Introduction
Prostate cancer is the second most frequently
diagnosed cancer in men worldwide, and with an increasing
incidence. It is also the fifth leading cause of cancer-related
death worldwide, and >250,000 men die of prostate cancer each
year (1,2). In prostate cancer patients, bone is the
most common site of distant metastasis, which is the predominant
cause of morbidity and mortality. In addition, bone destruction can
increase the risk of death by >20% in patients with bone
metastasis (3,4). However, at present, few drugs are able
to delay the skeletal events (such as bone destruction or pain
caused by bone metastasis), and the available treatments for bone
pain are unsatisfactory (5).
Therefore, the molecular mechanisms of bone metastasis from
prostate cancer must be further explored, and more efficacious
therapeutic targets are urgently required.
Nucleus accumbens-associated protein 1 (NAC1)
belongs to a family of proteins with the Pox virus and zinc finger
(POZ; also known as BTB) domain, a versatile protein-protein
interaction motif in the N-terminus that mediates either homodimer
or heterodimer formation (6–8). Moreover, NAC1 is a transcriptional
co-factor that participates in self-renewal and maintenance of
pluripotency in embryonic stem cells (9,10). Recent
studies have revealed that NAC1 is markedly up-regulated in several
types of human carcinomas, and is involved in the proliferation,
autophagy, senescence, chemotherapy resistance, cytokinesis,
migration and motility of cancer cells (11–16). An
abundance of NAC1 protein seems to correlate with poorer prognosis
in certain carcinomas in humans (17). However, the explicit mechanism of NAC1
in prostate cancer has not yet been reported. Given the significant
and diverse roles of NAC1 in cancer biology, the purpose of the
present study was to investigate the role of NAC1 in the
development of prostate cancer and bone metastasis.
Materials and methods
Patients and sampling
The protocol utilized in this study was approved by
the Central South University Institutional Review Board. For
western blot analysis, eight pairs of prostate cancer and adjacent
normal tissue specimens were collected from the patients who
underwent surgery at the Third Xiangya Hospital of the Central
South University in 2016. In addition, 84 paraffin-embedded samples
of prostate cancer and 84 specimens of adjacent non-cancerous
prostate tissue were collected between 2014 and 2016 for use in
immunohistochemical assays. All tumors were histologically and
clinically diagnosed by the Cancer Center of Central South
University. For the use of these clinical materials, prior patient
consent and approval from the institutional research ethics
committee (approval nο: 2016-S064) were obtained.
Immunohistochemistry
Immunohistochemistry was performed on deparaffinized
sections using a NAC1 antibody (Abcam, Cambridge, MA, USA) at a
dilution of 1:100 and an UltraSensitive™ S-P hypersensitivity kit
(Fuzhou Maixin Biotech Co., Ltd., China). Following antigen
retrieval in a sodium citrate buffer, sections were incubated
overnight at 4°C with the antibody. The slides for all samples were
evaluated with a light microscope by two researchers. The antibody
staining intensity was then analyzed using the H-SCORE system with
modifications, as follows: H-SCORE=ΣPi(i), where i is the intensity
of staining (0=undetectable, 1=weakly positive, 2=moderately
positive, and 3=intensely positive).
Cell lines and culture conditions
The prostate cancer cell lines PC-3, DU-145 and
LNCaP were kindly provided by the Cancer Center of Sun Yat-sen
University, and RAW264.7 was kindly provided by the Central
Laboratory of Central South University. Cells were cultured in
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
or DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal calf serum (Biological Industries, Cromwell, CT, USA) and
100 units/ml penicillin-streptomycin (Gibco; Thermo Fisher
Scientific, Inc.).
siRNA knockdown of NAC1 gene
expression
Two small interfering RNAs (siRNAs) targeting NAC-1
were designed, with the sequences 5′-UGAUGUACACGUUGGUGCCUGUCACCA-3′
and 5′-UGUAGCAGAAGCUGAGGAUCUGCUG-3′ (16). The NAC-1 siRNAs and negative control
siRNA (luciferase siRNA) were synthesized by Shanghai GenePharma
Co., Ltd. (Shanghai, China). Cells were seeded into 6-well plates
or 96-well plates and transfected with siRNAs using siRNA-Mate
(Shanghai GenePharma Co., Ltd.). At 24 h after transfection, cells
were harvested and prepared for assays.
Western blot analysis
Cells or tissues were lysed in RIPA Lysis Buffer
supplemented with a cocktail of protease inhibitors (Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China), followed by centrifugation at
14,000 × g for 10 min. After centrifugation, the cell lysates were
collected and the protein concentrations measured. Proteins (20–30
µg) were resolved by SDS-PAGE, and then transferred to PVDF
membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
membranes were blocked with 5% BSA at room temperature for 3 h, and
then incubated with primary antibodies in 3% BSA/TBST at 4°C
overnight, followed by incubation with secondary antibodies
(Auragene Bioscience Corp., Inc., Changsha, China) at room
temperature for 1 h. The protein signals were detected by the ECL
method. The following antibodies were used for western blotting:
NAC1, GAPDH, β-actin, IFNβ, RANK, MMP9 and PKM2 antibodies, all
purchased from Abcam.
MTT assay
DU-145 cell growth ability was measured by MTT
assay. Briefly, cells subjected to different treatments were plated
at a density of 3×103 cells per well in 96-well plates,
and then incubated at 37°C in a humidified atmosphere containing 5%
CO2/95% air. After 24, 48, 72 and 96 h of incubation,
MTT reagent was added to each well, and the samples were incubated
for another 4 h. The resultant formazan product was dissolved in
DMSO and absorbance was read at 490 nm on a Victor3 Multi Label
plate reader (PerkinElmer, Boston, MA, USA).
Colony formation assay
At 24 h after transfection, DU-145 cells
(1.5×103 cells per well were plated in 6-well plates and
incubated at 37°C in a humidified atmosphere containing 5%
CO2/95% air for 14 days to allow for the formation of
colonies. The colonies were washed twice with PBS, and the cells
were then stained with 0.1% crystal violet for 20 min and washed
with water before the colonies (each ≥50 cells) were counted.
Wound healing assay and Transwell
assay
To study cell migration, DU-145 cells were cultured
to confluence in 6-well plates, and a 2-mm-wide single,
uninterrupted scratch in the cell layer was made across the
diameter of each well. The culture plates were incubated with
RPMI-1640 for 24 h and then washed with PBS, and photomicrographs
were captured to assess the change in the size of the wound
area.
Transwell assays were also performed to quantify the
migration of DU-145 cells. Uncoated inserts (cat. no. 354578; BD
Biosciences, San Jose, CA, USA) were used for the migration assays.
Duplicate wells were assessed. DU-145 cells were seeded on top of
the inserts in RPMI-1640 and cultured for 24 h, after which the
cells that had migrated to the underside of the membrane were
visualized. Cells were counted in five individual fields on each
insert.
Microarray analysis
DU-145 cells were transfected with the two NAC1
siRNAs or a negative control siRNA. At 48 h after transfection, the
cells were harvested for microarray analysis. An Arraystar Human
messenger RNA (mRNA) Microarray v3.0 was used and run by the
service provider. The array contained 30,215 protein-coding
transcripts. A specific exon or splice junction probe which could
identify an individual transcript accurately was used to represent
each transcript. Positive probes for housekeeping genes and
negative probes were also included in the array for hybridization
quality control. The microarray hybridization and bioinformatics
analysis were performed by KangChen Bio-tech (Shanghai, China).
RT-qPCR
Total RNA was extracted from the cells with TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. The relative expression levels of
the mRNAs were determined by real-time RT-qPCR using a standard
SYBR Green RT-PCR Kit (Takara Bio, Inc., Otsu, Japan). The specific
primer pairs used in this study are listed in Table I. Averages of the Ct values from
duplicate measurements were obtained. The relative expression
levels of mRNAs were quantified using GraphPad Prism v5.0 software
(GraphPad Software, Inc., San Diego, CA, USA) and the
2−ΔΔCt method.
 | Table I.Oligonucleotide primers used for
reverse transcription-quantitative polymerase chain reaction. |
Table I.
Oligonucleotide primers used for
reverse transcription-quantitative polymerase chain reaction.
| Primer | Sequence |
|---|
| NAC1 F |
5′-CCAGACACTGCAGATGGAGA-3′ |
| NAC1 R |
5′-AAGCTGAGGATCTGCTGGAA-3′ |
| IFNβ F |
5′-GCTCTCCTGTTGTGCTTCTCCACT-3′ |
| IFNβ R |
5′-AGCTGCTTAATCTCCTCAGGGATG-3′ |
| IL-24 F |
5′-TTCTCTGGAGCCAGGTATC-3′ |
| IL-24 R |
5′-TAGAATTTCTGCATCCAGGT-3′ |
| NFATc1 F |
5′-GTCCCACCACCGAGCCCACTACG-3′ |
| NFATc1 R |
5′-GACCATCTTCTTCCCGCCCACGAC-3′ |
| S100A9 F |
5′-CACCCAGACACCCTGAACCA-3′ |
| S100A9 R |
5′-CCTCGAAGCTCAGCTGCTTG-3′ |
| GAB1 F |
5′-ATCAGAAACGCCAGCGAAGA-3′ |
| GAB1 R |
5′-TCAGATACCACAAAGCACCA-3′ |
| MMP9 F |
5′-TGTACCGCTATGGTTACACTCG-3′ |
| MMP9 R |
5′-GGCAGGGACAGTTGCTTCT-3′ |
| VEGFA F |
5′-GAGCCTTGCCTTGCTGCTCTA-3′ |
| VEGFA R |
5′-CACCAGGGTCTCGATTGGATG-3′ |
| RANK F |
5′-CATCGGGTTCCCATAAAG-3′ |
| RANK R |
5′-GAAGCAAATGTTGGCGTA-3′ |
| GAPDH F |
5′-TTGGTATCGTGGAAGGACTC-3′ |
| GAPDH R |
5′-ACAGTCTTCTGGGTGGCAGT-3′ |
Co-culture studies
Boyden chambers (3-µm hole inserts; Corning
Incorporated, Corning, NY, USA) were used to assess the influence
of PC-3 cells on osteoclasts using RAW 264.7 mouse monocytes. RAW
264.7 cells (1×105) were plated in 6-well plates for 24
h, then PC-3 cells (1×105) transfected with NAC1 siRNA
or negative control siRNA were added into the upper chamber.
RPMI-1640 supplemented with 10% fetal bovine serum was used to
culture the PC-3 cells and DMEM supplemented with 10% fetal bovine
serum was used to culture the RAW264.7 cells. IFNβ concentrations
were detected by ELISA of the co-culture medium (as described
below), and osteoclast formation was evaluated by RT-qPCR and
western blot analyses of the total RNA and protein extracted from
RAW 264.7 cells after culturing for 72 h.
Enzyme-linked immunosorbent assay
(ELISA)
The protein level of IFNβ in the conditioned medium
was assessed using human immunoassay ELISA kits (CUSABIO, Wuhan,
China). The optical densities were measured at 450 nm and the
detection limits were set in accordance with the log-log
correlative coefficient of the standard curve.
Statistical analysis
Data were expressed as the mean ± SEM. Student's
t-test (for comparison between two groups) was used to evaluate the
numeric data. The χ2 test was used for comparisons of
categorical data. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
NAC1 expression is up-regulated in the
progression of prostate cancer
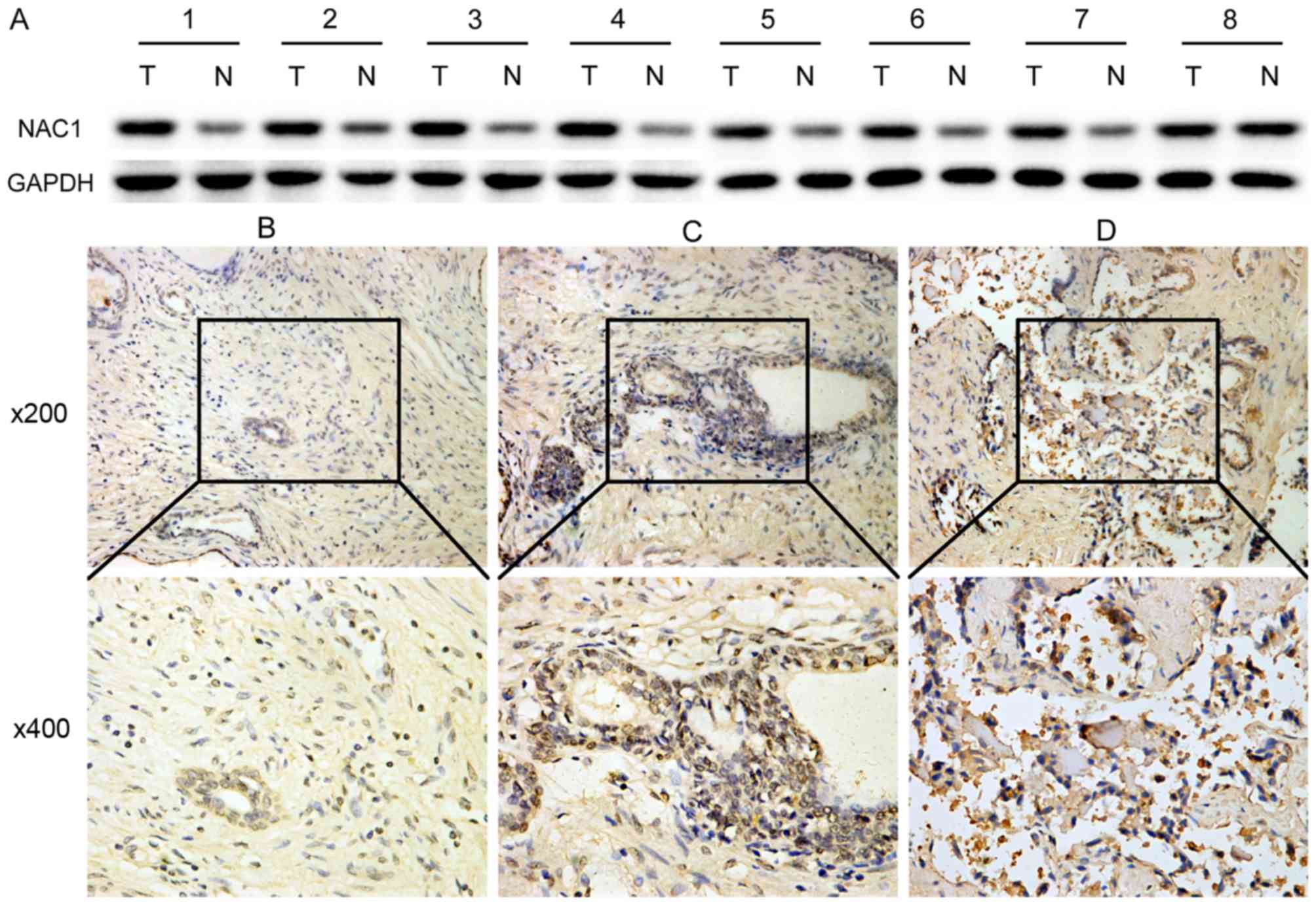
Initially, we examined the expression of NAC1
protein by western blotting in 8 prostate cancer and 8 adjacent
normal tissues taken from the same patients. The results revealed
that NAC1 expression was markedly higher in tumor tissues (Fig. 1A). To validate this result,
immunohistochemistry was used to analyze the correlation of NAC1
expression with several clinicopathological characteristics in 84
prostate cancer and 84 non-cancerous tissue samples. In contrast to
non-cancerous prostate tissues, prostate cancer demonstrated a
higher NAC1 immunoreactivity, with 79% of the cases showing
reactivity scores of 2+ or 3+ (χ2 test, P<0.001)
(Fig. 1B-D and Table II). It also revealed that the NAC1
immunoreactivity was closely related to the clinical stage of
prostate cancer (P<0.05). However, there were no significant
associations with patient age, preoperative prostate-specific
antigen level or Gleason score (Table
II).
| Table II.Correlation of NAC1 expression with
clinicopathologic factors in prostate cancer. |
Table II.
Correlation of NAC1 expression with
clinicopathologic factors in prostate cancer.
|
|
| Immunointensity, n
(%) |
|
|---|
|
|
|
|
|
|---|
| Factors | Case no. | 0 or 1+ | 2+ or 3+ | P-value |
|---|
| Histologic
type |
|
|
| <0.001 |
|
Prostate cancer | 84 | 18 (21) | 66 (79) |
|
| Normal
prostate | 84 | 62 (74) | 22 (26) |
|
| Patients' age |
|
|
| 0.09 |
| <60
y | 20 | 7
(35) | 13 (65) |
|
| ≥60
y | 64 | 11 (17) | 53 (83) |
|
| Clinical
stages |
|
|
| 0.002 |
|
≤IIb | 38 | 14 (37) | 24 (63) |
|
|
≥IIc | 46 | 4 (1) | 42 (99) |
|
| Preoperative |
|
|
| 0.878 |
| PSA (ng/ml) |
|
|
|
|
|
≤20 | 48 | 10 (21) | 38 (79) |
|
|
>20 | 36 | 8
(22) | 28 (78) |
|
| Gleason score |
|
|
| 0.437 |
| ≤7 | 62 | 12 (19) | 50 (81) |
|
| ≥8 | 22 | 6
(27) | 16 (73) |
|
Silencing NAC1 expression decreases
the migratory ability of prostate cancer cells
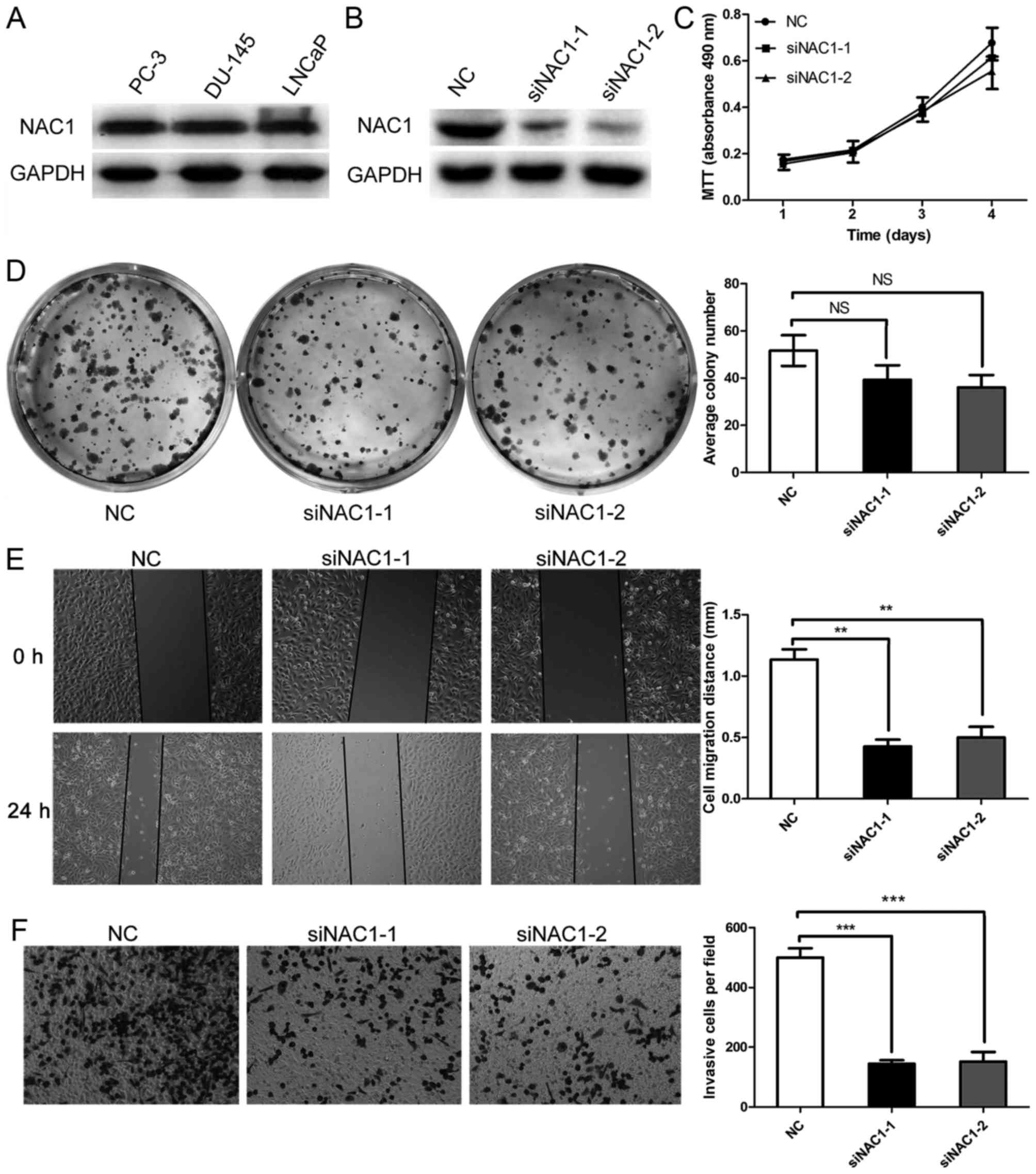
Since NAC1 was up-regulated in prostate cancer at
advanced clinical stages, we next assessed the biological function
of NAC1 in prostate cancer cells. NAC1 was expressed in the highly
aggressive prostate cancer cell lines PC-3, DU-145 and LNCaP
(Fig. 2A). DU-145 cells, which have
the highest degree of malignancy, were selected for further study.
Two NAC1 siRNAs (designated siNAC1-1 and siNAC1-2) were transfected
into the cells. Treatment with siNAC1-1 or siNAC1-2 significantly
reduced NAC1 protein expression compared with that in DU-145 cells
transfected with negative control siRNA (NC) (Fig. 2B). However, siNAC1-1 and siNAC1-2 did
not significantly reduce the numbers of DU145 cells compared with
the NC group (Fig. 2C). To confirm
the result, a colony formation assay was performed, which revealed
a slight but non-significant difference between the NAC1 siRNA and
NC groups (Fig. 2D).
Subsequently, we investigated whether NAC1 played a
causal role in the migration characteristics of prostate cancer
cells by employing a wound-healing assay. A wound was scratched on
the surface of DU-145 cells transfected with siNAC1-1, siNAC1-2 or
NC siRNA, and the width of the gap was measured after 24 h. The
cells in the siNAC-1 and siNAC1-2 groups migrated less than those
in the NC group, as shown in Fig. 2E.
Furthermore, Transwell assays demonstrated that down-regulation of
NAC-1 by siNAC1 in DU-145 cells decreased the cell migration
abilities compared with NC siRNA (Fig.
2F). These results revealed that silencing of NAC1 expression
inhibited the migration of prostate cancer cells.
Identification of NAC1-regulated genes
in prostate cancer cells
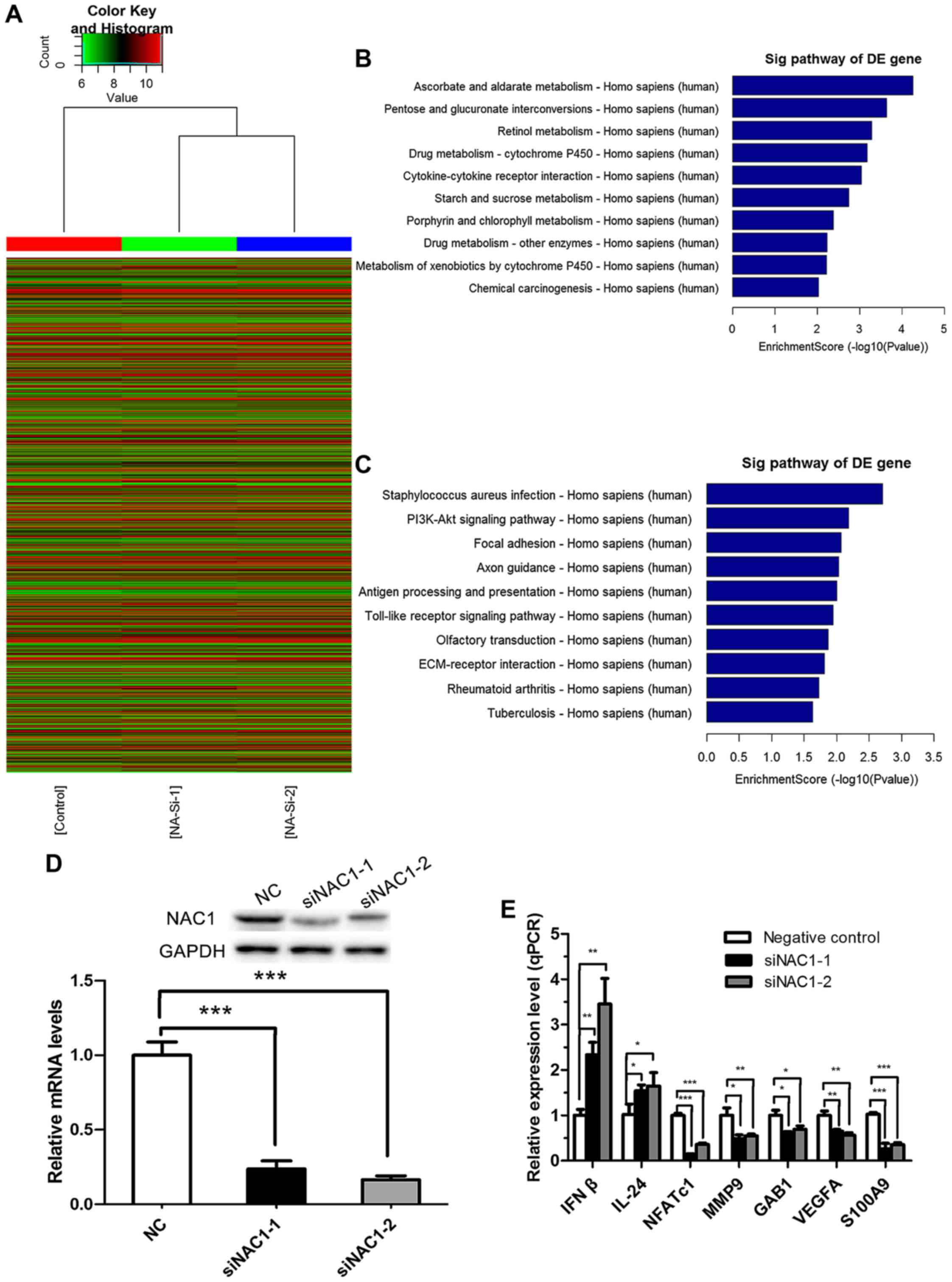
According to the preceding data, we performed a
microarray analysis to identify the potential genes regulated by
NAC1 in the DU-145 cell line. mRNA expression profiles were
determined in DU145 cells transfected with NAC1 or NC siRNA. In
total, 291 up-regulated mRNAs and 488 down-regulated mRNAs (with
fold change |>2| and P<0.05) were identified consistently in
the DU145 cells transfected with siNAC1-1 or siNAC1-2 compared with
the NC siRNA-transfected cells (data not shown). The results of a
Hierarchical Clustering analysis are shown in Fig. 3A, and the top ten pathways associated
with the up-regulated and down-regulated genes are shown in
Fig. 3B and C respectively.
To validate the microarray results, qPCR was
performed in cells transfected with the two independent NAC1
siRNAs, in order to assess the levels of seven genes identified as
being associated with the biological process of cancer development
based on the microarray analysis. All genes were validated by both
siRNAs (P<0.05). NAC1 knockdown efficiency was verified by
RT-qPCR and western blotting (Fig.
3D). Furthermore, we selected β-actin and PKM2 whose expression
was unchanged in the microarray to verify the efficiency by western
blotting (data not shown). The results revealed that NAC1 siRNAs
did not affect the expression of β-actin and PKM2, but effectively
down-regulated NAC1. As shown in Fig.
3E, the seven genes were IFNβ and IL-24, whose transcription
was repressed by NAC1, and NFATc1, MMP9, GAB1, VEGFA and S100A9,
whose transcription was promoted by NAC1.
NAC1 participates in
osteoclastogenesis by negatively regulating IFNβ
Interestingly, upon further exploration of the
microarray data, a GO analysis of biological processes revealed
that NAC1 negatively regulates IFNβ (data not shown). A previous
study reported that IFNβ has the function of regulating
osteoclastogenesis (18); therefore,
we hypothesized that NAC1 may mediate osteoclastogenesis by
negatively regulating IFNβ expression in prostate cancer. To verify
this hypothesis, PC-3 cells, which are isolated from a bone
metastatic site of prostate cancer, were employed for further
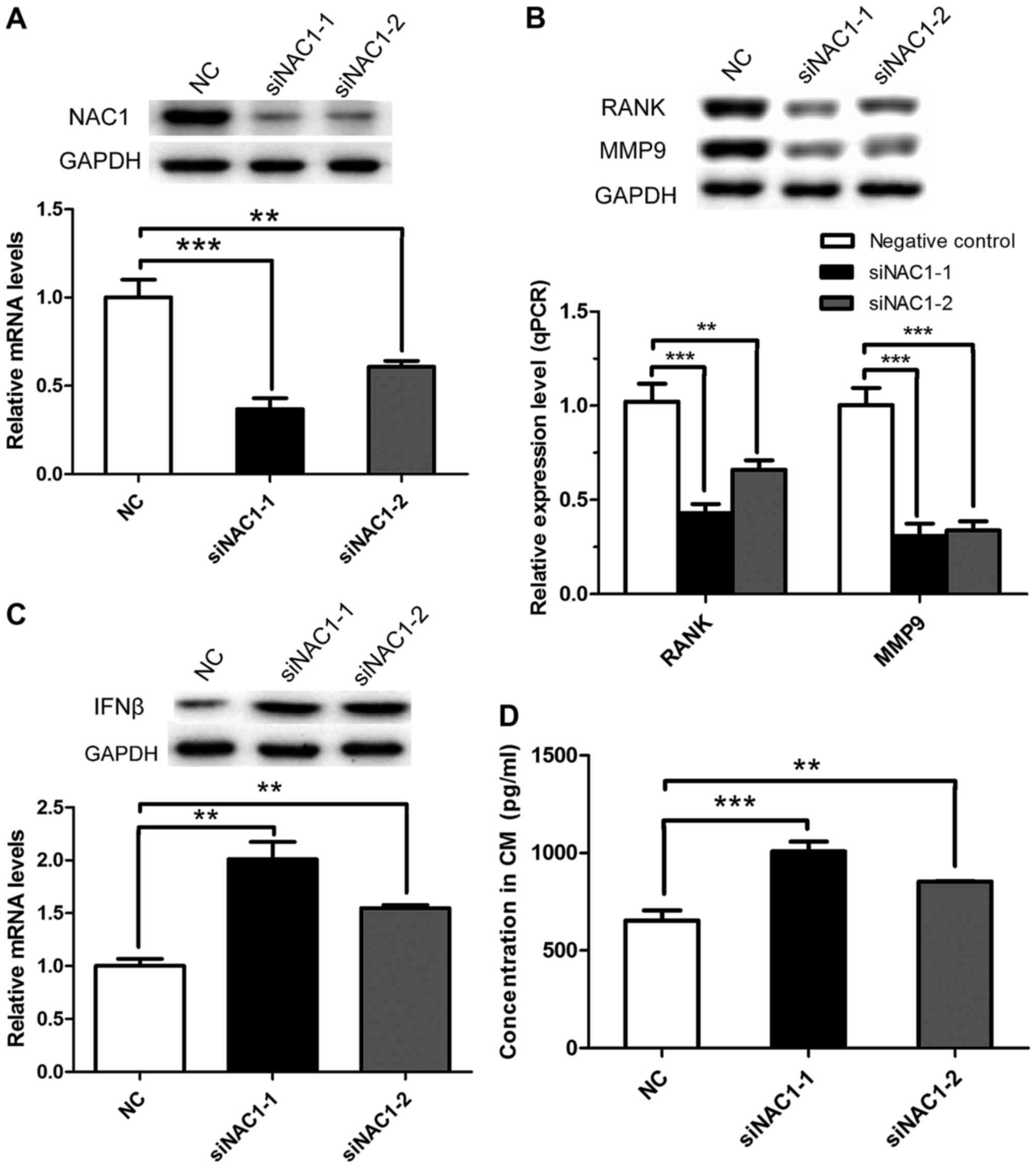
study. PC-3 cells were transfected with the two NAC1 siRNAs or NC
siRNA and after 24 h were seeded into the upper compartment of a
Boyden chamber, while RAW 264.7 cells were cultured in the bottom
chamber to allow exchange of secreted factors that generate
osteoclasts. When NAC1 was silenced in PC-3 cells (Fig. 4A), the expression of genes associated
with the activation of osteoclasts (such as RANK and MMP9) was
significantly decreased in RAW 264.7 cells, as identified by
RT-qPCR and western blot analyses (Fig.
4B). The mRNA and protein levels of IFNβ were increased in PC-3
cells transfected with NAC1 siRNAs compared with the NC siRNA
(Fig. 4C). Furthermore, the protein
level of IFNβ, detected by ELISA, was increased in the co-culture
medium in the NAC1 siRNA-transfected group compared with the
control group (Fig. 4D). This series
of results confirmed that NAC1 participates in osteoclastogenesis
through the negative regulation of IFNβ.
Discussion
The key findings of the present study were that, as
a cancer-associated gene, NAC1 promotes the migration of prostate
cancer cells and participates in osteoclastogenesis. NAC1 is
up-regulated at advanced clinical stages of prostate cancer,
indicating that it may be useful biomarker for tracking the
progression of prostate cancer. At present, the treatment of bone
metastasis in patients with prostate cancer is unsatisfactory. As
an upstream regulatory gene of osteoclastogenesis, NAC1 may
represent a novel potential therapeutic target.
The results of previous studies have implicated NAC1
in cell proliferation and survival in certain gynecological tumors,
such as ovarian cancer, endometrial cancer, cervical carcinomas and
uterine sarcoma (11,17,19,20).
However, interestingly, the results of the colony formation and MTT
assays in the present study showed that NAC1 expression had no
significant effects on cell proliferation in prostate cancer
(Fig. 2C and D), which is similar to
our previous studies in bladder urothelial carcinoma (In press).
However, in the Transwell and wound healing assays (Fig. 2E and F), NAC1-silencing resulted in a
decreased capacity for cell migration in the DU-145 cell line,
indicating that NAC1 may be essential for metastasis in prostate
cancer. Similarly, NAC1 has been shown to promote cell invasion and
motility through FOXQ1 in ovarian cancer (16). Furthermore, NAC1 participate in
multitudinous cancer-associated pathways like autophagy, apoptosis
and epithelial mesenchymal transition in different cancer cells
(21–23), indicating that NAC1 exert different
effects in different cancer types. This may account for why NAC1
inhibition suppresses proliferation in some cancer cell types
(11,19), but not in the prostate cancer, where
it suppresses migration.
On the basis of comparing the transcripts present in
NAC1-knockdown and control DU-145 prostate cancer cells by
microarray analysis, we validated a set of target genes whose
expression levels depend on NAC1. After NAC1 silencing, IFNβ and
IL-24 were up-regulated, and NFATc1, S1009, GAB1, MMP9 and VEGFA
were down-regulated (Fig. 3E). In
these genes, IFNβ is particularly interesting, as previous studies
have shown that IFNβ can negatively regulate osteoclastogenesis;
this mechanism involves the secretion of IFNβ by osteocytes, which
then acts on osteoclast precursors and down-regulates the protein
level of c-Fos, a key factor in osteoclast formation (20,24,25).
Combined with the GO analysis (data not shown), which demonstrated
that NAC1 can negatively regulate IFNβ, we hypothesized that NAC1
may participate in osteoclastogenesis by reducing IFNβ secretion.
To validate this hypothesis, PC-3 cells isolated from prostate
cancer bone metastasis were co-cultured with RAW 264.7 cells. RANK
and MMP9, which are associated with the activation of osteoclasts,
were down-regulated in RAW 264.7 cells when NAC1 was significantly
silenced in PC-3 cells (Fig. 4B).
Furthermore, the protein level of IFNβ was increased in the culture
medium of PC-3 cells (Fig. 4D). This
finding is consistent with the supposition that NAC1 may play an
important role in promoting osteoclastogenesis.
However, it should be noted that this study was only
an in vitro examination. Therefore, further studies in
vivo are needed to clarify the relationship between NAC1 and
osteoclastogenesis. Moreover, other underlying mechanisms of
prostate cancer migration and its downstream target genes must be
further explored. For example, VEGFA, the most important factor
among the angiogenic cytokines (26–28), was
identified to be positively regulated by NAC1, suggesting that NAC1
may play an important role in promoting tumor angiogenesis in
prostate cancer. Indeed, several NAC1-regulated genes identified in
the microarray analysis, including IL-24, GAB1 and S100A9, are also
involved in cell motility or invasion (29–31).
In conclusion, we have demonstrated that NAC1
promotes the migration of prostate cancer cells, and identified a
significant correlation between NAC1 and osteoclastogenesis. These
findings suggest that NAC1 may serve as a biomarker for the
diagnosis of prostate cancer in the future, and that a NAC1-based
molecular-targeting therapy is a potentially novel treatment for
prostate cancer.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rucci N and Angelucci A: Prostate cancer
and bone: The elective affinities. Biomed Res Int. 2014:1670352014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saad F, Lipton A, Cook R, Chen YM, Smith M
and Coleman R: Pathologic fractures correlate with reduced survival
in patients with malignant bone disease. Cancer. 110:1860–1867.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zustovich F and Fabiani F: Therapeutic
opportunities for castration-resistant prostate cancer patients
with bone metastases. Crit Rev Oncol Hematol. 91:197–209. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Albagli O, Dhordain P, Deweindt C, Lecocq
G and Leprince D: The BTB/POZ domain: A new protein-protein
interaction motif common to DNA- and actin-binding proteins. Cell
Growth Differ. 6:1193–1198. 1995.PubMed/NCBI
|
|
7
|
Stogios PJ, Downs GS, Jauhal JJ, Nandra SK
and Privé GG: Sequence and structural analysis of BTB domain
proteins. Genome Biol. 6:R822005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bardwell VJ and Treisman R: The POZ
domain: A conserved protein-protein interaction motif. Genes Dev.
8:1664–1677. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang J, Rao S, Chu J, Shen X, Levasseur
DN, Theunissen TW and Orkin SH: A protein interaction network for
pluripotency of embryonic stem cells. Nature. 444:364–368. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim J, Chu J, Shen X, Wang J and Orkin SH:
An extended transcriptional network for pluripotency of embryonic
stem cells. Cell. 132:1049–1061. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakayama K, Nakayama N, Davidson B, Sheu
JJ, Jinawath N, Santillan A, Salani R, Bristow RE, Morin PJ, Kurman
RJ, et al: A BTB/POZ protein, NAC-1, is related to tumor recurrence
and is essential for tumor growth and survival. Proc Natl Acad Sci
USA. 103:pp. 18739–18744. 2006; View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jinawath N, Vasoontara C, Yap KL,
Thiaville MM, Nakayama K, Wang TL and Shih IM: NAC-1, a potential
stem cell pluripotency factor, contributes to paclitaxel resistance
in ovarian cancer through inactivating Gadd45 pathway. Oncogene.
28:1941–1948. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Y, Cheng Y, Ren X, Hori T,
Huber-Keener KJ, Zhang L, Yap KL, Liu D, Shantz L, Qin ZH, et al:
Dysfunction of nucleus accumbens-1 activates cellular senescence
and inhibits tumor cell proliferation and oncogenesis. Cancer Res.
72:4262–4275. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Cheng Y, Ren X, Zhang L, Yap KL,
Wu H, Patel R, Liu D, Qin ZH, Shih IM and Yang JM: NAC1 modulates
sensitivity of ovarian cancer cells to cisplatin by altering the
HMGB1-mediated autophagic response. Oncogene. 31:1055–1064. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yap KL, Fraley SI, Thiaville MM, Jinawath
N, Nakayama K, Wang J, Wang TL, Wirtz D and Shih IeM: NAC1 is an
actin-binding protein that is essential for effective cytokinesis
in cancer cells. Cancer Res. 72:4085–4096. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao M, Wu RC, Herlinger AL, Yap K, Kim JW,
Wang TL and Shih IeM: Identification of the NAC1-regulated genes in
ovarian cancer. Am J Pathol. 184:133–140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yeasmin S, Nakayama K, Ishibashi M,
Katagiri A, Iida K, Purwana IN, Nakayama N and Miyazaki K:
Expression of the bric-a-brac tramtrack broad complex protein NAC-1
in cervical carcinomas seems to correlate with poorer prognosis.
Clin Cancer Res. 14:1686–1691. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takayanagi H, Kim S, Matsuo K, Suzuki H,
Suzuki T, Sato K, Yokochi T, Oda H, Nakamura K, Ida N, et al: RANKL
maintains bone homeostasis through c-Fos-dependent induction of
interferon-beta. Nature. 416:744–749. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ishikawa M, Nakayama K, Yeasmin S,
Katagiri A, Iida K, Nakayama N and Miyazaki K: NAC1, a potential
stem cell pluripotency factor expression in normal endometrium,
endometrial hyperplasia and endometrial carcinoma. Int J Oncol.
36:1097–1103. 2010.PubMed/NCBI
|
|
20
|
Rahman MT, Nakayama K, Ishikawa M, Rahman
M, Katagiri H, Katagiri A, Ishibashi T, Iida K, Yamada T and
Miyazaki K: NAC1, a BTB/POZ protein overexpressed in uterine
sarcomas. Anticancer Res. 32:3841–3845. 2012.PubMed/NCBI
|
|
21
|
Zhang Y, Yang JW, Ren X and Yang JM: NAC1
and HMGB1 enter a partnership for manipulating autophagy.
Autophagy. 7:1557–1558. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ju T, Jin H, Ying R, Xie Q, Zhou C and Gao
D: Overexpression of NAC1 confers drug resistance via HOXA9 in
colorectal carcinoma cells. Mol Med Rep. 16:3194–3200. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Faiola F, Yin N, Fidalgo M, Huang X,
Saunders A, Ding J, Guallar D, Dang B and Wang J: NAC1 regulates
somatic cell reprogramming by controlling Zeb1 and E-cadherin
expression. Stem Cell Rep. 9:913–926. 2017. View Article : Google Scholar
|
|
24
|
Hayashida C, Ito J, Nakayachi M, Okayasu
M, Ohyama Y, Hakeda Y and Sato T: Osteocytes produce interferon-β
as a negative regulator of osteoclastogenesis. J Biol Chem.
289:11545–11555. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofstetter W, Felix R, Fleisch HA and Wagner EF: c-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kieran MW, Kalluri R and Cho YJ: The VEGF
pathway in cancer and disease: Responses, resistance, and the path
forward. Cold Spring Harb Perspect Med. 2:a0065932012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Potente M, Gerhardt H and Carmeliet P:
Basic and therapeutic aspects of angiogenesis. Cell. 146:873–887.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Panneerselvam J, Jin J, Shanker M,
Lauderdale J, Bates J, Wang Q, Zhao YD, Archibald SJ, Hubin TJ and
Ramesh R: IL-24 inhibits lung cancer cell migration and invasion by
disrupting the SDF-1/CXCR4 signaling axis. PLoS One.
10:e01224392015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang J, Song W, Shen W, Yang X, Sun W, Qu
S, Shang R, Ma B, Pu M, Tao K, et al: MicroRNA-200a suppresses cell
invasion and migration by directly targeting GAB1 in hepatocellular
carcinoma. Oncol Res. 25:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lim SY, Yuzhalin AE, Gordon-Weeks AN and
Muschel RJ: Tumor-infiltrating monocytes/macrophages promote tumor
invasion and migration by upregulating S100A8 and S100A9 expression
in cancer cells. Oncogene. 35:5735–5745. 2016. View Article : Google Scholar : PubMed/NCBI
|