Introduction
Chronic inflammation is a well-established hallmark
of cancer proliferation (1). The
cellular stress caused by inflammation induces release of cytokine
and chemokine factors which induce tumor progression and metastasis
(2). Several agents have been
suggested to induce chronic inflammation (3). Recent evidence from our lab have
demonstrated that breast cancer cells cultured under high salt
conditions (Δ0.05 M NaCl, 50% above basal culture conditions) were
able to upregulate reactive nitrogen species (4,5).
Importantly, sodium-MRI studies in breast cancer patients have
demonstrated an increased sodium content, of up to 63% above the
surrounding soft tissue, in the breast tumors (6,7). These
support a notion that high salt exerts an effector role on tumor
progression, either working individually or synergistically to
enhance an inflammatory tumor microenvironment. Phospho-proteomic
based studies from our laboratory have demonstrated that high salt
(Δ0.05 M) synergized with sub-minimal stimulation of IL-17 (0.1
ng/ml) induced upregulation of SIK-3, salt inducible kinase-3, a
serine-specific protein kinase in breast cancer cells (8). Mechanistic studies have demonstrated
that SIK3 played a crucial role in induction of G1/S-phase release
of cell cycle, along with enhanced expression of metastasis
specific chemokine CXCR4 in breast cancer cells. Histone
deacetylase (HDAC4) is a well-documented downstream target of SIK3
(9). The phosophorylation of HDAC4 is
known to induce cell proliferation and malignancy. SIK3 induced
phosphorylation of HDAC4. Further, equimolar treatment with
mannitol or sucrose did not exert similar pro-cancer effect, thus
strongly suggesting that high sodium chloride specifically induces
a pro-cancer effect (8).
Natural plant product prostratin (12-deoxyphorbol
13-acetate) is widely studied for it's activation of latent T-cells
infected with HIV. Prostratin is identified as a pharmacologically
active ingredient in the Samoan medicinal plant Homolanthus
nutans (10). Molecular studies
have demonstrated that prostratin exerts its anti-HIV effect
through activation of protein kinase C (11). Importantly, prostratin is shown to
induce downregulation of chemokine receptor CXCR4 in CD4+T
lymphocytes there by preventing the entry of HIV-1 virus into
lymphocytes (12).
The C-X-C chemokine receptor type 4 (CXCR4), belongs
to the superfamily of the heterotrimeric G protein-coupled
receptors and is expressed on the cell surface of various types of
metastatic breast cancer cells. It is important to note that the
role of CXCR4 in cell proliferation and metastasis was obtained
from the elegant studies performed by Dewan et al (13), when less invasive MCF-7 breast cancer
lines with low expression of CXCR4 formed much smaller tumor mass
the SCID-mice compared to those larger tumor size the highly
invasive MDA-MB-231 cell line with high expression of CXCR4. This
is further corroborated with the studies by Lapteva et al
(14), where in, siRNA based
knock-down of CXCR4 in breast cancer cells tumors cells decrease
the tumor growth and size in the murine breast cancer models. Taken
together, as prostratin has shown to mediate CXCR4 downregulation
in HIV infected cells and CXCR4 is important for cancer
proliferation, in our current communication, we studied the
anti-cancer effect of prostratin through modulation of CXCR4
expression on cancer cells.
Materials and methods
Cell cultures and plasmids
Five breast tissue related cell lines were used in
our studies, of these, four breast cancer cells (MCF7, MDA-MB-231,
BT20, AU565) and one normalized breast epithelial cell line
(MCF10A) were utilized and obtained from the American Type Culture
Collection (ATCC, Manassas, VA, USA). The cells were cultured in
cell basal essential media (RPMI-1640 media; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) along with the media
supplements such as fetal bovine serum, penicillin/streptomycin,
fungizone, HEPES and glutamine, as recommended by the manufacturer
and as previously described (4,15). Cell
lines were frozen in liquid vapor nitrogen at −130°C until use.
Upon thawing, cells were maintained in 5% CO2 incubator
in sterile essential media at 37°C. For salt and interleukin-17
treatment conditions, cell culture media was supplemented with 0.05
M NaCl (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 0.1
ng/ml IL-17 (Thermo Fisher Scientific, Inc.). We have previously
performed a dose-response for salt (0–0.1 M NaCl) and IL-17
(0–1,000 ng/ml) and found-out that 0.05 M NaCl provided highest
cell proliferation (4,5) and 0.1 ng/ml of IL-17 induced
sub-effective inflammatory cytokine response (4,16). All
chemicals unless mentioned were obtained either from Sigma-Aldrich;
Merck KGaA (Darmstadt, Germany) or Thermo Fisher Scientific, Inc.
For siRNA knock down of SIK3 we have used the following two siRNA
sequences: SIK3-siRNA-1: 5′-GUGCAGAGUGUUGGAGUCC-3′; scramble
SIK3-siRNA-1: 5′-UGGAGGCGAGUCAGUUUGC-3′.
Western blot/immunoprecipitation
Total proteins were extracted from cells with lysis
buffer for western blot analysis as previously described (17,18). Total
proteins were separated on a 4–12% sodium dodecyl
sulfate-polyacrylamide gradient gel and transferred onto a
nitrocellulose membrane. The membranes were blocked overnight at
4°C in Tris-buffered saline with 0.05% Tween 20 (5% nonfat milk in
10 mM Tris-HCl-100 mM NaCl-0. 1% Tween 20, pH 7.4). The membranes
were incubated first with Abs specific for total and phosphorylated
forms at room temperature with primary Abs diluted 1 in 1,000 in
blocking buffer for 2 h, and then with a horseradish
peroxide-conjugated secondary IgG mAb diluted 1 in 5,000 for 1 h.
All primary and secondary Abs were obtained from Santa Cruz
Biotechnology, Inc., (Dallas, TX, USA). All primary and secondary
antibodies for western blot and immunoprecipitation were obtained
from either Abcam (Cambridge, MA, USA) or Santa Cruz Biotechnology,
Inc. The following specific primary antibodies to SIK3 (ab211424;
Abcam) GADPH (sc-47724), Actin (sc-8432), HDAC4 (sc-46672; all from
Santa Cruz Biotechnology, Inc.), pHDAC4-S632 (ab39408; Abcam). The
membrane was developed using the chemiluminescence kit (EMD
Millipore, Billerica, MA, USA) and analyzed on using Universal Hood
II by Bio-Rad Laboratories, Inc., (Hercules, CA, USA). Morphometric
analysis was done using the software provided by the company.
For SIK3 immunoprecipitation, the cultured cells
were washed with cold PBS, and lysed for 30 min on ice with 0.5 ml
of lysis buffer as previously mentioned (16,19). To
the lysis buffer 0.5 ml of dilution buffer was added and
centrifuged at 17,000 × g for 30 min. The supernatant was
transferred and 1 µg normal chicken IgY (ab97135; Abcam) or chicken
anti-SIK3 were added. After overnight incubation at 4°C, 30 µl
carbolink beads (Pierce, Thermo Fisher Scientific, Inc.) were added
to lysates and incubated for 2 h for chicken antibody
immunoprecipitation as per manufacturer's protocol. Beads were
washed with 700 µl of wash ice cold buffer four times, 3 min each
time followed by centrifugation at 1,800 × g for 3 min at 4°C.
Beads were then washed with cold PBS and bound proteins were eluted
by boiling with 30 µl of 2X SDS buffer for 10 min. Proteins were
subjected to SDS-PAGE (4–12% gel) and immunoblotting.
Phosphorylation of SIK3 were detected with a mouse monoclonal
phospho-serine antibody (sc-81514; Santa Cruz Biotechnology,
Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Expression profiles of genes at mRNA level in the
breast cancer cell lines were analyzed using the TaqMan FAM-labeled
RT-PCR primers for SIK3 (Hs00228549_m1), GADPH (Hs402869), and
Actin (Hs4333762T), obtained from Applied Biosystems; Thermo Fisher
Scientific, Inc. as per the manufacturer's recommendation. Briefly,
total RNA was extracted from 106 cells using TRIzol
reagent (Sigma-Aldrich; Merck KGaA) and analyzed as mentioned
previously (20–22). RNA samples were quantified by
absorbance at 260 nm. The RNA was reverse-transcribed and RT-PCR
(real time PCR) was performed in a final reaction volume of 20 µl
using CFX96 (Bio-Rad Laboratories, Inc.). Each sample was analyzed
in triplicate. Cycling conditions consisted of an initial
denaturation of 95°C for 15 min, followed by 40 cycles of 95°C for
30 sec, followed by 61°C for 1 min.
Cell proliferation assay
Cell viability was measured by trypan blue dye
exclusion (Sigma-Aldrich; Merck KGaA) and MTT assay (Thermo Fisher
Scientific, Inc.) as previously described (4). Briefly, the viability of breast cancer
cells was assessed by measuring mitochondrial activity using MTT
(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium bromide) assay.
For various treatment conditions the cancer cells were plated in 96
well plate for 48–72 h, the cells were incubated with 5 mg/ml MTT
in PBS for 2 h, latter lysed with manufacturer provided reagents.
Detection at 570 nm was performed using EMax Plus spectrophotometer
and data analysis was carried out using software provided by the
manufacturer (Molecular Devices, LLC, Sunnyvale, CA, USA).
Viability was calculated as percentage compared to untreated cells.
Drug dose-reponse parameters were obtained using the following
equation:
y=100–[A*x/(K+x)]
where, y is cell viability (%); x is
prostratin concentration (µM); A is defined as the maximum
cell viability following highest drug treatment, this data will be
reported as maximal prostratin cytoxicity (Cmax), as
represented as (100-A); K is defined as the concentration of
prostratin at which there is 50% of Cmax loss of cell
viability and is reported as IC50. While Cmax and IC50 are obtained
by best curve fit (with R-square value >0.95), the highest drug
treatment within the limitation of our experimental data collection
is shown at approximately three fold concentration above IC50. The
best curve fit was analyzed using Microcal Origin v7.0 (Microcal
Software, Westborough, MA, USA).
HDAC4 assay kit
The HDAC4 activity analysis (Epigentek, Farmingdale,
NY, USA) was performed on the nuclear fragments of the cell lysates
under various assay conditions as per manufacturer's instructions.
Calorimetry detection at 450 nm was performed using EMax Plus
spectrophotometer and data analysis was carried out using software
provided by the manufacturer (Molecular Devices, LLC). The data
analysis was performed based on a standard curve obtained using the
positive controls provided by the manufacturer as previously
described.
CXCR4 membrane expression assay
CXCR4 expression was analyzed by flow cytometry as
previously described (23). Briefly,
the CXCR4 protein was labeled by mouse anti-CXCR4 primary antibody
(Santa Cruz Biotechnology, Inc.) in 1:20 dilution to a 200 µl final
volume of cells (1×105 cells/ml). Antibodies used for
flow cytometry included anti-mouse-FITC (BD Biosciences, San Jose,
CA, USA), and the samples were latter analyzed using a FACS
Calibur/LSRII flow cytometer (BD Biosciences). Data were analyzed
using BD FACSDiva software. Gates were set according to isotype
controls.
Statistical analysis
All data were presented as mean values ± SEM from
four independent experiments. Student t-test performed for
statistical analysis. P<0.05 was considered to indicate a
statistically significant difference. All data analysis were
performed using Origin 6 software (Origin Labs, Northampton, MA,
USA) or SPSS software v21 (IBM Corp., Armonk, NY, USA).
Results
Enhanced cytotoxic effect of
prostratin on breast cancer cells in the presence of high salt
environment
To study the effect on prostratin on breast cancer
cells in the presence on high salt environment, we first determined
the drug's cytotoxicity. We have previously demonstrated that high
salt (Δ0.05 M NaCl) synergized with subminimal IL-17 (0.1 pg/ml)
(16) to induce a 24% enhanced cell
proliferation (4,5,8,22), which will be referred to as
stimulating conditions in our current communication. Importantly,
Na-MRI based human studies have revealed that breast tumors
accumulate high salt for yet unknown reasons (6,7). As shown
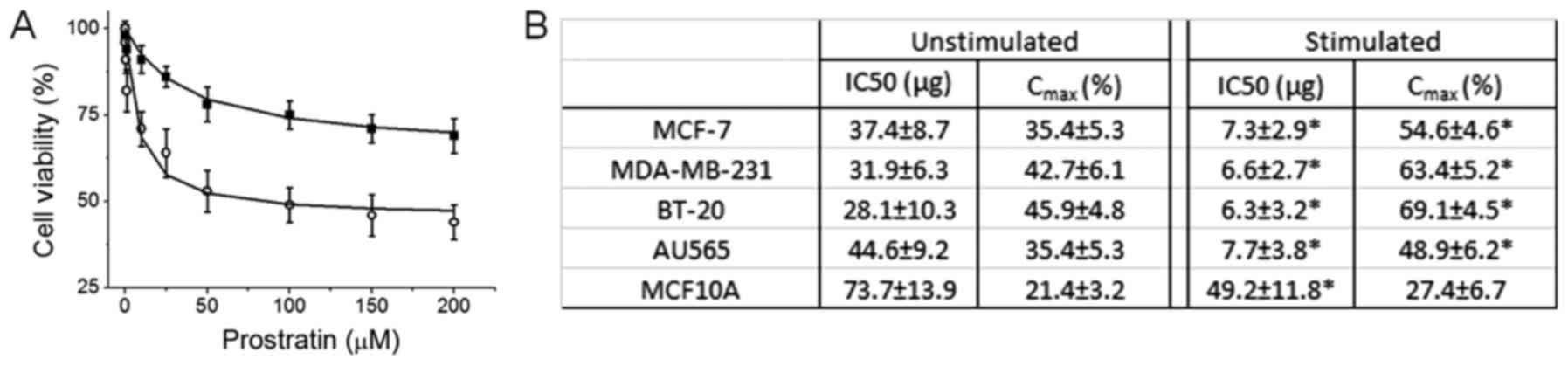
in Fig. 1A, prostratin induced
inhibitory effect on cell proliferation in MCF-7 breast cancer
cells, however, this cytotoxic effect was higher when cells were
culture in high salt culture environment. Further data analysis has
revealed that (Fig. 1B) the IC50 of
prostratin on MCF-7 cells under basal culture conditions was
37.4±8.7 µg, while under high salt stimulating conditions the IC50
was determined to be 7.3±2.9 µg. This suggests that natural plant
product prostratin has more efficient anti-tumor activity under
tumor microenvironment conditions similar to real human cancer
patients. We have further verified this cytotoxic effect on other
breast related cell lines. In our current study we have used four
breast cancer cell lines namely, MCF-7 (ER/PR double positive),
MDA-MB-231 (triple negative), BT-20 (Triple negative), AU-565 (Her2
positive); and one non-malignant breast epithelial cell line,
MCF-10A. As shown in Fig. 1B,
prostratin has upto seven fold higher cytotoxicity (as determined
by low IC50) on breast cancer cells over non-malignant cells. Of
the four breast cancer cell lines, although statistically
insignificant, prostratin seemed to exert higher effect (as
determined by Cmax) on the two highly invasive
(metastatic) cell lines, MDA-MD-231 and BT-20 (Fig. 1B). These data strongly suggest that
prostratin can selectively exert it's effect on breast tumor with
minimal effect on normal breast epithelium.
Prostratin inhibits SIK3 expression
and phosphorylation
As we have previously demonstrated that SIK3 plays a
critical role in mediating high salt induced cancer cell
proliferation (8). In our current
study, we studied the potential cytotoxic effect of prostratin
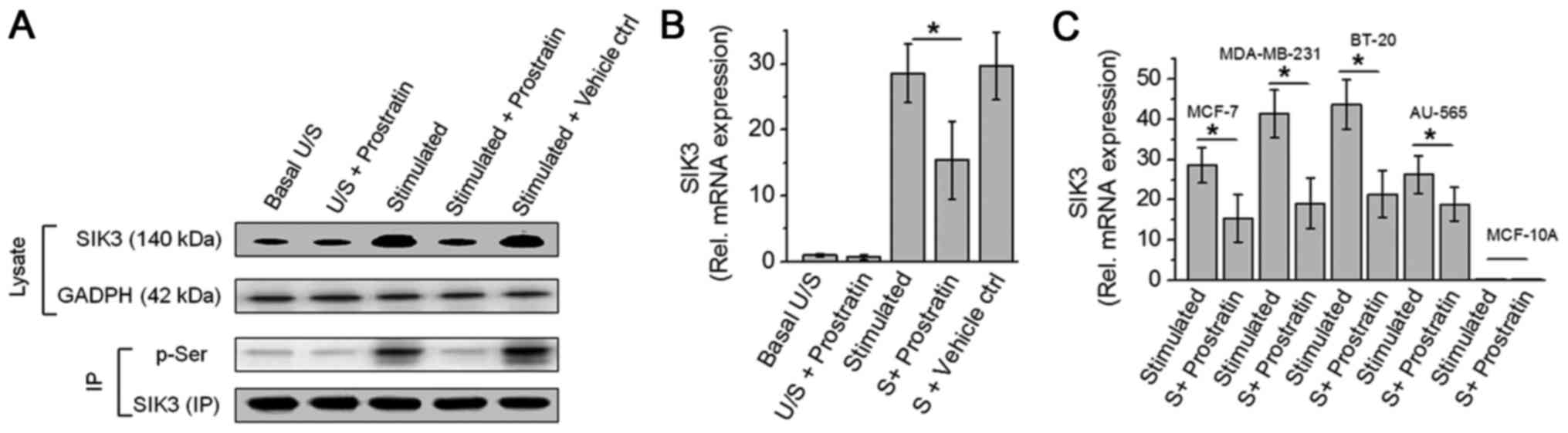
through inhibition of SIK3. As shown in Fig. 2, prostratin induced a 46% inhibition
in the SIK3 mRNA expression following stimulation with high salt on
MCF-7 cells. Prostratin was able to exert this SIK3 inhibition on
all four breast cancer cells (MCF-7, MDA-MB-231, BT-20 and AU-565).
Similar to evidence from cytotoxicity studies, prostratin exerted
slightly greater inhibition (54% vs. 46% inhibition, P>0.05,
statistically non-significant) of SIK3 expression in highly
invasive MDA-MB-231 and BT-20, over less invasive MCF-7 and AU-565
breast cancer cell lines. Furthermore, along with reduced SIK3
expression prostratin also inhibited the phosphorylation of SIK3
suggesting an inhibition of SIK3 mediated downstream signaling.
Prostratin inhibits activation of
HDAC4, a SIK3 downstream element
Previous studies in our laboratory have demonstrated
that high salt stimulating conditions induce SIK3 mediated
phosphorylation of HDAC4, and siRNA mediated knock-down of SIK3
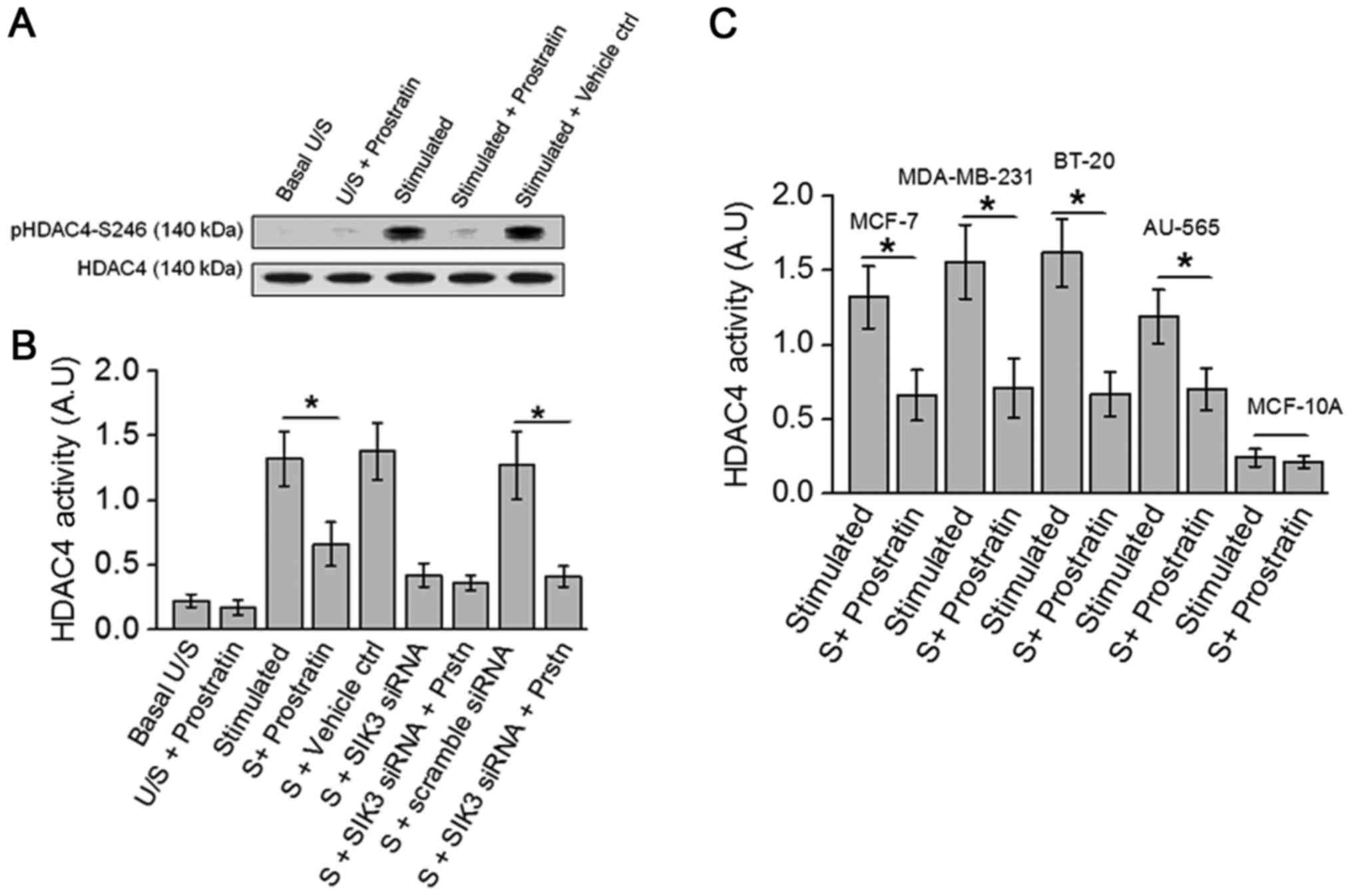
completely abrogated HDAC4 phosphorylation (8). Towards this, as prostratin inhibited
SIK3, we studied the drug effect on HDAC4 phosphorylation, a SIK3
downstream element. As shown in Fig.
3A, prostratin inhibited the phosphorylation of HDAC4. Further,
ELISA based biochemical analysis of HDAC4 activity (Fig. 3B) demonstrated that prostratin treated
cell lysate obtained following treatment with high salt stimulation
induced a 50% inhibition of HDAC4 specific substrate conversion.
Taken together, these data indicate that prostratin exerts its
anti-tumor effect through inhibition of SIK3/HDAC4 mediated cell
proliferation.
Prostratin downregulates CXCR4 in SIK3
dependent manner
As our cytotoxicity and SIK3 functional mechanistic
data suggested that prostratin exerted a slightly higher (but
statistically insignificant) inhibitory effect on highly invasive
MDA-MB-231 and BT-20 cancer cell lines, we tested if prostratin
could exerted anti-cancer effect through inhibition of metastasis.
CXCR4 is a well-established metastatic factor which is known to
promote cancer cell invasion and spreading to other organs in
patients (24). Importantly,
prostratin has previously been demonstrated to inhibit CXCR4
expression in T-lymphocytes. Therefore, in the present study we
tested if prostratin inhibited CXCR4 expression in breast cancer
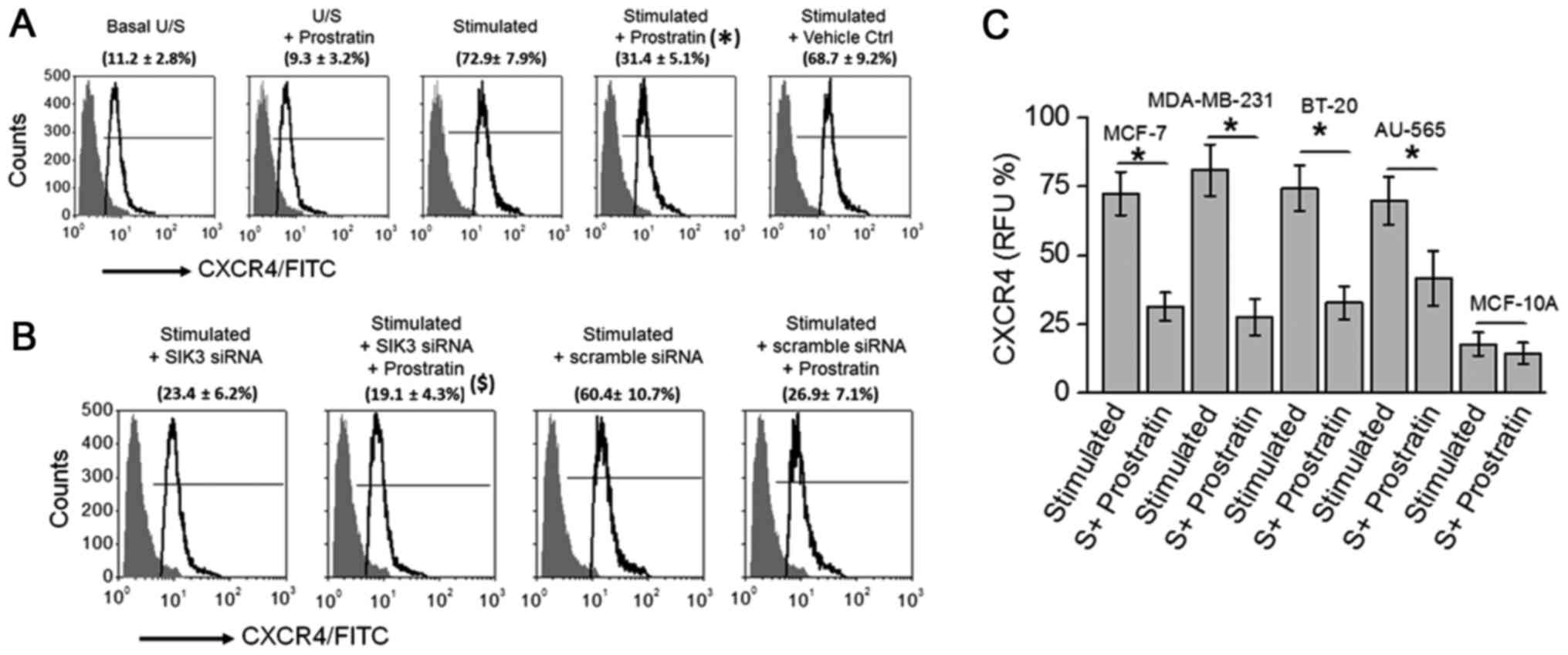
cells. As shown in Fig. 4A,
prostratin treatment inhibited the CXCR4 expression (from 72.9±7.9%
to 31.4±5.1%; P<0.05) following high salt stimulation.
Prostratin exerted no effect following siRNA mediated SIK3 knock
down, suggesting prostratin inhibits SIK3 induced CXCR4 metastatic
factor expression. Further, prostratin was able to inhibit CXCR4 in
all four breast cancer cell lines (Fig.
4C) with no effect on non-malignant breast epithelial cell
line. These data suggest that prostratin is exerts strong
anti-cancer effect through potential inhibition of metastasis.
Discussion
The natural product prostratin is extensively
researched in the context of therapy against HIV infection and
activation of CD4+T lymphocytes. However, there is limited evidence
to determine the effect of prostratin on cancer cells. Various
ligand-drug interaction studies have suggested that prostratin
exerts its anti-viral effect through the diacylglycerol (DAG)
binding domain of protein kinase C (PKC), leading to PKC enzymatic
activation (11). This logic behind
ligand mechanistics of prostratin, a phorbol ester, seems to have
to been inspired from the understanding of another structurally
related phorbol ester, phorbol 12-myristate 13-acetate (PHA), which
also stimulates PKC through allosteric activation by DAG. However,
inspite of structural similarity, while PHA is known to promote
tumor growth, prostratin is not considered to promote tumor growth.
Szallasi et al (25) have
demonstrated that prostratin was able to inhibit tumor growth in
CD1 murine cutaneous tumor models. However, to-date limited
literature on the exact cytotoxicity doses of prostratin on cancer
cell lines. In our current study we report that the IC50 of
prostratin on four different breast cancer cells under basal
culture conditions was around 35 µM (Fig.
1) however, upon stimulating culture conditions, mimicking the
real tumor microenvironment, the IC50 was around 7 µM (Fig. 1), thus suggesting that prostratin
might have an anticancer effect under in vivo solid tumor
environment in animal models and humans. Further, it is important
to note that the IC50 of prostratin on non-malignant breast cells
(MCF-10A) under basal culture conditions (which is close to basal
physiological conditions within human body) is 73 µM (Fig. 1). This data strongly suggests that
prostratin has a 10 fold higher therapeutic index and therefore
could specifically target malignant cells while being safe on
normal cells. This data could set a stage for future preclinical
studies in murine tumor models to exactly decipher the anticancer
effect and therapeutic dose of prostratin.
To date, extensive research with prostratin as an
anti-viral drug has demonstrated that the drug exerts its effect
through activation of PKC (11).
However, using fibroblast cell lines stably transfected with H-Ras
and K-Ras, Wang et al (26)
have demonstrated yet another mechanism for prostratin, wherein the
researchers have demonstrated that prostratin inhibits
K-Ras/calmodulin interaction and there by inhibiting the oncogenic
potential of their transfected cell lines. This evidence suggests,
in addition to PKC activation, prostratin could exert its
anti-cancer effect through other signaling pathways. In the present
study as prostratin demonstrated higher cytotoxicity when cells
were cultured under stimulating conditions (sub-minimal IL-17, 0.1
ng/ml, synergized high salt, Δ0.05 M NaCl, culture conditions)
commonly found in solid tumors. This data suggest that prostratin
might act through salt-specific signaling mechanism. Importantly,
we have recently demonstrated that salt-inducible kinase, SIK3, is
specifically upregulated and mediated proliferative signaling
following high salt stimulating conditions. Therefore, we tested
for the potential effect of prostratin on SIK3 expression. Previous
studies from our laboratory have demonstrated that (8), while IL-17 induced both mTORC1 and
mTORC2 pathways, only mTORC2 is a direct upstream signaling
molecule for SIK3. High salt treatment could directly induce SIK3
phosphorylation, which is further enhanced following co-treatment
with subminimal-IL-17. These studies clearly point out that SIK3 is
a direct downstream factor for high salt synergized inflammatory
stress. In our current study as we found prostratin could
down-regulate the activation of SIK3, we wanted to study under
treatment conditions wherein we had highest stimulation of SIK3 and
then quantitatively analyze for the prostratin inhibition of SIK3.
Therefore, as the focus of the current study is SIK3, to have
highest stimulation of SIK3 in our current work, we have used high
salt and sub-minimal IL-17 co-treatment conditions. Our current
studies demonstrate that prostratin inhibits SIK3 expression and
phosphorylation (Fig. 2). Also, the
phosphorylation and activity of histone deacetylase (HDAC4), a
direct downstream molecule of SIK3 (9), was inhibited by prostratin (Fig. 3). Taken together, these data strongly
suggest that prostratin inhibits SIK3 mediated pro-cancer
signaling. However, more elaborate studies in the preclinical SIK3
knock-out animal models and detailed cell cycle studies would be
warranted to delineate the effect of prostratin on SIK3 mediated
signaling in solid tumors.
Prostratin had been shown to elicit anti-HIV effect
by inducing down-regulation of CD4, C-X-C chemokine receptor type 4
(CXCR4), and thereby protecting CD4+T cells from HIV-1
entry (12). In acutely infected
cells, prostratin is thought to enhance cellular protection
possibly due to cytostatic effects (27). In the context of cancer CXCR4 is
well-known to induce cancer cell migration and metastasis (24). Elevated membrane expression of CXCR4
is observed in several cancers. Further CXCR4 is positively
correlated with cancer progression and considered a poor prognostic
biomarker (28). Several factors
contribute to the upregulation of CXCR4 in malignant cells.
Previous studies in our laboratory have demonstrated that, under
high salt stimulating conditions, enhanced SIK3 signaling through
MMP-9 pathway mediates upregulation of CXCR4 membrane expression
(8). In our current studies we
demonstrate that prostratin inhibits CXCR4 expression in breast
cancer cell lines (Fig. 4).
Furthermore, siRNA mediated knockdown of SIK3 under high salt
stimulating culture conditions did not induce expression of CXCR4,
thus suggesting that, atleast that under our experimental
conditions, prostratin mediated CXCR4 inhibition is mediated by the
direct drug induced downregulation of SIK3.
In conclusion, we demonstrated a novel mechanism of
action for prostratin. SIK3 inhibition could be a novel area of
drug-target anti-cancer studies. In addition to PKC activation,
prostratin could exert its anti-cancer effect through inhibition of
SIK3. This data could provide a mechanistic basis for further
research to study the potential application of prostratin as an
add-on drug to the anti-cancer chemotherapeutic regimen.
Acknowledgements
This study was supported by NIH grant no.
2U54-CA163066-06 (cancer partnership grant). The authors would like
to thank Department of Biological Sciences, Tennessee State
University for their financial support.
Glossary
Abbreviations
Abbreviations:
|
SIK3
|
salt inducible kinase-3
|
|
IL
|
interleukin
|
|
NaCl
|
sodium chloride/salt
|
|
HIV
|
human immunodeficiency virus
|
|
CXCR4
|
C-X-C chemokine receptor type 4
|
References
|
1
|
Crusz SM and Balkwill FR: Inflammation and
cancer: Advances and new agents. Nat Rev Clin Oncol. 12:584–596.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Landskron G, De la Fuente M, Thuwajit P,
Thuwajit C and Hermoso MA: Chronic inflammation and cytokines in
the tumor microenvironment. J Immunol Res. 2014:1491852014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Amara S, Ivy MT, Myles EL and Tiriveedhi
V: Sodium channel γENaC mediates IL-17 synergized high salt induced
inflammatory stress in breast cancer cells. Cell Immunol. 302:1–10.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amara S, Zheng M and Tiriveedhi V:
Oleanolic acid inhibits high salt-induced exaggeration of
warburg-like metabolism in breast cancer cells. Cell Biochem
Biophys. 74:427–434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ouwerkerk R, Jacobs MA, Macura KJ, Wolff
AC, Stearns V, Mezban SD, Khouri NF, Bluemke DA and Bottomley PA:
Elevated tissue sodium concentration in malignant breast lesions
detected with non-invasive 23Na MRI. Breast Cancer Res Treat.
106:151–160. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zaric O, Pinker K, Zbyn S, Strasser B,
Robinson S, Minarikova L, Gruber S, Farr A, Singer C, Helbich TH,
et al: Quantitative sodium MR Imaging at 7 T: Initial results and
comparison with diffusion-weighted imaging in patients with breast
tumors. Radiology. 280:39–48. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amara S, Majors C, Roy B, Hill S, Rose KL,
Myles EL and Tiriveedhi V: Critical role of SIK3 in mediating high
salt and IL-17 synergy leading to breast cancer cell proliferation.
PLoS One. 12:e01800972017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Walkinshaw DR, Weist R, Kim GW, You L,
Xiao L, Nie J, Li CS, Zhao S, Xu M and Yang XJ: The tumor
suppressor kinase LKB1 activates the downstream kinases SIK2 and
SIK3 to stimulate nuclear export of class IIa histone deacetylases.
J Biol Chem. 288:9345–9362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gustafson KR, Cardellina JH II, McMahon
JB, Gulakowski RJ, Ishitoya J, Szallasi Z, Lewin NE, Blumberg PM,
Weislow OS, Beutler JA, et al: A nonpromoting phorbol from the
samoan medicinal plant homalanthus nutans inhibits cell killing by
HIV-1. J Med Chem. 35:1978–1986. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen X, Xiong GL, Jing Y, Xiao H, Cui Y,
Zhang YF, Shan YJ, Xing S, Yang M, Liu XL, et al: The protein
kinase C agonist prostratin induces differentiation of human
myeloid leukemia cells and enhances cellular differentiation by
chemotherapeutic agents. Cancer Lett. 356:686–696. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hezareh M, Moukil MA, Szanto I,
Pondarzewski M, Mouche S, Cherix N, Brown SJ, Carpentier JL and
Foti M: Mechanisms of HIV receptor and co-receptor down-regulation
by prostratin: Role of conventional and novel PKC isoforms. Antivir
Chem Chemother. 15:207–222. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dewan MZ, Ahmed S, Iwasaki Y, Ohba K, Toi
M and Yamamoto N: Stromal cell-derived factor-1 and CXCR4 receptor
interaction in tumor growth and metastasis of breast cancer. Biomed
Pharmacother. 60:273–276. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lapteva N, Yang AG, Sanders DE, Strube RW
and Chen SY: CXCR4 knockdown by small interfering RNA abrogates
breast tumor growth in vivo. Cancer Gene Ther. 12:84–89. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tiriveedhi V, Gelman AE and Mohanakumar T:
HIF-1α signaling by airway epithelial cell K-α1-tubulin: Role in
fibrosis and chronic rejection of human lung allografts. Cell
Immunol. 273:59–66. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Amara S, Lopez K, Banan B, Brown SK,
Whalen M, Myles E, Ivy MT, Johnson T, Schey KL and Tiriveedhi V:
Synergistic effect of pro-inflammatory TNFα and IL-17 in periostin
mediated collagen deposition: Potential role in liver fibrosis. Mol
Immunol. 64:26–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tiriveedhi V, Tucker N, Herndon J, Li L,
Sturmoski M, Ellis M, Ma C, Naughton M, Lockhart AC, Gao F, et al:
Safety and preliminary evidence of biologic efficacy of a
mammaglobin-a DNA vaccine in patients with stable metastatic breast
cancer. Clin Cancer Res. 20:5964–5975. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tiriveedhi V, Angaswamy N, Brand D, Weber
J, Gelman AG, Hachem R, Trulock EP, Meyers B, Patterson G and
Mohanakumar T: A shift in the collagen V antigenic epitope leads to
T helper phenotype switch and immune response to self-antigen
leading to chronic lung allograft rejection. Clin Exp Immunol.
167:158–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sarma NJ, Tiriveedhi V, Crippin JS,
Chapman WC and Mohanakumar T: Hepatitis C virus-induced changes in
microRNA 107 (miRNA-107) and miRNA-449a modulate CCL2 by targeting
the interleukin-6 receptor complex in hepatitis. J Virol.
88:3733–3743. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tiriveedhi V, Takenaka M, Ramachandran S,
Gelman AE, Subramanian V, Patterson GA and Mohanakumar T: T
regulatory cells play a significant role in modulating MHC class I
antibody-induced obliterative airway disease. Am J Transplant.
12:2663–2674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tiriveedhi V, Takenaka M, Sarma NJ, Gelman
AG and Mohanakumar T: Anti-major histocompatibility complex-induced
obliterative airway disease: Selective role for CD4 and CD8 T cells
in inducing immune responses to self-antigens. J Heart Lung
Transplant. 32:714–722. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Amara S, Alotaibi D and Tiriveedhi V:
NFAT5/STAT3 interaction mediates synergism of high salt with IL-17
towards induction of VEGF-A expression in breast cancer cells.
Oncol Lett. 12:933–943. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Platt D, Amara S, Mehta T, Vercuyssee K,
Myles EL, Johnson T and Tiriveedhi V: Violacein inhibits matrix
metalloproteinase mediated CXCR4 expression: Potential anti-tumor
effect in cancer invasion and metastasis. Biochem Biophys Res
Commun. 455:107–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu C, Zhao H, Chen H and Yao Q: CXCR4 in
breast cancer: Oncogenic role and therapeutic targeting. Drug Des
Devel Ther. 9:4953–4964. 2015.PubMed/NCBI
|
|
25
|
Szallasi Z, Krsmanovic L and Blumberg PM:
Nonpromoting 12-deoxyphorbol 13-esters inhibit phorbol 12-myristate
13-acetate induced tumor promotion in CD-1 mouse skin. Cancer Res.
53:2507–2512. 1993.PubMed/NCBI
|
|
26
|
Wang MT, Holderfield M, Galeas J,
Delrosario R, To MD, Balmain A and McCormick F: K-Ras Promotes
Tumorigenicity through Suppression of Non-canonical Wnt Signaling.
Cell. 163:1237–1251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Biancotto A, Grivel JC, Gondois-Rey F,
Bettendroffer L, Vigne R, Brown S, Margolis LB and Hirsch I: Dual
role of prostratin in inhibition of infection and reactivation of
human immunodeficiency virus from latency in primary blood
lymphocytes and lymphoid tissue. J Virol. 78:10507–10515. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hiller D and Chu QD: CXCR4 and axillary
lymph nodes: Review of a potential biomarker for breast cancer
metastasis. Int J Breast Cancer. 2011:4209812011. View Article : Google Scholar : PubMed/NCBI
|