Introduction
Colorectal cancer (CRC), the third leading cause of
cancer-related mortality and morbidity globally, is a common
digestive tract tumor (1). Epigenetic
variations, for instance, hyper-methylation of tumor suppressor
genes, were associated with the development and progression of CRC
(2). After years of researches,
unfortunately, the molecular mechanisms of CRC metastasis have not
been completely understood yet.
microRNAs (miRNAs), a cluster of non-coding RNA
molecules that were at the length of about 19–24 nucleotides, could
suppress gene expression via inducing target mRNA degradation
and/or blocking translation (3)
through base-pairing to bind to the 3′ untranslated region (UTR) of
target mRNA (4,5). Up to now, more than 300 miRNAs have been
identified from various organisms (6–9). miRNAs
modulate numerous cell behaviors, for instance, cell
differentiation, proliferation, apoptosis and metastasis (10–12).
Phosphatase and tensin homolog deleted on chromosome
ten (PTEN) is a tumor suppressor gene located at 10q23, and plays a
pivotal role in the pathogenesis of various human cancers.
Evidences demonstrated the involvement of PTEN deactivation in
tumorigenesis, whose loss increased the activity of PI3K/AKT and
was correlated with cell proliferation, migration, invasion and
apoptosis (13,14). Moreover, another study showed that
PTEN, whose loss was positively correlated with malignant
progression including tumor size and TNM advanced stage, played a
crucial role at early/late stages of CRC (15); meanwhile, restoration of PTEN was
reported to reduce the ratio of metastases in an orthotopic model
of CRC (16). Current study aimed to
investigate whether there are miRNAs that could target PTEN and
regulate the progression of CRC, thus providing a therapeutic
target for CRC. In our study, dual luciferase reporter assay was
applied for the verification of interaction between miR-106a and
PTEN 3′ UTR which was predicted by targetscan and PicTar. qRT-PCR
was adopted to determine miR-106a expression level in tissues and
cell lines. HT29 cells were used for the present study, separated
into control group, miR-NC antagomiR group and miR-106a antagomiR
group. HT29 cell proliferation and apoptosis were tested by
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(MTT) and FCM assay, respectively. Western blot was used for the
detection of PTEN, p-PI3K/p-AKT protein level.
Materials and methods
Clinical samples
Forty tumor tissues and the adjacent tissues were
separated from CRC patients who underwent surgery in our hospital.
Tissues were kept in liquid nitrogen as quick as possible. Informed
consent was acquired from all the patients that participated in our
research. Our study was approved by the ethics committee of Central
Hospital of Chengde (Chengde, China).
Cell culture
The normal human colon epithelial cells (NCM640) and
immortalized human colon epithelial cell lines including SW620 and
HT29 were grown in RPMI-1640 medium (Hyclone; GE Healthcare, Logan,
UT, USA) containing 10% fetal bovine serum (FBS; Invitrogen; Thermo
Fisher Scientific Inc.) and 1% antibiotics at 37°C in a humidified
chamber with 95% air and 5% CO2. Cells that expressed
the highest miR-106a level was chosen for the following study.
Plasmid transfection
HT29 cells which showed the highest miR-106a level
were used for the following experiments, and seeded into 24-well
plates at the concentration of 1×105 cells/well.
miR-106a antagomiR was synthesized by GenePharma (Shanghai, China).
The antagomiR, a single-stranded RNA analogue was complementary to
mature miR-106a (5′-CAAAGUGCUAACAGUGCAGGUAG-3′). A mismatched
miR-negative control (NC) antagomiR (5′-UUGUACUACACAAAAGUACUG-3′)
was also synthesized. Transfection of HT29 cells with miR-106a
antagomiR or miR-NC antagomiR was conducted by Lipofectamine 2000
transfection reagent (Thermo Fisher, Inc., Waltham, MA, USA) in
accordance with manufacturer's instruction.
HT29 cells were separated into 3 different groups
including control group, miR-NC antagomiR group and miR-106a
antagomiR group. Forty-eight hours after the transfection of
plasmid, HT29 cells were collected.
Dual-luciferase reporter assay
The sequences of 3′UTR PTEN were amplified by the
following primers (PTEN forward, 5′-cac aac tcg agT GGC AAT AGG ACA
TTG TGTCA-3′ and reverse, 5′-aag gat ccA ACA ACA AGC AGT GAC
AGCG-3′), then digested with BamHI/XhoI and constructed with pLuc
vector. Downstream of the firefly luciferase stop codon was cloned
in the pLuc control vector (Promega Corporation, Madison, WI, USA).
HT29 cells were seeded into 48-well plates and incubated for 24 h,
thereafter, co-transfected 1 mg 3′UTR-PTEN or 3′ UTR mut-PTEN with
the control oligonucleotide (80 nM) or miR-106a antagomiR (80 nM)
by Lipofectamine 2000 reagents (Invitrogen; Thermo Fisher
Scientific Inc.) according to the manufacturer's protocol.
Forty-eight h after transfection, luciferase activity in each group
was determined with the Dual Luciferase Reporter Assay System
(Promega Corporation).
MTT assay
HT29 cells (1×105 cells/ml) were seeded
onto 12-well plates and incubated for 24 h, then incubated with 20
µl of MTT solution (5 mg/ml in PBS) for another 1 h. Followed by
the removal of culture medium, converted purple formazan dye from
MTT was solubilized in dimethyl sulfoxide (DMSO; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). Finally, optical densities from
all the samples were measured at 595 nm by a mocroplate reader
(BioRad, Hercules, CA, USA).
Cell apoptosis analysis
A total of 1×105 HT29 cells were
collected by centrifugation at the speed of 3,000 rpm/min for 5 min
and washed three times with PBS. Cells were re-suspended in RPMI
medium with 1% bovine serum albumin (BSA) and 10% FBS, mixed with
Annexin V-enhanced green fluorescent protein (FITC) and propidium
iodide (PI), and then incubated for 20 min at room temperature in
the dark. Assay results were determined with flow cytometry.
Western blot
Cells were seeded into 6-well plate at the
concentration of 1×105 cells/ml/plate and grown at 37°C
incubator in a 5% CO2 atmosphere for 24 h. Cells were
washed with cold PBS twice, scraped with radioimmunoprecipitation
buffer (RIPA; Beyotime, Shanghai, China), then subjected to western
blot analysis. Primary antibody reacted at 4°C overnight and the
second antibody reacted at room temperature for 2 h. At room
temperature, after washing the samples with Tris Buffered Saline
with Tween-20 (TBST) three times with each time for 10 min. At
last, bands were detected with ECL kit (Beyotime). Bands were
quantified by Odyssey infrared imaging (LICOR, Lincoln, NE, USA)
and GAPDH acted as an internal reference.
Quantitative real time polymerase
chain reaction
Total RNA was extracted from HT29 cells with RNAiso
Plus (Takara, Bio, Inc., Otsu, Japan). To assess the expression
level of miR-106a, a stemloop reverse transcription-polymerase
chain reaction (RT-PCR) was carried out. The relative expression
level of miR-106a was calculated via the comparative cycle
threshold (Ct) method and normalized to U6. Data were processed
using the 2−ΔΔCT method.
Statistical analysis
Student's t test and one-way ANOVA were carried out
to assess statistical significance. Results were presented as mean
± standard deviation (SD). P<0.05 was considered to indicate a
statistically significant difference.
Results
miRNA that could target PTEN
To explore the molecule by which PTEN was targeted,
we used two algorithms (PicTar and Target Scan). Seed sequence of
miR-106a was identified in 3′ UTR of PTEN, the wild-type and mutant
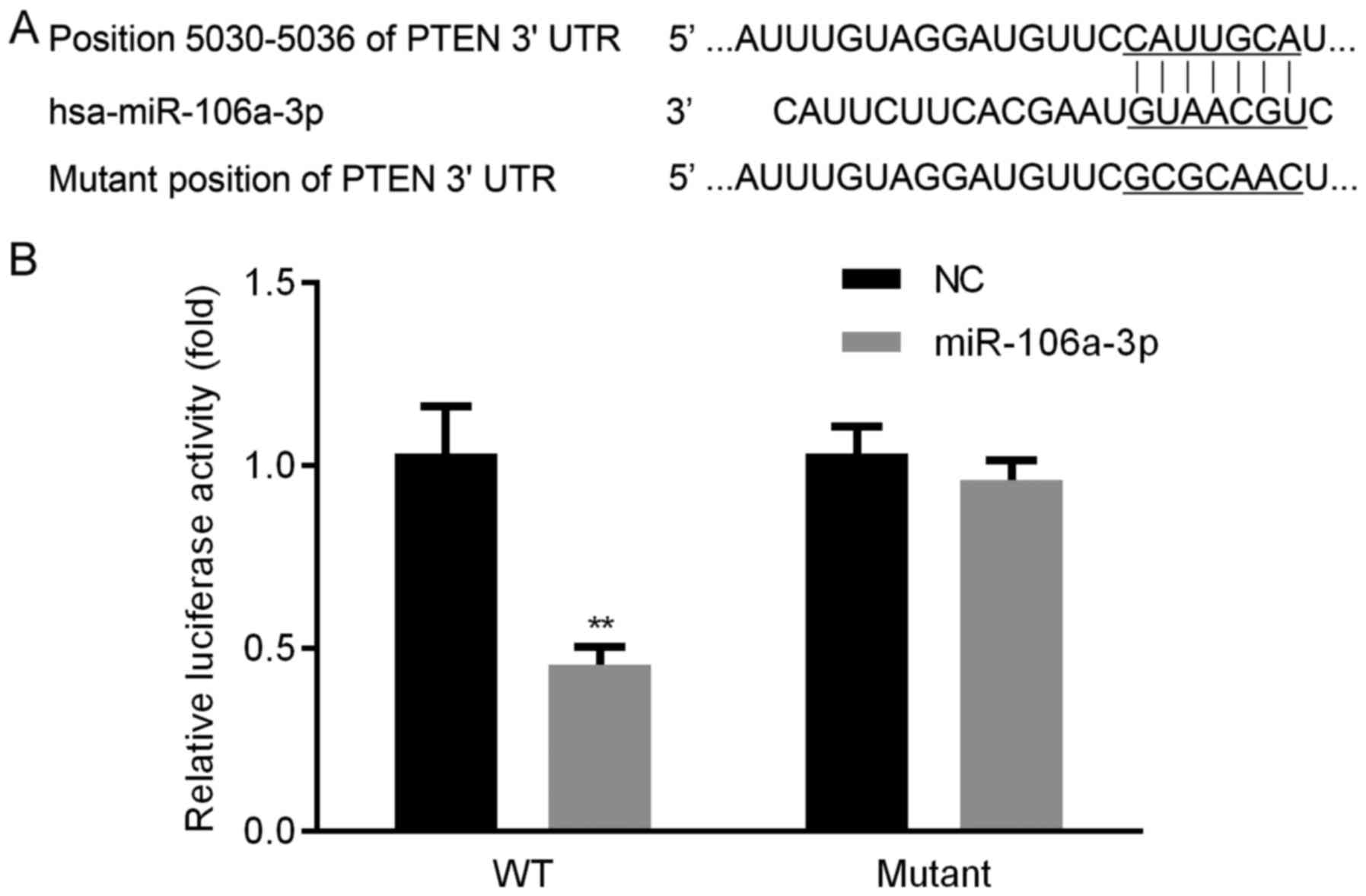
seed sequences between PTEN and miR-106a were displayed (Fig. 1A). We found that, luciferase activity
in WT group was obviously lower, whereas, there was no significant
change of luciferase activity in mutant group (Fig. 1B). These results proved that PTEN
could be targeted by miR-106a.
Expression level of miR-106a in
patients' tissues
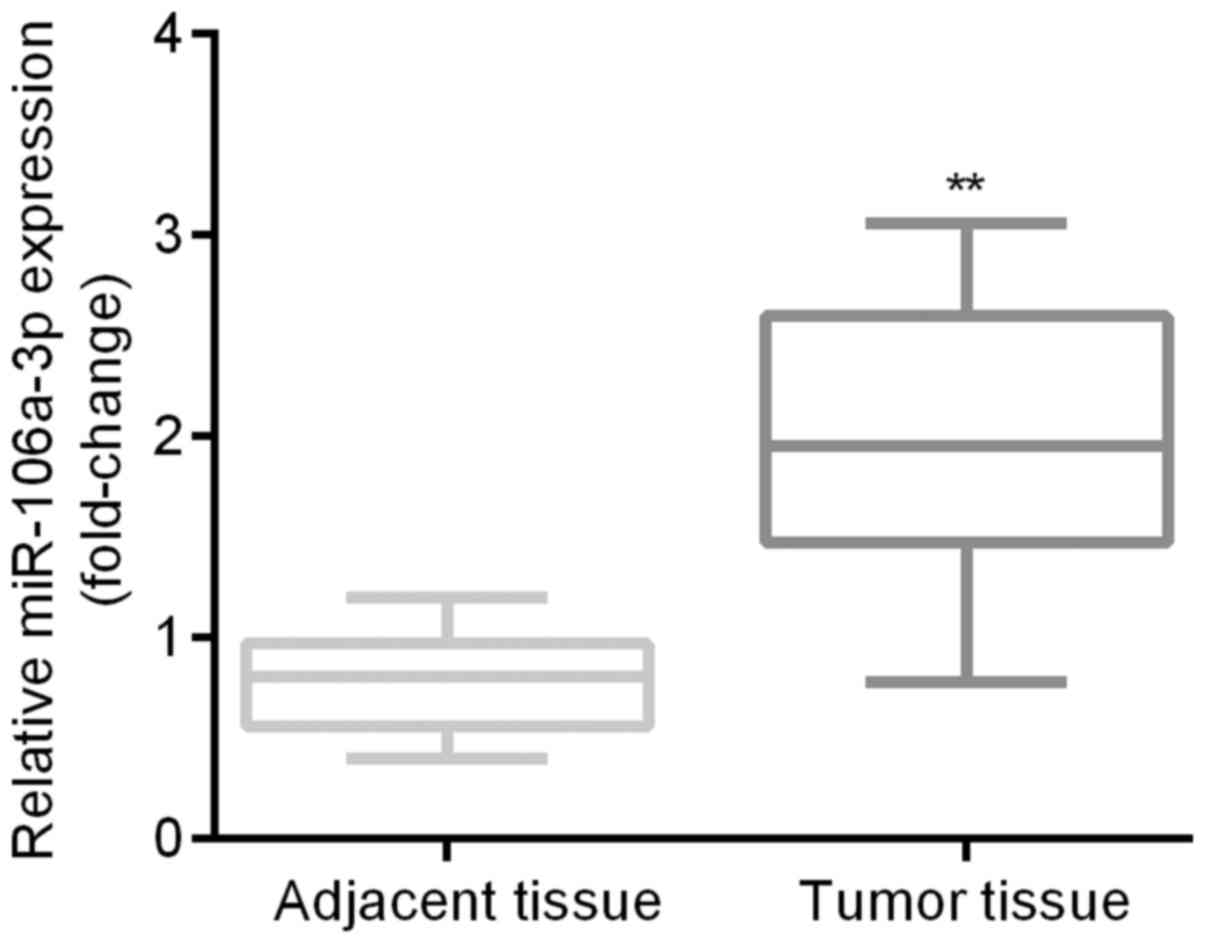
qRT-PCR was applied for the examination of
expression differences of miR-106a between patients' tumor tissues
and the adjacent tissues. Results indicated that miR-106a was
significantly higher in patients' tumor tissues than in the
adjacent tissues (Fig. 2).
Correlations between miR-106a-3p or
PTEN and CRC patients' characteristics
PTEN and CRC gene expression levels in patients'
tumor tissues and the adjacent tissues were tested by qRT-PCR as
presented in Table I. There were 15
male patients and 25 female patients in our study, with 19 patients
aged <60 years old and 21 patients aged ≥60 years old; there
were 18 patients whose tumor size was ≥5 cm and 22 patients whose
tumor size was <5 cm; well-intermediately differentiation was
found in 18 patients and poor differentiation was found in 22
patients; while metastasis was found in 17 patients. As for sex and
age, there was no significant difference of miR-106a-3p or PTEN.
Significant differences were discovered in tumor size, histological
size and metastasis.
 | Table I.The correlations between miR-106a-3p
or PTEN and colorectal cancer patients' characteristics (mean ±
SD). |
Table I.
The correlations between miR-106a-3p
or PTEN and colorectal cancer patients' characteristics (mean ±
SD).
| Factor | Case | miR-106a-3p | P-value | PTEN | P-value |
|---|
| Sex |
|
| 0.32 |
| 0.30 |
| Male | 15 |
2.58±0.34 |
|
2.12±0.51 |
|
|
Female | 25 |
2.45±0.39 |
|
1.96±0.42 |
|
| Age (years) |
|
| 0.18 |
| 0.58 |
|
<60 | 19 |
2.45±0.68 |
|
1.86±0.48 |
|
| ≥60 | 21 |
2.76±0.73 |
|
1.95±0.53 |
|
| Tumor size (cm) |
|
| 0.01 |
| 0.05 |
| ≥5 | 18 |
2.81±0.42 |
|
1.79±0.62 |
|
|
<5 | 22 |
2.21±0.67 |
|
2.22±0.68 |
|
| Histological
grade |
|
| 0.02 |
| 0.01 |
|
Well-intermediately
differentiation | 18 |
2.65±0.54 |
|
1.82±0.52 |
|
| Poor
differentiation | 22 |
2.26±0.46 |
|
2.25±0.38 |
|
| Metastasis |
|
| 0.01 |
| 0.02 |
| No | 23 |
2.28±0.63 |
|
2.19±0.51 |
|
| Yes | 17 |
2.76±0.48 |
|
1.81±0.47 |
|
Expression level of miR-106a in
NCM640, SW620 and HT29
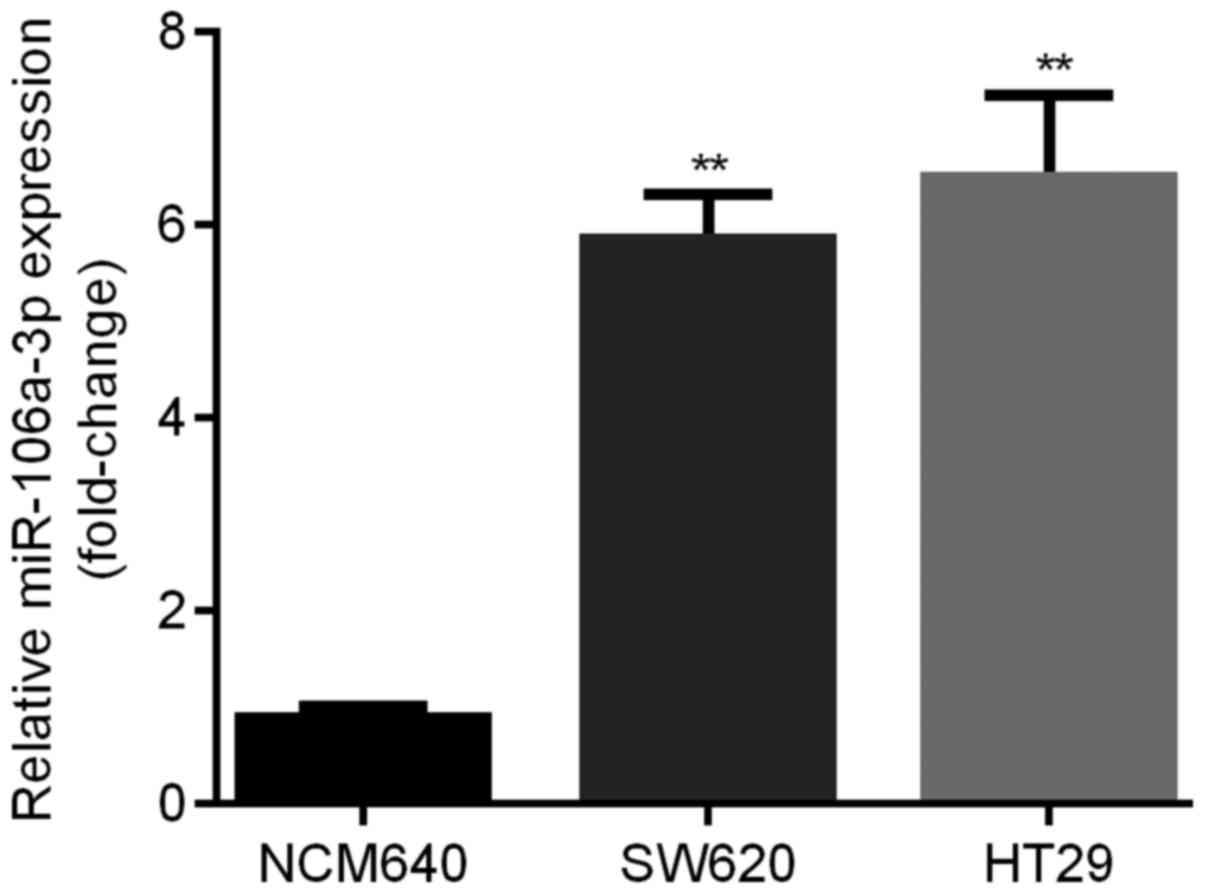
qRT-PCR was applied for the examination of
expression levels of miR-106a in 2 different human CRC cell lines
compared with NCM640. We found that relative expression level of
miR-106a was significantly elevated in SW620 and HT29 cell lines
(about 6-fold). Moreover, HT29 cells showed the highest miR-106a
level (Fig. 3).
Expression level of miR-106a in HT29
cells after transfection
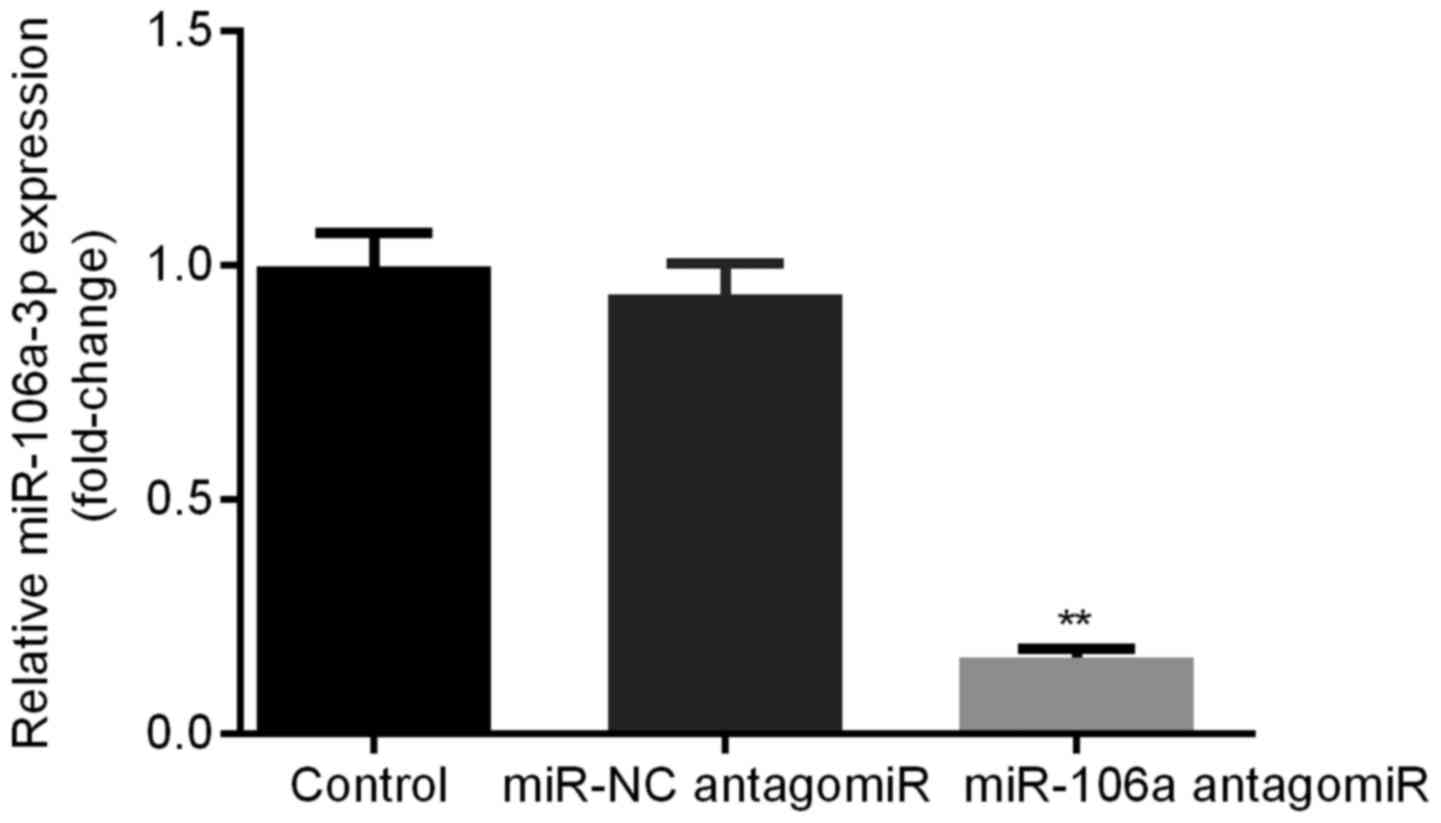
HT29 cells were transfected with miR-NC antagomiR or
miR-106a antagomiR. Relative miR-106a expression levels were
detected by qRT-PCR. We found that, there was no significant
difference of miR-106a expression level between control group and
miR-NC antagomiR group, however, miR-106a was obviously decreased
in miR-106a antagomiR group compared with other two groups. This
result verified the successful transfection of miR-106a antagomiR
into HT29 cells (Fig. 4).
Consequently, the transfected HT29 cells were used for the
conduction of the experiments in our study.
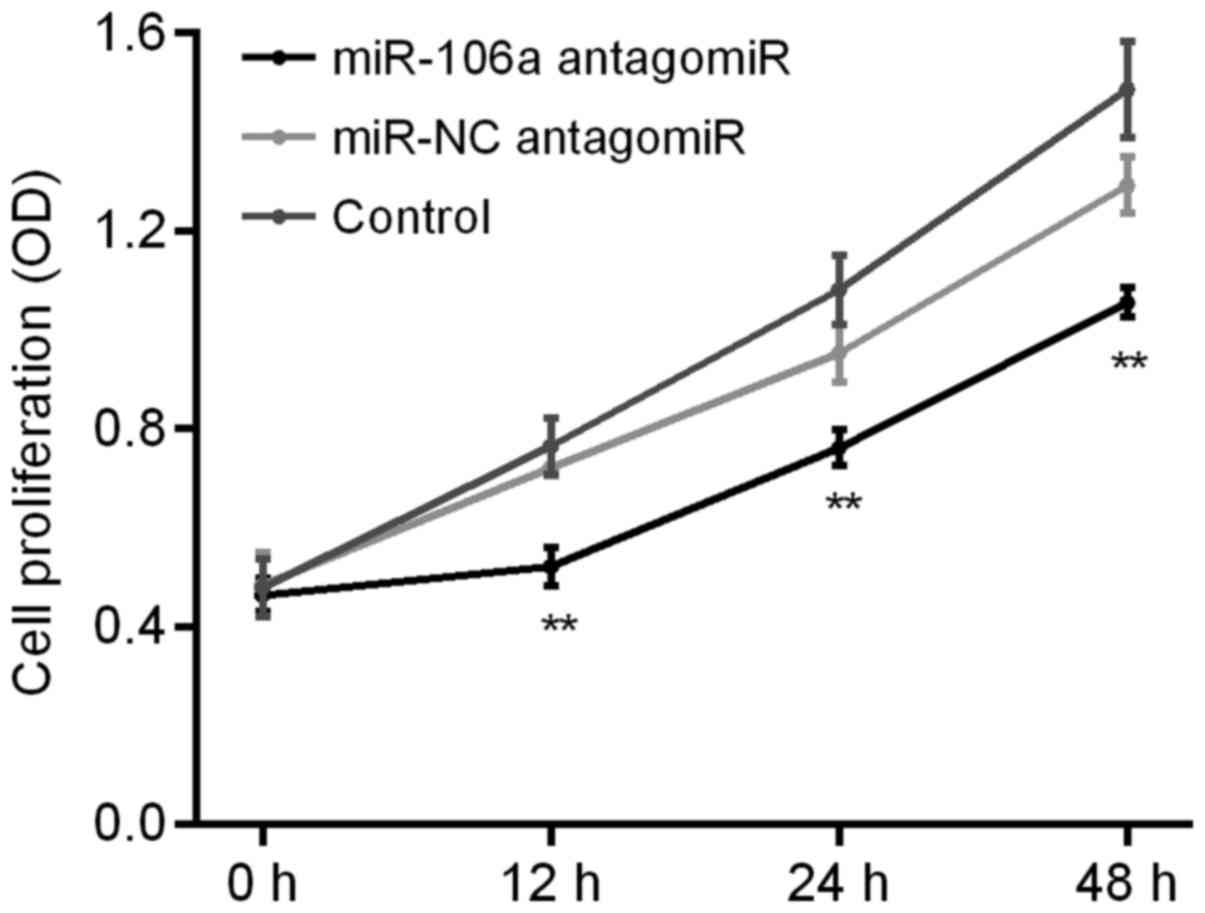
Influence of miR-106a antagomiR on
HT29 cell proliferation
We carried out MTT assay to detect the influence of
miR-106a antagomiR on the cell proliferation of HT29 cells. Results
showed that, there was no significant difference of HT29 cell
proliferation between control group and miR-NC antagomiR group,
however, which was obviously decreased by miR-106a antagomiR
(Fig. 5).
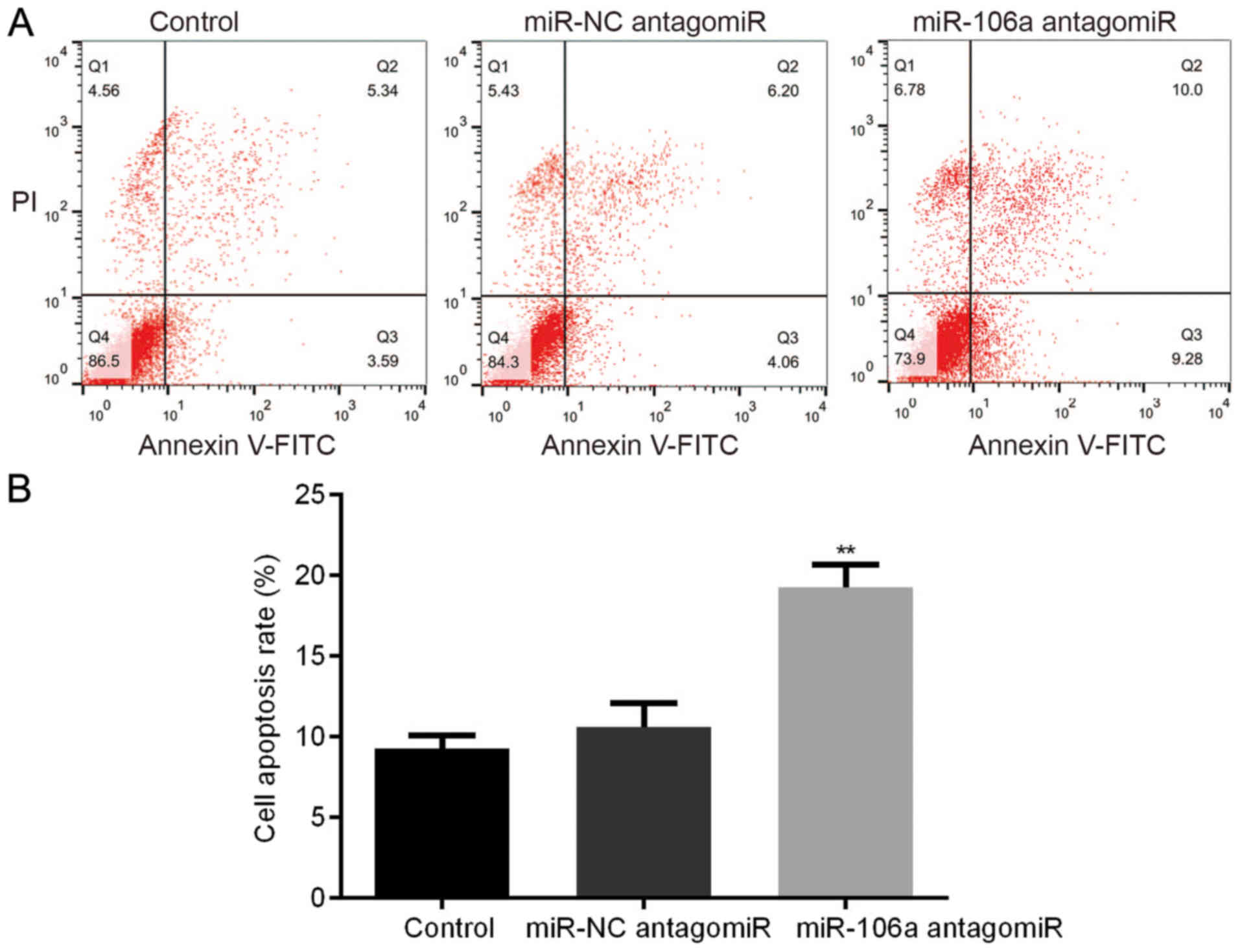
Influence of miR-106a antagomiR on
HT29 cell apoptosis
We carried out FCM assay to detect the influence of
miR-106a antagomiR on the cell apoptosis of HT29 cells. Results
showed that, there was no significant difference of HT29 cell
apoptosis between control group and miR-NC antagomiR group,
however, miR-106a antagomiR treatment led to the highest cell
apoptosis rate among the 3 groups (Fig.
6).
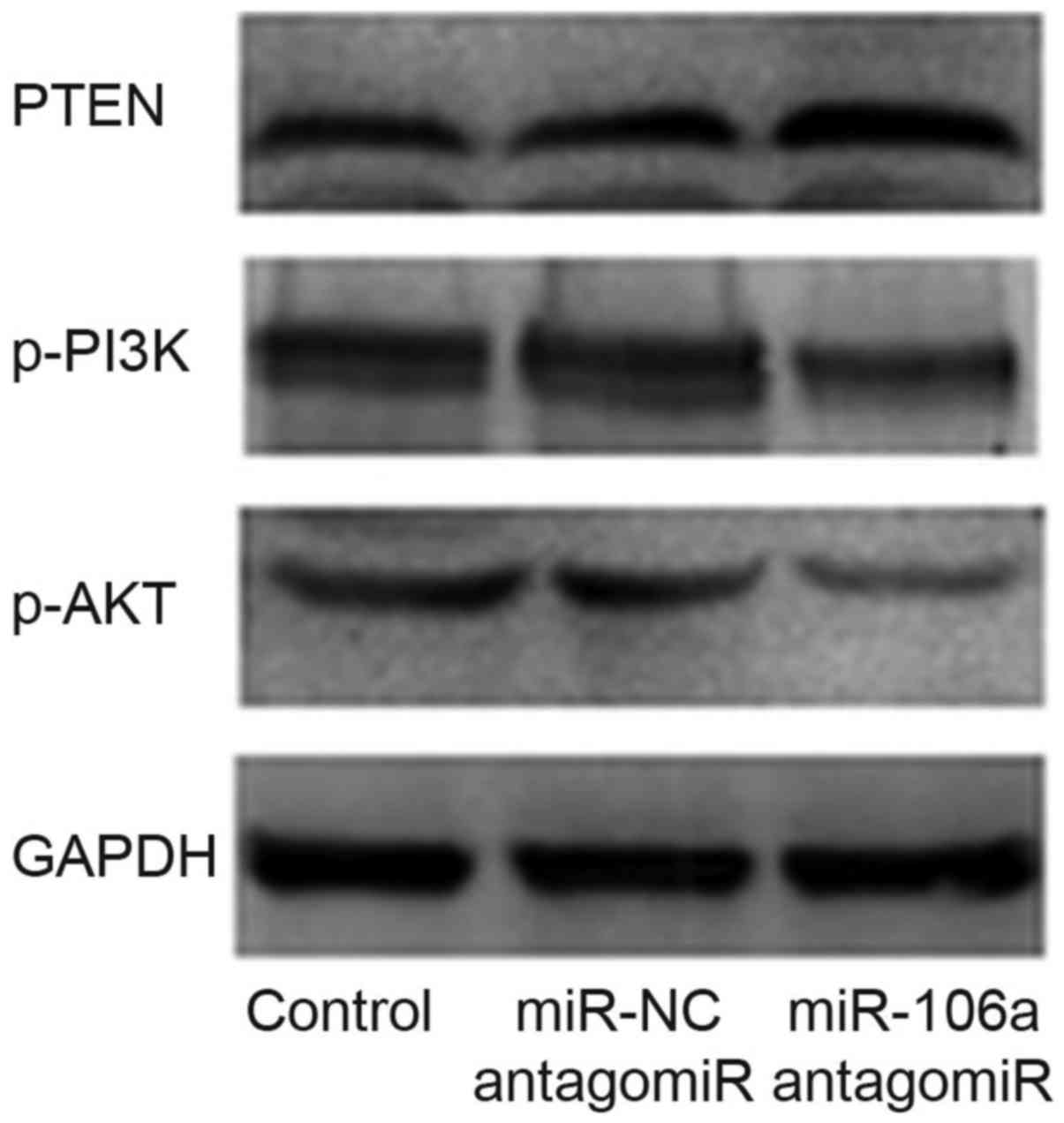
Influence of miR-106a antagomiR on
protein levels of PTEN, p-PI3K and p-AKT
We performed western blot to detect the influence of
miR-106a antagomiR on the expression level of PTEN, p-PI3K and
p-AKT. Results showed that, there was no significant difference of
the protein level between control group and miR-NC antagomiR group,
however, p-PI3K/p-AKT protein levels were obviously decreased and
PTEN protein level was obviously increased by the treatment of
miR-106a antagomiR (Fig. 7).
Discussion
CRC is a common digestive tract tumor (1). miRNAs were found to regulate cell
differentiation, proliferation, apoptosis and metastasis (10–12).
Loss of PTEN was positively correlated with
malignant progression of CRC (15),
restoration of PTEN reduced the ratio of metastases of CRC
(16). Current study aimed to
investigate whether there were miRNAs that targeted PTEN and
regulated the progression of CRC, thus providing a therapeutic
target for CRC.
To explore the molecule by which PTEN was targeted,
we used two algorithms (PicTar and TargetScan). Seed sequence of
miR-106a was identified in 3′ UTR of PTEN (Fig. 1A). The interaction between miR-106a
and PTEN were confirmed by dual luciferase activity (Fig. 1B). qRT-PCR indicated that, in
patients' tumor tissues miR-106a was obviously upregulated compared
to the adjacent tissues (Fig. 2),
which was in consistent with previous studies that reported high
expression of miR-106a in cancer tissues of CRC patients (17,18);
meanwhile, compared to NCM640 cells, miR-106a was significantly
elevated in SW620 and HT29 cell lines (Fig. 3), which was in line with a previous
study (19). These results together
suggested the oncogenetic role of miR-106a in CRC patients and in
SW620/HT29 cells.
Successful transfection of miR-106a antagomiR into
HT29 cells was confirmed by the lowest miR-106a level in the group
treated with miR-106a antagomiR among the 3 groups (Fig. 4). Consequently, HT29 cells
successfully transfected with miR-106a antagomiR were used for
carrying out the following experiments.
As for the influences of miR-106a on HT29 cell
proliferation and apoptosis, MTT assay and FCM assay were carried
out, respectively. Results showed that, there was no significant
difference of HT29 cell proliferation between control group and
miR-NC antagomiR group, however, which was significantly decreased
by the treatment of miR-106a antagomiR (Fig. 5). Otherwise, there was no significant
difference of HT29 cell apoptosis between control group and miR-NC
antagomiR group, which was notably increased by miR-106a antagomiR
(Fig. 6). Whereas, the signaling
pathways that were responsible for the changes remained to be
investigated.
Activation of PI3K phosphorylated and activated AKT,
thus localizing AKT to the plasma membrane; PI3K/AKT, an
intracellular signaling pathway, was directly associated with
cellular proliferation, cancer and longevity (20). And there was a natural inhibitor that
named PTEN to inhibit the activation of PI3K/AKT signaling pathway,
thus limiting cell proliferation and cancer progression;
furthermore, knocking out of PTEN was shown to elevate the mass of
brain on account of the corresponding unregulated proliferation
(21). Administration of PTEN
inhibitor was found to temporarily and safely impact the activation
of PI3K/AKT signaling pathway thus influencing cell survival
(22) and proliferation (23).
Consequently, we carried out western blot and found
that, there was no significant difference of the protein level
between control group and miR-NC antagomiR group, however,
p-PI3K/p-AKT were obviously decreased and PTEN was obviously
increased by the treatment of miR-106a antagomiR (Fig. 7).
Taken together, miR-106a antagomiR upregulated PTEN
protein level and HT29 cell apoptosis, downregulated p-PI3K/p-AKT
protein levels and HT29 cell proliferation via targeting 3′ UTR of
PTEN, which suggested the potential role of miR-106a as a
therapeutic target for the treatment of CRC.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Toyota M, Ahuja N, Ohe-Toyota M, Herman
JG, Baylin SB and Issa JP: CpG island methylator phenotype in
colorectal cancer. Proc Natl Acad Sci USA. 96:pp. 8681–8686. 1999;
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pasquinelli AE and Ruvkun G: Control of
developmental timing by microRNAs and their targets. Annu Rev Cell
Dev Biol. 18:495–513. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abrahante JE, Daul AL, Li M, Volk ML,
Tennessen JM, Miller EA and Rougvie AE: The Caenorhabditis elegans
hunchback-like gene lin-57/hbl-1 controls developmental time and is
regulated by microRNAs. Dev Cell. 4:625–637. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dostie J, Mourelatos Z, Yang M, Sharma A
and Dreyfuss G: Numerous microRNPs in neuronal cells containing
novel microRNAs. RNA. 9:180–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lagos-Quintana M, Rauhut R, Lendeckel W
and Tuschl T: Identification of novel genes coding for small
expressed RNAs. Science. 294:853–858. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lim LP, Glasner ME, Yekta S, Burge CB and
Bartel DP: Vertebrate microRNA genes. Science. 299:15402003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Reinhart BJ, Weinstein EG, Rhoades MW,
Bartel B and Bartel DP: Micro-RNAs in plants. Genes Dev.
16:1616–1626. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Croce CM and Calin GA: miRNAs, cancer and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ventura A and Jacks T: MicroRNAs and
cancer: Short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bermúdez Brito M, Goulielmaki E and
Papakonstanti EA: Focus on PTEN regulation. Front Oncol. 5:1662015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Miao Y, Zheng W, Li N, Su Z, Zhao L, Zhou
H and Jia L: Micro-RNA-130b targets PTEN to mediate drug resistance
and proliferation of breast cancer cells via the PI3K/Akt signaling
pathway. Sci Rep. 7:419422017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Waniczek D, Śnietura M, Mlynarczyk-Liszka
J, Piglowski W, Kopeć A, Lange D, Rudzki M and Arendt J: PTEN
expression profiles in colorectal adenocarcinoma and its
precancerous lesions. Pol J Pathol. 64:15–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chowdhury S, Ongchin M, Wan G, Sharratt E,
Brattain MG and Rajput A: Restoration of PTEN activity decreases
metastases in an orthotopic model of colon cancer. J Surg Res.
184:755–760. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schetter AJ, Leung SY, Sohn JJ, Zanetti
KA, Bowman ED, Yanaihara N, Yuen ST, Chan TL, Kwong DL, Au GK, et
al: MicroRNA expression profiles associated with prognosis and
therapeutic outcome in colon adenocarcinoma. JAMA. 299:425–436.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Diaz R, Silva J, Garcia JM, Lorenzo Y,
Garcia V, Peña C, Rodríguez R, Muñoz C, García F, Bonilla F and
Domínguez G: Deregulated expression of miR-106a predicts survival
in human colon cancer patients. Genes Chromosomes Cancer.
47:794–802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feng B, Dong TT, Wang LL, Zhou HM, Zhao
HC, Dong F and Zheng MH: Colorectal cancer migration and invasion
initiated by microRNA-106a. PLoS One. 7:e434522012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
King D, Yeomanson D and Bryant HE: PI3King
the Lock: targeting the PI3K/Akt/mTOR pathway as a novel
therapeutic strategy in neuroblastoma. J Pediatr Hematol Oncol.
37:245–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rafalski VA and Brunet A: Energy
metabolism in adult neural stem cell fate. Prog Neurobiol.
93:182–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lai JP, Dalton JT and Knoell DL:
Phosphatase and tensin homologue deleted on chromosome ten (PTEN)
as a molecular target in lung epithelial wound repair. Br J
Pharmacol. 152:1172–1184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wyatt LA, Filbin MT and Keirstead HS: PTEN
inhibition enhances neurite outgrowth in human embryonic stem
cell-derived neuronal progenitor cells. J Comp Neurol.
522:2741–2755. 2014. View Article : Google Scholar : PubMed/NCBI
|