Introduction
Lung cancer is the most prevalent and lethal type of
cancer worldwide (1). Non-small cell
lung cancer (NSCLC) is the most common subtype of lung cancer,
accounting for 85% of cases globally in 2012 (2,3). Gefitinib
is an epidermal growth factor receptor (EGFR)-associated tyrosine
kinase inhibitor (TKI) (4), and is an
anticancer targeted drug that was developed as an effective
clinical therapy for patients with NSCLC. However, data from
clinical studies indicate that numerous patients with cancer
develop gefitinib resistance, leading to poorer treatment outcomes
(5), and this acquired resistance
limits the further applications of the drug. Therefore, improving
gefitinib efficacy and to increasing the benefits of TKI therapy
conferred to patients with NSCLC is of clinical importance.
Sulforaphane (SFN) is an isothiocyanate that exists
as conjugates in the genus Brassica of cruciferous
vegetables; SFN has been demonstrated to inhibit the malignant
growth of various cancer cell types with little or no toxicity
towards normal cells (6,7). SFN is a potent anticancer agent, but its
underlying mechanisms and molecular targets remain unclear.
Hedgehog signaling has an essential role in the
control of stem cell growth in embryonic tissues, and it is
essential for the development of tissues and organs (8). The sonic hedgehog (SHH) signaling
pathway controls cell proliferation and differentiation during
embryonic development, and contributes to tumorigenesis when
mutated or dysregulated (9).
Furthermore, aberrant activation of the SHH signaling pathway
serves a critical role in the tumorigenesis and progression of lung
cancer (10–12).
Cancer stem cells (CSCs) (13) are a rare population of
undifferentiated tumorigenic cells responsible for tumor
initiation, maintenance and metastasis. These cells exhibit
unlimited proliferation potential, self-renewal and the capacity to
generate a progeny of differentiated cells that constitute the
major tumor population. CSCs are more resistant to standard
chemotherapy drugs, and employ various signaling pathways (14–16).
Rodova et al (17) reported
that SHH signaling regulates the self-renewal of pancreatic CSCs.
Eramo et al (18) dissociated
CD133+ cells from lung tumor tissue specimens, and
confirmed these cells have characteristics of CSCs. Additionally,
they identified that CD133+ cells are resistant to
conventional chemotherapy. CD44 was also defined as a surface
marker of CSCs (15). Therefore, the
membrane antigens CD133 and CD44 are shared among CSCs (19–22).
Cross-talk between the SHH and epidermal growth
factor receptor (EGFR) pathways involved in carcinogenesis have
recently been examined; these pathways co-operate during disease
initiation and progression, resulting in aggressive metastasis
(23). A prior study investigated the
co-targeting of the SHH and EGFR signaling pathways as a novel
approach to overcoming treatment resistance and eliminating CSCs
(24,25).
It has reported that SFN inhibits the expression of
key SHH factors in numerous cancer types (7,17,26). However, it is unclear whether SFN can
reverse gefitinib resistance in human lung cancer cells. The
present study was based on the hypothesis that SFN reverses
gefitinib tolerance in lung cancer cells by modulating the SHH
signaling pathway. The present study aimed to explore the potential
molecular mechanisms of SFN, and the feasibility of utilizing SFN
to reverse gefitinib resistance in human lung cancer cells via
targeting the SHH signaling pathway. The current study also aimed
to provide experimental evidence for subsequent clinical
applications, and to identify novel agents effective in the
treatment of gefitinib-resistant lung cancer.
Materials and methods
Cell culture
The NSCLC PC9 cells were provided by the Cancer
Institute of Southern Medical University (Guangzhou, China), and
maintained in RPMI-1640 medium (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA) at 37°C in a
humidified atmosphere of 5% CO2.
Establishment of a gefitinib-tolerant
cell line
Gefitinib-tolerant cells were developed through
chronic, repeated exposure to gefitinib. Briefly, PC9 cells were
exposed to 0.002 µmol/l gefitinib for 48 h in RPMI-1640 medium
containing 10% fetal bovine serum. Cells were then washed and
cultured in drug-free medium until they reached the logarithmic
growth phase. Subsequently, the cells were re-exposed to increasing
concentrations (0.002, 0.020, 0.050, 0.100, 0.250, 0.500, 1.000,
2.000 µmol/l) of gefitinib. Resistant cells can survive in 2 µmol/l
gefitinib; gefitinib-tolerant cells were obtained 48 days after
initial exposure. The established resistant cell line was
maintained in RPMI-1640 medium containing 1 µmol/l gefitinib. For
all studies, resistant cells were cultured in drug-free medium for
1 week to eliminate gefitinib. Gefitinib-tolerant cells are
referred as PC9GT cells.
Cell proliferation assay
The PC9 and PC9GT cells, used in the logarithmic
phase of growth, were seeded into 96-well plates (3,000
cells/well). When the cells had adhered, various concentrations of
gefitinib were added to the wells; the final concentrations were
0.002, 0.020, 0.050, 0.100, 0.250, 0.500, 1.000 and 2.000 µmol/l.
Following incubation for 48 h, 10 µl MTT reagent was added into
each well of the plates. After an additional incubation period of 4
h, 150 µl DMSO reagent was added, and then the optical density (OD)
at 490 nm was determined using a micro plate reader (Beckman
Coulter, Inc., Brea, CA, USA). Each drug concentration was
evaluated in triplicate wells. The number of cells was counted
using Countstar (version IC-1000; Rui Yu Biological Technology Co.,
Ltd., Shanghai, China). The density of cells was observed with a
inverted microscope system (magnification, ×40; Eclipse Ti-s,
Nikon, Tokyo, Japan).
Western blot analysis
Total protein was extracted from 2–5×106
PC9 and PC9GT cells with an ice-cold SDS protein lysis buffer at
100°C for 10 min. Protein concentration was measured using the
Micro BCA Protein Assay Reagent kit (Sangon Biotech Co., Ltd.,
Shanghai, China). Proteins were separated using a 12.5% gel and
SDS-PAGE and transferred onto polyvinylidene difluoride membranes
(EMD Millipore, Billerica, MA, USA). The membranes were blocked
with 5% non-fat dry milk in TBS buffer for 1 h at room temperature
and subsequently incubated with primary antibodies against CD133,
CD44, SHH, SMO, GLI1, GLI2 and GAPDH (1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA) overnight at 4°C, washed three
times with TBST (0.05% Tween 20), and then incubated with the
secondary antibodies conjugated with horseradish peroxidase at
1:3,000 dilutions in TBS for 2 h at room temperature. Membranes
were then washed again in TBS-Tween 20 three times at room
temperature. Protein bands were visualized on an X-ray film using
chemiluminescent horseradish peroxidase substrate (SurModics, Inc.,
Eden Prairie, MN, USA). A Tanon v.3500 Gel Imaging System (Tanon
Co., Shanghai, China) was used for the semi-quantitative analysis
of proteins.
Statistical analysis
Statistical analysis was performed using SPSS
version 16.0 software (SPSS, Inc., Chicago, IL, USA). The data are
expressed as the means ± standard deviation. The Student's t-test
was used for comparisons between two groups. Multigroup comparisons
of the means were performed using one-way analysis of variance and
post hoc analysis with a Student-Newman-Keuls test. P<0.05 was
considered to indicate a statistically significant difference.
Results
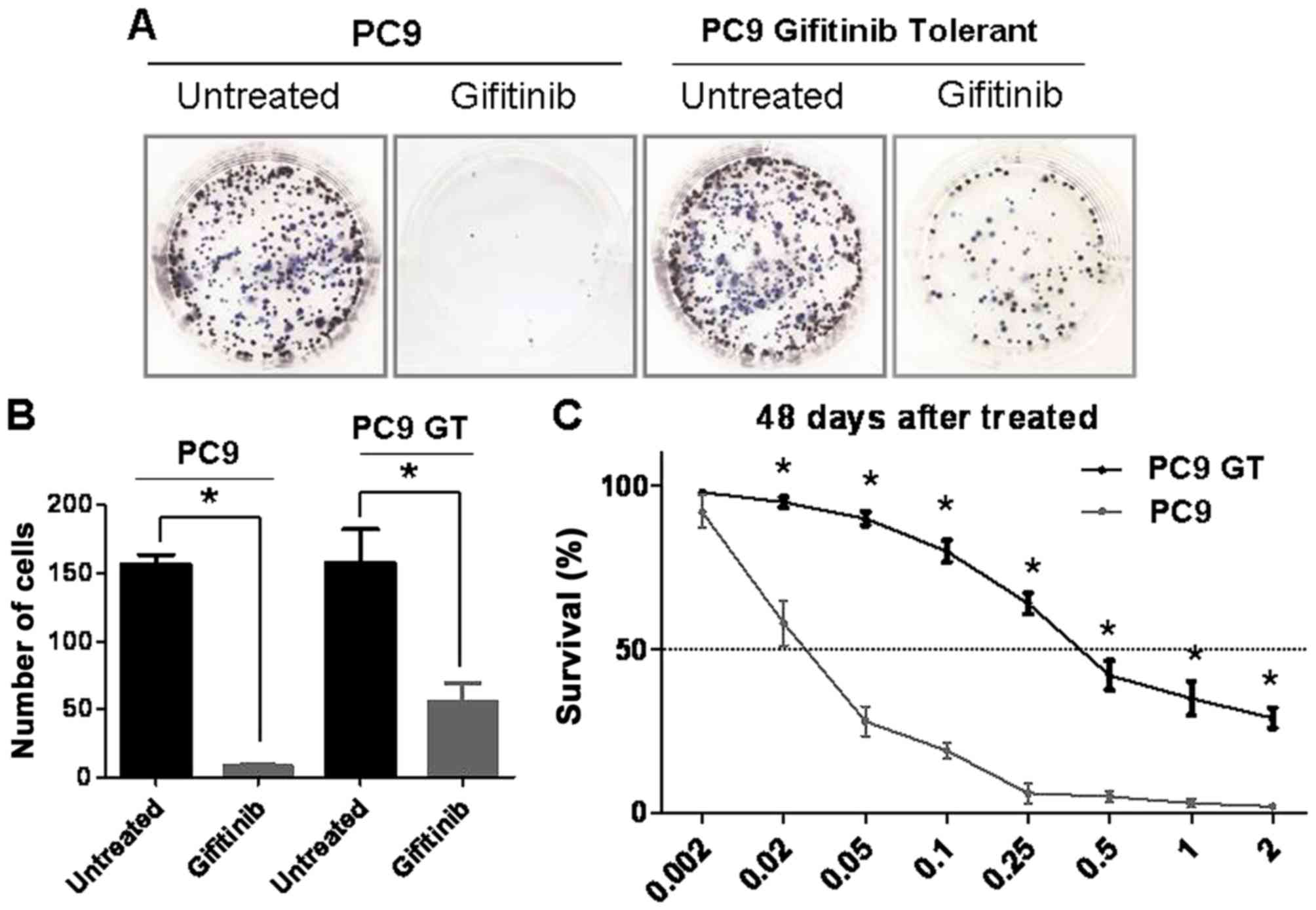
PC9GT cell lines were established
To generate a gefitinib-tolerant subline of PC9
cells, the cells were cultured with increasing concentrations of
gefitinib over a period of 48 days. To confirm that the new cell
line was established successfully, PC9GT and PC9 cells were divided
into two groups. One group was treated with gefitinib and the other
remained untreated (Fig. 1A). The
cell count demonstrated that PC9GT cells continued to proliferate,
contrary to the PC9 cells (P<0.05; Fig. 1B). The PC9GT cell line could be
serially passed in gefitinib-free media for >15 generations
without a change in sensitivity to gefitinib. The MTT assay showed
that the PC9GT cells could proliferate in response to exposure to 2
µmol/l gefitinib, compared with PC9 cells (P<0.05; Fig. 1C).
Activation of the SHH signaling
pathway affects gefitinib sensitivity
Aberrant activation of the SHH signaling pathway
serves a critical role in the tumorigenesis and progression of lung
cancer. To determine whether the SHH signaling pathway is
associated with gefitinib resistance, the key proteins of the SHH
signaling pathway in PC9 and PC9GT cells were detected via
western-blot analysis. Notably, robust expression of SHH, SMO, GLI1
and GLI2 was observed in PC9GT cells, as compared with in the PC9
cells (Fig. 2A).
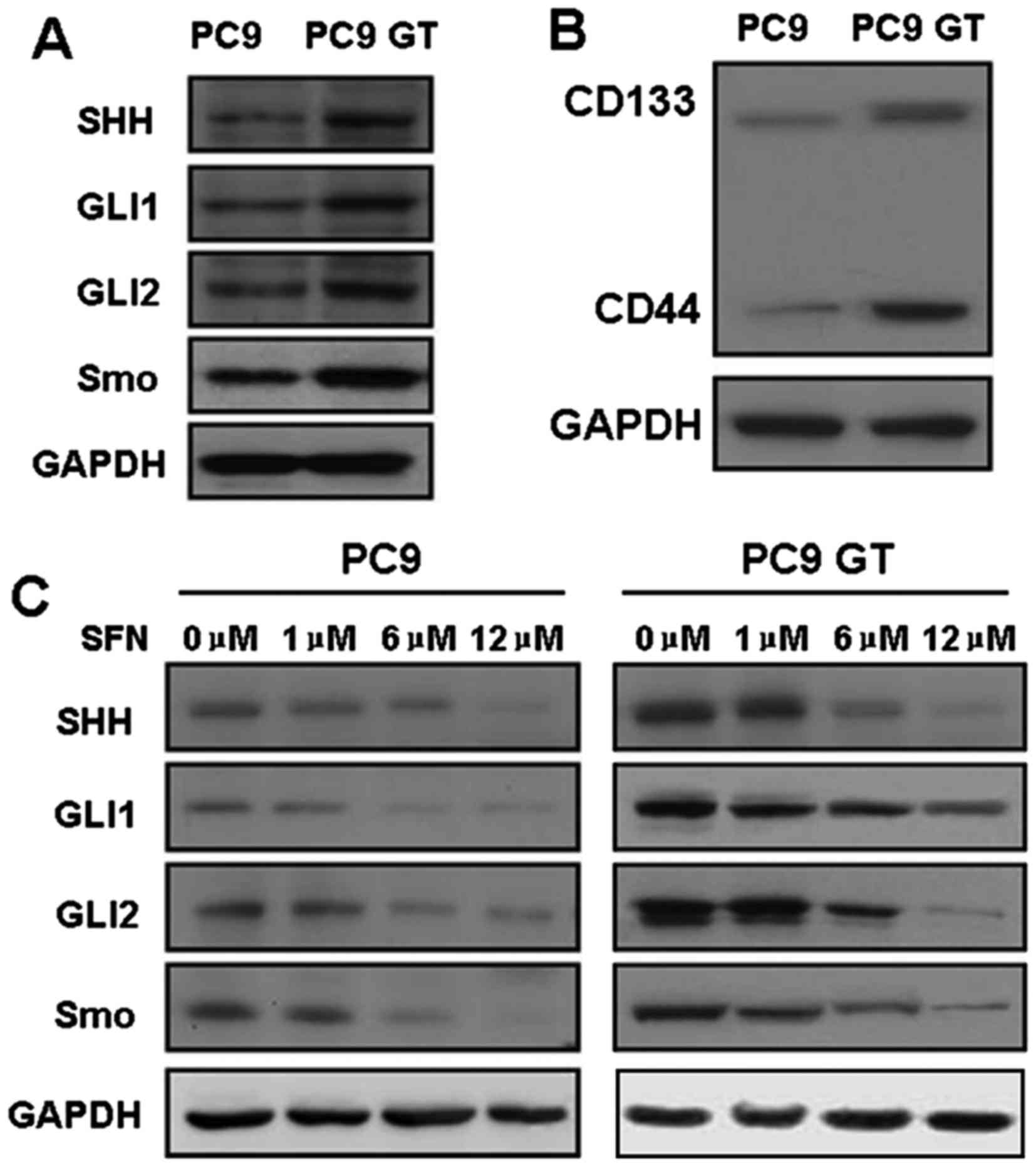 | Figure 2.The expression of key SHH and the
marker of LSCs in lung cancer cells was analyzed via western
blotting. (A) The expression of SHH, SMO, GLI1 and GLI2 was high in
PC9GT cells, as compared with in PC9 cells. (B) The expression of
CD133 and CD44 was upregulated in PC9GT cells, compared with in PC9
cells. (C) SFN markedly inhibited the expression of SHH, SMO and
GLI1 in PC9GT and PC9 cells, in a dose-dependent manner. SFN,
sulforaphane; PC9GT, gefitinib-tolerant PC9 cells; EGFR, epidermal
growth factor receptor; SHH, sonic hedgehog; SMO, smoothened;
GLI1/2, zinc finger protein GLI1/2. |
The existence of CSCs affects
gefitinib sensitivity
The presence of CSCs in lung cancer may render
standard chemotherapy ineffective and aid the cancer cells in
resisting anti-cancer drugs (27).
Previous reports suggest that a CD133- and CD44-positive
subpopulation of multipotent cells, with extensive proliferative
and self-renewal characteristics, have the biological features of
CSCs (19,20). Mizugaki et al (21) reported that the expression of CD133 is
a potential prognostic marker for NSCLC. CD133 and CD44 have been
established as representative markers for CSC in diverse types of
cancer (28). To determine whether
the CSCs were associated with gefitinib resistance, the protein
expression of CD133 and CD44 were examined using western blotting.
The expression of CD133 and CD44 was upregulated in PC9GT cells,
compared with in PC cells (Fig. 2B).
Upregulated CD133and CD44 were also revealed to be associated with
significantly poorer prognosis for NSCLC. Upregulated expression of
CD133 and CD44 could therefore be associated with gefitinib
resistance.
Sulforaphane inhibits PC9 and PC9GT
cell proliferation and enhances the chemotherapeutic effect of
gefitinib
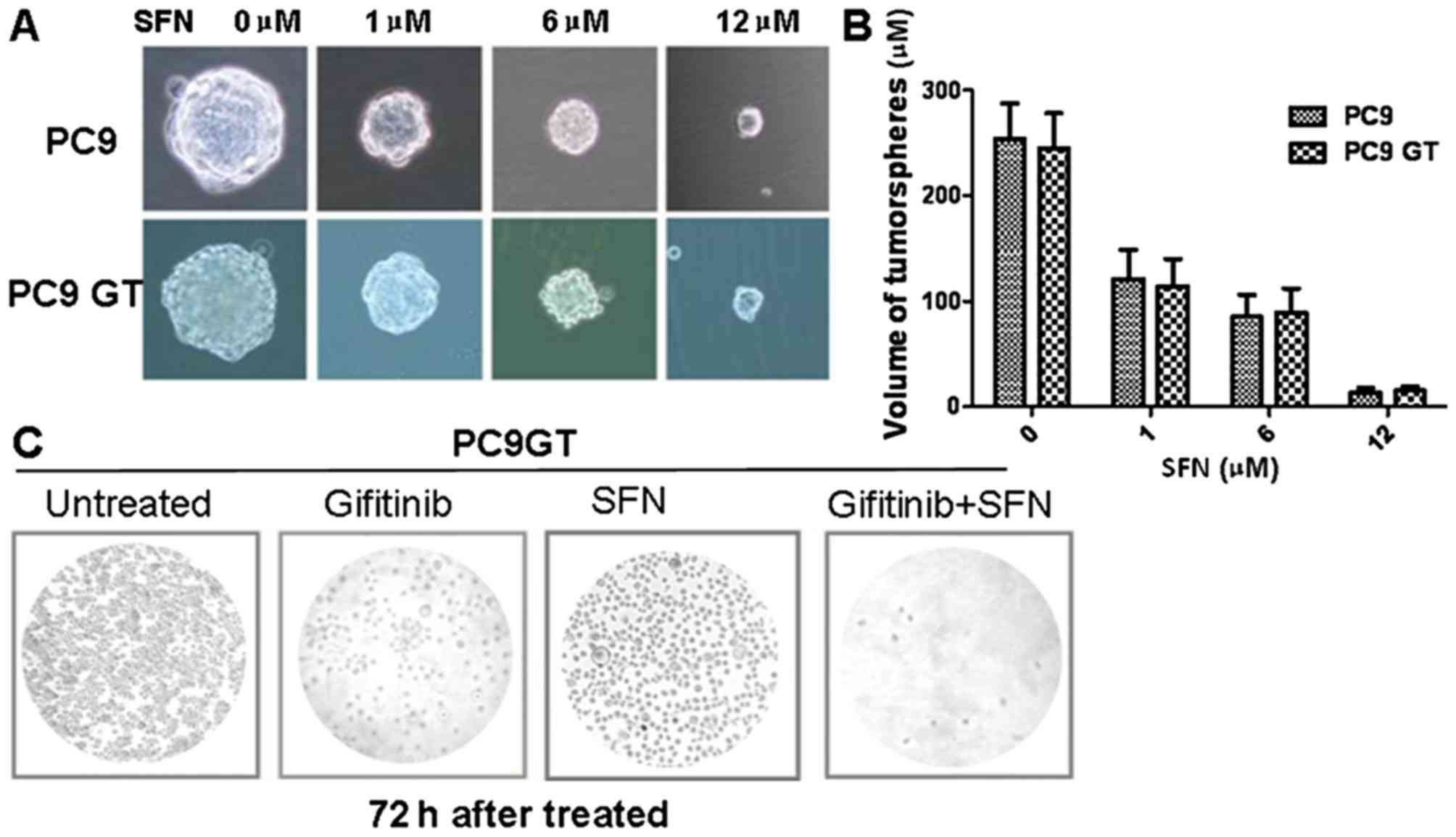
A sphere formation assay was performed to
investigate whether SFN sensitizes gefitinib-tolerant lung cancer
cells. The PC9GT cells and PC9 cells were treated with 0, 1, 6 and
12 µmol/l SFN. After 1 week, the total number of spheres and the
tumorsphere volumes were measured; the tumorsphere volumes were
observed to gradually decrease with the increasing dose of SFN
(Fig. 3A). SFN markedly inhibited the
proliferation of PC9GT and PC9 cells in a dose-dependent manner,
although there was no significant difference between the PC9GT and
PC9 cells (Fig. 3B), indicating that
SFN can effectively inhibit lung cancer cell proliferation, and is
unaffected by gefitinib-resistance.
The effects of combined therapy with SFN and
gefitinib were then assessed in PC9GT cells. The PC9GT cells were
treated with SFN, gefitinib or combined SFN/gefitinib. The combined
treatment was observed to markedly decrease PC9GT cell
proliferation, when compared with SFN or gefitinib administered
alone (Fig. 3C). The combined
treatment significantly decreased PC9GT cell proliferation, when
compared with SFN or gefitinib alone, after 48 and 72 h (P<0.05;
Fig. 4A).
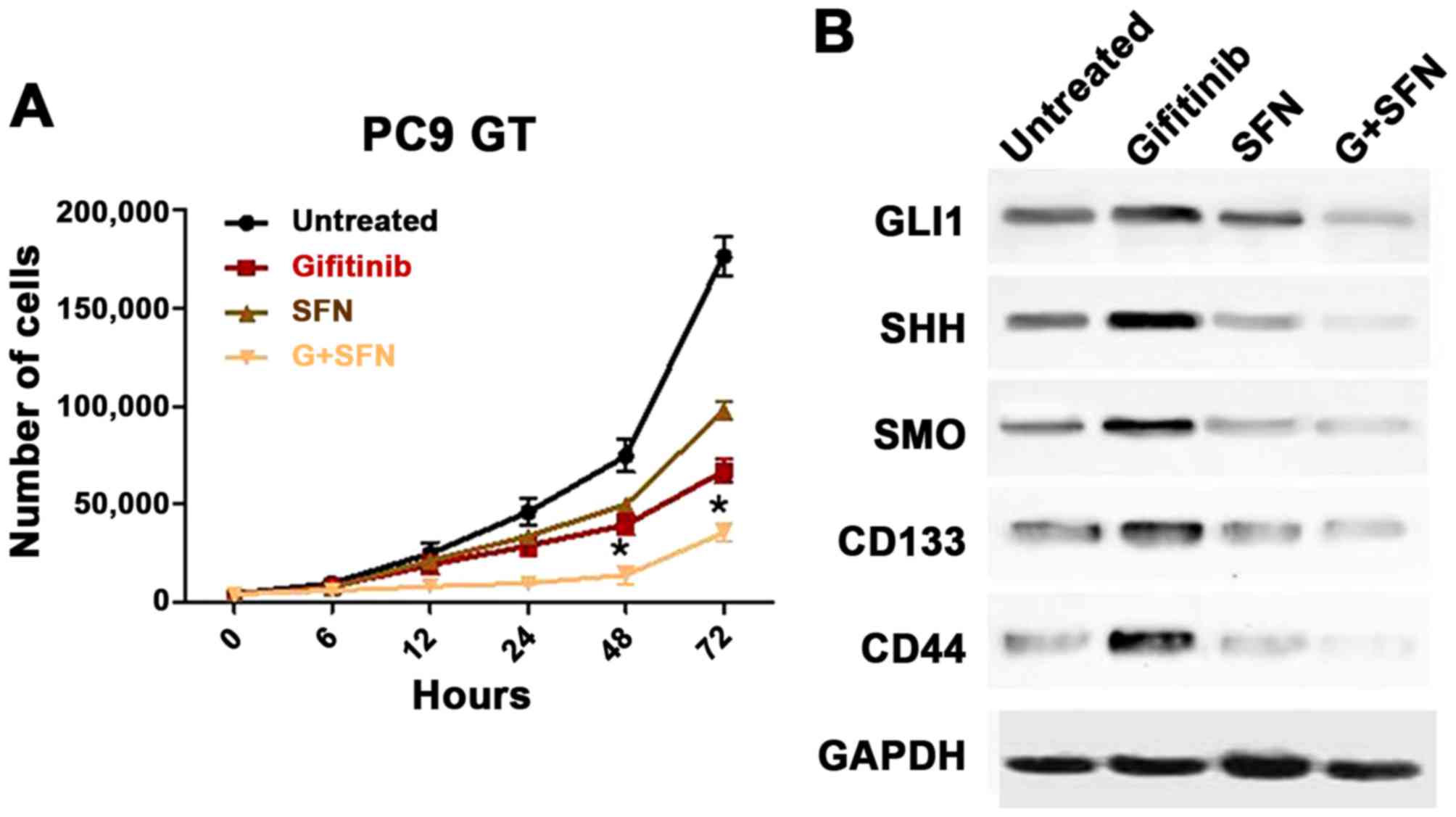 | Figure 4.The effect of combined SFN and
gefitinib therapy on PC9GT cells was determined using a cell
proliferation assay and western blotting. (A) Combined treatment
markedly decreased PC9GT cell proliferation, when compared with SFN
or gefitinib monotherapy, at 48 and 72 h (*P<0.05, compared with
SFN, gefitinib and the control). (B) SFN combined with gefitinib
markedly inhibited the expression of SHH, SMO, GLI1, CD133 and CD44
in PC9GT cells, as compared with SFN or gefitinib administered
alone. SFN, sulforaphane; PC9GT, gefitinib-tolerant PC9 cells;
EGFR, epidermal growth factor receptor; SHH, sonic hedgehog; SMO,
Smoothened; GLI1, zinc finger protein GLI1. |
Sulforaphane reverses gefitinib
resistance via modulation of the SHH signaling pathway
To explore the mechanism underlying the
anti-proliferative effect of SFN on PC9GT cells, the SHH signaling
pathway proteins CD133 and CD44 were examined via western blotting
in SFN-treated PC9 and PC9GT cells. Notably, SFN markedly inhibited
the expression of SHH, SMO and GLI1 in PC9GT and PC9 cells in a
dose-dependent manner. Furthermore, the expression of these
proteins did not significantly differ between the PC9GT and PC9
cells (Fig. 2C). The effects of
combined SFN/gefitinib therapy PC9GT cells were then assessed. The
results indicate that SFN combined with gefitinib markedly inhibits
the expression of SHH, SMO, GLI1, CD133 and CD44 in PC9GT cells,
when compared with SFN or gefitinib administered alone (Fig. 4B). The results suggested that SFN may
reverse gefitinib resistance by inhibiting the SHH pathway. In the
present study, it was also identified that SFN can inhibit the
expression of CD133 and CD44 in PC9GT cells.
Discussion
Gefitinib is effective for patients with NSCLC, but
numerous patients develop gefitinib resistance, which leads to poor
treatment outcomes. Acquired resistance to gefitinib is a serious
clinical problem; therefore, the clinical use of gefitinib has been
limited.
SFN exists as conjugates in cruciferous vegetables,
and has been demonstrated to inhibit the malignant growth of cancer
cells with little or no toxicity towards normal cells. The present
study evidenced that SFN may inhibit the proliferation of PC9
cells, but also inhibit the proliferation of PC9 GT cells. The
present study could be important in searching for a novel potential
agent against gefitinib-resistant lung cancer.
Further studies are required to clarify the
mechanism underlying SFN-mediated inhibition of gefitinib
resistance. The results indicated that components of the SHH
signaling pathway are highly expressed in PC9GT cells. To identify
the function of SFN in the SHH signaling pathway, the expression of
SHH, GLI1, GLI2 and SMO was measured in PC9GT cells following
treatment with SFN, gefitinib and SFN and gefitinib. The present
study suggested that SFN may inhibit the expression of certain SHH
signaling pathway-associated proteins, including SHH, GLI1, GLI2
and SMO, in PC9GT cells. Gefitinib is an inhibitor of the
EGFR-associated TK, and exerts antitumor effects via the blockade
of EGFR-associated TK activity (29).
It is evident the SHH signaling pathway co-operates with EGFR
during cancer initiation, and the progression to aggressive,
invasive and metastatic disease stages (23,24).
Concordantly, combined therapy with the SHH signaling pathway
antagonist and the EGFR inhibitor would be more efficient in
inhibiting invasiveness and inducing cancer cell apoptosis compared
with either drug alone. Therefore, combining SFN and gefitinib
meets the aforementioned requirements; the effectiveness of SFN in
inhibiting PC9GT cell growth may be associated with activation of
the SHH signaling pathways.
Recent studies have suggested that SHH serves a
critical role in maintaining the CSC pool (30). CD44 and CD133 are defined as CSC
surface markers (18,31). Lung cancer may resist conventional
chemotherapy due to the presence of the rare CSC (31). CSCs are not only resistant to standard
chemotherapy drugs, but are also associated with multiple signaling
pathways, including HH, Notch and Wnt signaling pathways (15,32,33). A
previous study demonstrated that increased CD133 expression was
associated with chemoresistance and poorer clinical outcomes in
lung cancer (34). The present study
revealed that the expression of CD44 and CD133 was increased in
PC9GT cells and examined whether the presence of lung CSCs could
help explain gefitinib resistance in lung cancer.
Recent studies underlining the therapeutic interest
of co-targeting the SHH and EGFR signaling pathways, and
eradicating CSCs/progenitor cells, may improve current clinical
therapies against aggressive and metastatic cancer cases. In the
current study, it was identified that SFN is effective against
gefitinib-resistant lung cancer cells and lung CSCs. SFN may
inhibit the proliferation of gefitinib-resistant lung cancer cells
via modulating the SHH signaling pathway. Furthermore, combined
treatment markedly decreases PC9GT cell proliferation when compared
with SFN or gefitinib monotherapy. A reason SFN inhibits the
proliferation of gefitinib-tolerant lung cancer cells may be due to
its modulation of SHH signaling pathway activation and the
inhibition of lung CSC self-renewal.
The results of the present study will facilitate the
identification of novel drug targets and the development of new
therapeutic strategies to block this tumorigenic cascade, thus
improving currently available cancer treatments.
To conclude, SFN was identified to be a potent
anticancer agent: In addition to inhibiting lung cancer cell
proliferation, it also kills gefitinib-tolerant cells. The data
demonstrated that SFN reverses gefitinib tolerance in human lung
cancer cells by modulating the SHH signaling pathway. Furthermore,
the combination of SFN and gefitinib could be an effective
treatment approach for lung cancer.
Acknowledgements
The present study was supported by The Program for
New Century Excellent Talents in University of China (grant no.
NCET-13-0990), The Project of Science and Technology Innovation
Talents in Universities of Henan Province (grant no. 2012HASTIT024)
and The Project of Science and Technology of Education Department
of Henan Province (grant nos. 15A320009 and 13B320220). Special
thanks are given to Dr Prajaya Shikhar Shrestha (National Academy
of Medical Science, Bir Hospital, Kathmandu, Nepal) for their
dedicated revision of the manuscript.
Glossary
Abbreviations
Abbreviations:
|
SFN
|
sulforaphane
|
|
PC9GT
|
gefitinib-tolerant PC9 cells
|
|
NSCLC
|
non-small cell lung cancer
|
|
EGFR
|
epidermal growth factor receptor
|
|
TKI
|
tyrosine kinase inhibitor
|
|
SHH
|
sonic hedgehog
|
|
CSC
|
cancer stem cell
|
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Meoni G, Cecere FL, Lucherini E and Di
Costanzo F: Medical treatment of advanced non-small cell lung
cancer in elderly patients: A review of the role of chemotherapy
and targeted agents. J Geriatr Oncol. 4:282–290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Puglisi M, Thavasu P, Stewart A, de Bono
JS, O'Brien ME, Popat S, Bhosle J and Banerji U: AKT inhibition
synergistically enhances growth-inhibitory effects of gefitinib and
increases apoptosis in non-small cell lung cancer cell lines. Lung
Cancer. 85:141–146. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jung SK, Lee MH, Lim DY, Lee SY, Jeong CH,
Kim JE, Lim TG, Chen H, Bode AM, Lee HJ, et al: Butein, a novel
dual inhibitor of MET and EGFR, overcomes gefitinib-resistant lung
cancer growth. Mol Carcinog. 54:322–331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ge X, Chen Q, Wu YP, Zhang Y, Xia H, Yuan
D, Chen Q, Leng W, Chen L, Tang Q, et al: Induced IGF-1R activation
contributes to gefitinib resistance following combined treatment
with paclitaxel, cisplatin and gefitinib in A549 lung cancer cells.
Oncol Rep. 32:1401–1408. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clarke JD, Dashwood RH and Ho E:
Multi-targeted prevention of cancer by sulforaphane. Cancer Lett.
269:291–304. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Priya DK, Gayathri R, Gunassekaran G,
Murugan S and Sakthisekaran D: Chemopreventive role of sulforaphane
by upholding the GSH redox cycle in pre- and post-initiation phases
of experimental lung carcinogenesis. Asian Pac J Cancer Prev.
12:103–110. 2011.PubMed/NCBI
|
|
8
|
Varjosalo M and Taipale J: Hedgehog:
Functions and mechanisms. Genes Dev. 22:2454–2472. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi S, Deng YZ, Zhao JS, Ji XD, Shi J,
Feng YX, Li G, Li JJ, Zhu D, Koeffler HP, et al: RACK1 promotes
non-small-cell lung cancer tumorigenicity through activating sonic
hedgehog signaling pathway. J Biol Chem. 287:7845–7858. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Raz G, Allen KE, Kingsley C, Cherni I,
Arora S, Watanabe A, Lorenzo CD, Edwards VD, Sridhar S, Hostetter G
and Weiss GJ: Hedgehog signaling pathway molecules and ALDH1A1
expression in early-stage non-small cell lung cancer. Lung Cancer.
76:191–196. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vestergaard J, Pedersen MW, Pedersen N,
Ensinger C, Tümer Z, Tommerup N, Poulsen HS and Larsen LA: Hedgehog
signaling in small-cell lung cancer: Frequent in vivo but a rare
event in vitro. Lung Cancer. 52:281–290. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maitah MY, Ali S, Ahmad A, Gadgeel S and
Sarkar FH: Up-regulation of sonic hedgehog contributes to
TGF-beta1-induced epithelial to mesenchymal transition in NSCLC
cells. PLoS One. 6:e160682011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Allegra A, Alonci A, Penna G, Innao V,
Gerace D, Rotondo F and Musolino C: The cancer stem cell
hypothesis: A guide to potential molecular targets. Cancer Invest.
32:470–495. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tanaka H, Nakamura M, Kameda C, Kubo M,
Sato N, Kuroki S, Tanaka M and Katano M: The Hedgehog signaling
pathway plays an essential role in maintaining the CD44+CD24-/low
subpopulation and the side population of breast cancer cells.
Anticancer Res. 29:2147–2157. 2009.PubMed/NCBI
|
|
15
|
Liu M, Inoue K, Leng T, Guo S and Xiong
ZG: TRPM7 channels regulate glioma stem cell through STAT3 and
Notch signaling pathways. Cell Signal. 26:2773–2781. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Arasada RR, Amann JM, Rahman MA, Huppert
SS and Carbone DP: EGFR blockade enriches for lung cancer stem-like
cells through Notch3-dependent signaling. Cancer Res. 74:5572–5584.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rodova M, Fu J, Watkins DN, Srivastava RK
and Shankar S: Sonic hedgehog signaling inhibition provides
opportunities for targeted therapy by sulforaphane in regulating
pancreatic cancer stem cell self-renewal. PLoS One. 7:e460832012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eramo A, Lotti F, Sette G, Pilozzi E,
Biffoni M, Di Virgilio A, Conticello C, Ruco L, Peschle C and De
Maria R: Identification and expansion of the tumorigenic lung
cancer stem cell population. Cell Death Differ. 15:504–514. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Okudela K, Woo T, Mitsui H, Tajiri M,
Masuda M and Ohashi K: Expression of the potential cancer stem cell
markers, CD133, CD44, ALDH1, and β-catenin, in primary lung
adenocarcinoma-their prognostic significance. Pathol Int.
62:792–801. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kitamura H, Okudela K, Yazawa T, Sato H
and Shimoyamada H: Cancer stem cell: Implications in cancer biology
and therapy with special reference to lung cancer. Lung Cancer.
66:275–281. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mizugaki H, Sakakibara-Konishi J, Kikuchi
J, Moriya J, Hatanaka KC, Kikuchi E, Kinoshita I, Oizumi S,
Dosaka-Akita H, Matsuno Y and Nishimura M: CD133 expression: A
potential prognostic marker for non-small cell lung cancers. Int J
Clin Oncol. 19:254–259. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shimada M, Sugimoto K, Iwahashi S,
Utsunomiya T, Morine Y, Imura S and Ikemoto T: CD133 expression is
a potential prognostic indicator in intrahepatic
cholangiocarcinoma. J Gastroenterol. 45:896–902. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Heo JS, Lee MY and Han HJ: Sonic hedgehog
stimulates mouse embryonic stem cell proliferation by cooperation
of Ca2+/protein kinase C and epidermal growth factor receptor as
well as Gli1 activation. Stem Cells. 25:3069–3080. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kasper M, Schnidar H, Neill GW, Hanneder
M, Klingler S, Blaas L, Schmid C, Hauser-Kronberger C, Regl G,
Philpott MP and Aberger F: Selective modulation of Hedgehog/GLI
target gene expression by epidermal growth factor signaling in
human keratinocytes. Mol Cell Biol. 26:6283–6298. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mimeault M and Batra SK: Frequent
deregulations in the hedgehog signaling network and cross-talks
with the epidermal growth factor receptor pathway involved in
cancer progression and targeted therapies. Pharmacol Rev.
62:497–524. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kalpana Deepa, Priya D, Gayathri R and
Sakthisekaran D: Role of sulforaphane in the anti-initiating
mechanism of lung carcinogenesis in vivo by modulating the
metabolic activation and detoxification of benzo(a)pyrene. Biomed
Pharmacother. 65:9–16. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
MacDonagh L, Gray SG, Breen E, Cuffe S,
Finn SP, O'Byrne KJ and Barr MP: Lung cancer stem cells: The root
of resistance. Cancer Lett. 372:147–156. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kinugasa Y, Matsui T and Takakura N: CD44
expressed on cancer-associated fibroblasts is a functional molecule
supporting the stemness and drug resistance of malignant cancer
cells in the tumor microenvironment. Stem Cells. 32:145–156. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee CG, McCarthy S, Gruidl M, Timme C and
Yeatman TJ: MicroRNA-147 induces a mesenchymal-to-epithelial
transition (MET) and reverses EGFR inhibitor resistance. PLoS One.
9:e845972014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yao J, An Y, Wie JS, Ji ZL, Lu ZP, Wu JL,
Jiang KR, Chen P, Xu ZK and Miao Y: Cyclopamine reverts acquired
chemoresistance and down-regulates cancer stem cell markers in
pancreatic cancer cell lines. Swiss Med Wkly.
141:w132082011.PubMed/NCBI
|
|
31
|
Dean M, Fojo T and Bates S: Tumour stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sarvi S, Mackinnon AC, Avlonitis N,
Bradley M, Rintoul RC, Rassl DM, Wang W, Forbes SJ, Gregory CD and
Sethi T: CD133+ cancer stem-like cells in small cell lung cancer
are highly tumorigenic and chemoresistant but sensitive to a novel
neuropeptide antagonist. Cancer Res. 74:1554–1565. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang J, Chen J, He J, Li J, Shi J, Cho WC
and Liu X: Wnt signaling as potential therapeutic target in lung
cancer. Expert Opin Ther Targets. 20:999–1015. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kubo T, Takigawa N, Osawa M, Harada D,
Ninomiya T, Ochi N, Ichihara E, Yamane H, Tanimoto M and Kiura K:
Subpopulation of small-cell lung cancer cells expressing CD133 and
CD87 show resistance to chemotherapy. Cancer Sci. 104:78–84. 2013.
View Article : Google Scholar : PubMed/NCBI
|