Introduction
It has long been considered that cancer cells
utilize glycolysis for their metabolism; they enhance glycolysis
even in the presence of adequate oxygen to meet the high energy
demands arising from their high proliferative status (1). This phenomenon, called the Warburg
effect, was proposed almost one hundred years ago (1). Additionally, many studies concerning
cancer metabolism have arisen from the development of metabolome
analysis. In other words, researchers have reported the presence of
metabolic heterogeneity even in the same tumor, and metabolic
reprogramming can occur in a context-dependent manner (2–8).
Ovarian cancer is a leading cause of death
worldwide, in part because most cases are identified late, when the
cancer has reached advanced stages (9). Patients in advanced stages almost always
have metastatic lesions in the omentum (9). The omentum itself contains a high
concentration of adipocytes. The omentum is only one site of
metastasis in the peritoneal cavity; however, it is reasonable to
consider that ovarian cancer could utilize fatty acids as an energy
resource and switch to lipid metabolism. Indeed, one research group
elegantly demonstrated that co-culturing ovarian cancer cells and
adipocyte tissues resulted in the enhancement of fatty acid
metabolism in cancer cells (10).
However, it remains unclear what type of cellular and molecular
processes specifically causes the metabolic reprogramming.
We investigated the switch to lipid metabolism in
the metabolic reprogramming of ovarian cancer cells.
We first considered the involvement of epigenetics.
Using an open database, we compared the methylation status between
the primary tumor site (ovary) and the metastatic tumor site
(omentum). However, we found no evidence of the involvement of
epigenetics (at least in terms of DNA methylation).
Then, we considered the influence of suspension in
ascites on the metabolism, and we performed a suspension culture
for the in vitro model (8). We
demonstrated that culturing cells in suspension caused an increase
in the expression of enzymes related to lipid metabolism.
Additionally, we found that those cells showed decreased expression
of cancer stem cell (CSC) markers, such as CD44 and c-kit. However,
we also found that the expression of CD44 and c-kit remained
considerably higher after those cells were exposed to starvation
and IL-6 (which is thought to be secreted by mesenchymal cells in
the peritoneal cavity), which might result in increased
phosphorylation of S6, the downstream product of mammalian target
of rapamycin (mTOR). In other words, we considered that i)
suspension in ascites, ii) exposure to starvation and iii) exposure
to the microenvironment (including IL-6) harmonically changed the
metabolic status of ovarian cancer cells and CSC markers, and we
hypothesized that the combination of these effects explained why
ovarian cancer cells were likely to disseminate to the peritoneal
cavity. The microenvironment in the body is likely to be more
complex; nonetheless, we proposed one process of ovarian cancer
metastasis. We suggest that ovarian cancer may be a good model for
the investigation of metabolic reprogramming.
Materials and methods
Cell line and cell culture
The cancer cell line SKOV3 was obtained from ATCC
(USA). Cells were cultured in DMEM (cat. no. 043-30085; Wako,
Japan), supplemented with 10% fetal bovine serum (FBS; Invitrogen,
Carlsbad, CA, USA) and 100 U/ml penicillin/100 µg/ml streptomycin
(Wako), and sub-cultured via 0.25% trypsin/EDTA (Wako) detachment.
The FBS lot number was 1706567. For the starvation culture,
low-glucose DMEM without glutamine (cat. no. 044-33555; Wako) that
was not supplemented with FBS was used. All cells were grown in a
humidified atmosphere at 37°C with 5% CO2.
Adherent culture and suspension
culture
Dissociated single cells (1×105 cells/ml)
were seeded into conventional adherent plates (Nalge Nunc
International; Thermo Fisher Scientific, Waltham, MA, USA) or
ultra-low attachment plates (Corning Inc., Acton, MA, USA) and
cultured for 48 h.
GSE 85293 and GSE 2109
Data analyses were performed by Subio, Inc.,
(Kagoshima, Japan) with Subio Platform.
GSE 85293 contained 16 samples (ovary, 4; omentum,
4; ascites, 4; normal, 4). Probe sets that were not changed
(|beta-value|<0.3) compared to normal were excluded. This gave
us 153,045 probe sets. After that, the omentum probe sets that were
changed compared to the ovary samples (|beta-value|>0.3) were
extracted.
GSE 2109 contained 2158 samples, including ovary (86
cases) and omentum (38 cases). The probe sets in which the ABS call
value was A (Absent) in more than 90% of cases were excluded. This
gave us 38877 probe sets. This set was called the QC1 list. Then,
the probe sets in which the processed signal average ranged from
−0.1 to 0.1 both in omentum and ovary samples were excluded from
the QC1 list. This gave us 16649 probe sets. This set was called
the QC2 list. From the QC2 list, the genes that differed
significantly in the omentum and ovary were extracted. The criteria
were i) |fold change| ≥1.5 and ii) P-value less than 0.1 according
to the Mann-Whitney U-test.
Reagent
IL-6 was purchased from Wako. The final
concentration of IL-6 was 50 ng/ml.
Western blotting
The same amount of protein from whole cell lysates
was subjected to SDS-polyacrylamide gel (Bio-Rad, Berkeley, CA,
USA) electrophoresis and then electrotransferred onto
polyvinylidene difluoride membranes (Bio-Rad). Blotting was
performed with Trans-Blot Turbo (Bio-Rad) according to the
manufacturer's instructions. From blocking to the secondary
antibody reaction, iBind Western System (Invitrogen) was used
according to the manufacturer's instructions. The secondary
antibodies were detected using Immobilon Western Chemiluminescent
HRP Substrate (EMD Millipore, Billerica, MA, USA) according to the
manufacturer's instructions.
Antibodies
Anti-PGC1α antibody, anti-ACOX1 antibody, anti-CPT1A
antibody, anti-CD44 antibody and anti-c-kit antibody were purchased
from Abcam (cat. no. ab54481, ab184032, ab128568, ab51037 and
ab32363, respectively). Anti-EHHADH antibody and anti-ACSS2
antibody were purchased from SantaCruz (cat. no. SC-393123 and
SC-398559, respectively). Anti-p-HSL (Ser 660) antibody,
anti-p-AMPK (Thr172) antibody and anti-S6 antibody were purchased
from Cell Signaling Technology (cat. no. 4126, 2535 and 2217,
respectively). Anti-LDHA antibody was purchased from Proteintech
(cat. no. 19987-1-AP). Anti-α-tubulin antibody was purchased from
Millipore (cat. no. CP06).
Results
DNA methylation might not be involved
in metabolic reprogramming during metastasis
It has been noted that epigenetics are involved in
the metabolic shift toward lipid metabolism (11). Thus, we speculated that epigenetic
regulation was also related to metabolic reprogramming in ovarian
cancer between the primary site (ovary) and the metastatic site
(omentum). GSE 85293 was the dataset of DNA methylation status from
sites including the ovary and omentum. GSE 2109 was the dataset of
gene expressions (microarray or RNA-sequencing) from many sites,
including the ovary and omentum. Using these datasets, we
investigated and compared the methylation status and gene
expression patterns, focusing on the PPAR signaling pathway, a
representative lipid metabolism pathway from the Kyoto Encyclopedia
of Genes and Genomes (KEGG). As shown in Table I, we could not find a correlation
between DNA methylation and gene expression, although the
investigation was limited because the datasets used were from
different patients and studies.
 | Table I.Methylation and expression status
(omentum/ovary) of the genes related to lipid metabolism. |
Table I.
Methylation and expression status
(omentum/ovary) of the genes related to lipid metabolism.
| Genes | Methylation (GSE
85293) | Expression (GSE
2109) |
|---|
| FATP | ↑ | – |
| ME1 | ↑ | – |
| LPL | – | ↑ |
| CD36 | – | ↑ |
| ACS | ↓ | – |
| EHHADH | ↑ | – |
| LCAD | ↑ | – |
| ACOX1 | ↑ | – |
| Perlipin | – | ↑ |
| FABP4 | – | ↑ |
| ADIPO | – | ↑ |
| SORBS1 | – | ↑ |
| MMP-1 | – | ↑ |
Cells cultured in suspension show
enhanced lipid metabolism and relatively less sensitivity to
starvation compared with cells cultured in adherent plates
We could not obtain evidence that epigenetics (at
least DNA methylation) were involved in metabolic reprogramming in
ovarian cancer. We speculated that a more direct phenomenon was
related to the metabolic shift toward lipid metabolism; in other
words, detachment from the primary site and suspension in ascites
itself affected metabolism (through morphological changes, for
instance). Indeed, we have already demonstrated that cancer cells
cultured in suspension are more likely to show an activated TCA
cycle than cells cultured in adherent plates (8). In the present study, we focused on lipid
metabolism. As shown in Fig. 1A,
cells cultured in suspension showed increased expression of PGC1α,
a sensor for lipid metabolism, compared with cells cultured in
adherent plates (12). Additionally,
the expression of some types of enzymes related to fatty acid
metabolism was also increased, although the level of increase
depended on the enzyme. While the expressions of these enzymes were
increased in cells cultured in adherent plates when they were
exposed to starvation, it seemed that cells cultured in suspension
showed less sensitivity to starvation. This could be interpreted to
indicate that cells in suspension already utilize fatty acid
metabolism and maintain a relative advantage against starvation.
Conversely, we considered the possibility that the phosphorylation
of AMPK, a sensor for glycolysis, was decreased in cells cultured
in suspension compared with cells cultured in adherent plates, as
shown in Fig. 1B (13). The expression of LDHA, the indicator
of glycolysis, was thought to be unchanged between the different
cell culture statuses. Taken with the result from Fig. 1A, this finding suggests that cells
cultured in suspension showed a metabolic shift from glycolysis
toward lipid metabolism. Although the phosphorylation of AMPK was
increased in cells cultured in suspension and exposed to
starvation, as shown in Fig. 1B, it
is possible that these cells were preparing for impending
nourishment.
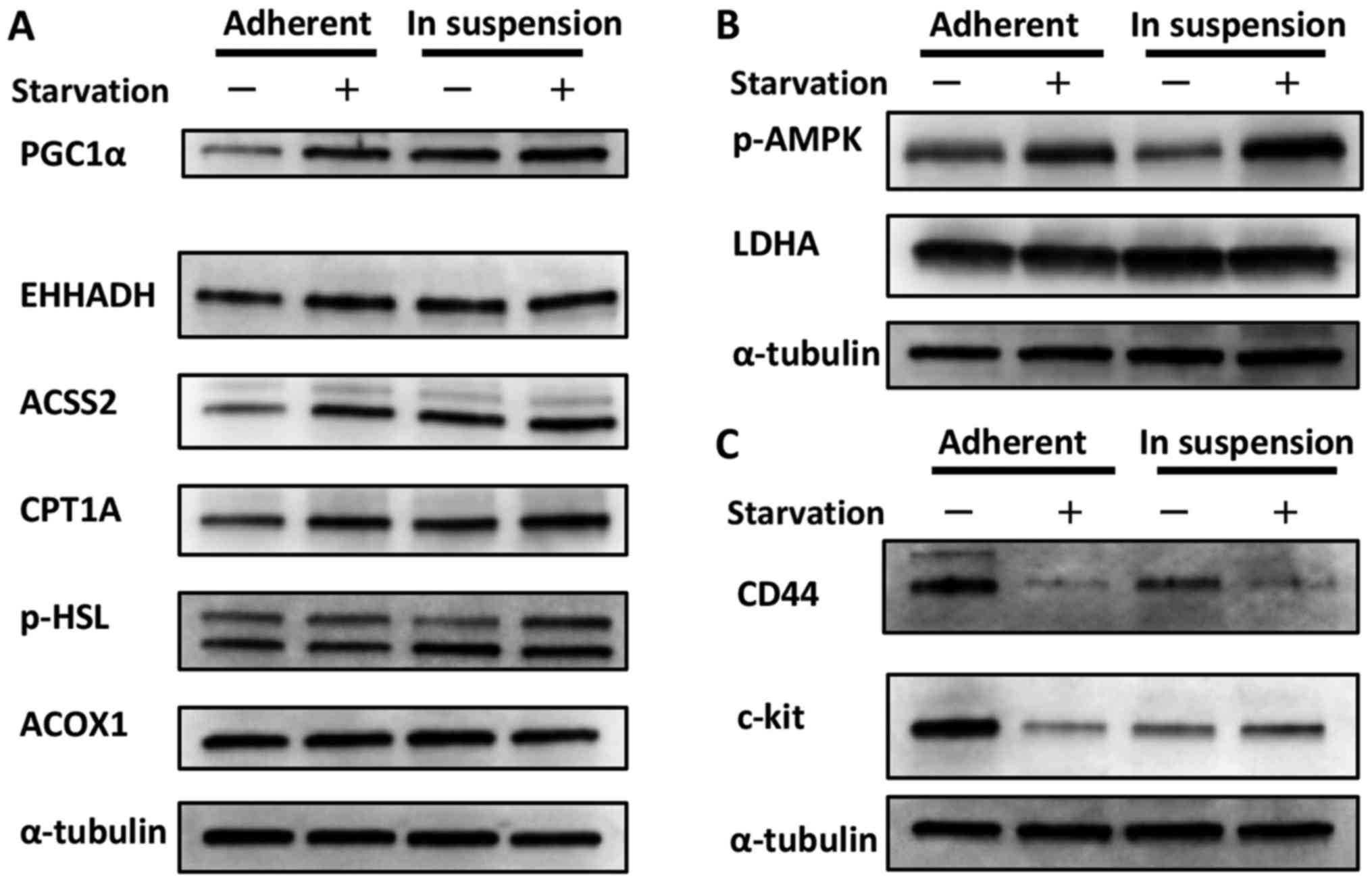 | Figure 1.Change in the metabolic status and CSC
marker expression of cells in adherent plates vs. suspension. (A)
Change in lipid metabolism. Cells cultured in suspension showed
increased expression of PGC1α, which is thought to be a sensor for
lipid metabolism, compared with cells cultured in adherent plates.
Additionally, the expression of some types of enzymes related to
fatty acid metabolism were also increased, although the level of
increase depended on the enzyme. The expressions of these enzymes
were increased in cells cultured in adherent plates when they were
exposed to starvation; however, the cells cultured in suspension
showed less sensitivity to starvation. This could be interpreted to
indicate that cells in suspension already utilize fatty acid
metabolism and maintain a relative advantage against starvation.
(B) Change in glycolysis. The phosphorylation of AMPK, a marker for
glycolysis, was decreased in cells cultured in suspension compared
with cells cultured in adherent plates. The expression of LDHA, the
indicator for glycolysis, was nearly unchanged between the two
groups. Although the phosphorylation of AMPK was increased in cells
cultured in suspension and exposed to starvation, it is possible
that these cells were preparing for upcoming nourishment. (C)
Change in CSC marker expression. Cells cultured in suspension
showed decreased expression of CD44 and c-kit compared with cells
cultured in adherent plates. Interestingly, the response during
starvation differed between the two culture groups. While the
expression of CD44 was decreased during starvation in cells
cultured in both suspension and adherent plates, the expression of
c-kit increased in cells cultured in suspension but decreased in
cells cultured in adherent plates. |
The expression of CSC markers changes
according to the cell status
At present, it is well-documented that CSCs are
related to metastasis (14). To add
to this knowledge, we investigated how CSC markers changed
according to the cell status. We considered CD44 and c-kit as CSC
markers for ovarian serous carcinoma, as previously described
(14). As shown in Fig. 1C, cells cultured in suspension showed
decreased expression of CD44 and c-kit compared with cells cultured
in adherent plates. Interestingly, the response during starvation
differed between the two culture environments. While the expression
of CD44 was decreased during starvation in both cells cultured in
suspension and those cultured in adherent plates, the expression of
c-kit increased in cells cultured in suspension but decreased in
cells cultured in adherent plates. This phenomenon was considered
advantageous for metastasis overall, as discussed below.
Cells cultured in suspension show
balanced high expression of CD44 and c-kit and possible mTOR
pathway activation after exposure to IL-6
Above, we describe an in vitro model that
mimics the condition of being detached from the primary site and
suspended in ascites. Then, we performed an experiment that
mimicked the status of ‘cells suspended in ascites and approaching
the omentum’. We focused on IL-6, which is thought to be secreted
from the omentum or other mesenchymal cells and is known to have
some impact on cancer (stem) cells (9). In other words, we investigated the
behavior of cancer cells in the presence or absence of IL-6. As
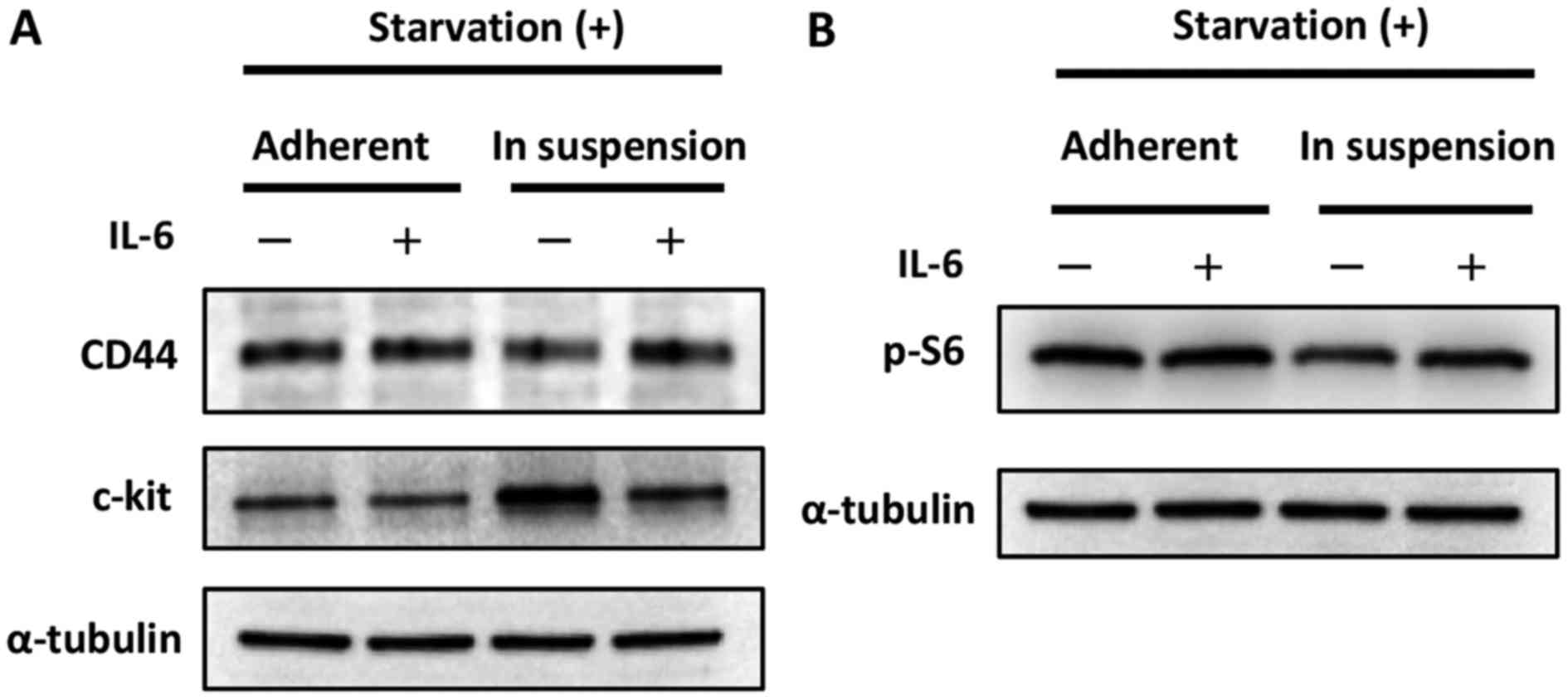
shown in Fig. 2A, the expression of
CD44 was increased in cells cultured in suspension when exposed to
IL-6, while the expression of c-kit was decreased. These results
seem paradoxical; however, they could be interpreted to indicate
that the cells expressed CD44 and c-kit in relatively high but
balanced manner. Indeed, the phosphorylation of S6, the indicator
of mTOR pathway activation, was increased in cells cultured in
suspension when exposed to IL-6, as shown in Fig. 2C.
Discussion
Here, we proposed an explanation of why ovarian
cancer cells are likely to disseminate to the peritoneal cavity,
focusing on metabolic reprogramming and CSCs.
It is considered that cancer cells utilize
glycolysis for metabolism even the presence of adequate oxygen to
satisfy the high energy demands that arise from their high rate of
proliferation. This phenomenon (aerobic glycolysis) is called the
Warburg effect, and it was first documented in the 1920s (1). However, more precise and specific
investigations of cancer cell metabolism have been performed since
the development of metabolome analysis. Metabolic reprogramming is
among these newly investigated processes, and the need to rethink
the Warburg effect is widely recognized (2–8).
Meanwhile, ovarian cancer is known to frequently
metastasize to the omentum (10). The
omentum itself contains a high concentration of adipocytes, and
ovarian cancer is thought to be a good research model for metabolic
reprogramming (especially for lipid metabolism). Indeed, a previous
report elegantly demonstrated that co-culturing ovarian cancer
cells and adipocyte tissues resulted in the enhancement of fatty
acid metabolism in ovarian cancer cells (10). However, an understanding of what
specific cellular and molecular processes cause the metabolic
reprogramming remains elusive. Collectively, we investigated the
switch in the metabolic reprogramming of ovarian cancer cells to
lipid metabolism.
While an unborn baby utilizes the glucose in blood
for its nutrition before birth, it begins to utilize lipid
metabolism during breast-feeding (i.e., upon exposure to fatty
acids) (11). Epigenetics is known to
be involved in the drastic shift (11). With this knowledge, we speculated that
epigenetics was also in involved in the metabolic reprogramming
during ovarian cancer. Using an open database, we compared the
methylation status between the primary tumor site (the ovary) and
the metastatic tumor site (the omentum). However, we did not obtain
any evidence of the involvement of epigenetics (at least in terms
of DNA methylation).
We then considered the influence of suspension in
ascites on metabolism, and we performed a suspension culture for
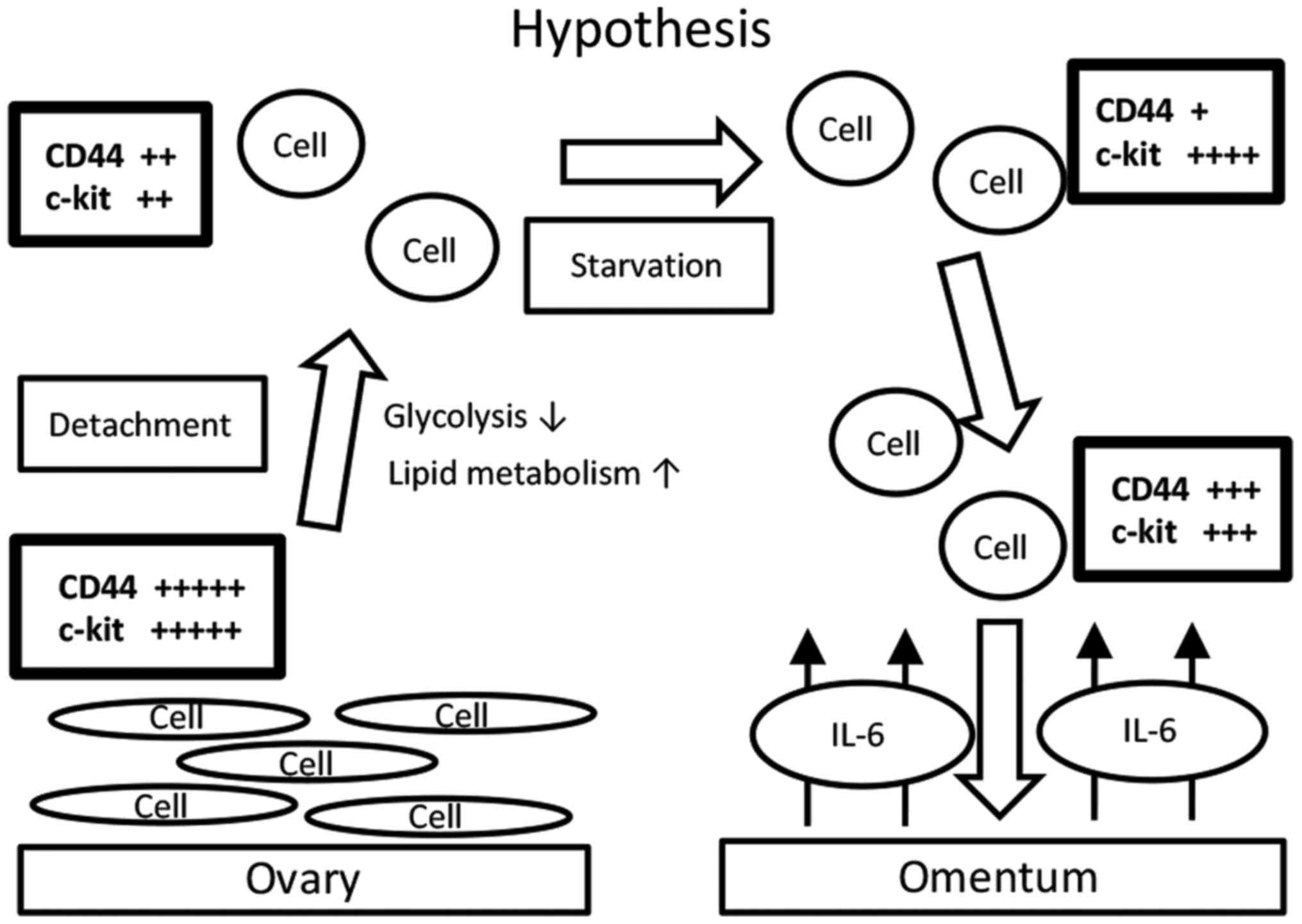
the in vitro model. A hypothetic schema of our results is
shown in Fig. 3. Ovarian cancer cells
that are detached from the primary site (the ovary) reprogram their
metabolism from glycolysis to lipid metabolism. In the process, the
expression levels of CD44 and c-kit decrease. During starvation,
the expression level of CD44 decreases and that of c-kit increases.
Upon exposure to IL-6, the expression level of CD44 further
increase, but that of c-kit decreases. These changing patterns are
believed to maintain an overall ‘CSC-like status’. Indeed, the mTOR
pathway is reactivated accordingly. We propose that this is why
ovarian cancer cells often metastasize to the omentum. Indeed, a
previous report showed that CD44-positive and c-kit-positive cells
isolated from the ascites of ovarian cancer patients presented
activation of lipid metabolism (14).
In the present study, we demonstrated that the reverse could also
be true; that is, that the expression of CD44 and c-kit in ovarian
cancer cells that are detached from the primary site and suspended
seems to change significantly in a balanced manner as the cells
approach the omentum. We do not know why detachment itself causes
metabolic changes; however, cell polarity and focal adhesion might
be responsible.
Although we only used one cell line and performed an
in vitro experiment, we proposed a possible explanation for
why ovarian cancer cells are likely to metastasize to the
peritoneal cavity, especially to the omentum.
It is quite challenging to determine whether CSCs
reprogram their own metabolism or the properties that could
contribute to metabolic reprogramming are CSCs, and further
investigations into this matter are expected. In that context,
ovarian cancer is thought to be a good model for the investigation
of metabolic reprogramming.
References
|
1
|
Dang CV: Rethinking the warburg effect
with Myc micromanaging glutamine metabolism. Cancer Res.
70:859–862. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Altieri DC: Mitochondria on the move:
Emerging paradigms of organelle trafficking in tumour plasticity
and metastasis. Br J Cancer. 117:301–305. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cliff TS and Dalton S: Metabolic switching
and cell fate decisions: Implications for pluripotency,
reprogramming and development. Curr Opin Genet Dev. 46:44–49. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: Metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fidoamore A, Cristiano L, Laezza C, Galzio
R, Benedetti E, Cinque B, Cinque B, Antonosante A, d'Angelo M,
Castelli V, et al: Energy metabolism in glioblastoma stem cells:
PPARα a metabolic adaptor to intratumoral microenvironment.
Oncotarget. 2017.
|
|
6
|
Kawada K, Toda K and Sakai Y: Targeting
metabolic reprogramming in KRAS-driven cancers. Int J Clin Oncol.
22:651–659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li X, Han G, Li X, Kan Q, Fan Z, Li Y, Ji
Y, Zhao J, Zhang M, Grigalavicius M, et al: Mitochondrial pyruvate
carrier function determines cell stemness and metabolic
reprogramming in cancer cells. Oncotarget. 8:46363–46380.
2017.PubMed/NCBI
|
|
8
|
Sato M, Kawana K, Adachi K, Fujimoto A,
Yoshida M, Nakamura H, Nishida H, Inoue T, Taguchi A, Takahashi J,
et al: Spheroid cancer stem cells display reprogrammed metabolism
and obtain energy by actively running the tricarboxylic acid (TCA)
cycle. Oncotarget. 7:33297–33305. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yeung TL, Leung CS, Yip KP, Au Yeung CL,
Wong ST and Mok SC: Cellular and molecular processes in ovarian
cancer metastasis. A review in the theme: Cell and molecular
processes in cancer metastasis. Am J Physiol Cell Physiol.
309:C444–C456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nieman KM, Kenny HA, Penicka CV, Ladanyi
A, Buell-Gutbrod R, Zillhardt MR, Romero IL, Carey MS, Mills GB,
Hotamisligil GS, et al: Adipocytes promote ovarian cancer
metastasis and provide energy for rapid tumor growth. Nat Med.
17:1498–1503. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ehara T, Kamei Y, Yuan X, Takahashi M,
Kanai S, Tamura E, Tsujimoto K, Tamiya T, Nakagawa Y, Shimano H, et
al: Ligand-activated PPARα-dependent DNA demethylation regulates
the fatty acid β-oxidation genes in the postnatal liver. Diabetes.
64:775–784. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davidson B, Hadar R, Stavnes HT, Trope CG
and Reich R: Expression of the peroxisome proliferator-activated
receptors-alpha, -beta and -gamma in ovarian carcinoma effusions is
associated with poor chemoresponse and shorter survival. Hum
Pathol. 40:705–713. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang CS, Hawley SA, Zong Y, Li M, Wang Z,
Gray A, Ma T, Cui J, Feng JW, Zhu M, et al:
Fructose-1,6-bisphosphate and aldolase mediate glucose sensing by
AMPK. Nature. 548:112–116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pastò A, Bellio C, Pilotto G, Ciminale V,
Silic-Benussi M, Guzzo G, Rasola A, Frasson C, Nardo G, Zulato E,
et al: Cancer stem cells from epithelial ovarian cancer patients
privilege oxidative phosphorylation and resist glucose deprivation.
Oncotarget. 5:4305–4319. 2014. View Article : Google Scholar : PubMed/NCBI
|