Introduction
Renal cell carcinoma (RCC) accounts for 2–3% of all
malignancies in adults and is the most common type of tumor of the
urinary system. RCCs may be divided into clear cell, papillary,
chromophobe, collecting duct and unclassified RCC (1). Renal clear cell carcinoma, originating
from renal tubular epithelial cells, is the most common type of
RCC, accounting for 80–90% of all cases (1). The effect of surgical treatment in
patients with circumscribed RCC is good and the 5-year survival
rate is 60–80% (2). However, the
prognosis of patients with distal metastasis remains poor and the
median survival time of such patients is only ~10 months (2). At the time of diagnosis, 20–35% of
patients already exhibit distal metastasis (3). Furthermore, following surgery, 20–40% of
patients exhibit metastasis, which greatly reduces survival rates.
Therefore, metastasis is the main reason for the poor prognosis of
patients with RCC and RCC-associated mortality. It has also been
hypothesized that RCC staging and pathology are significant factors
affecting the prognosis of patients with RCC (4).
The metastasis of malignant cells is dominated by a
series of complex physiological procedures. It has been suggested
that cell migration serves a key function throughout the process of
tumor metastasis, although the mechanism by which this occurs
remain unknown (5). In normal
epithelial tissue and benign epithelial tumors, epithelial cells
are tightly associated with adjacent cells and the basement
membrane via tight and adhering junctions, as well as hemidesmosome
and desmosome connections (6). In
malignant epithelial tumors however, tumor cells are stimulated by
external signals and consequently acquire migratory and invasive
capabilities. Tumor cells may then degrade the basement membrane,
invade the surrounding extracellular matrix and penetrate
microvascular or lymphatic vessels. The epithelial-mesenchymal
transition (EMT) serves a key function in carcinoma progression,
invasion and metastasis. During the EMT, epithelial functions are
lost and the expression and distribution of proteins, including
E-cadherin, which mediates cell-cell and cell-matrix contacts and
the cytoskeletal organization responsible for normal epithelial
polarity, become disordered (7). The
concurrent gain of mesenchymal characteristics includes the ability
of cells to migrate and invade the surrounding matrix due to the
increased expression of proteins, including N-cadherin and vimentin
(8). However, few studies have
investigated the EMT mechanisms that occur in RCC (5,9).
Therefore, determining the regulatory mechanisms of the EMT in RCC
should improve understanding of how metastasis occurs.
F-box and WD repeat domain containing 7 (Fbxw7) is
an F-box protein that belongs to the SKP1-CUL1-F-box protein E3
ligase complex and is responsible for transferring the ubiquitin
molecule to the substrate, resulting in its recognition and
subsequent degradation by proteasomes (10). It identifies a network of signal
proteins that function in cell growth, differentiation and
apoptosis (11,12). Several specific substrates of Fbxw7
have been identified, including cyclin E (13), c-Jun (14), c-Myc (15), mechanistic target of rapamycin
(16) and Notch (17), which are all overexpressed in
malignant tumors. Therefore, Fbxw7 is an important tumor suppressor
gene, as it negatively regulates the expression of these proteins.
The mutation rate of Fbxw7 in cholangiocarcinoma is 35% (18,19).
Furthermore, 30% of patients with acute T cell lymphatic leukemia
(11), 16% of patients with primary
endometrial cancer (20) and 11% of
patients with colorectal cancer (21)
exhibit an Fbxw7 mutation, and the overall mutation frequency of
Fbxw7 is ~6% in human primary tumors (19). In addition, previous studies have
demonstrated that Fbxw7 may affect the migration and invasion of
tumors, including gastric cancer and melanoma, and serve as a
prognostic marker (22,23). However, to the best of our knowledge,
there have been no reports on the association between Fbxw7 and
metastasis in RCC.
The aim of the present study was to identify the
function of Fbxw7 in the migration and invasion of renal cancer and
to investigate its underlying molecular mechanisms of action.
Immunohistochemistry (IHC) was used to examine the expression of
the Fbxw7 protein in RCC tissues compared with non-cancerous
tissue. Subsequently, the potential mechanism of Fbxw7 and the
involvement of the EMT in the migration and invasion of RCC cells
was explored in vitro.
Materials and methods
Tissue samples
A total of 70 renal cancer samples and paired normal
tumor-adjacent samples were taken from patients (42 males and 28
females, mean age 52.5 years) with RCC who underwent radical
nephrectomy or nephron-sparing surgery at the Department of
Urology, Ruijin Hospital, Shanghai Jiao Tong University (Shanghai,
China) between July 2008 and April 2014. All tumors were
pathologically diagnosed as RCC by experienced pathologists (Ruijin
Hospital, Shanghai Jiao Tong University School of Medicine,
Shanghai, China) by applying the TNM classification (7th edition)
(24) following surgery and no
patients had received chemo- or radiotherapy prior to surgery. All
clinicopathological parameters were acquired from the archive of
the pathology department and confirmed by the medical records of
the hospital. The present study was approved by the Ethics
Committee of the Ruijin Hospital, Shanghai Jiaotong University
(Shanghai, China) and all patients gave informed consent for the
use of their tissue in the current study. The follow-up period
began on the date of tumor resection and ended in February 2014 or
once the patient succumbed.
Cell culture
The human RCC cell lines 786-O and ACHN were
obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China) and grown in the
laboratory. Cells were cultured in RPMI-1640 medium supplemented
with 10% fetal bovine serum (FBS), 100 U/ml penicillin and
streptomycin (all Gibco; Thermo Fisher Scientific Inc., Waltham,
MA, USA), and incubated at 37°C in a humidified atmosphere with 5%
CO2.
Western blot analysis
Standard methods were used for western blotting.
Cells were lysed in radioimmunoprecipitation assay buffer (Cell
Signaling Technology, Inc., Danvers, MA, USA). Protein
concentration was determined using a bicinchoninic acid assay kit
(Pierce; Thermo Fisher Scientific, Inc.). The total extracted
protein contents (50 µg) were separated via SDS-PAGE (10% gel) and
then blotted onto a PVDF membrane (EMD Millipore, Billerica, MA,
USA). The membrane was blocked with 5% non-fat milk in PBS with
Tween-20 at room temperature for 2 h and followed by incubation at
4°C overnight with primary antibodies against Fbxw7 (cat. no.
ab109617; 1:300 dilution; Abcam, Cambridge, MA, USA), E-cadherin
(cat. no. 3195; 1:1,000 dilution; Cell Signaling Technology, Inc.),
N-cadherin (cat. no. 13116; 1:1,000 dilution; Cell Signaling
Technology, Inc.), vimentin (cat. no. 5741; 1:1,000 dilution; Cell
Signaling Technology, Inc.) and β-actin (cat. no. sc70319; 1:1,000
dilution; Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
Subsequently, blots were probed with appropriate secondary antibody
(cat. no. LI-COR; 1:8,000 dilution; Lincoln, NE, USA) for 1 h at
room temperature and visualized using the Bio-Rad Gel imaging
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The results
were analyzed using Quantity One software (Version 4.62; GelDoc
XR+; Bio-Rad Laboratories, Inc.) as specified by Bio-Rad.
Wound healing assay
Cells were seeded to 90–100% confluence of monolayer
cells overnight prior to serum starvation for 8 h in 6-well plates
at 37°C. Following wounding with a sterile pipette tip, cells were
rinsed three times with PBS to eliminate floating cells. Images of
the wounds were captured at 0, 24 and 48 h post-wound using a
phase-contrast microscope. The migration rate was calculated as
(%)=[W (24 or 48 h)-W (0 h)]/W (0 h). All assays were independently
performed in triplicate.
Transwell migration and invasion
assays
Transwell assays were performed as previously
described (25). Cell invasion assays
were performed in Matrigel-coated Transwell inserts containing
polycarbonate filters with 6-µm pores. Cell suspension
(2×105 cells in 200 µl serum-free RPMI-1640 medium) was
plated onto the upper chamber. In the lower chamber, RPMI 1640
medium containing 10% FBS was used as a chemoattractant. Following
incubation for 24 h at 37°C, non-migrating cells were scraped using
a cotton swab and cells on the lower surface of the filter were
fixed with 4% methanol for 15 min at room temperature and stained
with 0.5% crystal violet for 10 min at room temperature. Cells were
counted under a light microscope (magnification, ×100) in five
different fields and in duplicate wells, in at least three
independent experiments. The protocol used for the migration assay
was the same as the Matrigel invasion assay, except that the
Transwell insert was not coated with Matrigel.
IHC
A total of 70 samples were taken from patients with
RCC for which complete clinicopathological information was
available. Formalin-fixed, paraffin-embedded tissue sections of 4
µm thickness were baked at 60°C for ≥3 h and were then incubated
twice in xylene for 10 min each time prior to rehydration in a
descending alcohol series. Subsequently, citrate buffer was used
for antigen retrieval by microwave heating for 15 min. The sections
were then blocked in 10% goat serum (Sp kit; Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) at 37°C for 30
min prior to incubation at 4°C overnight in anti-Fbxw7 primary
antibody (cat. no. ab109617; 1:300, Abcam, Cambridge, MA, USA). The
biotinylated secondary antibody (Sp kit, Beijing, China) was used
to detect the primary antibody at 30°C for 30 min. Horseradish
peroxidase-streptavidin conjugate was used to connect with
biotinylated secondary antibody at 37°C for 30 min. The rest of the
staining was performed following the manufacturer's protocol. For
detection of bound antibody, 3,3′-diaminobenzidine was used for 2–5
min depending on the degree of the color at room temperature. The
measurement of positive staining density was determined using an
immunohistochemical scoring system to assess the proportion of
tumor cells exhibiting specific staining. Staining intensity was
assessed using four grades: 0 for none; 1 for weak; 2 for moderate
and 3 for strong. The percentage of positive cells was divided into
five degrees: 1 for <5%; 2 for 5–25%; 3 for 26–50%; 4 for 50–75%
and 5 for 75–100%. Total scores ≤4 were defined as low expression
and scores of >4 were considered to indicate high expression of
Fbxw7 (26). Staining was evaluated
under a light microscope. The overall intensity and percentage were
evaluated by Image-Pro plus software (version 6.0; Media
Cybernetics, Rockville, MD, USA) at five independent fields
(magnification, ×200).
Establishment of knockdown cells and
stable expression of Fbxw7
Small interfering RNA (siRNA) specific to human
Fbxw7 and scramble siRNA (NC) were purchased from Biomics
Biotechnologies Co., Ltd. (Nantong, China). 786-O cells
(1×105 cells) were plated into 6-well plates and allowed
to adhere for 24 h and cultured to 60–70% confluence and cells were
then transfected with 100 nM siRNA (Fbxw7 siRNA or NC siRNA) using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Cells were collected for
further investigation 24 h after transfection.
The plasmid expressing Fbxw7 and the control plasmid
(Biomics Biotechnologies, Nantong, China) were transfected into
ACHN cells by Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) when cells were cultured to 80% confluence
according to the manufacturer's protocol. Cells transfected with
empty vector (control plasmid) were used as controls.
Plasmid-Lipofectamine 2000 complex was prepared as follows: i)
Diluted DNA (0.8 g) with 50 ul serum-free medium and mixed gently;
ii) diluted 2 µl Lipofectamine 2000 with 48 µl serum-free medium,
mixed gently and incubate for 5 min at 4°C; iii) the two solutions
were mixed together and incubated for 20 min at 4°C. A total of 100
ul complex was added to the culture plate. The transfected cells
were selected and incubated with Geneticin (G418) from Invitrogen
(Thermo Fisher Scientific, Inc.) at a dose of 600 ug/ml for 2 weeks
in order to produce overexpression clones. Stable transfected
clones were selected and Fbxw7 expression assessed using western
blot analysis.
Statistical analysis
All data are presented as the mean ± standard
deviation from ≥3 separate experiments. A χ2 test was
used to analyze the association between Fbxw7 expression and
patient clinicopathological characteristics. The Kaplan-Meier
survival curve was used to analyze overall survival (OS) rates of
patients and differences were compared using the Log-rank test. Cox
proportional hazard analysis was employed for univariate and
multivariate analyses. *P<0.05 was considered to indicate a
statistically significant difference. Numerical data were
calculated using Microsoft Excel and analyzed using SPSS 18.0
(SPSS, Inc., Chicago, IL, USA).
Results
Fbxw7 is frequently downregulated in
RCC and is associated with poor RCC prognosis
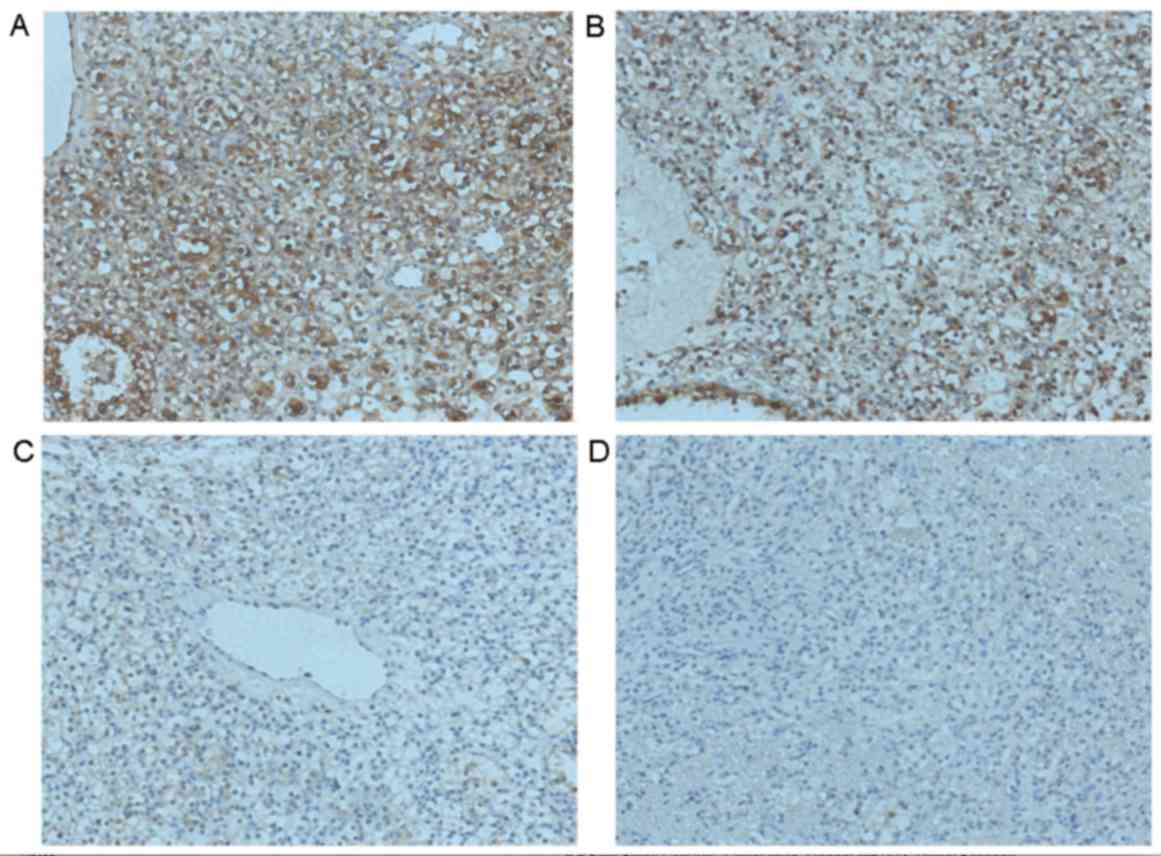
IHC was performed to investigate the expression of
Fbxw7 in tumor and adjacent non-tumor tissues collected from 70
patients with RCC to determine the clinical significance of Fbxw7
(Fig. 1). Of the 70 tissues analyzed,
Fbxw7 expression was downregulated in 17 RCC samples (24.3%)
compared with adjacent non-tumor tissues. Additionally, to
determine the association between Fbxw7 expression and the
clinicopathological features of patients with RCC, 70 patients with
RCC were separated into Fbxw7 low and high expression groups,
according to their IHC scores. The results revealed a significant
association between Fbxw7 expression and tumor node metastasis
(TNM) (24) stage (P=0.005), lymph
node metastasis (P=0.003) and distant metastasis (P=0.006).
However, there were no significant associations between Fbxw7
expression and other clinicopathological factors, including age,
gender, tumor size and histological grade (Table I). These data indicate that the
decreased expression of Fbxw7 stimulates RCC progression.
 | Table I.Association of Fbxw7 expression with
clinicopathological factors in 70 patients with renal cell
carcinoma. |
Table I.
Association of Fbxw7 expression with
clinicopathological factors in 70 patients with renal cell
carcinoma.
|
|
| Fbxw7 expression |
|
|---|
|
|
|
|
|
|---|
| Patient
clinicopathological parameters | Number of
cases | Low expression | High
expression | P-values |
|---|
| Age (years) |
|
|
| 0.994 |
|
≤60 | 33 | 8 | 25 |
|
|
>60 | 37 | 9 | 28 |
|
| Sex |
|
|
| 0.306 |
|
Male | 42 | 12 | 30 |
|
|
Female | 28 | 5 | 23 |
|
| Histological
grade |
|
|
| 0.262 |
|
Well | 45 | 9 | 36 |
|
|
Moderate and poor | 25 | 8 | 17 |
|
| Tumor size |
|
|
| 0.856 |
| ≤5
cm | 44 | 11 | 33 |
|
| >5
cm | 26 | 6 | 20 |
|
| Tumor stage |
|
|
| 0.005a |
| I and
II | 41 | 5 | 36 |
|
| III and
IV | 29 | 12 | 17 |
|
| Lymph node
metastasis |
|
|
| 0.003a |
|
Positive | 21 | 10 | 11 |
|
|
Negative | 49 | 7 | 42 |
|
| Distant
metastasis |
|
|
| 0.006a |
|
Positive | 12 | 7 | 5 |
|
|
Negative | 58 | 10 | 48 |
|
Low Fbxw7 expression is associated
with the poor survival of patients with RCC
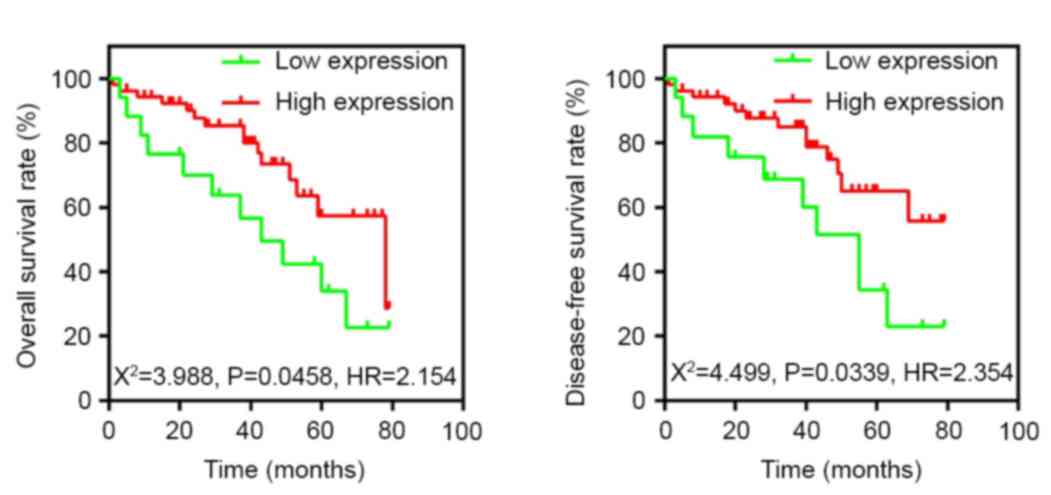
To determine whether the decreased expression of
Fbxw7 in RCC was associated with poor patient prognosis, a
Kaplan-Meier survival analysis was performed using the overall
5-year survival and disease-free survival data from 70 patients
with RCC (Fig. 2). The results
revealed that patients with low Fbxw7 expression had a
significantly decreased postoperative survival time compared with
those with high Fbxw7 expression (P=0.0458). Furthermore, Fbxw7
downregulation was associated with a significantly shorter
disease-free survival duration (P=0.0339). To control for potential
confounders, a Cox proportional hazards regression analysis was
performed, which revealed that histological grade (P=0.013), Fbxw7
expression (P<0.001), tumor stage (P<0.001), lymph node
metastasis (P<0.001) and distant metastasis (P<0.001) were
significantly associated with the OS of patients with RCC (Table II). Multivariate analysis further
confirmed that reduced Fbxw7 expression was an independent
predictor for a shorter OS in patients with RCC (HR=0.623;
P<0.001; Table II). These data
indicated that reduced Fbxw7 expression is associated with the
poorer prognosis of patients with RCC.
| Table II.Univariate and multivariate Cox
regression analyses of overall survival in 70 patients with renal
cell carcinoma. |
Table II.
Univariate and multivariate Cox
regression analyses of overall survival in 70 patients with renal
cell carcinoma.
| Tumor
characteristics | HR (95% CI) | P-value |
|---|
| Univariate
analysis |
|
|
| Age
(≥60 vs. <60) years | 1.461
(0.943–2.265) | 0.090 |
| Sex
(male vs. female) | 0.978
(0.575–1.664) | 0.935 |
|
Histological grade
(poor/well) | 1.784
(1.130–2.818) | 0.013a |
| Tumor
size (>5 vs. ≤5) cm | 1.454
(0.950–2.227) | 0.085 |
| Tumor
stage (III, IV/I, II) | 2.512
(2.112–2.987) |
<0.001a |
| Lymph
node metastasis (positive vs. negative) | 2.336
(1.966–2.776) |
<0.001a |
| Distant
metastasis (positive vs. negative) | 2.621
(1.984–3.462) |
<0.001a |
| Fbxw7
(high vs. low) | 0.465
(0.360–0.601) |
<0.001a |
| Multivariate
analysis |
|
|
|
Histological grade
(poor/well) | 1.436
(0.906–2.275) | 0.124 |
| Tumor
stage (III, IV/I, II) | 1.962
(1.626–2.369) |
<0.001a |
| Lymph
node metastasis (positive vs. negative) | 1.784
(1.459–2.181) |
<0.001a |
| Distant
metastasis (positive vs. negative) | 1.621
(1.225–2.145) |
<0.001a |
| Fbxw7
(high vs. low) | 0.623
(0.479–0.810) |
<0.001a |
Fbxw7 regulates the migration and
invasion of RCC cells
The ability to migrate and invade is important for
RCC metastasis. Currently, little is known regarding whether Fbxw7
is able to regulate the migration and invasion of RCC cells. Thus,
to identify whether Fbxw7 regulates these processes in RCC, the
786-O and ACHN cell lines were employed, since it has been
demonstrated that the expression of Fbxw7 is high in 786-O cells
and low in ACHN cells (26). The
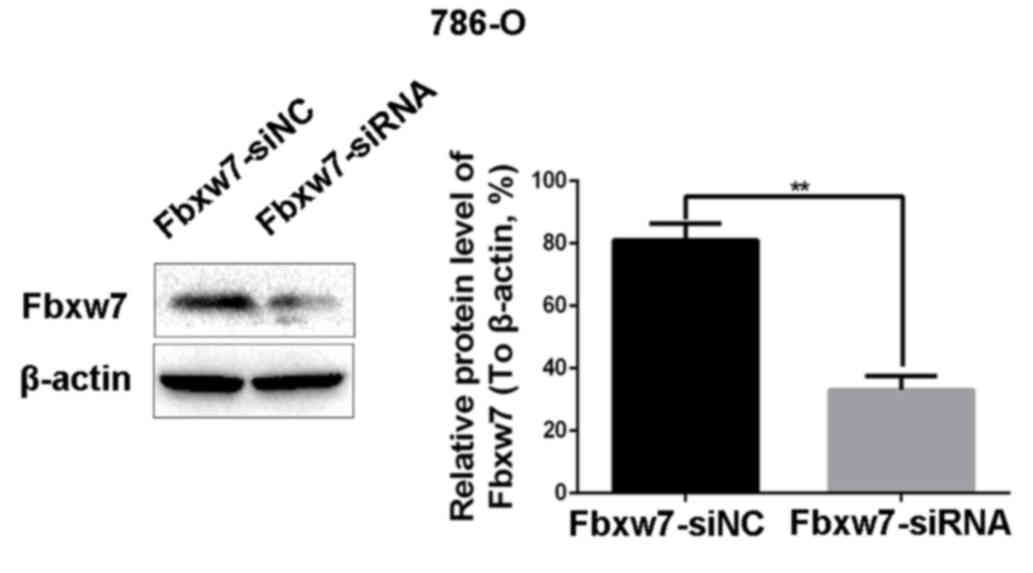
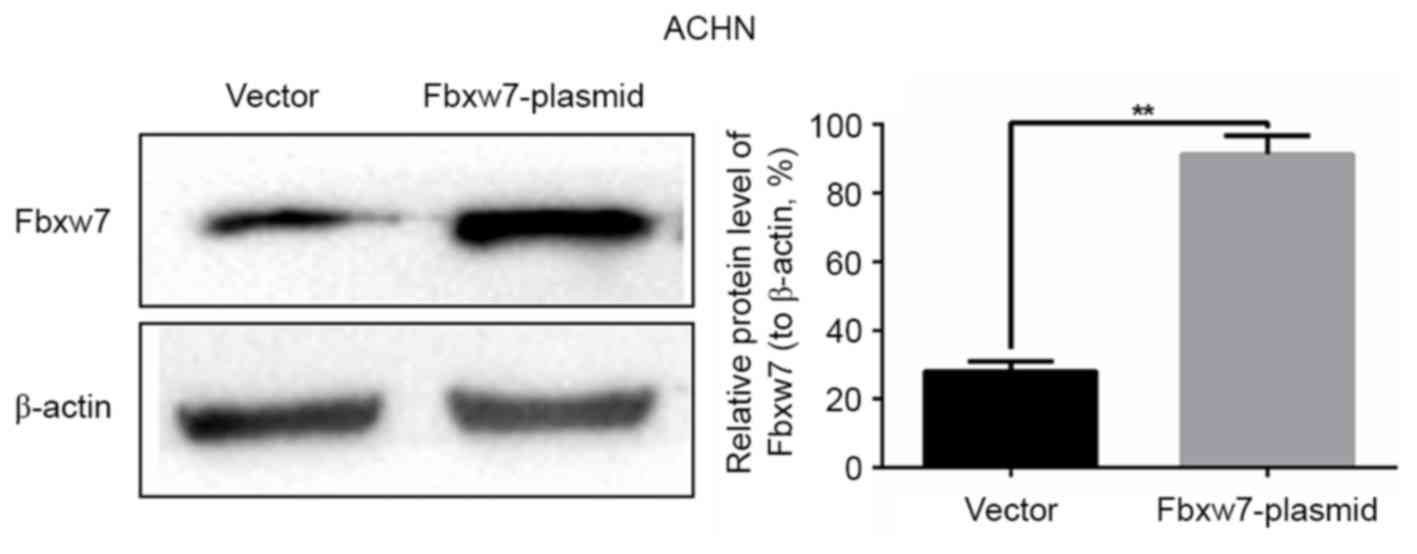
expression of Fbxw7 in the 786-O cell line was downregulated using
Fbxw7-siRNA and upregulated in the ACHN cell line using an Fbxw7
expressing plasmid. Their respective negative controls were also
established. Changes in Fbxw7 expression in the two cell lines
following transfection were confirmed by western blotting (Figs. 3 and 4).
Subsequently, wound healing and Transwell migration assays were
used to assess the migration ability of transfected cells and the
Transwell invasion assay was employed to assess the invasive
ability of the cells.
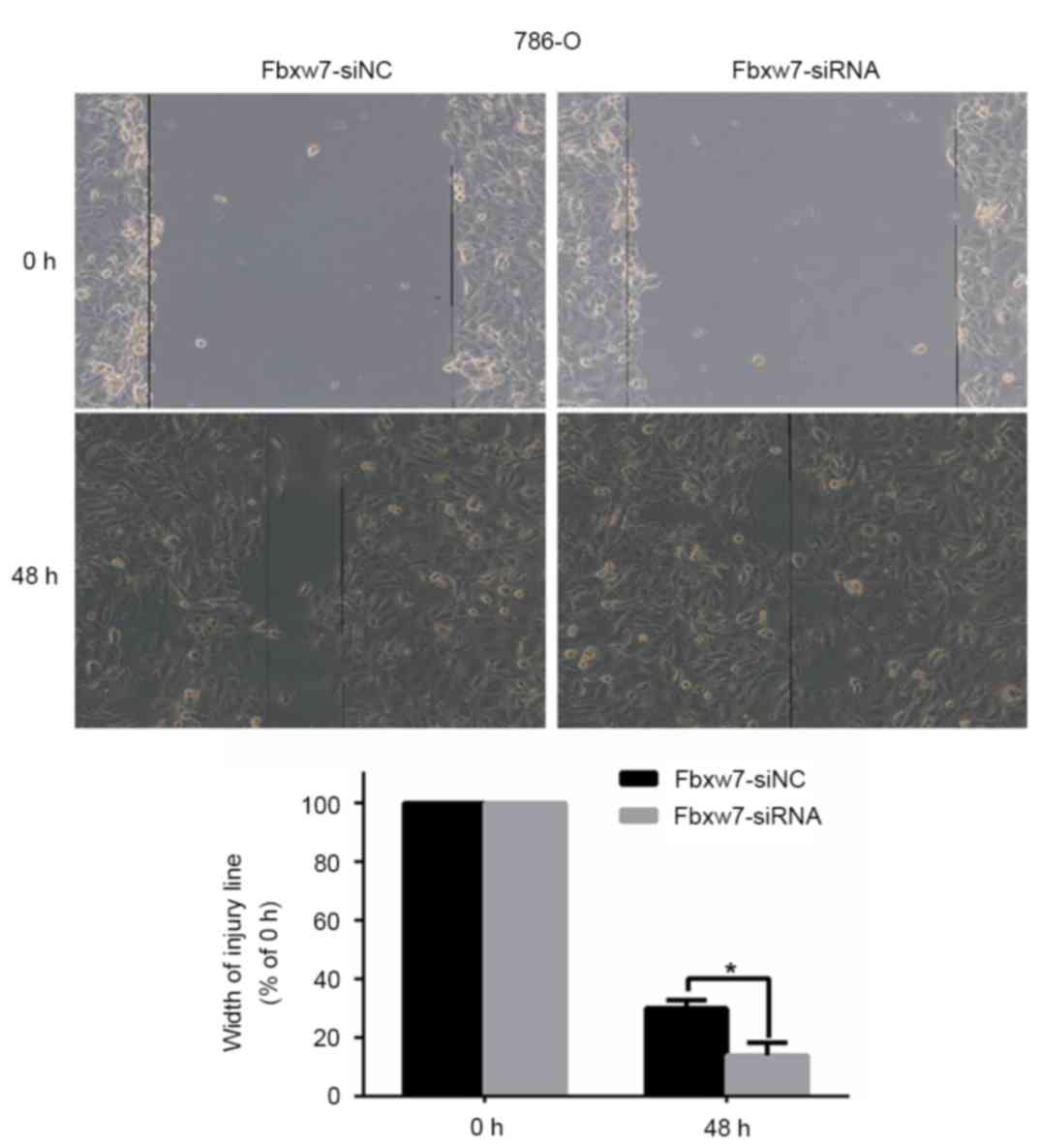
To determine whether Fbxw7 affects RCC cell
migration, a wound healing assay was performed, which revealed that
Fbxw7 silencing significantly reduced the wound healing time in
786-O cells compared with negative controls at 48 h (P=0.047;
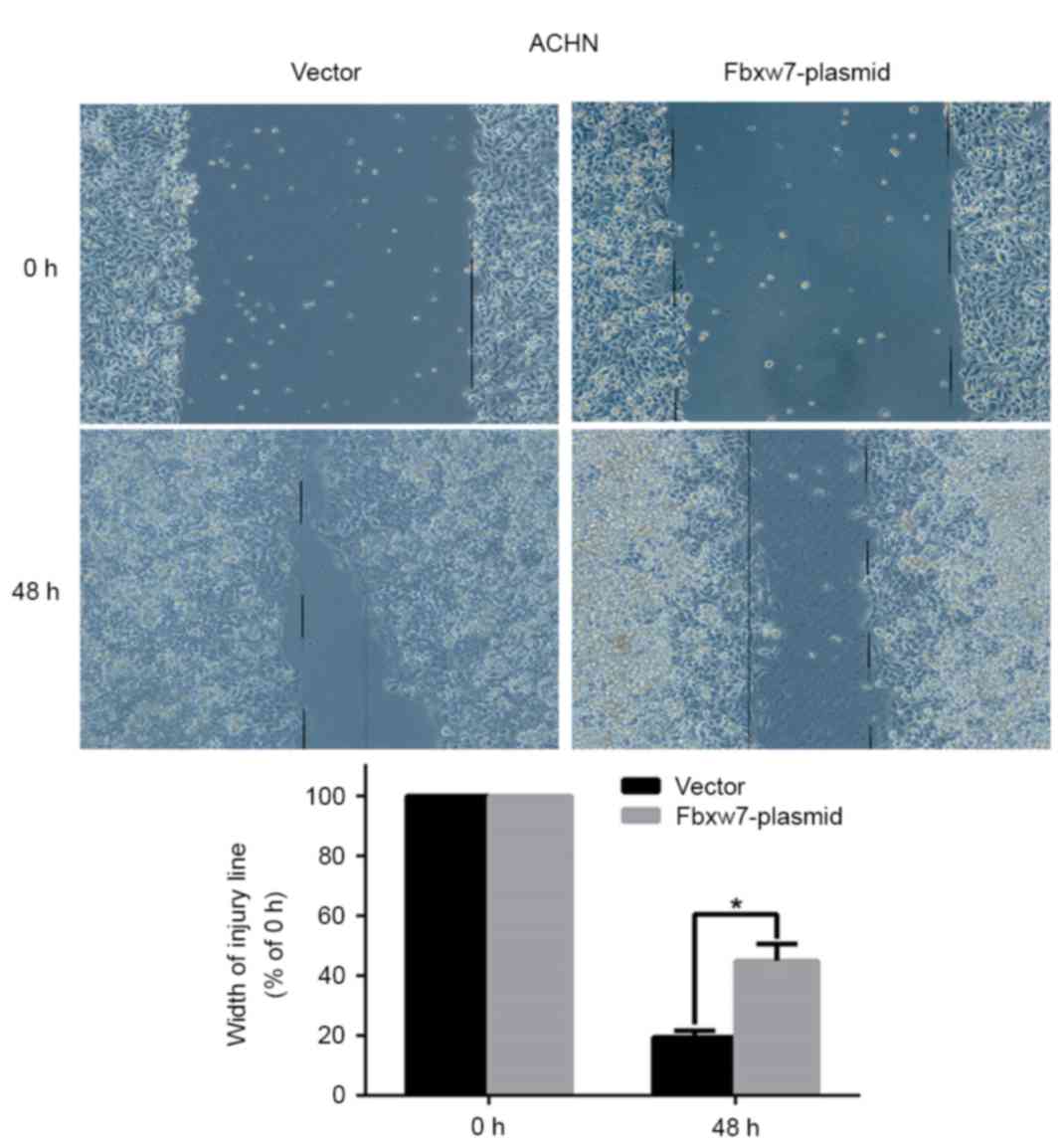
Fig. 5). By contrast, the wound
healing time in ACHN cells significantly increased following Fbxw7
overexpression compared with cells in the control group at 48 h
(P=0.027; Fig. 6). Furthermore, a
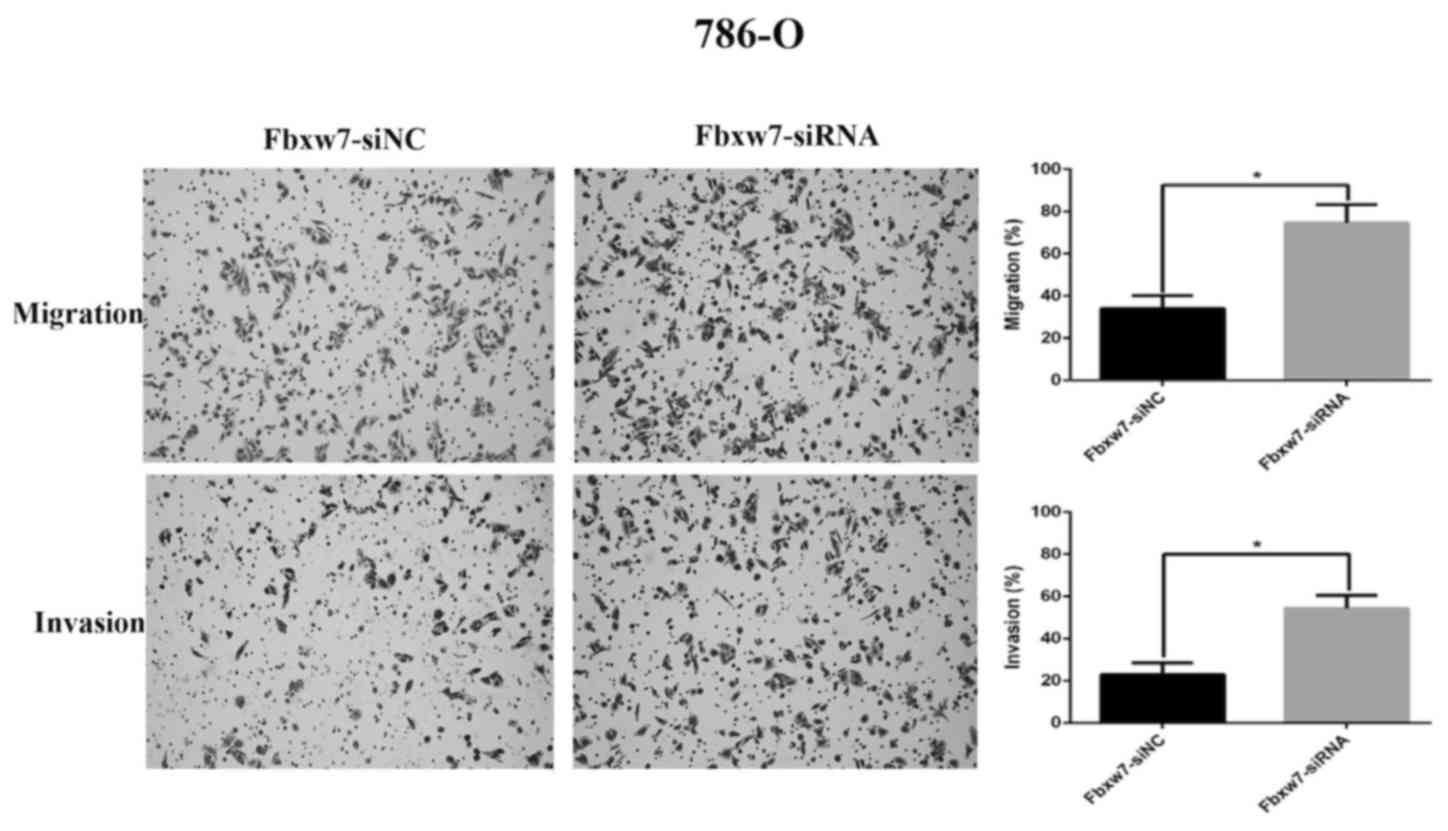
Transwell assay without Matrigel was performed, which revealed that
the number of Fbxw7-siRNA-transfected 786-O cells migrating through
the membrane partition into the lower chamber was significantly
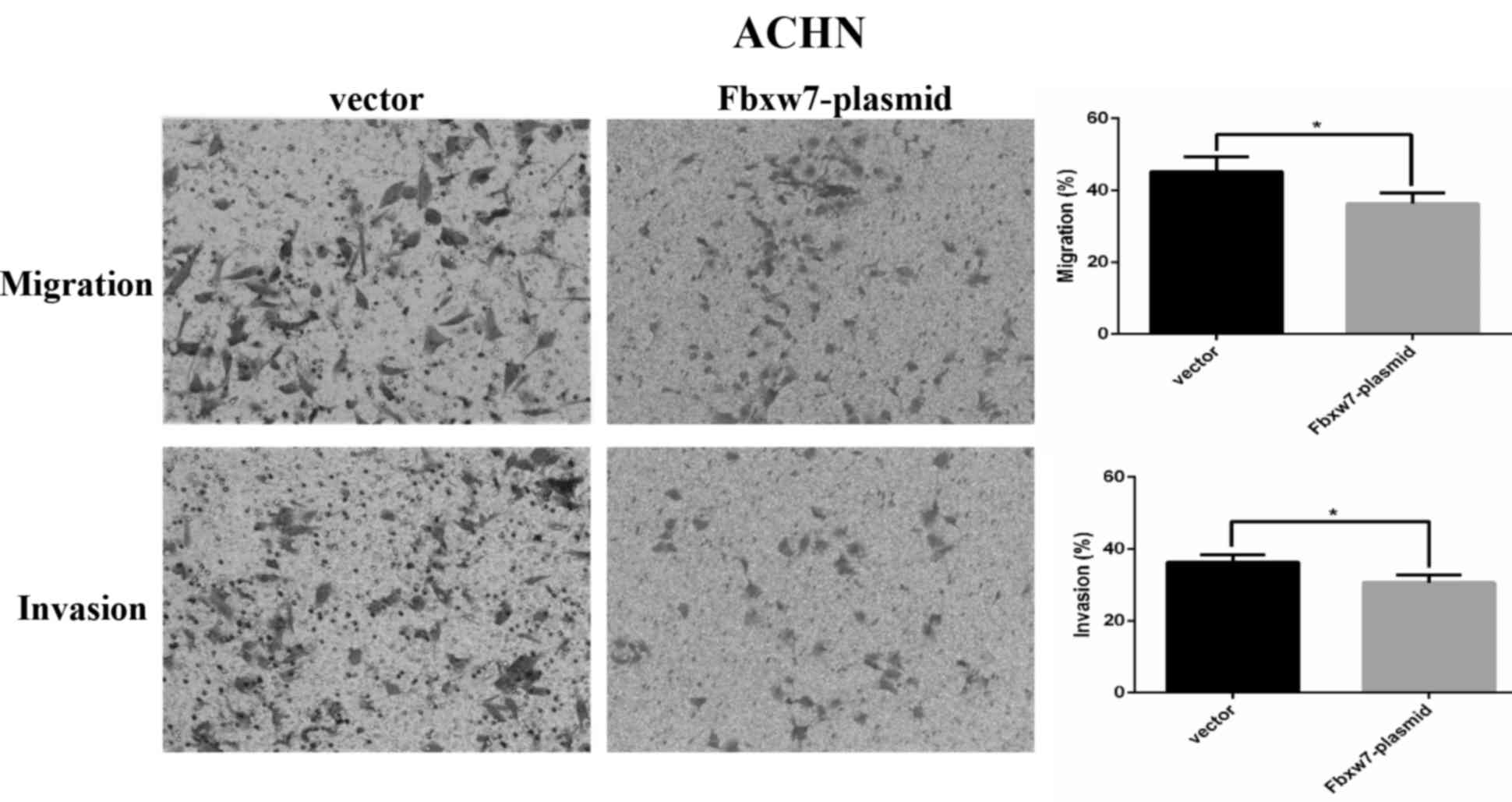
higher than the vector group (P=0.0027; Fig. 7). By contrast, Fbxw7-overexpressing
ACHN cells exhibited a significantly lower migration rate compared
with the control group (P=0.037; Fig.
8).
A Transwell assay with Matrigel was also performed
to elucidate the function of Fbxw7 in tumor cell invasion. There
were significantly more 786-O cells invading through the Matrigel
in the Fbxw7-siRNA group than in the Fbxw7-NC siRNA group
(P=0.0028; Fig. 7). However, elevated
Fbxw7 expression significantly reduced the number of ACHN cells
that invaded through the Matrigel compared with the control cells
(P=0.029; Fig. 8). These results
indicate that Fbxw7 inhibits the migration and invasion of RCC
cells.
Fbxw7 regulates the EMT in RCC
cells
From the aforementioned results, it was concluded
that the decreased expression of Fbxw7 stimulates the metastasis of
RCC cells. Therefore, it was investigated whether Fbxw7 influences
the invasive behavior of RCC cells via regulation of the EMT.
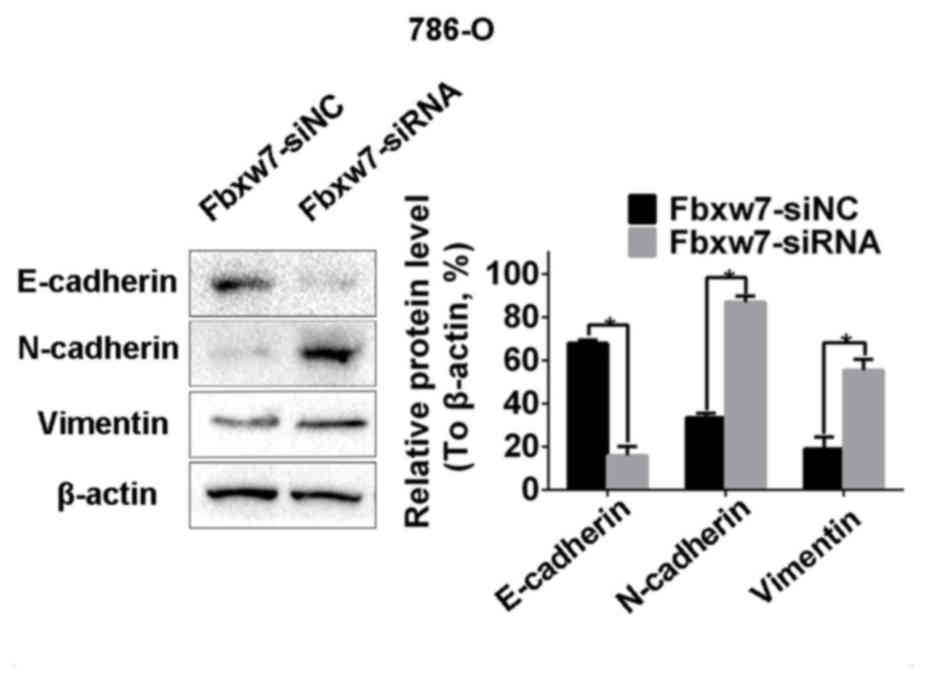
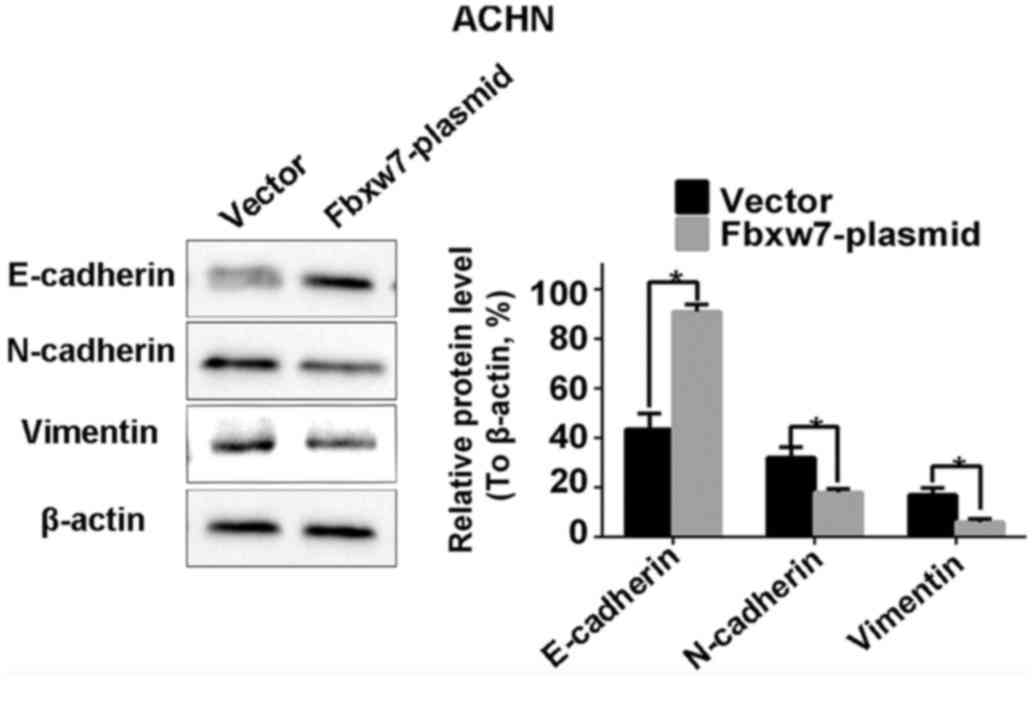
Western blotting identified a significant decrease in the
expression of the epithelial marker protein E-cadherin in 786-O
cells following Fbxw7 silencing. It also identified a significant
increase in the expression of the mesenchymal marker proteins
N-cadherin and vimentin (P<0.05; Fig.
9). By contrast, the overexpression of Fbxw7 in ACHN cells
significantly increased E-cadherin expression and significantly
reduced the expression of N-cadherin and vimentin (P<0.05;
Fig. 10). This suggests that Fbxw7
inhibits the EMT. Overall, these results suggest that the
downregulation of Fbxw7 in RCC accelerates cell migration and
invasion by inducing the EMT.
Discussion
RCC is a common human malignancy, accounting for
2–3% of adult malignant tumors. Treatment is effective when RCC is
limited to the kidney and the postoperative 5-year survival rate is
60–80% in such cases. However, following the onset of metastasis,
the prognosis of patients worsens and such patients exhibit a
median survival time of ~10 months. Therefore, metastasis is an
important cause of poor prognosis (4). However, the mechanisms underlying the
progression and metastasis of RCC remain unclear.
Previous studies have demonstrated that Fbxw7 serves
a key role as a suppressor gene in the cell cycle, proliferation,
differentiation, apoptosis, tumor metastasis and drug resistance
(11,12). Clinical and basic studies have
demonstrated that the decreased expression or mutations of Fbxw7
are common in all types of human tumors, including T cell acute
lymphoblastic leukemia, pancreatic, gastric, colorectal and
prostate cancer, cholangiocarcinoma and endometrial cancer
(11,19–21), with
an overall mutation rate of ~6% (19,27). In
addition, there are a number of studies investigating the role of
Fbxw7 in tumor cell migration and metastasis, including in gastric
cancer, hepatocellular carcinoma and hepatocholangiocarcinoma
(22,23). It has been demonstrated that the level
of Fbxw7 is decreased in renal clear cancer cells and that the
ectopic expression of Fbxw7 may inhibit the proliferation and
induce the apoptosis of tumor cells (26). In the present study, the role of Fbxw7
in RCC cell migration and invasion was investigated. Initially, the
expression of Fbxw7 in RCC tissues was detected by IHC and in
combination with clinical pathological parameters, the results
indicated that low Fbxw7 expression in tumor tissues was
significantly associated with tumor stage, lymph node and distant
metastases. In addition, clinical prognostic analysis revealed a
significantly improved OS rate in patients with high expression of
Fbxw7. Additionally, multivariate Cox regression analyses
demonstrated that the low expression of Fbxw7 was associated with
poor patient prognosis and may be an independent predictor of poor
prognosis for patients with RCC. Due to the limited variables in
the current study, multivariate analysis cannot control all the
potential confounders. Therefore, potential limitations existed in
the results that require verification in further studies. Although
these observations highlight the role of Fbxw7 in the progression
of RCC, to the best of our knowledge, the current study is the
first to investigate this function of Fbxw7 and its precise
mechanisms in the regulation of migration and invasion in RCC
cells. Therefore, the results of the current study indicate that
Gbxw7 may be used to diagnose and treat patients with RCC.
In order to verify the influence of Fbxw7 on the
ability of RCC cells to invade and migrate, Fbxw7 knockdown and
overexpression were performed in 786-O and ACHN cells,
respectively. The migration and invasion of tumor cells was
evaluated using Transwell and wound healing assays. The Transwell
assay is a common method to detect the ability of cell invasion and
migration in vitro. The principle is that cells in the upper
chamber are cultured in a serum-free medium and culture medium
containing 10% FBS is added to the lower chamber (28). There are two main reasons why low
nutrient culture medium is added to the upper chamber: Cells in the
relatively low nutrient culture exhibit a reduction in
proliferation; by contrast, tumor cells are spontaneously move to
the lower chamber containing higher FBS, which acts as a
chemoattractant. Under the same conditions, more tumor cells with a
higher migration ability may migrate to the lower chamber than
tumor cells and with a weaker ability to migrate. Adding Matrigel
to the upper chamber means that tumor cells must degrade the
Matrigel through the chamber to simulate the metastatic process to
allow tumor cells to invade through the basement membrane.
Therefore, it can evaluate the invasive ability of cells. In the
present study, the results of the wound healing and Transwell
migratory assays revealed that the migratory ability of 786-O cells
following Fbxw7 knockdown increased significantly. Furthermore, the
results of the Transwell invasion assay demonstrated that the
invasive ability of 786-O cells increased significantly following
Fbxw7 knockdown. By contrast, the invasion and migration ability of
ACHN cells with high levels of Fbxw7 was evidently weakened. These
results indicate that Fbxw7 inhibits RCC cell invasion and
metastasis. To the best of our knowledge, the present study is the
first to report that Fbxw7 downregulation promotes the invasion and
metastasis of RCC cells.
It has been suggested that tumor metastasis is a
multi-step biochemical process with numerous molecular events
(29). Firstly, adhesion of tumor
cells decreases, resulting in tumor cells separating from each
other. Subsequently, tumor cells adhere to the extracellular matrix
by binding to receptors on the cell surface; eventually, tumor
cells may migrate and invade to the surrounding tissues by
degrading the extracellular matrix (30,31). The
proliferation, adhesion and migration of tumor cells serve an
important role during the different stages of invasion. The EMT is
one of the most important mechanisms of tumor invasion and
metastasis in which the adhesion protein E-cadherin is critical. In
the present study, enhanced Fbxw7 expression significantly
attenuated the migration and invasion of RCC cells. Additionally,
when Fbxw7 expression was repressed using a specific siRNA, the
migration and invasion of tumor cells was significantly increased.
Furthermore, the association between Fbxw7 and the EMT process was
also explored. The results revealed that the EMT phenotypes were
reversed in the RCC cells following overexpression of Fbxw7 in ACHN
cells or knockdown of Fbxw7 in 786-O cells, as indicated by the
altered expression of epithelial and mesenchymal biomarkers. These
results reveal that Fbxw7 regulates RCC cell metastasis by
modulating the EMT. Consistent with the data analysis of the
clinical characteristics in the present study, these results
support the hypothesis that Fbxw7 serves a significant role in
inhibiting the migration and invasion of RCC cells.
In conclusion, the present study demonstrated that
the decreased expression of Fbxw7 is associated with RCC
progression and poor patient prognosis and indicated that its
underlying mechanism of action is associated with regulating the
EMT. Therefore, based on the results of in vitro experiments
investigating the migration and invasion of RCC cells, the results
of the present study indicate that Fbxw7 may be used as a potential
target against tumor metastasis in RCC. Furthermore Fbxw7 may be
used as an additional indicator to improve the prognosis of
patients with RCC. However, the function of Fbxw7 in RCC remains
unclear and further studies are required.
Glossary
Abbreviations
Abbreviations:
|
RCC
|
renal cell carcinoma
|
|
EMT
|
epithelial-mesenchymal transition
|
|
Fbxw7
|
F-box and WD repeat domain containing
7
|
|
IHC
|
immunohistochemistry
|
|
siRNA
|
small interfering RNA
|
|
OS
|
overall survival
|
References
|
1
|
Siegel R, DeSantis C, Virgo K, Stein K,
Mariotto A, Smith T, Cooper D, Gansler T, Lerr C, Fedewa S, et al:
Cancer treatment and survivorship statistics, 2012. CA Cancer J
Clin. 62:220–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun BL, Chen L, Fu H, Guo L, Guo H and
Zhang N: Upregulation of RICTOR gene transcription by the
proinflammatory cytokines through NF-κB pathway contributes to the
metastasis of renal cell carcinoma. Tumour Biol. 37:4457–4466.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Finley DS, Pantuck AJ and Belldegrun AS:
Tumor biology and prognostic factors in renal cell carcinoma.
Oncologist. 16 Suppl 2:S4–S13. 2011. View Article : Google Scholar
|
|
4
|
Xiong J, Liu Y, Jiang L, Zeng Y and Tang
W: High expression of long non-coding RNA lncRNA-ATB is correlated
with metastases and promotes cell migration and invasion in renal
cell carcinoma. Jpn J Clin Oncol. 46:378–384. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song J, Chen X, Bai J, Liu Q, Li H, Xie J,
Jing H and Zheng J: Discoidin domain receptor 1 (DDR1), a promising
biomarker, induces epithelial to mesenchymal transition in renal
cancer cells. Tumour Biol. 37:11509–11521. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lauffenburger D and Horwitz AF: Cell
migration: A physically integrated molecular process. Cell.
84:359–369. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee J: The epithelial-mesenchymal
transition: New insights in signaling, development and disease. J
Cell Biol. 172:973–981. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Q, Hou L, Ding G, Li Y, Wang J, Qian B,
Sun J and Wang Q: KDM6B induces epithelial-mesenchymal transition
and enhances clear cell renal cell carcinoma metastasis through the
activation of SLUG. Int J Clin Exp Pathol. 8:6334–6344.
2015.PubMed/NCBI
|
|
10
|
Wang Z, Inuzuka H, Fukushima H, Wan L, Gao
D, Shaik S, Sarkar FH and Wei W: Emerging roles of the FBW7 tumour
suppressor in stem cell differentiation. EMBO Res. 13:36–43. 2011.
View Article : Google Scholar
|
|
11
|
Welcker M and Clurman BE: FBW7 ubiquitin
ligase: A tumour suppressor at the crossroads of cell division,
growth and differentiation. Nat Rev Cancer. 8:83–93. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Z, Inuzuka H, Zhong J, Wan L,
Fukushima H, Sarkar FH and Wei W: Tumor suppressor functions of
FBW7 in cancer development and progression. FEBS Lett.
586:1409–1418. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Koepp DM, Schaefer LK, Ye X, Keyomarsi K,
Chu C, Harper JW and Elledge SJ: Phosphorylation-dependent
ubiquitination of cyclin E by the SCFFbw7 ubiquitin ligase.
Science. 294:173–177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei W, Jin J, Schlisio S, Harper JW and
Kaelin WG Jr: The v-Jun point mutation allows c-Jun to escape
GSK3-dependent recognition and destruction by the Fbw7 ubiquitin
ligase. Cancer Cell. 8:25–33. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Welcker M, Orian A, Jin J, Grim JE, Harper
JW, Eisenman RN and Clurman BE: The Fbw7 tumor suppressor regulates
glycogen synthase kinase 3 phosphorylation-dependent c-Myc protein
degradation. Proc Natl Acad Sci USA. 101:pp. 9085–9090. 2004;
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mao JH, Kim IJ, Wu D, Climent J, Kang HC,
DelRosario R and Balmain A: FBXW7 targets mTOR for degradation and
cooperates with PTEN in tumor suppression. Science. 321:1499–1502.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gupta-Rossi N, Le Bail O, Gonen H, Brou C,
Logeat F, Six E, Ciechanover A and Israël A: Functional interaction
between SEL-10, an F-box protein, and the nuclear form of activated
Notch1 receptor. J Biol Chem. 276:34371–34378. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thompson BJ, Buonamici S, Sulis ML,
Palomero T, Vilimas T, Basso G, Ferrando A and Aifantis I: The
SCFFBW7 ubiquitin ligase complex as a tumor suppressor in T cell
leukemia. J Exp Med. 204:1825–1835. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Akhoondi S, Sun D, von der Lehr N,
Apostolidou S, Klotz K, Maljukova A, Cepeda D, Fiegl H, Dafou D,
Marth C, et al: FBXW7/hCDC4 is a general tumor suppressor in human
cancer. Cancer Res. 67:9006–9012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hubalek MM, Widschwendter A, Erdel M,
Gschwendtner A, Fiegl HM, Müller HM, Goebel G, Mueller-Holzner E,
Marth C, Spruck CH, et al: Cyclin E dysregulation and chromosomal
instability in endometrial cancer. Oncogene. 23:4187–4192. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee JW, Soung YH, Kim HJ, Park WS, Nam SW,
Kim SH, Lee JY, Yoo NJ and Lee SH: Mutational analysis of the hCDC4
gene in gastric carcinomas. Eur J Cancer. 42:2369–2373. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li J, Guo Y, Liang X, Sun M, Wang G, De W
and Wu W: MicroRNA-223 functions as an oncogene in human gastric
cancer by targeting FBXW7/hCdc4. J Cancer Res Clin Oncol.
138:763–774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng Y, Chen G, Martinka M, Ho V and Li
G: Prognostic significance of Fbw7 in human melanoma and its role
in cell migration. J Invest Dermatol. 133:1794–1802. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Santiago JM, Sasako M and Osorio J:
TNM-7th edition 2009 (UICC/AJCC) and Japanese Classification 2010
in Gastric Cancer. Towards simplicity and standardisation in the
management of gastric cancer. Cir Esp. 89:275–281. 2011.(In
Spanish).
|
|
25
|
Xu W, Wang Z, Zhang W, Qian K, Li H, Kong
D, Li Y and Tang Y: Mutated K-ras activates CDK8 to stimulate the
epithelial-to-mesenchymal transition in pancreatic cancer in part
via the Wnt/β-catenin signaling pathwa. Cancer Lett. 356:613–627.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fu Y, Lin Y, Yang Z, Yang G, Li G, Liu Y,
Tan X, Huang Y, Wu X, Wang Y, et al: FBXW7 overexpression
suppresses renal cancer cell proliferation and induces apoptosis.
Med Oncol. 32:2152015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Liu Y, Lu J, Zhang P, Wang Y, Xu
Y, Wang Z, Mao JH and Wei G: Rapamycin inhibits FBXW7 loss-induced
epithelial-mesenchymal transition and cancer stem cell-like
characteristics in colorectal cancer cells. Biochem Biophys Res
Commun. 434:352–356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao J, Liu J, Xu R, Zhu X, Liu L and Zhao
X: MicroRNA-21 stimulates epithelial-to-mesenchymal transition and
tumorigenesis in clear cell renal cells. Mol Med Rep. 13:75–82.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Song Y, Washington MK and Crawford HC:
Loss of FOXA1/2 is essential for the epithelial-to-mesenchymal
transition in pancreatic cancer. Cancer Res. 70:2115–2125. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schmalhofer O, Brabletz S and Brabletz T:
E-cadherin, beta-catenin, and ZEB1 in malignant progression of
cancer. Cancer Metastasis Rev. 28:151–166. 2009. View Article : Google Scholar : PubMed/NCBI
|