing and affect the mRNA level (45). In the present study, the shRNAs
targeting exons and introns effectively inhibited the expression of
survivin in HeLa cells and increased the apoptosis rate, inducing
marked apoptotic morphological changes. This indicates that introns
were not ‘junk DNA’.
The survivin gene contains four exons and three
introns, and the survivin protein is the smallest member of the
mammalian IAP family (2). To the best
of our knowledge, few reports concerning the use of shRNA/siRNA
targeted against introns to silence oncogenes exist, as the
siRNA/shRNAs generated tend to target exons or promoters. As such,
shRNAs in the present study were designed to block survivin
expression by targeting exons and introns. A U6-promoter-mediated
shRNA recombinant plasmid (pGP-U6-GFP-neo-Survivin) was utilized to
inactivate survivin at multiple sites via multiple shRNA sequences.
These plasmids were transfected into HeLa cells to evaluate the
effectiveness of the survivin gene in vitro. All the shRNAs
generated significantly downregulated expression of the target gene
and protein, as assessed by RT-qPCR and western blot analysis.
Notably, both FCM detection and Hochest 33258 staining revealed an
increased degree of apoptosis in HeLa cells induced by survivin
inactivation.
RNAi is an evolutionary conserved regulatory
mechanism by which siRNAs induce the cleavage and degradation of
homologous mRNA molecules specifically by transcriptional gene
silencing (TGS) and post-TGS and (15). The shRNAs that target exons can
decrease expression of survivin mRNA and protein in the cytoplasm;
however, the intron-specific shRNAs cannot find the homologous
mRNAs in cytoplasm, so they cannot have mediated their inhibitory
role in this way. Previous reports have demonstrated that RNAi can
suppress gene expression through TGS in plants (46,47),
Drosophila melanogaster (48),
Caenorhabditis elegans (49,50), and
Schizosaccharomyces pombe (51). Many studies have revealed that
RNAi-mediated TGS in mammalian cells does occur at the nuclear
level, via RNA-directed DNA methylation, RNAi-mediated
heterochromatin formation and the modification of histones
(52–54). In the present study, we hypothesize
that the shRNAs targeting introns may have blocked survivin gene
expression through TGS.
Upon entry into the nucleus, shRNAs are processed by
the microprocesor complex containing the RNase-III enzyme Drosha
and DiGeorge Syndrome Critical Region 8 (55,56) into a
form that can be recognized by exportin-5, a Ran-GTP-dependent
nucleocytoplasmic transporter (57,58). The
modified shRNAs are then exported from the nucleus into the
cytoplasm by exportin-5. In the cytoplasm, the functional siRNAs
are cleaved by Dicer in a complex with protein kinase R activator
of transcription and Tar-RNA-binding protein. The siRNAs are loaded
into Argonaute proteins (59,60), forming the pre-RNA-induced initiator
of transcriptional gene silencing (pre-RISC) complex. With the
assistance of partners, such as Dicer and Argonaute proteins
(49,61), the pre-RISC complex translocates into
the nucleus, where the RISC complex matures. Single stranded siRNAs
directly bind to complementary DNA sequences, leading to DNA
methylation, heterochromatin formation and the post-translational
modification of histones, such as the trimethylation of Lysine 27
of histone H3 (H3K27me3), histone deacetylation, dimethylation of
lysine 9 of histone H3 Lys9 (H3K9me2) (62,63).
Several proteins may be involved in this model, including
histone-lysine-methyltransferases, Histone-deacetylases, DNA
methyltransferases, and histone protein 1.
Another alternative explanation for this
experimental phenomenon is that exogenous siRNAs that target intron
sequences may control alternative splicing, in a process similar to
that of TGS (64). In this silencing
pathway, the mature siRNAs directly recognize nascent pre-mRNAs by
targeting intronic sequences, leading to heterochromatin formation
(H3K9 dimethylation and H3K27 trimethylation) and DNA methylation
by recruiting associated enzymes, subsequently modulating
alternative splicing. As in TGS, Argonaute proteins, particularly
Argonaute 1 (AGO1) and Dicer, are also involved in this mechanism
(63). It is possible that once the
siRNAs bind to the nascent pre-mRNAs with AGO1 and Dicer, they may
cleave them directly to trigger transcriptional change. However,
the other possible mechanisms cannot be excluded as explanations of
the results of the present study. Whether the shRNAs target intron
sequences to suppress survivin expression in vitro through
the aforementioned mechanism remains unclear and will be revealed
by further experiments.
In summary, the results of the present study
demonstrate that the inhibition of survivin expression by shRNA may
be a potential tool for cancer therapy and may provide further
options for the design of interference target sites.
Acknowledgements
The authors would like to thank Professor Hou and
Professor Huang for their guidance and Li Guodong for assistance
with statistical analysis. The present study was supported by the
Science and Technology Program of Higher Learning Institutions
(Dongguan, China; grant nos. 200910815264 and 2012108102016).
References
|
1
|
Fulda S: Inhibitor of Apoptosis (IAP)
proteins in hematological malignancies: Molecular mechanisms and
therapeutic opportunities. Leukemia. 28:1414–1422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wheatley S and McNeish IA: Survivin: A
protein with dual roles in mitosis and apoptosis. Int Rev Cytol.
247:35–88. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soleimanpour E and Babaei E: Survivin as a
potential target for cancer therapy. Asian Pac J Cancer Prev.
16:6187–6191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sanhueza C, Wehinger S, Bennett Castillo
J, Valenzuela M, Owen GI and Quest AF: The twisted survivin
connection to angiogenesis. Mol Cancer. 14:1982015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mobahat M, Narendran A and Riabowol K:
Survivin as a preferential target for cancer therapy. Int J Mol
Sci. 15:2494–2516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lladser A, Sanhueza C, Kiessling R and
Quest AF: Is survivin the potential Achilles' heel of cancer? Adv
Cancer Res. 111:1–37. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Altieri DC: Targeting survivin in cancer.
Cancer Lett. 332:225–228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li F, Ambrosini G, Chu EY, Plescia J,
Tognin S, Marchisio PC and Altieri DC: Control of apoptosis and
mitotic spindle checkpoint by survivin. Nature. 396:580–584. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao J, Tenev T, Martins LM, Downward J
and Lemoine NR: The ubiquitin-proteasome pathway regulates survivin
degradation in a cell cycle-dependent manner. J Cell Sci.
113:4363–4371. 2000.PubMed/NCBI
|
|
11
|
Chen X, Duan N, Zhang C and Zhang W:
Survivin and tumorigenesis: Molecular mechanisms and therapeutic
strategies. J Cancer. 7:314–323. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Altieri DC: Validating survivin as a
cancer therapeutic target. Nat Rev Cancer. 3:46–54. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tran J, Master Z, Yu JL, Rak J, Dumont DJ
and Kerbel RS: A role for survivin in chemoresistance of
endothelial cells mediated by VEGF. Proc Natl Acad Sci USA.
99:4349–4354. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Coumar MS, Tsai FY, Kanwar JR, Sarvagalla
S and Cheung CH: Treat cancers by targeting survivin: Just a dream
or future reality? Cancer Treat Rev. 39:802–811. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fire A, Xu S, Montgomery MK, Kostas SA,
Driver SE and Mello CC: Potent and specific genetic interference by
double-stranded RNA in Caenorhabditis elegans. Nature.
391:806–811. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Y, Chang S, Sun J, Zhu S, Pu C, Li
Y, Zhu Y, Wang Z and Xu RX: Targeted microbubbles for ultrasound
mediated short hairpin RNA plasmid transfection to inhibit survivin
gene expression and induce apoptosis of ovarian cancer A2780/DDP
cells. Mol Pharm. 12:3137–3145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Scholz C and Wagner E: Therapeutic plasmid
DNA versus siRNA delivery: Common and different tasks for synthetic
carriers. J Control Release. 161:554–565. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Paddison PJ, Caudy AA, Bernstein E, Hannon
GJ and Conklin DS: Short hairpin RNAs (shRNAs) induce
sequence-specific silencing in mammalian cells. Genes Dev.
16:948–958. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ling X and Li F: Silencing of
antiapoptotic survivin gene by multiple approaches of RNA
interference technology. Biotechniques. 36(450–454): 456–460.
2004.
|
|
20
|
Hannon GJ and Rossi JJ: Unlocking the
potential of the human genome with RNA interference. Nature.
431:371–378. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mohr SE and Perrimon N: RNAi screening:
New approaches, understandings, and organisms. Wiley Interdiscip
Rev RNA. 3:145–158. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pan Q, van der Laan LJ, Janssen HL and
Peppelenbosch MP: A dynamic perspective of RNAi library
development. Trends Biotechnol. 30:206–215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu G and Luo J: A primer on using pooled
shRNA libraries for functional genomic screens. Acta Biochim
Biophys Sin (Shanghai). 44:103–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brummelkamp TR, Bernards R and Agami R: A
system for stable expression of short interfering RNAs in mammalian
cells. Science. 296:550–553. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li QX, Zhao J, Liu JY, Jia LT, Huang HY,
Xu YM, Zhang Y, Zhang R, Wang CJ, Yao LB, et al: Survivin stable
knockdown by siRNA inhibits tumor cell growth and angiogenesis in
breast and cervical cancers. Cancer Biol Ther. 5:860–866. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Igney FH and Krammer PH: Death and
anti-death: Tumour resistance to apoptosis. Nat Rev Cancer.
2:277–288. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mita AC, Mita MM, Nawrocki ST and Giles
FJ: Survivin: Key regulator of mitosis and apoptosis and novel
target for cancer therapeutics. Clin Cancer Res. 14:5000–5005.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Altieri DC: New wirings in the survivin
networks. Oncogene. 27:6276–6284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Grossman D, McNiff JM, Li F and Altieri
DC: Expression and targeting of the apoptosis inhibitor, survivin,
in human melanoma. J Invest Dermatol. 113:1076–1081. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yuan QZ, Wang CT, Mao YQ, Zhang P, Shi HS,
Li ZY, Pan L, Yu DD, Leng F, Chen X, et al: Enhanced tumor
radiosensitivity by a survivin dominant-negative mutant. Oncol Rep.
23:97–103. 2010.PubMed/NCBI
|
|
32
|
Pennati M, Binda M, Colella G, Folini M,
Citti L, Villa R, Daidone MG and Zaffaroni N: Radiosensitization of
human melanoma cells by ribozyme-mediated inhibition of survivin
expression. J Invest Dermatol. 120:648–654. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pennati M, Binda M, De Cesare M, Pratesi
G, Folini M, Citti L, Daidone MG, Zunino F and Zaffaroni N:
Ribozyme-mediated down-regulation of survivin expression sensitizes
human melanoma cells to topotecan in vitro and in vivo.
Carcinogenesis. 25:1129–1136. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang Z, Wang L, Wang H, Shang X, Niu W, Li
J and Wu Y: A novel mimovirus vaccine containing survivin epitope
with adjuvant IL-15 induces long-lasting cellular immunity and high
antitumor efficiency. Mol Immunol. 45:1674–1681. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang X, Chen Y, Ren J and Qu X: Small
interfering RNA for effective cancer therapies. Mini Rev Med Chem.
11:114–124. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Joo MK, Yhee JY, Kim SH and Kim K: The
potential and advances in RNAi therapy: Chemical and structural
modifications of siRNA molecules and use of biocompatible
nanocarriers. J Control Release. 193:113–121. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Carvalho A, Carmena M, Sambade C, Earnshaw
WC and Wheatley SP: Survivin is required for stable checkpoint
activation in taxol-treated HeLa cells. J Cell Sci. 116:2987–2998.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guo K, Song W, Gong Y, Hu S, Zhong W and
Qiu W: Down-Regulation of survivin expression by siRNA suppresses
proliferation and enhances chemosensitivity in human pancreatic
cancer cell line Panc-1. In: Proceedings of the 2015 Seventh
International Conference on Measuring Technology and Mechatronics
Automation. IEEE, Nanchang. 400–402. 2015.
|
|
39
|
Habib R, Akhtar J, Taqi M, Yu C and Zhang
C: Lentiviral vector-mediated survivin shRNA delivery in gastric
cancer cell lines significantly inhibits cell proliferation and
tumor growth. Oncol Rep. 34:859–867. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zheng H, Tang C and Yin C: Oral delivery
of shRNA based on amino acid modified chitosan for improved
antitumor efficacy. Biomaterials. 70:126–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Berget SM, Moore C and Sharp PA: Spliced
segments at the 5′terminus of adenovirus 2 late mRNA. Proc Natl
Acad Sci USA. 74:3171–3175. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chow LT, Gelinas RE, Broker TR and Roberts
RJ: An amazing sequence arrangement at the 5′ ends of adenovirus 2
messenger RNA. Cell. 12:1–8. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pan Q, Shai O, Lee LJ, Frey BJ and
Blencowe BJ: Deep surveying of alternative splicing complexity in
the human transcriptome by high-throughput sequencing. Nat Genet.
40:1413–1415. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chorev M and Carmel L: The function of
introns. Front Genet. 3:552012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lunghi M, Spano F, Magini A, Emiliani C,
Carruthers VB and Di Cristina M: Alternative splicing mechanisms
orchestrating post-transcriptional gene expression: Intron
retention and the intron-rich genome of apicomplexan parasites.
Curr Genet. 62:31–38. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Matzke MA, Primig M, Trnovsky J and Matzke
AJ: Reversible methylation and inactivation of marker genes in
sequentially transformed tobacco plants. EMBO J. 8:643–649.
1989.PubMed/NCBI
|
|
47
|
Zilberman D, Cao X and Jacobsen SE:
ARGONAUTE4 control of locus-specific siRNA accumulation and DNA and
histone methylation. Science. 299:716–719. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Pal-Bhadra M, Bhadra U and Birchler JA:
RNAi related mechanisms affect both transcriptional and
posttranscriptional transgene silencing in Drosophila. Mol
Cell. 9:315–327. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dernburg AF, Zalevsky J, Colaiácovo MP and
Villeneuve AM: Transgene-mediated cosuppression in the C.
elegans germ line. Genes Dev. 14:1578–1583. 2000.PubMed/NCBI
|
|
50
|
Grishok A, Sinskey JL and Sharp PA:
Transcriptional silencing of a transgene by RNAi in the soma of
C. elegans. Genes Dev. 19:683–696. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lippman Z, May B, Yordan C, Singer T and
Martienssen R: Distinct mechanisms determine transposon inheritance
and methylation via small interfering RNA and histone modification.
PLoS Biol. 1:E672003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Matzke MA and Birchler JA: RNAi-mediated
pathways in the nucleus. Nat Rev Genet. 6:24–35. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Morris KV: RNA-mediated transcriptional
gene silencing in human cells. Curr Top Microbiol Immunol.
320:211–224. 2008.PubMed/NCBI
|
|
54
|
Morris KV, Chan SW, Jacobsen SE and Looney
DJ: Small interfering RNA-induced transcriptional gene silencing in
human cells. Science. 305:1289–1292. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J,
Lee J, Provost P, Radmark O, Kim S, et al: The nuclear RNase III
Drosha initiates microRNA processing. Nature. 425:415–419. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mendez C, Ahlenstiel CL and Kelleher AD:
Post-transcriptional gene silencing, transcriptional gene silencing
and human immunodeficiency virus. World J Virol. 4:219–244. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lund E, Güttinger S, Calado A, Dahlberg JE
and Kutay U: Nuclear export of microRNA precursors. Science.
303:95–98. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ohrt T, Merkle D, Birkenfeld K, Echeverri
CJ and Schwille P: In situ fluorescence analysis demonstrates
active siRNA exclusion from the nucleus by Exportin 5. Nucleic
Acids Res. 34:1369–1380. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Gagnon KT and Corey DR: Argonaute and the
nuclear RNAs: New pathways for RNA-mediated control of gene
expression. Nucleic Acid Ther. 22:3–16. 2012.PubMed/NCBI
|
|
60
|
Ohrt T, Mutze J, Staroske W, Weinmann L,
Hock J, Crell K, Meister G and Schwille P: Fluorescence correlation
spectroscopy and fluorescence cross-correlation spectroscopy reveal
the cytoplasmic origination of loaded nuclear RISC in vivo in human
cells. Nucleic Acids Res. 36:6439–6449. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Verdel A, Vavasseur A, Le Gorrec M and
Touat-Todeschini L: Common themes in siRNA-mediated epigenetic
silencing pathways. Int J Dev Biol. 53:245–257. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Castel SE and Martienssen RA: RNA
interference in the nucleus: Roles for small RNAs in transcription,
epigenetics and beyond. Nat Rev Genet. 14:100–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Burger K and Gullerova M: Swiss army
knives: Non-canonical functions of nuclear Drosha and Dicer. Nat
Rev Mol Cell Biol. 16:417–430. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Allo M, Buggiano V, Fededa JP, Petrillo E,
Schor I, de la Mata M, Agirre E, Plass M, Eyras E, Elela SA, et al:
Control of alternative splicing through siRNA-mediated
transcriptional gene silencing. Nat Struct Mol Biol. 16:717–724.
2009. View Article : Google Scholar : PubMed/NCBI
|