Introduction
Metastasis to the regional lymph nodes is a major
factor indicating the negative prognosis of patients with cancer.
Whether or not metastasis has occurred influences whether a patient
receives postoperative therapy, including chemo- and radiotherapy,
to reduce the risk of metastatic recurrence (1). Carcinoma-induced lymphangiogenesis
within the tumor tissue is one of the causes of lymph node
metastasis and it has been revealed that stromal and cancer cells
serve a critical role in the induction of lymphangiogenesis by
secreting lymphangiogenic factors, including vascular endothelial
growth factor (VEGF) and platelet-derived growth factor (PDGF)
(2). Carcinoma-associated fibroblasts
(CAFs) are the most abundant stromal cells in the tumor
microenvironment (TME) and promote tumor growth, angiogenesis and
the malignant progression of various solid tumors (3). In addition, CAFs use the
epithelial-mesenchymal transition, a major biological process that
can stimulate cancer progression and metastasis, to regulate the
invasiveness of tumor cells (4,5).
The hepatocyte growth factor (HGF) receptor c-Met is
encoded by the proto-oncogene MET. MET exhibits tyrosine-protein
kinase activity and is overexpressed in numerous types of cancer,
such as head and neck squamous cell carcinoma (6,7).
Furthermore, the HGF/c-MET complex is widely expressed in cells of
epithelial-endothelial origin and regulates multiple cellular
processes in cancer, including cell proliferation, invasion and
angiogenesis (7). Although a
correlation between CAFs and cancer progression and metastasis has
been determined (4,5), it remains unknown whether CAFs
contribute to lymphangiogenesis in oral squamous cell carcinoma
(OSCC) via the HGF/c-Met complex. Therefore, identifying the
complex interactions between CAFs and lymphatic endothelial cells
may help to identify the underlying mechanisms of CAFs in the
lymphangiogenesis of OSCC.
CAFs and normal fibroblasts (NFs) have been
successfully isolated and characterized from OSCC as well as from
normal oral mucosal tissues (8). A
co-culture experiment in a Transwell system revealed that CAFs
significantly promote the proliferation, migration, invasion and
tube formation of human lymphatic endothelial cells (HLECs)
(8). The present study used a
chemiluminescent immunoassay (CLIA) to measure the secretion of HGF
by CAFs and to evaluate the expression of c-Met in HLECs
co-cultured with CAFs. Furthermore, the influence of HGF on the
proliferation, migration, invasion and tube formation of HLEC was
evaluated using recombinant human HGF (rhHGF). An inhibitor of
c-Met, JNJ-38877605, suppressed the proliferation, migration and
invasion, as well as the tube formation, of HLECs by downregulating
phosphatidylinositol 3-kinase (PI3K) and phosphorylated protein
kinase B (AKT) levels. Thus, the results of the current study
indicate that the c-Met/PI3K/AKT signaling pathway is one mechanism
by which CAFs regulate lymphangiogenesis.
Materials and methods
Cell culture
The present study was approved by the Medical Ethics
Committee of the West China Hospital of Stomatology of Sichuan
University (Chengdu, China). Specimens of OSCC and normal oral
mucosal tissue were obtained from 5 patients (2 males, 3 females;
mean age, 61.6 years) from November 2015 to December 2015 at the
Department of Head and Neck Oncology, West China Hospital of
Stomatology, and all patients provided informed consent prior to
inclusion in the current study. The isolation and characterization
of primary CAFs and NFs was performed according to a previously
established protocol (8). CAFs and
NFs were cultured in high-glucose Dulbecco's modified Eagle's
medium (DMEM; GE Healthcare, Chicago, IL, USA) containing 10% fetal
bovine serum (FBS; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), penicillin (100 U/ml) and streptomycin (100 µg/ml; GE
Healthcare) at 37°C in a humidified incubator with 5%
CO2. HLECs were purchased from ScienCell Research
Laboratories (Carlsbad, CA, USA) and cultured in endothelial cell
medium (ECM; ScienCell Research Laboratories) supplemented with
endothelial cell growth supplement (ScienCell Research
Laboratories), 10% FBS, penicillin (100 U/ml) and streptomycin (100
µg/ml) at 37°C in a 5% CO2 humidified atmosphere. Cell
culture duration is described in each specific experiment as
follows.
CLIA
CAFs and NFs (third passage, 2×105
cells/well) were plated in 6-well plates in high-glucose DMEM
containing 10% FBS. When cells reached 60–70% confluence, they were
incubated further at 37°C in FBS-free DMEM-high glucose for 24 h.
Culture medium were collected and centrifuged at 1,000 × g, at 4°C
for 20 min and the supernatant was evaluated using an HGF CLIA kit
(USCN Life Science, Inc., Wuhan, China) following the
manufacturer's protocol.
Cell proliferation assay
Cell Counting kit-8 (CCK-8) assays (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) were used to
evaluate the proliferation of HLECs. HLECs induced by rhHGF
(PeproTech, Inc., Rocky Hill, NJ, USA) or JNJ-38877605 (400 nM;
Selleck Chemicals, Houston, TX, USA) were seeded in 96-well plates
(2,000 cells/well, 100 µl) and incubated at 37°C for 12 or 24 h.
For rhHGF, the blank control was the culture medium without cells.
The negative control used the vehicle, deionized water for rhHGF
and DMSO for JNJ-38877605. Cells were incubated with 10 µl CCK-8
for 2 h, and optical density (OD) was, measured at 450 nm using
Varioskan Flash microplate reader (Thermo Fisher Scientific,
Inc.).
Cell migration and invasion
assays
HLEC migration and invasion assays were performed in
a 24-well Transwell system (8 µm pore size; Corning, Corning, NY,
USA). In these assays, rhHGF, CAFs or CAFs and JNJ-38877605 (400
nM) were plated in the lower chamber. HLECs (3×105 cells
in 300 µl medium) were seeded in the upper chamber coated with
Matrigel (BD Biosciences, San Jose, CA, USA) for the invasion
assay. The same protocol was used to carry out the migration assay
except without coating Matrigel in the upper chamber. Deionized
water and CAFs without JNJ-38877605 were taken as the respective
negative control in the two assays. Following incubation for 24 h,
the non-penetrating cells on the upper surface of the filters were
removed using a cotton swab. The migrated and invaded cells were
fixed in 4% paraformaldehyde for 5 min at room temperature and
stained with 0.1% crystal violet for 15 min. Following each step,
filters were washed 3 times in PBS. Three random fields were
selected and the cells on each filter were photographed using an
inverted phase contrast microscope (Olympus Corporation, Tokyo,
Japan) at a magnification of ×100.
Matrigel in vitro HLEC tube formation
assay
The Matrigel in vitro assay was used to
evaluate HLEC tube formation following rhHGF treatment or
co-culture with CAFs that were supplemented with JNJ-38877605 or
not. Matrigel was plated at the bottom of a 24-well plate or the
lower chamber of the 24-well Transwell system (0.4 µm; Corning)
following thawing overnight at 4°C and then kept at 37°C for 60
min. HLEC cells (1×105) in 500 µl ECM containing 5% FBS
and endothelial cell growth supplement were seeded in the
Matrigel-coated wells and treated with rhHGF (3, 30 ng/ml) at 37°C
for 14 h, or co-cultured with CAFs (upper chamber, 1×105
cells) supplemented with JNJ-38877605 (400 nM) at 37°C for 14 h.
Following fixation with 4% paraformaldehyde at room temperature for
30 min, images of the cells were captured using an inverted phase
contrast microscope (Olympus Corporation) at a magnification of
×100.
Western blot analysis
CAFs (2×105) or NFs in 1 ml FBS-free DMEM
supplemented with JNJ-38877605 or not were seeded in the upper
chamber of a 6-well Transwell system (0.4 µm; Corning), and
2×105 HLEC cells in 2 ml FBS-free ECM were seeded in the
lower chamber. Following 24 or 48 h incubations at 37°C, total
proteins were isolated from HLECs using RIPA lysis buffer [50 mM
Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1% sodium
deoxycholate, 0.1% SDS, containing protease and phosphatase
cocktails] (Beyotime Institute of Biotechnology, Haimen, China).
Protein concentration was determined using a bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology).
Subsequently, 20 µg proteins were separated using 8% SDS-PAGE and
electrophoretically transferred onto a nitrocellulose membrane (0.2
µm; EMD Millipore, Billerica, MA, USA). Following blocking with 5%
skim milk in Tris-buffered saline and 0.1% Tween-20 (TBST) for 1 h,
membranes were incubated with primary antibodies against c-Met
(1:2,000), phosphorylated (p)-c-Met (1:500) AKT1 (1:5,000), p-AKT1,
(1:5,000), β-tubulin (1:1,000; all from Abcam, Cambridge, UK); PI3K
(1:1,000; Tianjin Saierbio Biotechnology, Tianjin, China).
Following overnight incubation with primary antibodies diluted in
5% skim milk in TBST at 4°C, membranes were incubated with
peroxidase-conjugated goat-anti mouse (cat. no. A0216) or goat-anti
rabbit (cat. no. A0208) secondary antibodies (1:2,000; Beyotime
Institute of Biotechnology) for 1 h at room temperature. Between
each incubation, the membranes were washed 3 times for 5 min each
in TBST. Western blots were visualized using Western Lightning
Chemiluminescence Reagent (PerkinElmer, Waltham, MA, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from HLECs was isolated using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. cDNA was synthesized using a PrimeScript
RT reagent kit (Takara Bio, Inc., Otsu, Japan). qPCR was performed
using a SYBR® Premix Ex gen, Taq™ II
kit (Takara Bio, Inc.) on an ABI 7300 Real-time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The thermocycling
conditions used were as follows: 1 cycle of 95°C for 30 sec, 40
cycles of 95°C for 5 sec and 60°C for 31 sec, and tubulin was used
as a reference gene. The primers for c-Met and β-tubulin were as
follows: c-Met forward, 5′-CATGCCGACAAGTGCAGTA-3′ and reverse,
5′-TCTTGCCATCATTGTCCAAC-3′; β-tubulin forward,
5′-TTGGCCAGATCTTTAGACCAGACAAC-3′ and reverse,
5′-CCGTACCACATCCAGGACAGAATC-3′. Data were analyzed using
2−ΔΔCq method (9). Data
were analyzed and compared using the following relative
quantification method: R=2−ΔΔCq,
ΔΔCq=(Cq1-Cq2)-(Cq3-Cq4), where R represents the relative amount of
the experimental group relative to the NC group; Cq1 represents the
cycle threshold of c-Met in the experimental group, Cq2 represents
the cycle threshold of tubulin in the experimental group, Cq3
represents the cycle threshold of c-Met in the NC group and Cq4
represents the cycle threshold of tubulin in the NC group (9).
Statistical analysis
All experiments were conducted in triplicate and
representative data are presented as the mean ± standard deviation.
The comparison of treatment effects was evaluated using a
two-tailed Student's t-test or one-way ANOVA followed by
Holm-Sidak's multiple comparisons test. P<0.05 was considered to
indicate a statistically significant difference. Image pro plus 6.0
(Media Cybernetics, Rockville, MD, USA) was used to count the cells
in the migration and invasion assays as well as the tubes in the
tube formation assay. Statistical analyses were performed using
GraphPad Prism 6.0 (GraphPad Software Inc., La Jolla, CA, USA).
Results
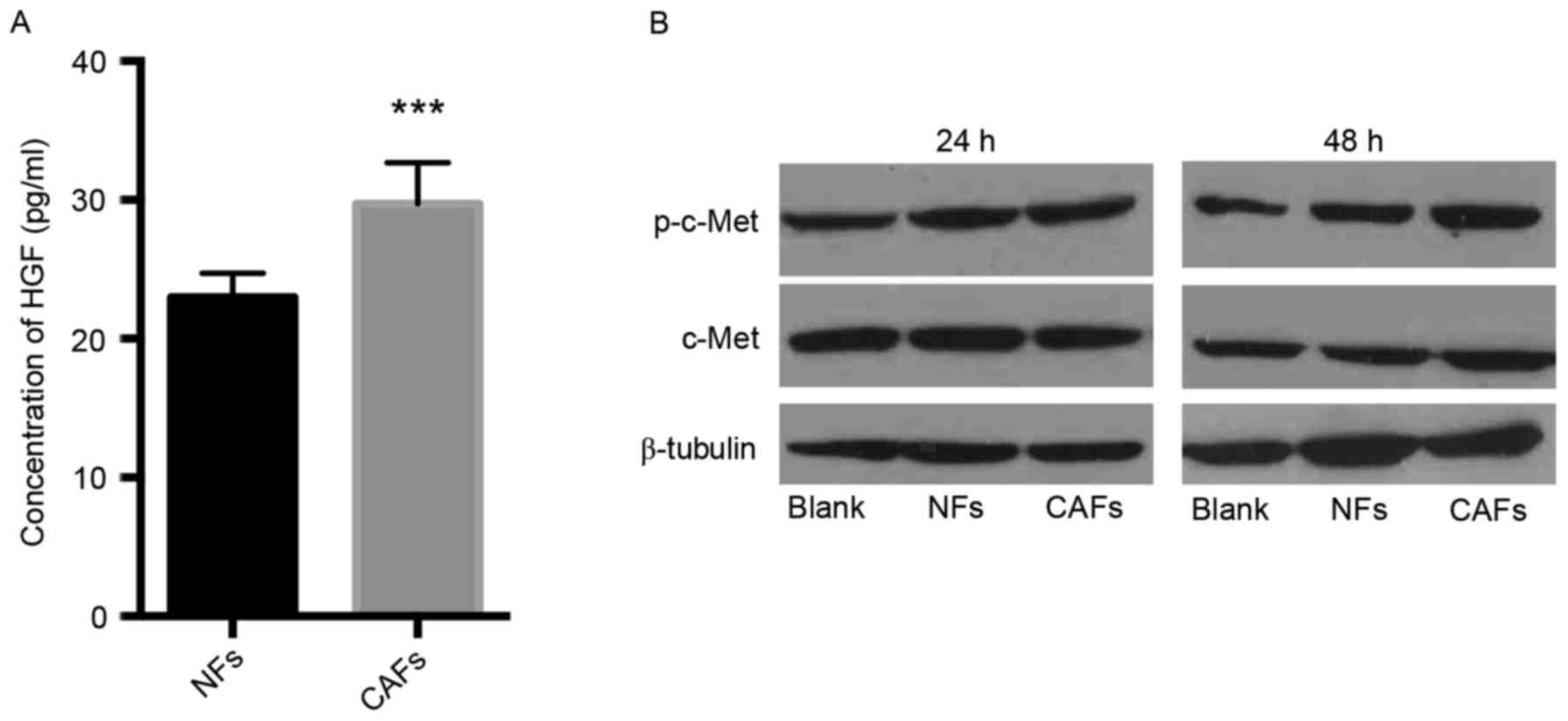
CAFs secrete higher level of HGF than
NFs
It is currently unknown whether the contribution of
CAFs to lymphangiogenesis in OSCC involves HGF/c-Met. Secreted HGF
in the supernatant of CAFs and NFs medium were examined using a
CLIA kit. Following 24 h, the secretion of HGF by CAFs (29.71±2.963
pg/ml) was significantly higher than the secretion of HGF by NFs
(22.99±1.717 pg/ml; P<0.001; Fig.
1A). These data indicate that, compared with NFs, CAFs taken
from the TME of OSCC secrete higher levels of HGF.
Overexpression of p-c-Met in HLECs
co-cultured with CAFs
To evaluate the expression of c-Met and its
phosphorylated derivative p-c-Met in HLECs co-cultured with CAFs or
NFs, western blot analysis was performed to identify the expression
of c-Met and p-c-Met following 24 and 48 h culture. Co-cultured
with CAFs for 48 h, levels of p-c-Met in HLECs were markedly higher
than with NFs (Fig. 1B). These data
indicate that substances secreted by CAFs promoted the expression
of p-c-Met of HLECs. Thus, CAFs may serve an important role in
lymphangiogenesis.
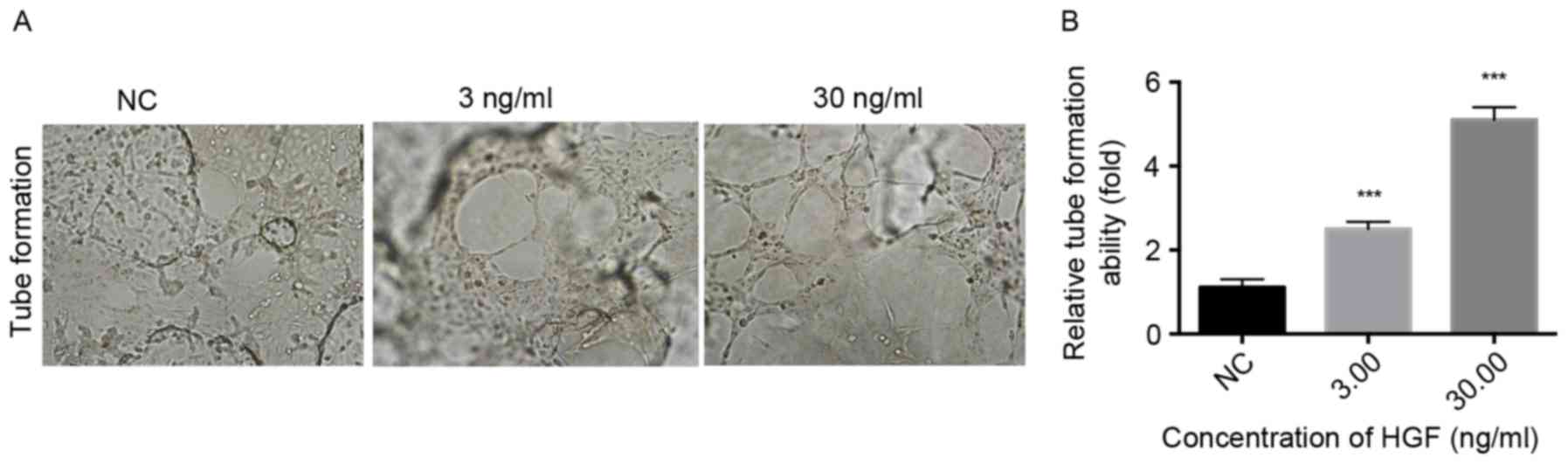
HGF promotes the proliferation,
migration, invasion and tube formation of HLECs
To determine the regulatory role of HGF on HLECs,
HLECs were treated with increasing concentrations of rhHGF and the
proliferation, migration, invasion and tube formation of HLECs was
subsequently evaluated. A significant dose-dependent difference was
observed in the proliferation of HLECs that were treated with rhHGF
(1.5625–200 ng/ml) for 12 and 24 h compared with the NC (P<0.05;
Fig. 2A). The HLECs were induced by
0, 0.3, 3.0 and 30 ng/ml rhHGF. The number of migrating and
invading cells significantly increased in a dose-dependent manner
following treatment with rhHGF (P<0.001; Fig. 2B and C), indicating that rhHGF
promotes the migration and invasion of HLECs. Furthermore,
following stimulation with 3.0 and 30 ng/ml rhHGF, HLECs exhibited
a significantly higher number of tube formation compared with the
NC group (P<0.001; Fig. 3),
indicating that rhHGF is involved in the tube formation of HLECs.
Taken together, HGF serves a critical role in modulating the
CAFs-associated proliferation, invasion, migration and tube
formation of HLECs.
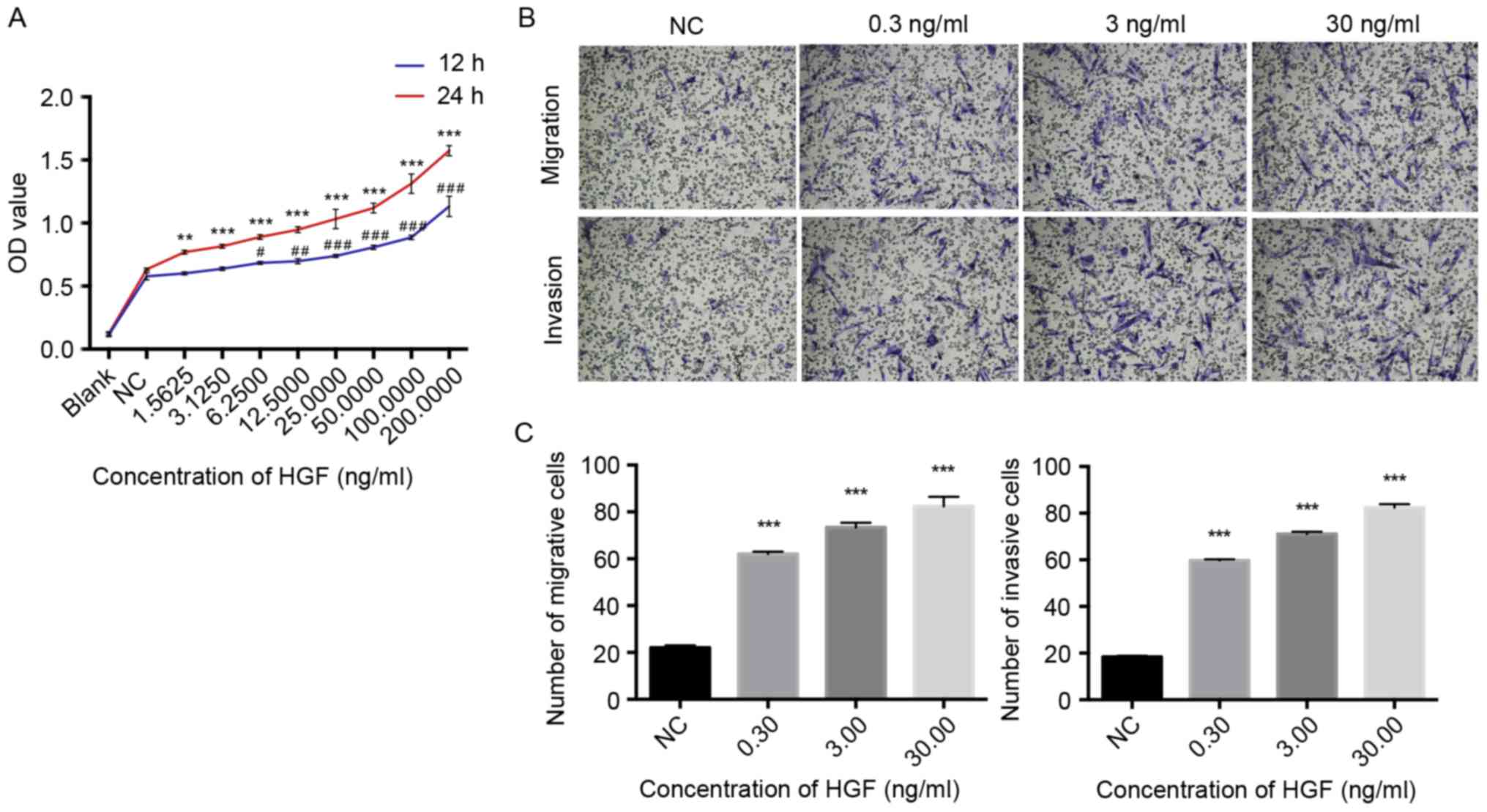 | Figure 2.rhHGF promotes the proliferation,
migration and invasion of HLECs. (A) Cell Counting kit-8 assay for
the growth curve of HLECs treated with 1.5625–200 ng/ml rhHGF for
12 or 24 h; (B) Images of migrating and invading cells and (C) the
number of migrating and invading HLECs treated with 0, 0.3, 3.0 or
30 ng/ml rhHGF for 24 h. Cells were identified using an inverted
phase contrast microscope (magnification, ×100). Results are the
representative of three independent experiments.
#P<0.05, ##P<0.01 and
###P<0.001 vs. NC after 12 h; **P<0.01 and
***P<0.001 vs. NC after 24 h. rhHGF, recombinant human
hepatocyte growth factor; HLECs, human lymphatic endothelial cells;
NC, negative control; OD, optical density; HGF, hepatocyte growth
factor. |
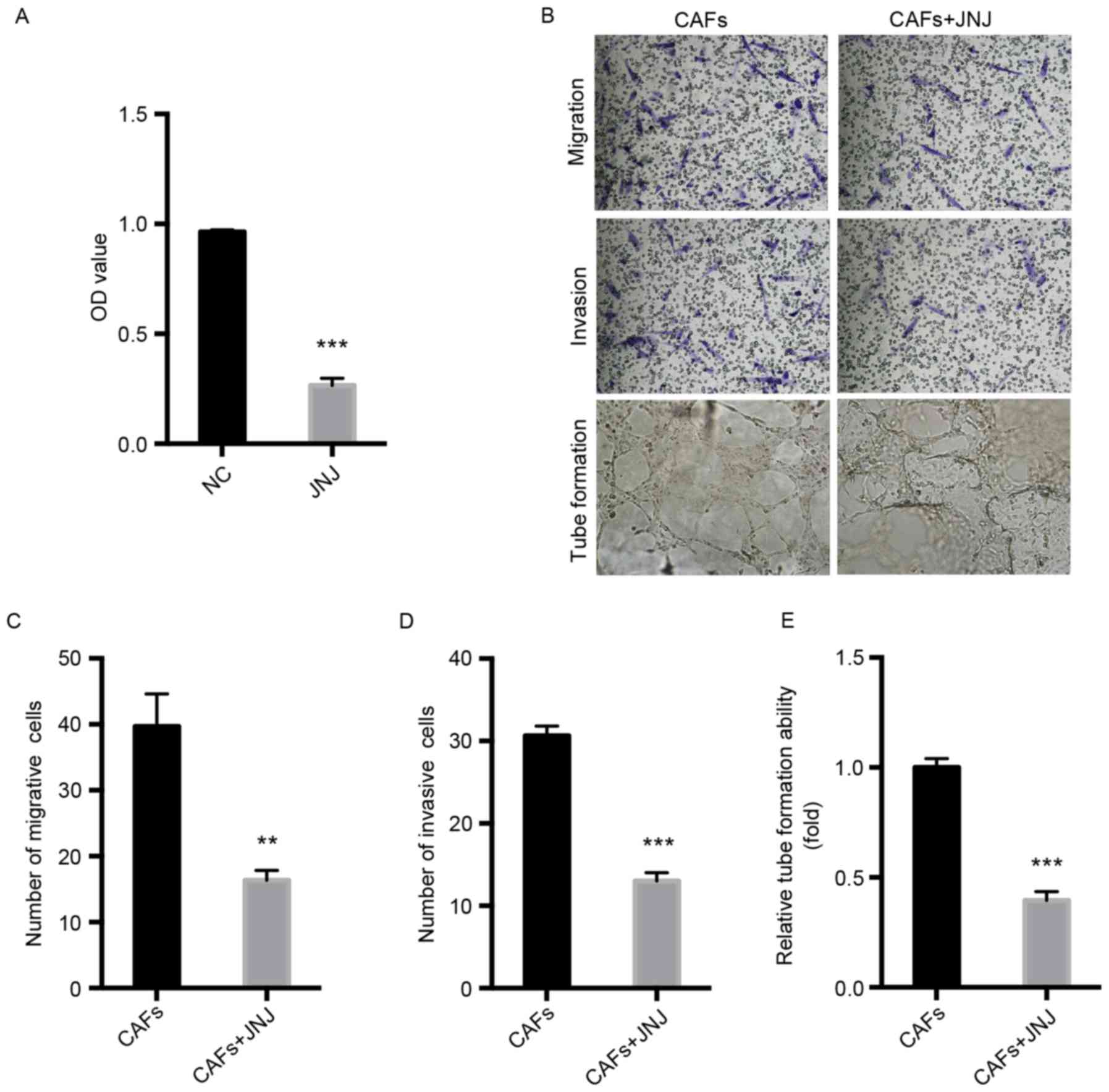
Inhibition of c-Met attenuates the
proliferation, migration, invasion and tube formation of HLECs
To know the role of c-Met in lymphangiogenesis, the
proliferation, invasion, migration and tube formation of HLECs
co-cultured with CAFs and JNJ-38877605 was investigated. The
results of the CCK-8 assay indicated that JNJ-38877605
significantly inhibited the proliferation of HLECs compared with
the NC group (P<0.001; Fig. 4A).
Furthermore, the results of the migration and invasion assays
indicated that JNJ-38877605 significantly repressed the migration
(P<0.01) and invasion (P<0.001) of HLECs compared with the NC
group (Fig. 4B-D).
JNJ-38877605-treated HLECs formed fewer tubes compared with the NC
group (P<0.001; Fig. 4B and E).
These results indicate that CAFs-modulated HGF/c-Met is crucial for
the proliferation, invasion, migration and tube formation of
HLECs.
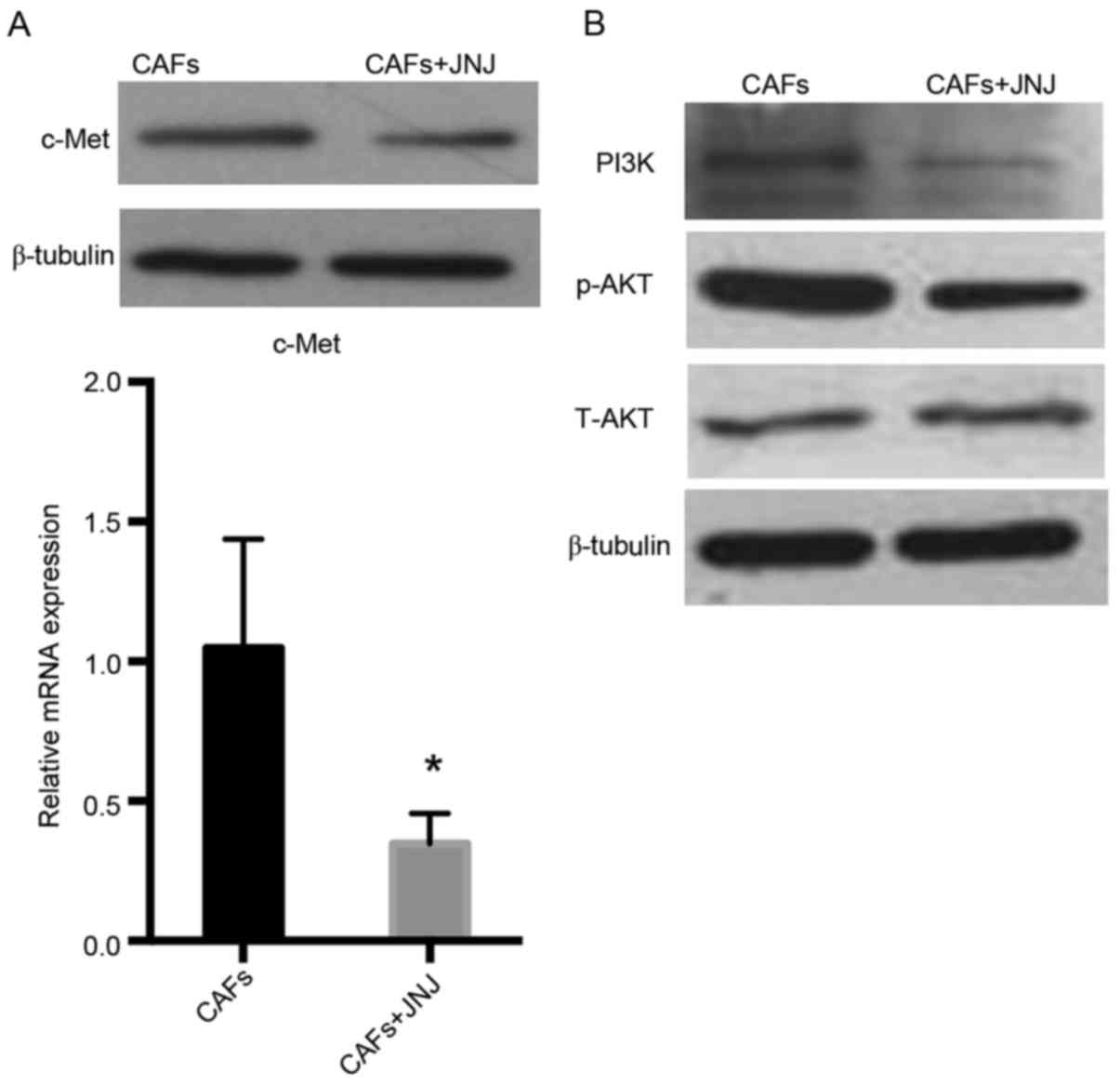
Inhibitor of c-Met downregulates the
expression of c-Met and PI3K/AKT in HLECs
To identify the molecules or proteins involved in
the signaling pathway, HLECs were co-cultured with CAFs
supplemented with or without the c-Met inhibitor, JNJ-38877605.
JNJ-38877605 reduced c-Met mRNA compared with the control group
(P<0.05) and markedly downregulated c-Met protein level
(Fig. 5A). In addition, western blot
analysis indicated that PI3K and p-AKT was also downregulated
(Fig. 5B). These data indicate that
c-Met/PI3K/AKT may regulate CAF-mediated lymphangiogenesis in the
TME of OSCC.
Discussion
Lymphangiogenesis or invasion by pre-existing
lymphatic vessels is the prerequisite of lymphatic metastasis
(10). The TME consists of various
secreted soluble cytokines or growth factors, non-cellular solid
material, ECM and stromal cells, and all of them surround the tumor
cells and support tumor hemangiogenesis, lymphangiogenesis and
metastasis (3). Stromal cells,
including fibroblasts, smooth muscle, endothelial and inflammatory
cells, interact with and modify the TME via the release of growth
factors and proteases (11–13). Stromal cells also regulate cell
proliferation, apoptosis, invasion and metastasis, as well as
hemangiogenesis and lymphangiogenesis (11–13). There
is increasing evidence that activated fibroblasts, primarily CAFs,
are vital to the TME as they promote tumor growth and
hemangiogenesis (3,14). Mantovani et al (15) reported that CAFs are recruited along
with hypoxic inducible factor-1α and promote tumor growth and
angiogenesis in the presence of hypoxia release growth factors,
including HGF, VEGF and PDGF. It remains unclear whether CAFs
contribute to lymphangiogenesis and the mechanism of CAFS in OSCC
remains unknown, although certain lymphangiogenic factors have been
identified, including fibroblast growth factor, insulin-like growth
factor, angiopoietin, VEGF, PDGF and HGF (16). Cao et al (16) successfully induced sprouting and the
growth of new lymphatic vessels in a mouse model of corneal
lymphangiogenesis via implantation of HGF. In addition, Gibot et
al (17) used a human 3D
lymphatic vascular construct to identify that HLECs, co-cultured
with fibroblasts, spontaneously formed a 3D lymphatic capillary
network without the use of any other supplemented factors. It was
further demonstrated that fibroblast-derived HGF and VEGF-C work
together in lymphangiogenesis by activating the extracellular
regulated protein kinase (ERK1/2) signaling pathway (17). Additionally, the roles of HGF/c-Met
and VEGF-C/VEGF receptor 3 (VEGFR-3) in the formation of lymphatic
vessels were identified (17).
The results of the present study revealed that HGF
levels were increased in CAFs and that c-Met was overexpressed in
HLECs co-cultured with CAFs. Furthermore, it was demonstrated that
rhHGF promotes the proliferation, migration, invasion and tube
formation of HLECs. A previous study by De Bacco et al
(18) determined that ionizing
radiation induced the overexpression and activation of the MET
oncogene that encodes c-Met via ataxia telangiectasia
mutated-nuclear factor-κB (NF-κB) signaling. This indicated that
MET promotes cell invasion and inhibits apoptosis, facilitating the
development of radioresistance. Chen et al (19) demonstrated that JNJ-38877605 decreased
the migration of retinal pigment epithelial cells, reduced the
phosphorylation of ERK and partially suppressed the activation of
protein kinase C (PKC) induced by HGF. These data implied that
c-Met, the HGF receptor, contributes to the stimulation of the
PKC/ERK signaling pathway. The present study used the co-culture
method to simulate crosstalk between CAFs and HLECs in the TME, and
the role of HGF/c-Met was subsequently investigated. The results
revealed that JNJ-38877605 downregulated the expression of c-Met at
the mRNA as well as the protein level.
PI3K, a family of enzymes involved in multiple cell
functions, can be phosphorylated and activated by several cell
membrane receptor tyrosine kinases, including Met, epidermal growth
factor receptors (EGFR), ErbB3 (a type of EGFR), platelet-derived
growth factor receptors, VEGFR and insulin-like growth factor 1
receptor (20). PI3K generated by
phosphatidylinositol (4,5)-bisphosphate, stimulates AKT via
translocation to the plasma membrane and subsequently
phosphorylates AKT. As a key effector of PI3K, AKT regulates
various cellular responses, including the cell cycle, cell
proliferation and cell survival, by phosphorylating the relevant
proteins (20–22). Yoo et al (23) reported that inhibition of PI3K/AKT
blocks sonic hedgehog-induced lymphangiogenesis in gastric cancer.
A variety of molecules are involved in the downstream signaling of
AKT, including mammalian target of rapamycin, NF-κB, glycogen
synthase kinase 3β, which regulate a varitey of cellular processes
including protein synthesis, cell survival and proliferation and
make AKT an attravtive therapuetic target for cancer (21). Finally, it was demonstrated that the
c-Met inhibitor decreased PI3K and p-AKT and attenuated the
proliferation, invasion, migration and tube formation of HLECs
co-cultured with CAFs.
In conclusion, our present study preliminarily
identified that CAFs serves a vital role on lymphangiogenesis via
the Met/PI3K/AKT signaling pathway. Future studies should focus on
evaluating the downstream mechanisms, which may help to identify
novel potential targets for therapies to inhibit metastasis.
Acknowledgements
The present study was supported by the Science and
Technology Fund of Sichuan Province of China (grant no.
2012JY0075).
References
|
1
|
Roberts TJ, Colevas AD, Hara W, Holsinger
FC, Oakley-Girvan I and Divi V: Number of positive nodes is
superior to the lymph node ratio and American Joint Committee on
Cancer N staging for the prognosis of surgically treated head and
neck squamous cell carcinomas. Cancer. 122:1388–1397. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sipos B, Klapper W, Kruse ML, Kalthoff H,
Kerjaschki D and Klöppel G: Expression of lymphangiogenic factors
and evidence of intratumoral lymphangiogenesis in pancreatic
endocrine tumors. Am J Pathol. 165:1187–1197. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alphonso A and Alahari SK: Stromal cells
and integrins: Conforming to the needs of the tumor
microenvironment. Neoplasia. 11:1264–1271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fullár A, Kovalszky I, Bitsche M, Romani
A, Schartinger VH, Sprinzl GM, Riechelmann H and Dudás J: Tumor
cell and carcinoma-associated fibroblast interaction regulates
matrix metalloproteinases and their inhibitors in oral squamous
cell carcinoma. Exp Cell Res. 318:1517–1527. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cirri P and Chiarugi P:
Cancer-associated-fibroblasts and tumour cells: A diabolic liaison
driving cancer progression. Cancer Metastasis Rev. 31:195–208.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kumar D, Kandl C, Hamilton CD, Shnayder Y,
Tsue TT, Kakarala K, Ledgerwood L, Sun XS, Huang HJ, Girod D and
Thomas SM: Mitigation of tumor-associated fibroblast-facilitated
head and neck cancer progression with anti-hepatocyte growth factor
antibody ficlatuzumab. JAMA Otolaryngol Head Neck Surg.
141:1133–1139. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sierra JR and Tsao MS: c-MET as a
potential therapeutic target and biomarker in cancer. Ther Adv Med
Oncol. 3(1 Suppl): S21–S35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen S, Gao P, Chang Z and Xuan M: Effects
of oral cancer-associated fibroblasts on the proliferation,
migration, invasion and tube formation to human lymphatic
endothelial cells. Hua Xi Kou Qiang Yi Xue Za Zhi. 33:524–528.
2015.(In Chinese). PubMed/NCBI
|
|
9
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao Y: Opinion emerging mechanisms of
tumour lymphangiogenesis and lymphatic metastasis. Nat Rev Cancer.
5:735–743. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sund M and Kalluri R: Tumor stroma derived
biomarkers in cancer. Cancer Metastasis Rev. 28:177–183. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
DeWever O and Mareel M: Role of tissue
stroma in cancer cell invasion. J Pathol. 200:429–447. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pupa SM, Ménard S, Forti S and Tagliabue
E: New insights into the role of extracellular matrix during tumor
onset and progression. J Cell Physiol. 192:259–267. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Routray S, Sunkavali A and Bari KA:
Carcinoma-associated fibroblasts, its implication in head and neck
squamous cell carcinoma: A mini review. Oral Dis. 20:246–253. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao R, Björndahl MA, Gallego MI, Chen S,
Religa P, Hansen AJ and Cao Y: Hepatocyte growth factor is a
lymphangiogenic factor with an indirect mechanism of action. Blood.
107:3531–3536. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gibot L, Galbraith T, Kloos B, Das S,
Lacroix DA, Auger FA and Skobe M: Cell-based approach for 3D
reconstruction of lymphatic capillaries in vitro reveals distinct
functions of HGF and VEGF-C in lymphangiogenesis. Biomaterials.
78:129–139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
De Bacco F, Luraghi P, Medico E, Reato G,
Girolami F, Perera T, Gabriele P, Comoglio PM and Boccaccio C:
Induction of MET by ionizing radiation and its role in
radioresistance and invasive growth of cancer. J Natl Cancer Inst.
103:645–661. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen YJ, Tsai RK, Wu WC, He MS, Kao YH and
Wu WS: Enhanced PKCδ and ERK signaling mediate cell migration of
retinal pigment epithelial cells synergistically induced by HGF and
EGF. PLoS One. 7:e449372012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Simpson DR, Mell LK and Cohen EE:
Targeting the PI3K/AKT/mTOR pathway in squamous cell carcinoma of
the head and neck. Oral Oncol. 51:291–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rogers SJ, Box C, Harrington KJ, Nutting
C, Rhys-Evans P and Eccles SA: The phosphoinositide 3-kinase
signalling pathway as a therapeutic target in squamous cell
carcinoma of the head and neck. Expert Opin Ther Targets.
9:769–790. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yoo YA, Kang MH, Lee HJ, Kim BH, Park JK,
Kim HK, Kim JS and Oh SC: Sonic hedgehog pathway promotes
metastasis and lymphangiogenesis via activation of Akt, EMT, and
MMP-9 pathway in gastric cancer. Cancer Res. 71:7061–7070. 2011.
View Article : Google Scholar : PubMed/NCBI
|