Introduction
Hepatocellular carcinoma (HCC) is one of the most
common types of tumor worldwide, particularly in less developed
countries (1,2). During 2012, ~745,500 hepatocellular
carcinoma-associated mortalities worldwide were reported, and the
mortality rate was predicted to increase in developed areas,
including the USA and Europe (1,2). It has
been demonstrated that chemotherapy resistance is one of the most
important reasons leading to HCC patient mortality (3). Therefore, investigating a novel
biomarker for HCC treatment is necessary.
Tribbles homolog 3 (TRB3) is a type of pseudokinase,
which contains a consensus serine/threonine kinase catalytic core
structure but does not exhibit kinase activity (4). Previous studies verified that TRB3 was
upregulated in hepatic carcinoma cells and was able to induce
apoptosis in hepatic carcinoma cell as a downstream target of
activating transcription factor 4 (ATF4)-CCAAT/enhancer binding
protein homologous protein (CHOP), a well-known signaling pathway,
which serves a key role in endoplasmic reticulum (ER) stress
(5,6).
Conversely, TRB3 inhibits activation of protein kinase B (AKT) by
binding with its threonine or serine activated site, which results
in the activation of the downstream signal to induce cell apoptosis
(7,8).
Furthermore, much of the research in the previous two decades has
indicated that the AKT signaling pathway is closely associated with
the occurrence and development of hepatic carcinoma (9,10).
On this basis, in the present study, it was
hypothesized that TRB3 possessed a novel function to connect ER
stress with the AKT signaling pathway and induce apoptosis and
chemotherapy resistance in HCC cells. In the present study, the
expression levels of TRB3, CHOP, AKT and phosphorylated (p)AKT were
primarily detected under normal conditions and ER stress
respectively. Subsequently, cell apoptosis and resistance to
chemotherapy induced by TRB3 knockdown were observed. Furthermore,
the associations between TRB3, the marker proteins of ER stress,
CHOP and AKT, in MHCC97H cells were investigated.
Materials and methods
Cell lines
MHCC97H and L-02 cell lines were provided by the
Central Laboratory of Shanghai First People's Hospital (Shanghai,
China). The cells were cultured in Dulbecco's modified Eagle's
medium (DMEM; Hyclone, GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in 5% CO2 at 37°C.
MHCC97H cells were treated with two drugs, thapsigargin (TG; 5
µmol/l; Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) and tunicamycin (TU; 5 µg/ml; Beijing Solarbio
Science & Technology Co., Ltd.) for 24 h, and DMSO (5 µg/ml;
Beijing Solarbio Science & Technology Co., Ltd.) was used as
the control.
Lentiviral transduction of short
hairpin (sh)RNA and transduction efficient examination
Transduction was performed using a GenePharma
Lentivirus Transduction kit (Shanghai GenePharma Co., Ltd.,
Shanghai, China) according to the manufacturer's protocol. MHCC97H
cells were plated in six-well plates at a density of
2×105 cells/well. Following 24 h at 37°C, lentiviruses
carrying four different TRB3/CHOP targeting shRNAs (TRB3-shRNA 778,
shRNA 1057, shRNA1185, shRNA1608; CHOP-shRNA 342, shRNA 807, shRNA
814, shRNA 929) and negative control (NC) shRNA (Shanghai
GenePharma Co., Ltd.) were added to the cells. The shRNA sequences
are shown in Table I. The
concentration of shRNA added was 1×108 U/ml (40 µl). The
viral titer was 1 ml/well. At the same time, polybrene was added to
each well at a final concentration of 6 µg/ml. Following incubation
at 37°C for 12 h, the viral supernatant was replaced with DMEM
supplemented with 10% FBS. The infected cells were selected using 2
mg/ml puromycin (Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
following incubation at 37°C for 48 h. Successful infection was
confirmed by the expression of green fluorescent protein using an
inverted fluorescence microscope (Leica DMI4000 B; Leica
Microsystems GmbH, Wetzlar, Germany). After 72 and 96 h at 37°C the
total RNA and total proteins were isolated using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) and SDS sample loading
buffer (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) respectively
to determine the expression levels of target genes and proteins by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blot analysis. The cells were infected every
12 h for 2 days.
 | Table I.Primer sequences for genes used in
reverse transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences for genes used in
reverse transcription-quantitative polymerase chain reaction.
| Primer | Sequence (5′-3′) |
|---|
| GAPDH |
|
|
Forward |
CATGAGAAGTATGACAACAGCCT |
|
Reverse |
AGTCCTTCCACGATACCAAAGT |
| TRB3 |
|
|
Forward |
GCTGCCAACAGTGGATTGA |
|
Reverse |
GCTTCTGCCTTTCTCCCTTCT |
| AKT |
|
|
Forward |
GGCACATTAAGATCACAGACTTCG |
|
Reverse |
TCATTGTCCTCCAGCACCTC |
| CHOP |
|
|
Forward |
TGAACGGCTCAAGCAGGAA |
|
Reverse |
AGGGTCACATCATTGGCACTA |
Cell Counting kit-8 (CCK-8) assay
A total of 1×104 MHCC97H cells in 100 µl
DMEM containing 10% FBS were seeded in a 96-well plate format in 5%
CO2 at 37°C. A total of 1 ml CCK-8 (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was mixed with 10 ml DMEM
supplemented with 10% FBS. Following transfection with TRB3-shRNA
1608 at 37°C for 48 h or treatment with cisplatin (4 µg/ml; Sangon
Biotech Co., Ltd., Shanghai, China) at 37°C for 24 h, the MHCC97H
cells were then transferred into DMEM containing 10% FBS at 37°C
for 48, 72 and 96 h, the 100 µl diluted CCK-8 was added to each
well and then absorbance at 450 nm following 1 h of cultivation at
37°C condition was determined. The absorbance at the 48 h was used
as the standard.
RT-qPCR
The total mRNA from L-02 and MHCC97H cells was
extracted using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) and reverse transcribed into cDNA using the
Reverse Transcription kit from Takara Biotechnology Co., Ltd.
(Dalian, China) according to the manufacturer's protocol. The
process of reverse transcription was as follows: 37°C for 15 min,
85°C for 5 sec, then held at 4°C hold. The forward and reverse
primers used are presented in Table
II. PCR reactions were performed using a FTC3000 Real-Time PCR
Detection System (Funglyn Biotech, Inc., Richmond Hill, ON, Canada)
with SYBR Premix Ex Taq™ II (Takara Biotechnology Co., Ltd.). The
conditions as follows: Pre-denaturation for 3 min at 95°C, followed
by 40 cycles of denaturation for 30 sec at 95°C, annealing for 30
sec at 52°C, extension for 30 sec at 72°C and a final cycle
extension at 72°C for 10 min. The level of relative gene expression
was evaluated using the ΔΔCq method (11), and expression was normalized to GAPDH.
All samples were analyzed three times.
| Table II.Sequences of shRNAs. |
Table II.
Sequences of shRNAs.
| shRNA | Sequence (5′-3′) |
|---|
| TRB3 |
|
| shRNA
778 |
GCACTGAGTATACCTGCAAGG |
| shRNA
1057 |
AGCTGTGTCGCTTTGTCTTCG |
|
shRNA1185 |
GGACCTGAGATACTCAGCTCA |
|
shRNA1608 |
GCCAACAGTGGATTGAGTTTG |
| CHOP- |
|
| shRNA
342 |
CAGCTGAGTCATTGCCTTTCT |
| shRNA
807 |
GAGCTCTGATTGACCGAATGG |
| shRNA
814 |
GATTGACCGAATGGTGAATCT |
| shRNA
929 |
GCCAATGATGTGACCCTCAAT |
Western blot analysis
The cells were lysed using SDS sample loading buffer
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 30 min on ice.
The protein concentration was determined using a BCA protein assay
kit (Beyotime Institute of Biotechnology, Haimen, China). Equal
amounts of protein extracts (30 µg) were separated
electrophoretically on 10 or 15% SDS-PAGE gels and transferred to a
polyvinylidene fluoride membrane. The membrane was blocked using 5%
w/v bovine serum albumin (Beyotime Institute of Biotechnology),
dissolved in TBS with Tween-20 (TBST; 0.1% Tween-20) at room
temperature for 1 h and incubated sequentially with primary
antibodies for 8 h at 4°C and peroxidase-conjugated secondary
antibodies for 1 h at 25°C. The antibodies used were as follows:
Rabbit TRB3 polyclonal antibody (1:2,000; cat no. A2346; ABclonal
Biotech Co., Ltd., Woburn, MA, USA), rabbit CHOP polyclonal
antibody (1:5,000; cat no. A0221; ABclonal Biotech Co., Ltd.),
rabbit AKT polyclonal antibody (1:1,000; cat no. A0001; ABclonal
Biotech Co., Ltd.), rabbit pAKT-S473 polyclonal antibody (1:1,000;
cat no. AP0098; ABclonal Biotech Co., Ltd.), rabbit GAPDH antibody
(1:10,000; cat no. AB21612; ABclonal Biotech Co., Ltd.) and
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:2,000; cat no. SC-2004; Santa Cruz Biotechnology Inc.,
Dallas, TX, USA). Following washing in TBST, immunobands were
visualized using an enhanced chemiluminescence kit (GE Healthcare,
Chicago, IL, USA) following exposure to X-ray films. The western
blot bands were evaluated using Fusion software (version 8.0;
Vilber Lourmat, Marne-la-Vallée, France) to determine the protein
expression levels and analyzed using the Quantity One program
(version 4.6.9; Bio-Rad Laboratories, Inc., Hercules, CA, USA). All
experiments were repeated three times.
Statistical analysis
Data are presented as the mean ± standard deviation.
All statistical analyses were performed using SPSS (version 19.0;
IBM Corp., Armonk, NY, USA). Cell proliferation analysis between
the experimental and control groups was performed using unpaired
t-test. The results from RT-qPCR and western blot analysis were
analyzed using one-way analysis of variance. Multiple comparisons
between the groups were performed using the Student-Newman-Keuls
method. P<0.05 was considered to indicate a statistically
significant difference.
Results
TRB3 was overexpressed in MHCC97H HCC
cells
The level of TRB3 mRNA expression was previously
demonstrated to be altered in various tumor tissues including liver
cancer tissue samples from patients and HCC cell lines (HepG2,
Hep3B, Huh-7, HLE and HCF) compared with normal cell lines
(12). The present study demonstrated
an elevated expression level of TRB3 in MHCC97H cells, ~3.8-fold
compared with the expression in the L-02 normal hepatic cell line
(Fig. 1A). It was revealed that TRB3
mRNA expression level was also altered in the MHCC97H HCC cell line
compared to L-02 cells.
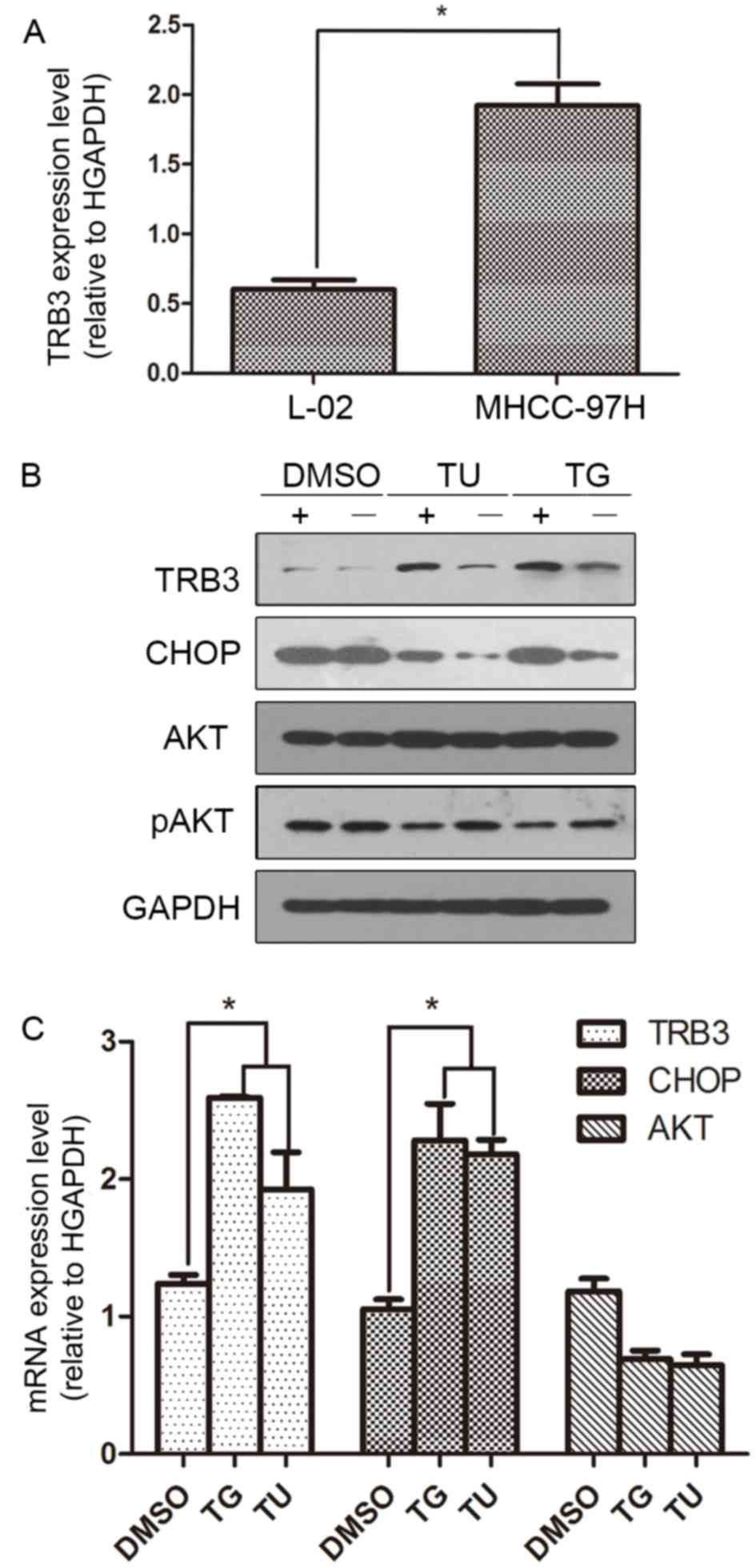 | Figure 1.Levels of TRB3 mRNA expression in HCC
cells and levels of TRB3, AKT and CHOP protein expression ER
stress. (A) TRB3 mRNA was overexpressed in MHCC97H HCC cells
compared with L-02 normal hepatic cells. (B) MHCC97H cells were
cultured with or without TG (5 µmol/l) or TU (5 µg/ml) for 24 h.
The levels of TRB3, CHOP, AKT, pAKT and GAPDH expression were
analyzed by western blotting. DMSO was used as control. (C) The
levels of mRNA expression of TRB3, CHOP and AKT in MHCC97H and L-02
cells were determined by reverse transcription-quantitative
polymerase chain rection. DMSO was used as control. *P<0.05. TG,
thapsigargin; TU, tunicmycin; DMSO, dimethylsulfoxide; TRB3,
tribbles homolog 3; AKT, protein kinase B; CHOP, CCAAT/enhancer
binding protein homologous protein; p, phosphorylated; HCC,
hepatocellular carcinoma. |
Expression of TRB3, AKT and CHOP is
induced by ER stress
To detect the changes in the expression of TRB3,
CHOP and AKT under ER stress, MHCC97H cells were treated with two
drugs, TG (5 µmol/l) and TU (5 µg/ml), which are known inducers of
ER stress. After 24 h, western blotting was performed to analyze
the levels of TRB3, CHOP, AKT and pAKT expression. Following
treatment with TU or TG, the expression of TRB3 and CHOP was
upregulated, whereas pAKT expression was decreased compared to the
DMSO control (Fig. 1B). The levels of
mRNA expression of TRB3, CHOP and AKT in these cells were also
determined by RT-qPCR. The results revealed that the expression
levels of CHOP and TRB3 mRNA were increased following treatment
with TG or TU. By contrast, the changes in the levels of AKT
expression following treatment with TG or TU were not statistically
significant (Fig. 1C).
Knockdown of TRB3 decreases apoptosis
of HCC cells and reverses resistance to chemotherapy
To investigate the function of TRB3 in apoptosis of
MHCC97H cells and resistance to chemotherapy, TRB3 was knocked down
in the present study by transduction of MHCC97H cells with specific
shRNAs. A total of four shRNA sequences targeting various regions
of TRB3 mRNA (shRNA-778, shRNA-1507, shRNA-1185 and shRNA-1608) and
a single NC shRNA sequence were used, and the knockdown efficiency
was subsequently determined. Following lentivirus-mediated
transduction for 24 h, the levels of TRB3 expression after 72 and
96 h were determined in the present study. Compared with the NC
control-transduced cells, transduction with shRNA-778 or shRNA-1608
was able to inhibit TRB3 expression at 72 and 96 h (Fig. 2A and B). In the present study, the
levels of TRB3 protein expression in the transducted cells were
also analyzed by western blotting and it was indicated that the
level of TRB3 protein expression in shRNA-1608-trasnfected cells
was markedly downregulated compared with the expression in the the
NC control-transduced cells. (Fig.
2C). Based on these results, the most efficient shRNA sequences
for inhibiting TRB3 expression were selected for further
experiments.
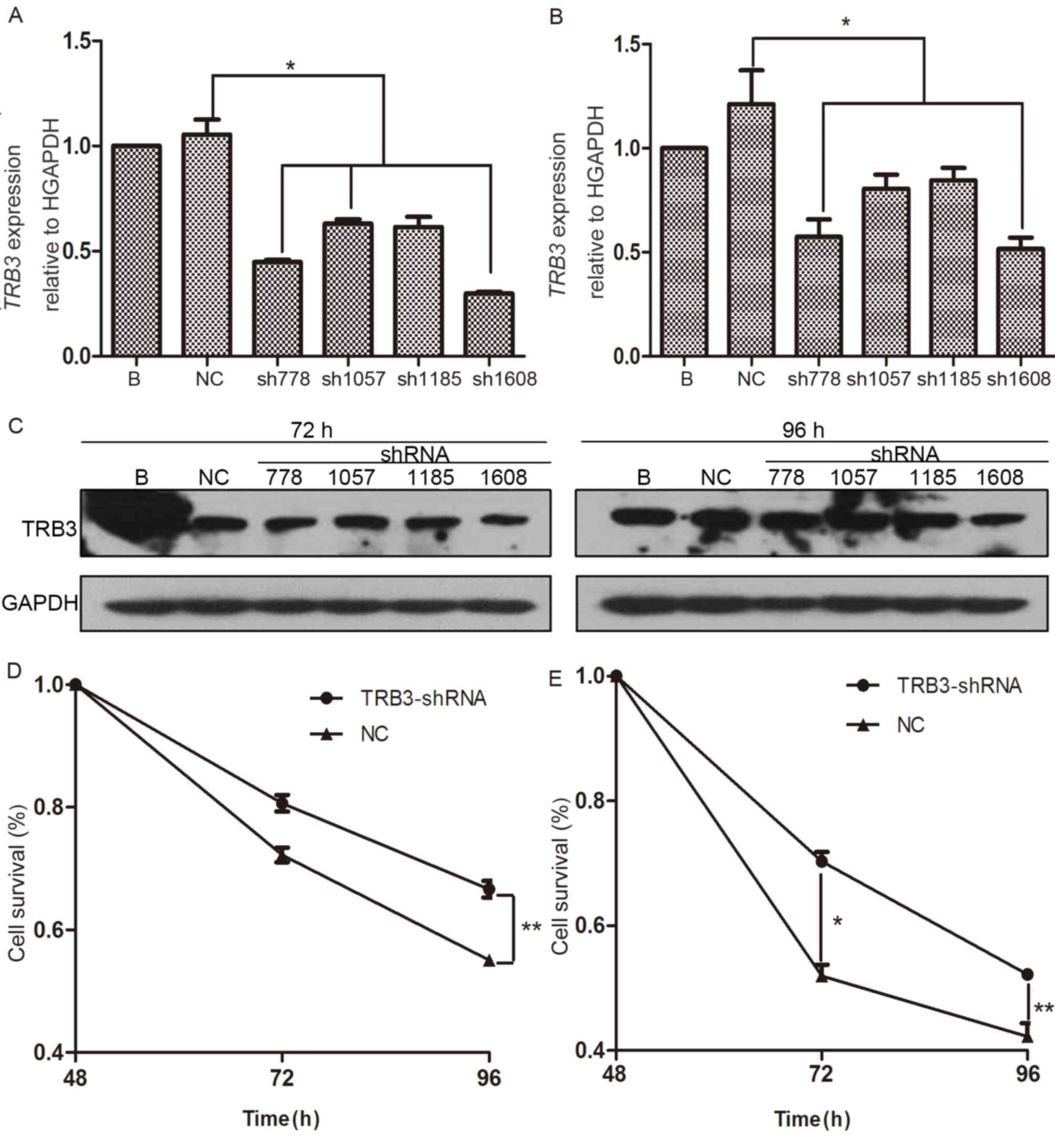 | Figure 2.Knockdown of TRB3 decreases apoptosis
of MHCC97H cells and reverses chemotherapy resistance. (A)
Following cultivation with DMEM supplemented with 10% FBS for 72
and 96 h, the levels of TRB3 mRNA expression in MHCC97H cells
transfected with lentivirus containing TRB3-sh778, TRB3-sh1057,
TRB3-sh1185, TRB3-sh1608 or NC-shRNA for 24 h were detected as
indicated by histogram. (B) Following cultivation with DMEM
supplemented with 10% FBS for 72 and 96 h, the level of TRB3 mRNA
expression in MHCC97H cells transfected with lentivirus containing
TRB3-sh778, TRB3-sh1057, TRB3-sh1185, TRB3-sh1608 or NC-shRNA for
24 h was detected as presented by histogram. (C) The levels of TRB3
protein expression in MHCC97H cells were evaluated by western blot
analysis. (D) Survival of MHCC97H cells transducted with the most
efficient lentivirus containing TRB3- and NC-shRNA for 24 h were
analyzed at 48, 72 and 96 h by CCK-8 assay. (E) TRB3- and NC-shRNA
MHCC97H cells were treated with cisplatin at 4 µg/ml for 24 h.
CCK-8 assay was used to determine cell survival at 48, 72 and 96 h.
*P<0.05, **P<0.01. TRB3, tribbles homolog 3; CCK-8, Cell
Counting kit-8; HCC, hepatocellular carcinoma; DMEM, Dulbecco's
modified Eagle's medium; FBS, fetal bovine serum; sh, short
hairpin; NC, negative control; B, blank control. |
Following shRNA transfection, CCK-8 assay was used
to determine cell survival. The 24 to 96 h cell survival curve
revealed that the number of living cells was markedly reduced when
TRB3 expression was knocked down compared with the
NC-shRNA-transduced cells (Fig. 2D).
In addition, to investigate whether the silencing of TRB3 altered
the sensitivity of MHCC97H cells to chemotherapeutic agents, TRB3-
and NC-shRNA-transduced cells were treated with 4 µg/ml cisplatin
for 24 h and cell survival was determined by CCK-8 assay. There was
a significantly higher number of surviving cells in the cells
transduced with TRB3-shRNA compared with cells transduced with
NC-shRNA following treatment with anticancer drug cisplatin for 96
h (Fig. 2E). These results suggested
that expression of TRB3 may increase apoptosis of MHCC97H cells and
reverse chemotherapy resistance.
Silencing of CHOP decreases the level
of TRB3 expression
The aforementioned ER stress activation experiment
confirmed the previous hypothesis that TRB3 expression was
upregulated under ER stress conditions and is associated with
activation of AKT. For the next part of the present study, CHOP, a
well-known molecule involved in ER stress, was knocked down using
the same methods for TRB3 knockdown (Figs. 3A-C). Following transduction of
CHOP-shRNA, the levels of TRB3 mRNA and protein expression in
MHCC97H cells were determined by RT-qPCR and western blotting,
respectively. The results indicated that TRB3 expression was
significantly downregulated compared with the control group
(Fig. 3D and E). These results do not
support the hypothesis that ER stress marker CHOP increases the
expression level of TRB3.
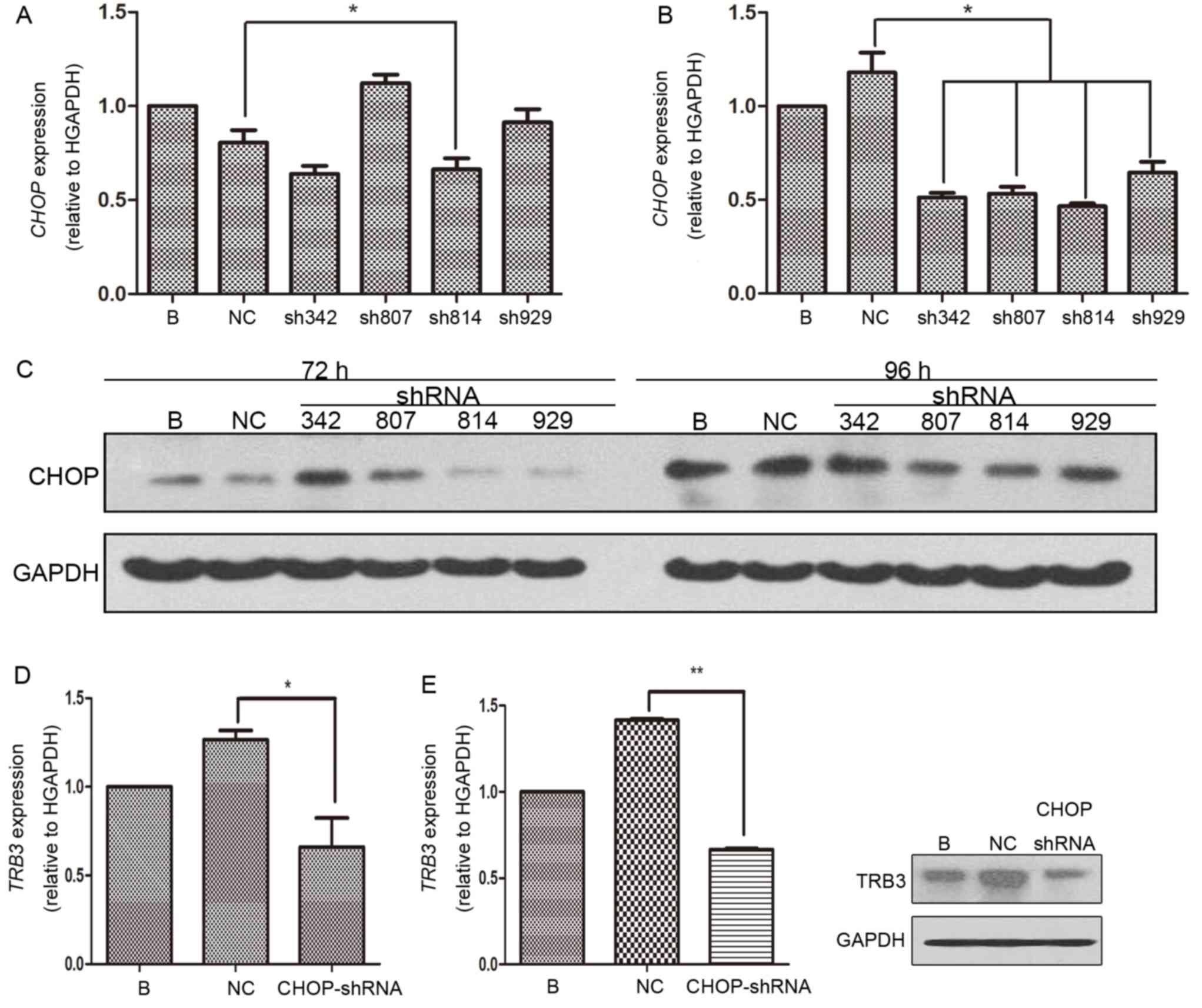 | Figure 3.Silencing of CHOP decreased the
expression level of TRB3. (A) Following cultivation with DMEM
supplemented with 10% FBS for 72 and 96 h, the level of TRB3 mRNA
expression in MHCC97H cells transfected with lentivirus containing
CHOP-sh342, CHOP-sh807, CHOP-sh814, CHOP-sh 929 or NC-shRNA for 24
h as indicated by histogram. (B) Following cultivation with DMEM
supplemented with 10% FBS for 72 and 96 h, the level of TRB3 mRNA
expression in MHCC97H cells transducted with lentivirus containing
CHOP-sh342, CHOP-sh807, CHOP-sh814, CHOP-sh 929 or NC-shRNA for 24
h was detected. The results are presented in a histogram. (C) The
levels of CHOP protein expression in MHCC97H cells were
investigated by western blot analysis. (D) The level of TRB3 mRNA
expression in MHCC97H cells transfected with the most effcient
lentivirus containing CHOP-shRNA and NC-shRNA for 24 h as presented
by histogram. (E) MHCC97H cells were analyzed by western blotting.
*P<0.05, **P<0.01. AKT, protein kinase B; TRB3, tribbles
homolog 3; CHOP, CCAAT/enhancer binding protein homologous protein;
DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum;
sh, short hairpin; NC, negative control; B, Blank control. |
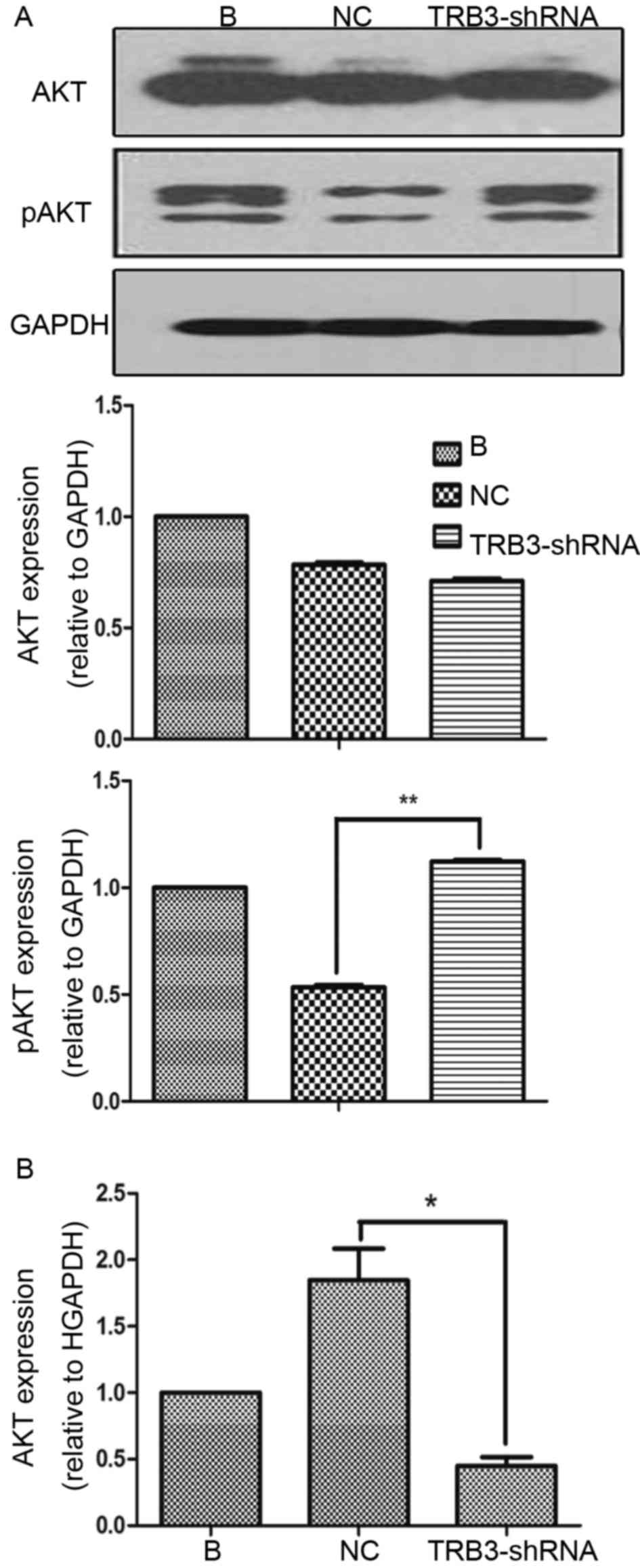
Loss of TRB3 increases the
phosphorylation of AKT in HCC cells
In the present study, it was also investigated
whether increased tumorigenic properties as a result of inhibition
of TRB3 expression, relied on increased AKT activation. Therefore,
TRB3 expression was knocked down in MHCC97H cells and activation of
AKT was investigated. It was indicated that knockdown of TRB3 did
not significantly affect the level of AKT expression but
significantly increased the level of pAKT compared with the
expression in NC shRNA-transducted cells, which suggested that TRB3
was able to prevent AKT activation (Fig.
4A). However, RT-qPCR results revealed that AKT mRNA decreased
in TRB3-knocked down MHCC97H cells (Fig.
4B). These results suggested that TRB3 exerted its inhibitory
effects on AKT activity at a post-transcriptional level.
Discussion
TRB3 has been recognized as an oncogene for various
types of tumor, including breast cancer, colorectal cancer, giloma,
oral tongue squamous cell carcinoma and HCC (12–16). A
previous study revealed that TRB3 was highly expressed in a number
of HCC cell lines under normal conditions or ER stress (12). To the best of our knowledge, the
present study identified the first time that the target gene TRB3
is overexpressed in MHCC97H HCC cells in normal conditions.
Furthermore, the levels of TRB3 protein and mRNA expression were
upregulated under ER stress.
Chemotherapeutic agents, including cisplatin, are
usually offered to unresected patients. However, the response rates
are low with rates of ~20% or lower (17). In the present study, it was revealed
that the downregulation of TRB3 resulted in increased survival of
HCC cells and that knockdown of TRB3 was able to reverse resistance
to chemotherapy in HCC cells, which suggests that poor chemotherapy
effects may be associated with TRB3 expression. To summarize, these
results implied that TRB3 may serve a key role in HCC.
Of note, the results from the present study
indicated that TRB3 was a mediator between ER stress and AKT
signaling pathways. ER stress is a protective response when the
microenvironment outside the cell has a low pH, oxygen or glucose
level. When these changes are more marked, the unfolded protein
response (UPR) then becomes a self-destructive signal for cell
death (18). Among the three UPR
downstream molecules, it is the PRKR-like endoplasmic reticulum
kinase, which is upstream of AFT4-CHOP, that serves an important
role in survival and apoptosis of HCC cells (19). A previous study in 2005 investigated
the mechanism underlying TRB3 and apoptosis of liver cells
(20). The study indicated that TRB3
is a novel target of CHOP and downregulates its own induction by
the repression of CHOP/ATF4 functions, and is involved in
CHOP-dependent cell death during ER stress (20). In addition, a previous study revealed
that TRB3 disrupts insulin signal by binding directly to Akt and
blocking PKB/AKT activation (8).
Recently, it was demonstrated that salinomycin stimulated
endoplasmic reticulum stress and mediated autophagy via the
ATF4-CHOP-TRIB3-AKT1-mammalian target of rapamycin cascades in
human non-small-cell lung cancer cells (21).
In the present study, it was revealed that CHOP and
TRB3 were co-overexpressed when under ER stress and stimulated by
thapsigargin and tunicamycin. Furthermore, the downregulation of
CHOP by shRNA inhibited the expression of TRB3. Finally, the
knockdown of TRB3 was able to increase the phosphorylation of AKT.
Conversely, the suppression of TRB3 was able to increase AKT
phosphorylation in MHCC97H cells, which suggested that TRB3 may
inhibit AKT activation. Of note, the present study demonstrated
that AKT mRNA was decreased in TRB3-knockdown MHCC97H cells. This
was in contrast to the findings of a previous study, which
suggested that the level of ATK mRNA expression may be upregulated
under TRB3 suppression (10). This
may be attributed to the specificity of the tumor type.
Consequently, the results of the present study revealed a potential
underlying mechanism between ER stress-dependent cell death and the
AKT signaling pathway of HCC cells.
In conclusion, the present study suggested a crucial
function of TRB3, which serves a key role in regulating apoptosis
and chemotherapy resistance in HCC cells. TRB3 may be a mediator
for regulating ER stress-induced tumor behaviors via modulation of
the AKT signal in HCC cells. The present study provided evidence
that TRB3 serves an important role in increasing tumorigenesis and
was a potential therapeutic target for the development of therapies
for HCC.
Acknowledgements
The present study was funded by the National Natural
Science Foundation of China (grant no. 81072007).
Glossary
Abbreviations
Abbreviations:
|
ER
|
endoplasmic reticulum
|
|
HCC
|
hepatocellular carcinoma
|
|
shRNA
|
short hairpin RNA
|
|
UPR
|
unfolded protein response
|
|
AKT
|
protein kinase B
|
|
pAKT
|
phosphorylated AKT
|
|
TG
|
thapsigargin
|
|
TU
|
tunicamycin
|
|
DMSO
|
dimethyl sulfoxide
|
|
CHOP
|
CCAAT/enhancer binding protein
homologous protein
|
|
ATF4
|
activating transcription factor 4
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 2016. View Article : Google Scholar
|
|
3
|
Mizukoshi E, Nakagawa H, Kitahara M,
Yamashita T, Arai K, Sunagozaka H, Iida N, Fushimi K and Kaneko S:
Phase I trial of multidrug resistance-associated protein 3-derived
peptide in patients with hepatocellular carcinoma. Cancer Lett.
369:242–249. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mayumi-Matsuda K, Kojima S, Suzuki H and
Sakata T: Identification of a novel kinase-like gene induced during
neuronal cell death. Biochem Biophys Res Commun. 258:260–264. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Meng X, Leyva ML, Jenny M, Gross I,
Benosman S, Fricker B, Harlepp S, Hébraud P, Boos A, Wlosik P, et
al: A ruthenium-containing organometallic compound reduces tumor
growth through induction of the endoplasmic reticulum stress gene
CHOP. Cancer Res. 69:5458–5466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu J, Lv S, Qin Y, Shu F, Xu Y, Chen J, Xu
BE, Sun X and Wu J: TRB3 interacts with CtIP and is overexpressed
in certain cancers. Biochim Biophys Acta. 1770:273–278. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bromati CR, Lellis-Santos C, Yamanaka TS,
Nogueira TC, Leonelli M, Caperuto LC, Gorjão R, Leite AR, Anhê GF
and Bordin S: UPR induces transient burst of apoptosis in islets of
early lactating rats through reduced AKT phosphorylation via
ATF4/CHOP stimulation of TRB3 expression. Am J Physiol Regul Integr
Comp Physiol. 300:R92–R100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du K, Herzig S, Kulkarni RN and Montminy
M: TRB3: A tribbles homolog that inhibits Akt/PKB activation by
insulin in liver. Science. 300:1574–1577. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meric-Bernstam F, Akcakanat A, Chen H, Do
KA, Sangai T, Adkins F, Gonzalez-Angulo AM, Rashid A, Crosby K,
Dong M, et al: PIK3CA/PTEN mutations and Akt activation as markers
of sensitivity to allosteric mTOR inhibitors. Clin Cancer Res.
18:1777–1789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Samarin J, Laketa V, Malz M, Roessler S,
Stein I, Horwitz E, Singer S, Dimou E, Cigliano A, Bissinger M, et
al: PI3K/AKT/mTOR-dependent stabilization of oncogenic far-upstream
element binding proteins in hepatocellular carcinoma cells.
Hepatology. 63:813–826. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Miyoshi N, Ishii H, Mimori K, Takatsuno Y,
Kim H, Hirose H, Sekimoto M, Doki Y and Mori M: Abnormal expression
of TRIB3 in colorectal cancer: A novel marker for prognosis. Br J
Cancer. 101:1664–1670. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wennemers M, Bussink J, Scheijen B,
Nagtegaal ID, van Laarhoven HW, Raleigh JA, Varia MA, Heuvel JJ,
Rouschop KM, Sweep FC and Span PN: Tribbles homolog 3 denotes a
poor prognosis in breast cancer and is involved in hypoxia
response. Breast Cancer Res. 13:R822011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carracedo A, Lorente M, Egia A, Blázquez
C, García S, Giroux V, Malicet C, Villuendas R, Gironella M,
González-Feria L, et al: The stress-regulated protein p8 mediates
cannabinoid-induced apoptosis of tumor cells. Cancer Cell.
9:301–312. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J, Wen HJ, Guo ZM, Zeng MS, Li MZ,
Jiang YE, He XG and Sun CZ: TRB3 overexpression due to endoplasmic
reticulum stress inhibits AKT kinase activation of tongue squamous
cell carcinoma. Oral Oncol. 47:934–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ord T, Ord D, Kõivomägi M, Juhkam K and
Ord T: Human TRB3 is upregulated in stressed cells by the induction
of translationally efficient mRNA containing a truncated 5′-UTR.
Gene. 444:24–32. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yeo W, Mok TS, Zee B, Leung TW, Lai PB,
Lau WY, Koh J, Mo FK, Yu SC, Chan AT, et al: A randomized phase III
study of doxorubicin versus cisplatin/interferon
alpha-2b/doxorubicin/fluorouracil (PIAF) combination chemotherapy
for unresectable hepatocellular carcinoma. J Natl Cancer Inst.
97:1532–1538. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang WA, Groenendyk J and Michalak M:
Endoplasmic reticulum stress associated responses in cancer.
Biochim Biophys Acta. 1843:2143–2149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mondal D, Mathur A and Chandra PK:
Tripping on TRIB3 at the junction of health, metabolic dysfunction
and cancer. Biochimie. 124:34–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ohoka N, Yoshii S, Hattori T, Onozaki K
and Hayashi H: TRB3, a novel ER stress-inducible gene, is induced
via ATF4-CHOP pathway and is involved in cell death. EMBO J.
24:1243–1255. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dim DC, Jiang F, Qiu Q, Li T, Darwin P,
Rodgers WH and Peng HQ: The usefulness of S100P, mesothelin,
fascin, prostate stem cell antigen and 14-3-3 sigma in diagnosing
pancreatic adenocarcinoma in cytological specimens obtained by
endoscopic ultrasound guided fine-needle aspiration. Diagn
Cytopathol. 42:193–199. 2014. View
Article : Google Scholar : PubMed/NCBI
|