Introduction
Gallbladder cancer is a relatively rare disease,
with an estimated annual incidence of between 2 and 3
individuals/100,000; however, it is the most frequently encountered
malignancy of the biliary tract (1,2). Complete
surgical resection is the only potentially curative approach in
early stages of the disease; however, the majority of cases are
diagnosed in advanced stages. Additionally, because the aggressive
malignancy tolerates traditional chemotherapy and radiotherapy,
these cases have a dismal prognosis with an overall 5-year survival
rate of <5% (3). Therefore,
improving the diagnostic accuracy during the early stages and
identifying the molecular mechanisms associated with chemotherapy
or radiation resistance may provide a potential therapeutic
approach for the treatment of gallbladder cancer.
Cyclin M, also known as FAM58A, is a member of the
cyclin family, which serves important regulatory functions in the
cell cycle and transcription (4).
Mutations in cyclin M cause toe syndactyly, telecanthus and
anogenital and renal malformations syndrome, a human developmental
anomaly (4,5). However, the function of cyclin M in
other diseases remains unclear. Cyclin M has been identified as the
binding partner for cyclin-dependent kinase 10 (CDK10) (5). CDK10 is involved in the regulation of
cell division and function in various types of tumor (6–10). It was
confirmed that CDK10 functions as a tumor suppressor gene and
regulates the survivability of biliary tract cancer cells,
including gallbladder cancer cells (8). These results suggest that cyclin M may
also be involved in cancer development through binding to
CDK10.
MicroRNAs (miRNAs) are key negative regulators of
gene expression in eukaryotes and abundant evidence indicates that
miRNAs are involved in human cancers (11,12).
miR-433 has been identified as a cancer-associated miRNA in various
types of tumor, including bladder cancer, ovarian cancer, gastric
cancer, hepatocellular carcinoma and oral squamous cell carcinoma
(13–17). Furthermore, aberrant expression levels
of miR-433 have been identified to be associated with
chemosensitivity and enhanced survival of ovarian cancer cells
(15,18).
In the present study, gemcitabine-resistant (GR)
subclones were established on the basis of stable transfectants of
CDK10 or cyclin M overexpression. To provide a possible mechanism
for cyclin M downregulation in acquired chemoresistance of
gallbladder cancer cells, miR-433 was predicted to target cyclin M.
Furthermore, the association of serum levels of miR-433 and the
chemotherapeutic response in gallbladder cancer was evaluated.
Collectively, the results of the present study suggest that
aberrant expression of miR-433 affects the chemoresistance of
gallbladder cancer cells by targeting cyclin M.
Materials and methods
Cell culture, plasmids, RNA
oligonucleotides and transfection
The GBC-SD gallbladder carcinoma cell line was
obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). Cells were cultured
at 37°C in 5% CO2 in RPMI-1640 medium with 10% fetal
bovine serum (both from Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), 100 IU/ml penicillin and 100 µg/ml streptomycin.
Human CDK10 and cyclin M coding sequences were cloned into
pBudCE4.1 vector (Invitrogen; Thermo Fisher Scientific, Inc.).
Total RNA was isolated from normal liver tissue using TRIzol
reagent (Thermo Fisher Scientific, Inc.) and cDNA was synthesized
using the PrimeScript reagent kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
instructions. The primer sequences for PCR amplification of
CDK10-open reading frame (ORF) were as follows:
5′-CCAAGCTTATGGCGGAGCCAGATCTGGA-3′ (forward) and
5′-CCGGATCCTCAGGGTTTACAGCGCTTG-3′ (reverse). The PCR products were
purified and cloned into the HindIII/BamHI sites of
the vector. The primer sequences for PCR amplification of cyclin
M-ORF were as follows: 5′-GGGTACCATGGAAGCCCCGGAGGG-3′ (forward) and
5′-CCCTCGAGTTAGGGGATCTCTGTGTCCA-3′ (reverse). The PCR products were
purified and cloned into the KpnI/XhoI sites of the
vector. CDK10-ORF was cloned into the pBudCE4.1 vector and cyclin
M-ORF was cloned into the pBudCE4.1 vector to produce
pBudCE4.1-CDK10 and pBudCE4.1-CycM, respectively. Finally,
CDK10-ORF was cloned into the pBudCE4.1-CycM vector and named
pBudCE4.1-Co. All the constructs were confirmed by DNA sequencing
and the pBudCE4.1 empty vector was used as a vector control.
GBC-SD cells were transfected with pBudCE4.1-CDK10,
pBudCE4.1-CycM, pBudCE4.1-Co or empty vector plasmid using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). At 48 h post-transfection, the culture medium
was replaced with medium containing Zeocin (Invitrogen; Thermo
Fisher Scientific, Inc.) and the medium containing Zeocin was
replaced every 3–4 days. After 2 weeks, isolated colonies began to
appear. After 3 weeks, stable transfectants expressing CDK10 or
cyclin M were selected for further study. The control clones
expressing the empty vector were isolated at the same time. The
stable transfectants were named as follows: GBC-Mock, GBC-CDK10,
GBC-CycM and GBC-Co, respectively.
Small interfering RNA (siRNA) targeting the
proto-oncogene serine/threonine-protein kinase c-Raf was obtained
from RiboBio (Guangzhou, China) and the sequence was
5′-GCUGCAUCUCUCCUACAAU-3′. Control siRNA was obtained from RiboBio
(8). Other RNA oligonucleotides
(including miR-433, anti-miR-433, miR-control and siRNA-control
groups) were also obtained from RiboBio. At 24 h before
transfection, cells were plated onto a 35-mm dish at 50–60%
confluence. RNA oligonucleotides transfection was performed at 50
nM using Lipofectamine® 2000, according to the
manufacturer's instructions. The transfected cells were resuspended
in RPMI-1640 medium and cultured for 48–72 h before analysis. Each
assay was performed in triplicate at least.
Establishment of GR subclones
Stable transfectants were exposed to increasing
concentrations (0.001, 0.0025, 0.005, 0.01, 0.025, 0.05, 0.1, 0.25,
0.3, 0.35, 0.4, 0.45 and 0.5 µg/ml) of gemcitabine (Lilly France
S.A., Lilly, Fegersheim, France) every 4 weeks over a 9-month
period. As cells adapted to the doses, the gemcitabine
concentration was increased. The cell subclones were named as
follows: GBC-Mock-GR, GBC-CDK10-GR, GBC-CycM-GR and GBC-Co-GR.
Western blot analysis
Total proteins were extracted from tissue samples or
cells using lysis buffer containing phenylmethyl sulfonylfluoride
(both from Beyotime Institute of Biotechnology, Haimen, China) at
25°C and protein concentration was determined by BCA Protein Assay
kit (Beyotime Institute of Biotechnology). A total of 20 µg protein
was separated by SDS-PAGE (10% gel) and transferred onto a
polyvinylidene fluoride membrane. Membranes were blocked with 5%
non-fat milk and incubated with anti-CDK10 (cat.no. 17182-1-AP,
1:500; ProteinTech Group, Inc., Chicago, IL, USA), anti-c-Raf (cat.
no. AP7816d, 1:500; Abgent, Inc., San Diego, CA, USA,) and
anti-cyclin M (cat. no. ab81062, 1:1,000; Abcam, Cambridge, UK)
primary antibodies at 4°C for 12 h. Anti-β-actin (cat. no.
sc-47778, 1:1,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
was used as a loading control. Membranes were then washed and
incubated with horseradish peroxidase-conjugated secondary antibody
(cat. no. A21020 and A21010, 1:8,000; Abbkine, Wuhan, China).
Immunoreactive bands were visualized using a chemiluminescence
solution with an ECL kit (Beyotime Institute of Biotechnology).
Quantity One (Bio-Rad Laboratories, Inc., Hercules, CA, USA) was
used as the software for quantification.
Cell counting Kit-8 (CCK-8) assay
Cells were seeded into a 96-well plate at a density
of 4×103 cells/well, and allowed to attach at 37°C
overnight. Cells were treated with various concentrations (0, 0.01,
0.05, 0.1, 0.5, 1 and 10 µg/ml) of gemcitabine in ≥6 replicate
wells and incubated at 37°C for 48 h. Cell proliferation was
determined using the CCK-8 assay (Beyotime Institute of
Biotechnology). In brief, a 1/10 volume of the CCK-8 solution was
added to each well, and the cells were cultured for another 2 h.
Cell viability was determined by measuring the absorbance at 450 nm
using a plate reader (BioTek Instruments, Inc., Winooski, VT,
USA).
Colony formation assay
Cells were seeded for colony formation in 24-well
dishes at a density of 500 viable cells/well. The medium contained
1 µg/ml gemcitabine and was exchanged every 2 days. After 14 days,
the cells were fixed in 4% paraformaldehyde for 15 min. Following
washing, the cells were stained with 0.005% crystal violet solution
for 1 h. The plates were aspirated, washed and allowed to air dry.
The dishes were photographed with a camera and the colonies were
counted (a single colony contained >20 cells) using a light
microscope (magnification, ×40. Each assay was performed in
triplicate.
Clinical tissue and blood samples
Gallbladder cancer tissue and paired normal tissue
specimens were obtained from 8 patients at the Shaoxing People's
Hospital (Shaoxing, China) from September 2012 to March 2016. All
participants provided written informed consent and the tissue
acquisition protocol was approved by the Zhejiang University
Institutional Review Board (Hangzhou, China). Blood samples were
collected from 23 patients with gallbladder cancer and the survival
time was >3 months. Among the patients, 17 patients underwent
chemotherapy with gemcitabine or 5-fluorouracil and were assigned
to the chemotherapy group. The remaining 6 patients did not receive
chemotherapy and were assigned to the non-chemotherapy group. Blood
samples from the patients were collected and serum was separated by
centrifugation (1,000 × g, for 5 min at 4°C) collected into
RNase/DNase-free tubes and stored at −80°C until use. Fresh tissues
were stored at −80°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from cells, tissues or serum
using TRIzol reagent, and RNA was reverse-transcribed into cDNA
using a High Capacity cDNA Reverse Transcription kit (Takara
Biotechnology Co., Ltd., Dalian, China), according to the
manufacturer's instructions. qPCR was performed using SYBR Green
PCR Master Mix (Takara Biotechnology Co., Ltd.) and the ABI 7500
Real-time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions. Specific
primers were designed for cyclin M, CDK10 and β-actin (Table I). The PCR conditions included 95°C
for 5 min, and a total of 40 cycles of 95, 60 and 72°C for 30 sec
and then a final extension at 72°C for 5 min. β-actin was used as
an endogenous control. miR-433 normalization was performed
according to a protocol described previously (19). Expression of miR-433 was normalized
using U6 RNA (for RNA from cells and tissues) and with
Caenorhabditis elegans miR-39 mimic (for RNA from culture
supernatant and serum samples) which was used as an RNA spike-in.
Specific primers for miR-433, U6 and C. elegans miR-39 RNA
were obtained from RiboBio (15,19). In
total, three individual experiments were performed. Relative
expression levels of miR-433 were determined using the
2−ΔΔCq method (20).
 | Table I.Primers designed for cyclin M, CDK10
and β-actin. |
Table I.
Primers designed for cyclin M, CDK10
and β-actin.
| Name | Symbol | Forward (5′-3′) | Reverse (5′-3′) |
|---|
| β-actin | ACTB |
AGAAGGAGATCACTGCCCTGGCACC |
CCTGCTTGCTGATCCACATCTGCTG |
| CDK10-ORF | CDK10 |
TGGACAAGGAGAAGGATG |
CTGCTCACAGTAACCCATC |
| CDK10-3′UTR | CDK10 |
GTGCAACCACTGCTTCTTGG |
AGCGTCCACCACAACTCAAA |
| Cyclin M-ORF | FAM58A |
ACTTCCGAGTGGCGAGGTTCAT |
TCATAGGCGTCCAGGTTGGTCT |
| Cyclin M-3′UTR | FAM58A |
CGCTTTGCCAGTCCCTAGAA |
GACAAACGCAGATGTGGCTG |
Dual-luciferase reporter assay
Predictions for the potential binding sites of
miR-433 in the 3′UTR of cyclin M was performed using 2 different
online programs: TargetScan 6.2 (http://www.targetscan.org/vert_61/) and microRNA.org (http://www.microrna.org). The 3′UTR of cyclin M was
amplified by PCR using human genomic DNA as a template. The primers
used were as follows: 5′-CGACGCGTTTGACTACAATACAGGCATGACA-3′
(forward) and 5′-CCAAGCTTTCCGTAGGGATTCAGTCTCA-3′ (reverse). The PCR
product, which contained a potential binding site for miR-433, was
digested with MluI and HindIII, and was inserted
downstream of the luciferase gene in the pMIR-REPORT system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
constructs were verified by DNA sequencing. The verified construct
was named pMIR-CycM-3′UTR. The mutant reporter plasmids were
constructed in accordance with the aforementioned criteria by
annealing the following oligonucleotides:
5′-CGACGCGTTTGACTACAATACAGGCATGACATCAATGAAAGGAAAGTCcatAAATCGATGAGACTGAATCCCTACGGAAAGCTTGG-3′
and
5′-CCAAGCTTTCCGTAGGGATTCAGTCTCATCGATTTatgGACTTTCCTTTCATTGATGTCATGCCTGTATTGTAGTCAAACGCGTCG-3′
(the bases in lower-case indicate the site mutations). The mutant
reporter plasmids were named mut-pMIR-CycM-3′UTR. For luciferase
assays, 293T cells were cultured in 6-well plates and transfected
with plasmids (including pMIR-CycM-3′UTR, mut-pMIR-CycM-3′UTR and
β-galactosidase reporter plasmids) and RNA oligonucleotides using
Lipofectamine® 2000. A β-galactosidase reporter plasmid
was designed to normalize transfection efficiency. At 48 h after
transfection, the cells were lysed, luciferase activity was
determined using a Dual-Luciferase Reporter assay system and
β-galactosidase activity was determined using the β-Galactosidase
Enzyme assay system (both from Promega Corporation, Madison, WI,
USA).
Statistical analysis
Data were analyzed using SPSS software (version
13.0; SPSS, Inc., Chicago, IL, USA). The relevant data are
expressed as the mean ± standard deviation (SD). Statistical
significance between two groups was determined using Student's
t-test. One-way analysis of variance followed by a Tukey-Kramer
test was used to examine differences among multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Decreasing expression of cyclin M
contributes to acquired chemoresistance of GR subclones
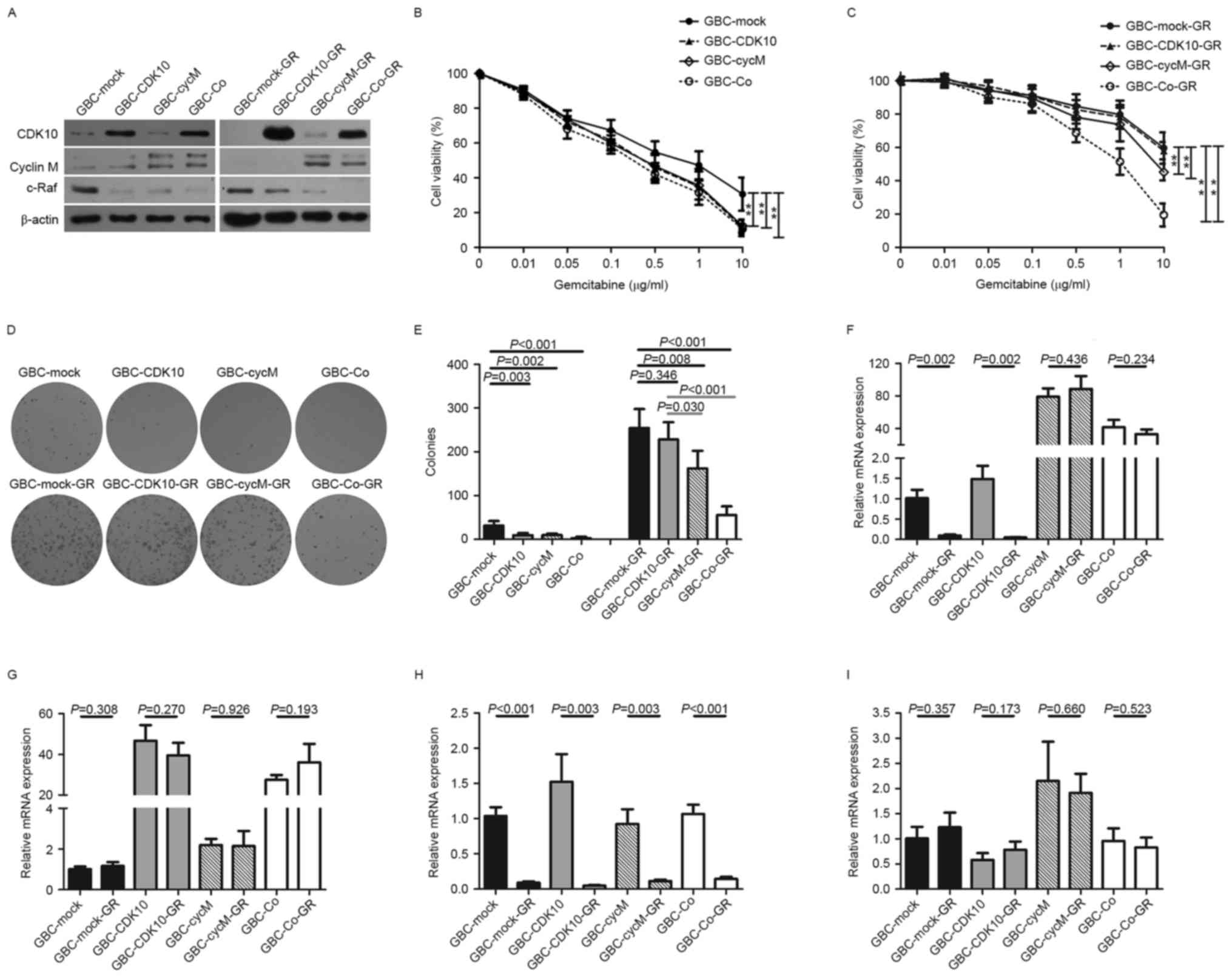
To generate stable cell lines for experimental
analyses, GBC-SD cells were transfected with pBudCE4.1-CDK10,
pBudCE4.1-CycM, pBudCE4.1-Co or with the empty vector plasmid.
After 4 weeks, stable transfectants were obtained and named
GBC-CDK10, GBC-CycM, GBC-Co and GBC-Mock, respectively. Western
blot analysis was performed to confirm positive clones (Fig. 1A). Next, the chemosensitivity of the
four stable cell lines to gemcitabine was examined. The results
demonstrated that GBC-CDK10 and GBC-CycM cells exhibited a
significantly increased sensitivity to gemcitabine (Fig. 1B). As expected, GBC-Co cells were also
more sensitive to gemcitabine than were GBC-Mock cells (Fig. 1B).
Next, the four stable cell lines (GBC-CDK10,
GBC-CycM, GBC-Co and GBC-Mock) were cultured for 9 months in the
presence of gemcitabine to establish GR subclones (Fig. 1B and C). The dose-response curves
indicated that GBC-CDK10-GR cells exhibited a similar
chemosensitivity to that of the GBC-Mock-GR cells and were more
resistant to gemcitabine compared with the GBC-Co-GR or GBC-CycM-GR
cells (Fig. 1C). Following extended
exposure to gemcitabine, although GBC-CDK10-GR cells maintained
high levels of CDK10 expression (Fig.
1A), GBC-CDK10-GR cells did not exhibit the increased
sensitivity to gemcitabine that was exhibited by GBC-CDK10 cells
(Fig. 1C).
Additionally, colony formation assays were performed
in all four-cell lines, which were cultured with 1 µg/ml
gemcitabine. After 14 days, GBC-CDK10, GBC-CycM and GBC-Co cells
were more sensitive to gemcitabine compared with GBC-Mock cells,
and GBC-Co-GR cells were more sensitive to gemcitabine compared
with GBC-Mock-GR and GBC-CDK10-GR cells (Fig. 1D and E). These results confirm the
results obtained from the dose-response curves (Fig. 1B and C).
Cyclin M is a binding partner for CDK10 (5). It was demonstrated that cyclin M protein
levels were too low to detect by western blot in the GBC-CDK10-GR
cells (Fig. 1A), different from the
GBC-CDK10 cells. It suggested that decreasing expression of cyclin
M may contribute to the decreased chemosensitivity of
CDK10-overexpressing GBC-SD cells.
Because of the transfection, both the transcripts
from plasmid and genome could influence the protein levels of CDK10
and cyclin M. The exogenous transcription product (from plasmid)
could cover up the practical change of endogenous expression.
Considering that only the open reading frames (ORFs), without
3′UTRs, were cloned into plasmid in our study, the expression of
the 3′UTRs may more accurately reflect the endogenous expression of
CDK10 and cyclin M (Fig. 1F-G). To
perform PCR, specific primers for the 3′UTRs of CDK10 and cyclin M
were designed. The results demonstrated that the 3′UTR of cyclin M
was significantly decreased in GR subclones (Fig. 1H). However, the 3′UTR of CDK10 did not
exhibit a similar change (Fig. 1I).
This result indicates that downregulating the expression of cyclin
M may be an important mechanism for the acquired resistance in GR
subclones.
Cyclin M is a target of miR-433 in
gallbladder cancer cells
The interactions between miRNAs and the 3′UTRs of
target genes are key regulatory mechanisms that are involved in
multiple biological processes and human diseases, including cancer
(21–23). To examine the molecular mechanism
underlying cyclin M downregulation in gallbladder cancer cells,
miRNAs targeting cyclin M using the TargetScan 6.2 and microRNA.org software were employed. Considering the
association between candidate miRNAs and chemoresistance (15,18),
miR-433 was hypothesized to act as a candidate target for cyclin M.
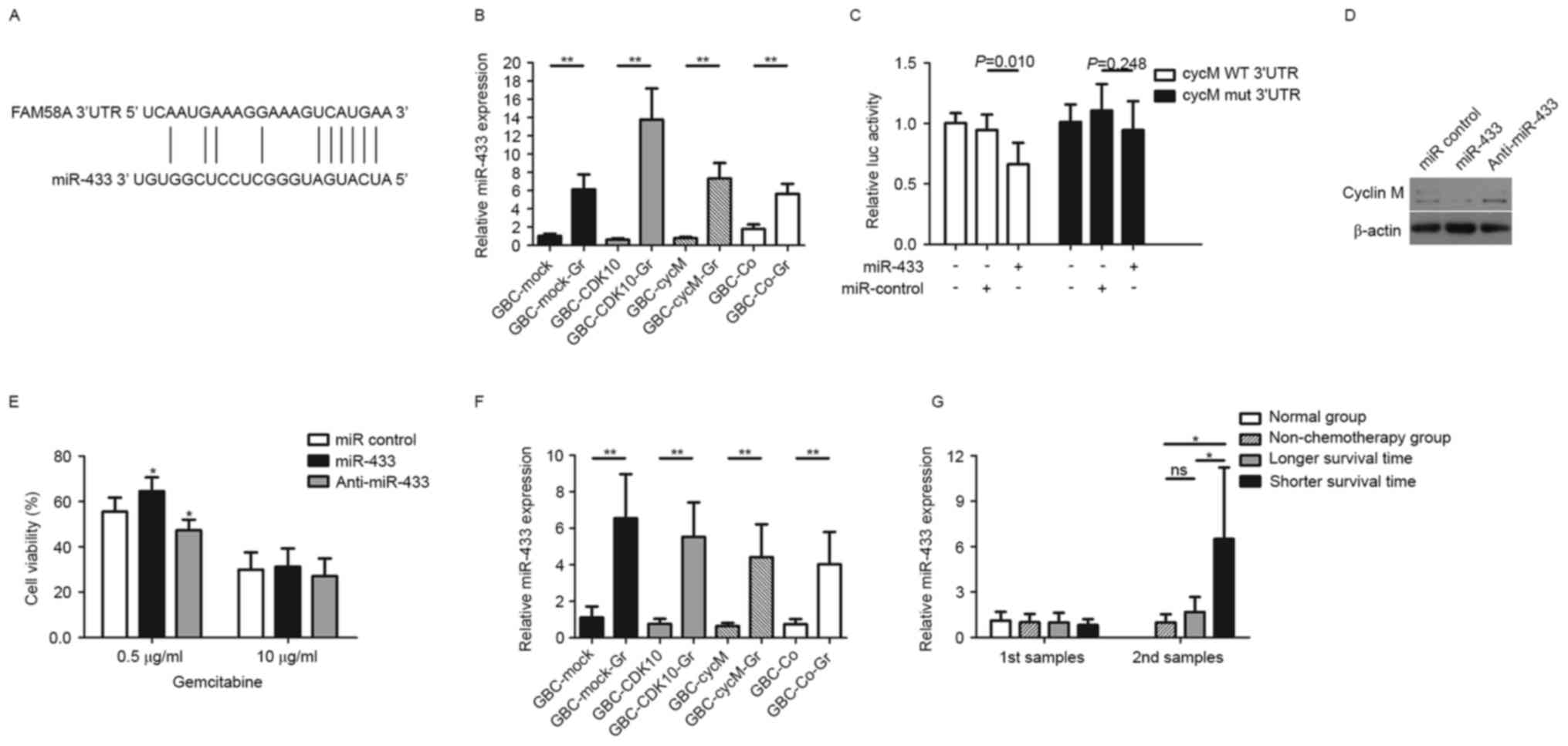
miR-433-binding sequences in the 3′UTR of cyclin M mRNA are
indicated in Fig. 2A. Examination of
miR-433 expression levels in the GBC subclones revealed that all
four GR subclones, i.e., GBC-Mock-GR, GBC-CDK10-GR, GBC-CycM-GR and
GBC-Co-GR, expressed significantly increased levels of miR-433
compared with their respective non-GR subclones (Fig. 2B).
 | Figure 2.miR-433 accelerates acquired
chemoresistance of gallbladder cancer cells by targeting cyclin M.
(A) Putative miR-433-binding sequences in the 3′UTR of cyclin M
(FAM58A) mRNA. (B) Relative expression of miR-433 in GR and non-GR
subclones. (C) Relative luciferase activity in CycM WT 3′UTR (cells
were transfected with pMIR-CycM-3′UTR) and CycM mut 3′UTR (cells
were transfected with mut-pMIR-CycM-3′UTR)-293T cells.
β-galactosidase activity was used as a normalization control. (D)
Western blot analysis of cyclin M expression in GBC-SD cells
following transfection with miR-433 or anti-miR-433. β-actin was
used as a loading control. (E) Cell viability (expressed as a
percentage) of GBC-SD cells following transfection with miR-433 or
anti-miR-433 in response to 0.5 and 1 µg/ml gemcitabine. (F)
Relative expression of miR-433 in supernatants from GR and non-GR
subclones. (G) Relative expression of miR-433 in serum samples.
Patients in the longer survival time group and shorter survival
time group underwent chemotherapy with gemcitabine or
5-fluorouracil. The first samples were collected from the patients
prior to receiving chemotherapy and the second samples were
collected following three chemotherapy cycles. *P<0.05;
**P<0.01. GR, gemcitabine-resistant; CDK10, cyclin-dependent
kinase 10; 3′UTR, 3′ untranslated region; miR, microRNA; luc,
luciferase; GBC, gallbladder cancer; CycM, cyclin M; WT, wild-type;
mut, mutant; Co, transfected with the plasmids which contained
CDK10-ORF and CycM-ORF. |
Luciferase activity assays indicated that the
miR-433 mimic repressed the luciferase activity of pMIR-CycM-3′UTR
reporter plasmid, but did not repress the activity of
mut-pMIR-CycM-3′UTR (Fig. 2C). The
miR-control had no impact on the luciferase activity of
pMIR-CycM-3′UTR (Fig. 2C). To further
confirm the results, cyclin M expression levels were investigated
following transfection with miR-433 in GBC-SD cells. Western blot
analysis was performed at 72 h post-transfection. It was revealed
that cyclin M was downregulated in GBC-SD cells following miR-433
transfection, whereas cyclin M was upregulated following
transfection with anti-miR-433 (Fig.
2D).
miR-433 expression enhances the
chemoresistance of gallbladder cancer cells
Considering that miR-433 was expressed at a high
level in GR subclones and targets cyclin M, the function of miR-433
in promoting resistance to gemcitabine in GBC-SD cells was examined
in vitro. The results indicated that the miR-433 mimic
enhanced the resistance of GBC-SD cells in response to a moderate
dose of gemcitabine (0.5 µg/ml), whereas anti-miR-433 treatment
reversed this change (Fig. 2E).
However, in response to a high gemcitabine dose (10 µg/ml), the
function of the miR-433 mimic or inhibitor did not induce any
significant effect.
Circulating miR-433 levels are
associated with chemotherapy in gallbladder cancer
A previous study demonstrated that ovarian cancer
cells with increased miR-433 expression are able to release miR-433
into the growth medium (15).
Therefore, the relative miR-433 expression was examined in the
culture supernatant of the subclones. The results indicated that
the culture supernatant from GR subclones exhibited significantly
increased relative miR-433 expression compared with that from
non-GR subclones (Fig. 2F).
Serum expression levels of miR-433 were examined in
patients with gallbladder cancer, from the chemotherapy group and
the non-chemotherapy group. The chemotherapy group comprised 17
patients with a median survival time of 9.8 months. Among these
patients, 9 patients had a shorter survival time compared with the
median (shorter survival time group, as presented in Fig. 2G) and the remaining 8 patients had a
longer survival time compared with the median (longer survival time
group, as presented in Fig. 2G). The
first serum samples were collected from the patients prior to
receiving chemotherapy. The results indicated no marked difference
in miR-433 levels between the non-chemotherapy group and
chemotherapy group, regardless of shorter or longer survival time
(Fig. 2G). Following three
chemotherapy cycles, additional serum samples were collected.
Interestingly, the patients in the chemotherapy group with a
shorter survival time exhibited significantly increased levels of
miR-433 in the serum, compared with the patients in the
non-chemotherapy group or with the patients in the chemotherapy
group with a longer survival time (Fig.
2G). However, although they underwent chemotherapy, the
patients with a longer survival time did not exhibit higher miR-433
levels in the serum compared with those in the non-chemotherapy
group.
Increased expression of c-Raf induced
by dysfunction of CDK10-cyclin M increases chemoresistance in
gallbladder cancer cells
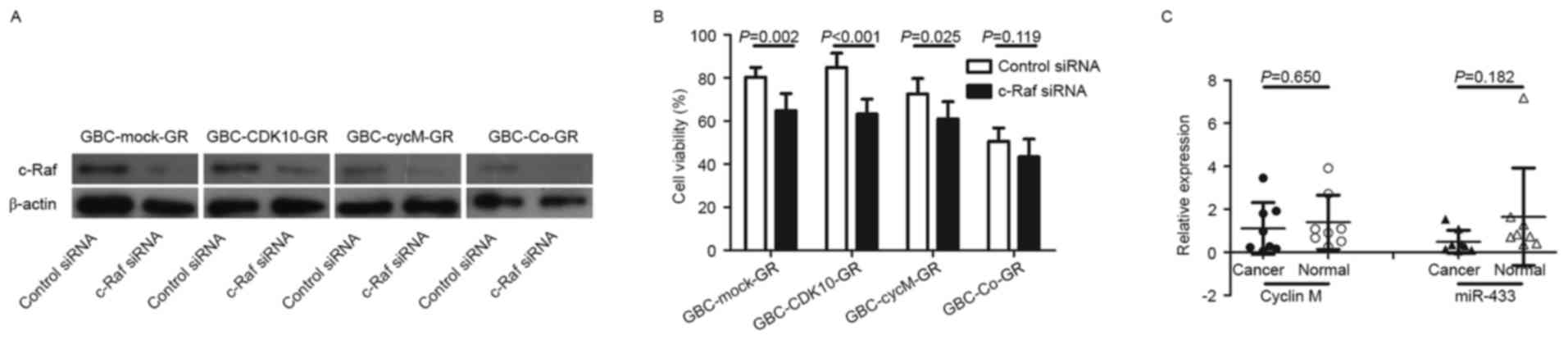
Considering that the CDK10-cyclin M complex
regulates the expression of c-Raf (5,6), c-Raf
protein levels were examined prior to and following the
establishment of GR subclones. The results demonstrated that c-Raf
protein expression was markedly increased in cells with decreased
expression of CDK10 or decreased expression of cyclin M (Fig. 1E). However, the expression of c-Raf
was at low levels under conditions of increased expression of CDK10
and cyclin M, such as in the case of GBC-Co cells and GBC-Co-GR
cells (Fig. 1E).
Subsequently, the involvement of c-Raf to
chemoresistance was assessed. All GR subclones, including
GBC-Mock-GR, GBC-CDK10-GR, GBC-CycM-GR and GBC-Co-GR, were
transfected with siRNA against c-Raf and c-Raf downregulation was
confirmed by western blot analysis (Fig.
3A). The results demonstrated that c-Raf silencing
significantly increased the sensitivity to gemcitabine in
GBC-Mock-GR, GBC-CDK10-GR and GBC-CycM-GR cells (Fig. 3B). Therefore, increased expression of
c-Raf is also an important factor that is involved in the
regulation of chemoresistance in cancer.
Discussion
Resistance to chemotherapy is a major concern in
gallbladder cancer (3). Acquired
chemoresistance of cancer cells may arise following extended
exposure to chemotherapy drugs. The majority of patients with
gallbladder cancer that undergo chemotherapy become resistant
following consecutive treatments and fail to benefit from
chemotherapy (24). Therefore, it is
important to clarify the molecular mechanism underlying acquired
chemoresistance for an improved treatment of gallbladder
cancer.
Gemcitabine is a deoxycytidine analogue with
antitumor activity and a key drug for chemotherapy of biliary tract
cancer (25,26). To investigate the acquired
chemoresistance of gallbladder cancer cells, GR subclones were
established. Drug concentrations were gradually increased to
establish subclones of GBC-SD cells that were able to grow in
medium with 0.5 µg/ml gemcitabine. This strategy may simulate the
process of acquired chemoresistance. Although CCK-8 or MTT assays
are used to examine sensitivity to chemotherapy during a relatively
short period (48 or 72 h), a colony formation assay is able to
accurately determine chemosensitivity and colony forming ability
under extended exposure to gemcitabine (2 weeks).
Similar to a previous study (8), the results of the present study
confirmed that GBC-CDK10 cells were more sensitive to gemcitabine
compared with GBC-Mock cells. Although GBC-CDK10-GR and GBC-CDK10
cells expressed high levels of CDK10, the gemcitabine resistance of
GBC-CDK10-GR cells was as high as that of GBC-Mock-GR cells in
contrast with that of GBC-CDK10 cells. This indicates that the
expression level of CDK10 alone was not able to explain the
acquired gemcitabine resistance of GBC-SD cells. However, GBC-Co-GR
cells and GBC-Co cells, which expressed increased CDK10 and cyclin
M, exhibited a relatively increased sensitivity to gemcitabine
compared with that of GBC-Mock-GR and GBC-Mock cells. Additionally,
the levels of cyclin M mRNA that contained the 3′UTR were
significantly downregulated in GR subclones, compared with that
observed in the non-GR subclones This suggests that cyclin M serves
an important function in the acquired gemcitabine resistance of
gallbladder cancer cells.
miRNAs regulate the expression of target genes by
interacting with the 3′UTRs of the mRNA. The results of the present
study demonstrated that the 3′UTR of cyclin M was downregulated in
all GR subclones and miR-433 was identified as a predicted miRNA to
target cyclin M. Overexpression of miR-433 was demonstrated to
promote resistance to paclitaxel in ovarian cancer cells (15). The results of the present study also
demonstrated that the acquired gemcitabine resistance of
gallbladder cancer cells was associated with an upregulation of
miR-433. However, other studies reported that miR-433 functions as
an anti-oncomiR in various tumors by regulating various targets
(13,14,27,28). The
results of the present study demonstrated that the expression of
miR-433 was decreased in the tumor samples, but this decrease was
not statistically significant. Furthermore, the expression of
cyclin M between tumor samples and normal tissues also did not
exhibit a significant difference. The expression of miR-433 in
serum samples was also examined. No differences in the expression
of miR-433 were identified in the serum of normal volunteers and
patients with gallbladder cancer. Nevertheless, the patients with
poor response to chemotherapy exhibited a significant increase in
the expression of serum miR-433, whereas the patients with an
improved response did not exhibit changes in the serum levels of
miR-433. These observations indicate that circulating miR-433
expression may not be useful as a biomarker for diagnosis, but it
may be a potential biomarker to evaluate the response to
chemotherapy.
The CDK10-cyclin M complex is a negative regulator
of c-Raf (5,6). The results of the present study also
identified that downregulated cyclin M expression or downregulated
CDK10 expression resulted in increasing c-Raf expression.
Considering that c-Raf silencing reversed the resistance to
gemcitabine in GBC-Mock-GR, GBC-CDK10-GR and GBC-CycM-GR cells, the
increasing gemcitabine resistance of GBC-SD cells may be attributed
to upregulated c-Raf expression, which was induced by
downregulation of cyclin M expression.
In conclusion, the results of the present study
suggest that the miR-433/cyclin M axis is associated with the
acquired chemoresistance of gallbladder cancer cells. Circulating
miR-433 expression represents a potential biomarker for evaluating
the chemosensitivity of gallbladder cancer cells.
Acknowledgements
The present study was supported by Zhejiang
Provincial Natural Science Foundation of China (grant no.
LQ14H160001), National Natural Science Foundation of China (grant
no. 81602044), Zhejiang Provincial Public Welfare Technology
Application Research Projects (grant nos. 2013C33214 and
2015C33293) and Public Welfare Technology Application Research
Projects of Shaoxing (grant no. 2014B70073).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lai CH and Lau WY: Gallbladder cancer - a
comprehensive review. Surgeon. 6:101–110. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goetze TO: Gallbladder carcinoma:
Prognostic factors and therapeutic options. World J Gastroenterol.
21:12211–12217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Unger S, Böhm D, Kaiser FJ, Kaulfuss S,
Borozdin W, Buiting K, Burfeind P, Böhm J, Barrionuevo F, Craig A,
et al: Mutations in the cyclin family member FAM58A cause an
X-linked dominant disorder characterized by syndactyly, telecanthus
and anogenital and renal malformations. Nat Genet. 40:287–289.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guen VJ, Gamble C, Flajolet M, Unger S,
Thollet A, Ferandin Y, Superti-Furga A, Cohen PA, Meijer L and
Colas P: CDK10/cyclin M is a protein kinase that controls ETS2
degradation and is deficient in STAR syndrome. Proc Natl Acad Sci
USA. 110:pp. 19525–19530. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iorns E, Turner NC, Elliott R, Syed N,
Garrone O, Gasco M, Tutt AN, Crook T, Lord CJ and Ashworth A:
Identification of CDK10 as an important determinant of resistance
to endocrine therapy for breast cancer. Cancer Cell. 13:91–104.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leman ES, Magheli A, Yong KM, Netto G,
Hinz S and Getzenberg RH: Identification of nuclear structural
protein alterations associated with seminomas. J Cell Biochem.
108:1274–1279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yu JH, Zhong XY, Zhang WG, Wang ZD, Dong
Q, Tai S, Li H and Cui YF: CDK10 functions as a tumor suppressor
gene and regulates survivability of biliary tract cancer cells.
Oncol Rep. 27:1266–1276. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhong XY, Xu XX, Yu JH, Jiang GX, Yu Y,
Tai S, Wang ZD and Cui YF: Clinical and biological significance of
Cdk10 in hepatocellular carcinoma. Gene. 498:68–74. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kasten M and Giordano A: Cdk10, a
Cdc2-related kinase, associates with the Ets2 transcription factor
and modulates its transactivation activity. Oncogene. 20:1832–1838.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee RC and Ambros V: An extensive class of
small RNAs in Caenorhabditis elegans. Science. 294:862–864. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yates LA, Norbury CJ and Gilbert RJ: The
long and short of microRNA. Cell. 153:516–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu X, Zhu Y, Liang Z, Li S, Xu X, Wang X,
Wu J, Hu Z, Meng S, Liu B, et al: c-Met and CREB1 are involved in
miR-433-mediated inhibition of the epithelial-mesenchymal
transition in bladder cancer by regulating Akt/GSK-3β/Snail
signaling. Cell Death Dis. 7:e20882016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang XC, Ma Y, Meng PS, Han JL, Yu HY and
Bi LJ: miR-433 inhibits oral squamous cell carcinoma (OSCC) cell
growth and metastasis by targeting HDAC6. Oral Oncol. 51:674–682.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weiner-Gorzel K, Dempsey E, Milewska M,
McGoldrick A, Toh V, Walsh A, Lindsay S, Gubbins L, Cannon A,
Sharpe D, et al: Overexpression of the microRNA miR-433 promotes
resistance to paclitaxel through the induction of cellular
senescence in ovarian cancer cells. Cancer Med. 4:745–758. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang Z, Tsuchiya H, Zhang Y, Hartnett ME
and Wang L: MicroRNA-433 inhibits liver cancer cell migration by
repressing the protein expression and function of cAMP response
element-binding protein. J Biol Chem. 288:28893–28899. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo LH, Li H, Wang F, Yu J and He JS: The
tumor suppressor roles of miR-433 and miR-127 in gastric cancer.
Int J Mol Sci. 14:14171–14184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Furlong F, Fitzpatrick P, O'Toole S,
Phelan S, McGrogan B, Maguire A, O'Grady A, Gallagher M, Prencipe
M, McGoldrick A, et al: Low MAD2 expression levels associate with
reduced progression-free survival in patients with high-grade
serous epithelial ovarian cancer. J Pathol. 226:746–755. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kroh EM, Parkin RK, Mitchell PS and Tewari
M: Analysis of circulating microRNA biomarkers in plasma and serum
using quantitative reverse transcription-PCR (qRT-PCR). Methods.
50:298–301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 50:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Tekcham DS and Tiwari PK: Non-coding RNAs
as emerging molecular targets of gallbladder cancer. Gene.
588:79–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Oliveto S, Mancino M, Manfrini N and Biffo
S: Role of microRNAs in translation regulation and cancer. World J
Biol Chem. 8:45–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu J, Zhang W, Tang H, Qian H, Yang J, Zhu
Z, Ren P and Lu B: Septin 2 accelerates the progression of biliary
tract cancer and is negatively regulated by mir-140-5p. Gene.
589:20–26. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Williams TM, Majithia L, Wang SJ and
Thomas CR Jr: Defining the role of adjuvant therapy:
Cholangiocarcinoma and gall bladder cancer. Semin Radiat Oncol.
24:94–104. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Phelip JM, Vendrely V, Rostain F, Subtil
F, Jouve JL, Gasmi M, Michel P, Le Malicot K, Smith D, Seitz JF, et
al: Gemcitabine plus cisplatin versus chemoradiotherapy in locally
advanced biliary tract cancer: Fédération Francophone de
Cancérologie Digestive 9902 phase II randomised study. Eur J
Cancer. 50:2975–2982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Valle J, Wasan H, Palmer DH, Cunningham D,
Anthoney A, Maraveyas A, Madhusudan S, Iveson T, Hughes S, Pereira
SP, et al: Cisplatin plus gemcitabine versus gemcitabine for
biliary tract cancer. N Engl J Med. 362:1273–1281. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li X, Yang L, Shuai T, Piao T and Wang R:
miR-433 inhibits retinoblastoma malignancy by suppressing Notch1
and PAX6 expression. Biomed Pharmacother. 82:247–255. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liang T, Guo Q, Li L, Cheng Y, Ren C and
Zhang G: MicroRNA-433 inhibits migration and invasion of ovarian
cancer cells via targeting Notch1. Neoplasma. 63:696–704. 2016.
View Article : Google Scholar : PubMed/NCBI
|