Introduction
All mature blood cells are derived from
hematopoietic stem cells, and this process is known as
hematopoiesis. In a healthy adult, 1011-1012
new blood cells are produced daily in order to maintain a steady
state in the peripheral circulation (1). Granulocytes are a class of white blood
cells marked with the presence of granules in their cytoplasm,
which serve important roles in the innate immune system and are
involved in chemotaxis, phagocytosis and bactericidal action
(2). Monocytes are the largest of all
leukocytes, characterized by a granulated cytoplasm. Monocytes also
have multiple roles in immune function, including replenishing
resident macrophages and differentiating into macrophages and
dendritic cells to elicit an immune response (3,4).
Granulocytes and monocytes are derived from the common myeloid
progenitors. Granulocytic and macrophage-like differentiation and
maturation are important components of normal hematopoiesis
(5). Blocking myeloid differentiation
results in acute myeloid leukemia (AML). AML is characterized by
the reduced production of functional blood cells and overproduction
and accumulation of immature white blood cells. The symptoms of AML
include fatigue, shortness of breath, easy bruising and bleeding
and an increased risk of infection (6). AML is the most common type of acute
leukemia affecting adults, and the incidence of AML increases with
age (7,8). The French-American-British (FAB)
classifies AML into eight subtypes (M0-M7) according to cell
morphology and maturation (9). From
M1 to M5, AML cases involving granulocytic and macrophage-like
differentiation blockage account for 85% of adult AML cases
(9,10). Granulocytic and monocytic
differentiation is regulated by multiple factors, including key
transcriptional factors (PU.1, CCAAT/enhancer-binding protein α,
interferon regulatory factor 8, TAL bHLH transcription factor 1 and
runt-related transcription factor 1) (11), cytokines [granulocyte
colony-stimulating factor (CSF), macrophage-CSF and interleukin-3]
(12) and non-coding RNAs (13).
MicroRNAs (miRNA) are a class of small non-coding
RNAs, which regulate gene expression post-transcriptionally through
degradation of mRNA or inhibition of mRNA translation. miRNAs have
been reported to be involved in numerous physiological and
pathological biological processes (14). A number of miRNAs have been indicated
to be involved in myeloid differentiation and AML progression,
including the miR-223, miR-142-3p, miR-29 and miR-181 families
(15–17). The miR-10 family consists of two
members, miR-10a and miR-10b, which are located at chromosome 17
and 2, respectively (18).
Accumulating evidence suggests that miR-10a/b may act as novel
oncogenes in various types of human cancer, including metastatic
breast cancer, pancreatic cancer, esophageal cancer, hepatocellular
carcinoma, nasopharyngeal carcinoma and colorectal cancer (19–28). These
findings suggest that miR-10a and miR-10b are strongly expressed in
highly metastatic cancer and serve important roles in cancer
metastasis. miR-10a and miR-10b have also been reported to be
upregulated in nucleophosmin (NPM1)-mutated AMLs (29,30).
However, the regulatory role of miR-10a/b in AML remains
unknown.
In the present study, the expression of miR-10a and
miR-10b was examined in a number of AML cases and healthy donor
controls. It was revealed that the expression of miR-10a and
miR-10b was increased in AML cases compared with healthy controls,
particularly in M1, M2 and M3 subtypes. Additionally, the levels of
miR-10a and miR-10b were upregulated in AML patients with mutated
NPM1, and also AML patients with t(8;21) and t(9;11), compared with
the normal controls. Furthermore, the roles of miR-10a and miR-10b
in regulating granulocytic and monocytic differentiation and AML
progression were investigated. The results indicated that miR-10a
and miR-10b were able to promote the proliferation of human
promyelocytic leukemia HL-60 cells, while suppressing the
granulocytic and monocytic differentiation of HL-60 cells. These
findings suggested that abnormal high expression of miR-10a and
miR-10b may result in unlimited proliferation of immature blood
progenitors and the repression of mature blood cell differentiation
and maturation, thus leading to the occurrence of AML. Therefore,
miR-10a and miR10b may be developed as new therapeutic targets of
AML.
Materials and methods
Human samples
The peripheral blood samples of patients with AML
(89 patients) and normal volunteers (65 patients) were obtained
from The First Affiliated Hospital of Wenzhou Medical University
(Nanbaixiang, Ouhai, Wenzhou, Zhejiang) from January to December
2015. The gender ratio of the patients and controls was
approximately 1:1 and aged between 2–60 years old. Written informed
consent to perform the biological studies was obtained from all
examined patients. The present study was approved by the Ethics
Committees of The First Affiliated Hospital of Wenzhou Medical
University. Mononuclear cells were isolated from the peripheral
blood samples of patients with AML and normal volunteers using
lymphocyte separation medium (Lonza Walkersville, Inc.,
Walkersville, MD, USA). The peripheral blood samples were first
diluted with phosphate-buffered saline (PBS; 1:4) and then plated
on the lymphocyte separation medium. The middle layer of
mononuclear cells was collected following centrifugation at 1,000 ×
g for 30 min at room temperature.
Cell culture and transfections
The human leukemia HL-60 cells (ATCC, Manassas, VA,
USA) were grown in Iscove's modified Dulbecco's medium (Invitrogen;
Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.,), 50
U/ml penicillin and 50 µg/ml streptomycin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) at 37°C in a 5% CO2 cell
culture incubator. All-trans-retinoic acid (ATRA; 50 ng/ml;
Sigma-Aldrich; Merck KGaA) and phorbol myristate acetate (PMA; 2
µM; Sigma-Aldrich; Merck KGaA) were used to induce HL-60 cells to
undergo granulocytic and monocytic differentiation. miR-10a mimics,
miR-10b mimics and scramble control were obtained from Dharmacon
(GE Healthcare Life Sciences, Little Chalfont, UK) and transfected
with DharmFECT1 (Dharmacon; GE Healthcare Life Sciences) into HL-60
cells at a final concentration of 50 nM.
May-Grünwald Giemsa staining
HL-60 cells undergoing granulocytic and monocytic
differentiation were harvested at indicated time. Cells were
smeared on glass slides, fixed in 100% methanol for 10 min at room
temperature, then stained with May-Grünwald/Giemsa (cat. no.
BA-4017; BaSO Diagnostics Inc., Zhuhai, China) for 5 min at room
temperature, and analyzed at ×400 magnification under an inverted
microscope (Nikon TE2000; Nikon Corporation, Tokyo, Japan) equipped
with a digital camera.
RNA extraction, cDNA synthesis and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) assays
Total RNA was extracted from cultured cells and
mononuclear cells isolated from peripheral blood using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. cDNA was synthesized using
high-capacity cDNA reverse transcription Kit (Thermo Fisher
Scientific, Inc.) from 1–5 µg of total RNA. For detection of
microRNA expression, a stem-loop RT primer was used for the reverse
transcription of miR-10a/b. For detection of mRNA expression,
oligodT primer was used for the reverse transcription of mRNA.
RT-qPCR using SYBR® Green qPCR Master Mix (Takara Bio,
Inc., Otsu, Japan) was performed in a Bio-Rad CFX96 real-time PCR
System (Bio-Rad Laboratories, Inc., CA, USA) according to the
manufacturer's protocols. The PCR conditions were as follows: 95°C
for 30 sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 34
sec. The data were normalized using the endogenous U6 small nuclear
RNA and GAPDH. The 2−ΔΔCq method (31) was used in the analysis of PCR data.
Primer sequences are presented in Table
I.
 | Table I.Sequences of primers used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Sequences of primers used in reverse
transcription-quantitative polymerase chain reaction.
| Primer | Sequence
(5′-3′) |
|---|
| miR-10a-RT |
GTCGTATCCAGTGCAGGGTCCGAGG |
|
|
TATTCGCACTGGATACGACCACAAAT |
| miR-10b-RT |
GTCGTATCCAGTGCAGGGTCCGAGG |
|
|
TATTCGCACTGGATACGCACAAATT |
|
miR-10a-forward | GCGC
TACCCTGTAGATCCG |
|
miR-10a-reverse |
GTGCTACCCTGTAGAAC |
|
miR-10b-forward |
AGCTGTTCAGTGCACTACAGA |
|
miR-10b-reverse |
GTGCTACCCTGTAGAAC |
| miR-10a-probe |
FAM-CCTGTAGATCCGAATTTG-MGB |
| miR-10b-probe |
FAM-CCTGTAGAACCGAATTTG-MGB |
| U6-RT |
AAAATATGGAACGCTTCACGAATTTG |
| U6-forward |
CTCGCTTCGGCAGCACATATACT |
| U6-reverse |
ACGCTTCACGAATTTGCGTGTC |
| U6-probe |
FAM-CCATGCTAATCTTCTCTGTA-MGB |
| CD14-forward |
GACCTAAAGATAACCGGCACC |
| CD14-reverse |
GCAATGCTCAGTACCTTGAGG |
| CD11b-forward |
CAGACAGGAAGTAGCAGCTCCT |
| CD11b-reverse |
CTGGTCATGTTGATGAAGGTGCT |
| GAPDH-forward |
TCAACGACCACTTTGTCAAGCTCA |
| GAPDH-reverse |
GCTGGTGGTCCAGGGGTCTTACT |
| CSF1R-forward |
GGGAATCCCAGTGATAGAGCC |
| CSF1R-reverse |
TTGGAAGGTAGCGTTGTTGGT |
| CSF3R-forward |
TCAAGTTGGTGCTATGGCAAGG |
| CSF3R-reverse |
GCTCCCAGTCTCCACAGAATC |
Flow cytometry and
fluorescence-activated cell sorting analysis (FACS)
HL-60 cells were harvested at 48 h after
transfection and were washed twice at 4°C in PBS containing 0.5%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.). ATRA-
or PMA-induced HL-60 cells were incubated with
phycoerythrin-conjugated cluster of differentiation molecule 11B
(CD11b) antibody (CD11b04-4; eBioscience; Thermo Fisher Scientific,
Inc.) or fluorescein isothiocyanate-conjugated anti-cluster of
differentiation CD14 antibody (11-0141-82; eBioscience; Thermo
Fisher Scientific, Inc.) at 1:100 dilution for 30 min at room
temperature. Flow cytometry was performed using a C6 flow cytometer
instrument (BD Biosciences, Franklin Lakes, NJ, USA) and
subsequently analyzed with CFlow Sampler Analysis 1.0.208.2 (BD
Biosciences).
Cell proliferation assay
To measure the effect of miRNA mimics on
proliferation of HL-60 cells, the cells were incubated in 10% cell
counting kit-8 solution (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) and diluted in normal culture medium at 37°C until
color change has taken place. Proliferation rates were determined
on day 1, 2, 3 and 4 post-transfection, and quantification was
performed on a microtiter plate reader (Spectra Rainbow, Tecan
Trading AG, Männedorf, Switzerland) at OD450 nm according to the
manufacturer's protocol.
Statistical analysis
Comparisons between multiple groups were analyzed
using one-way analysis of variance and the least significant
difference post hoc test using GraphPad Prism 5 v5.01 (Graph Pad
Software, Inc., La Jolla, CA, USA). Comparisons between two groups
were evaluated by independent sample t-test (two-tailed). P≤0.05
was considered to indicate a statistically significant difference.
Data are presented as the mean ± standard deviation.
Results
miR-10a and miR-10b are significantly
increased in patients with AML
To accurately examine the expression of miR-10a and
miR-10b in AML, TaqMan quantitative PCR was used to detect the
levels of miR-10a and miR-10b in peripheral blood mononuclear cells
(PBMNCs) derived from 89 patients with AML and 65 healthy donors
(Fig. 1).
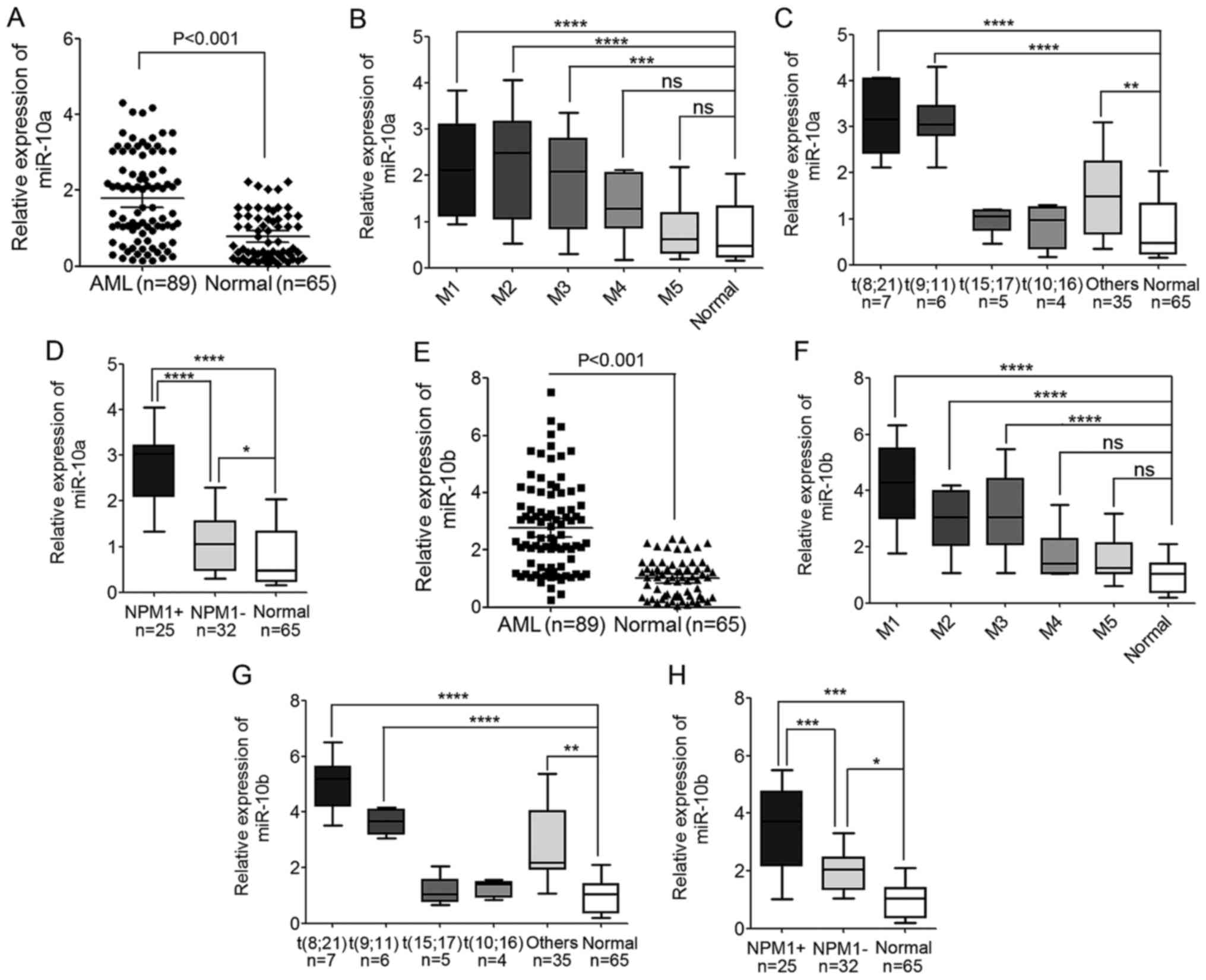 | Figure 1.Levels of miR-10a and miR-10b
expression are significantly increased in AML cases. (A) The
expression level of miR-10a in PBMNCs derived from 89 patients with
AML and 65 healthy donors detected by TaqMan quantitative
polymerase chain reaction. (B) The expression level of miR-10a in
different AML subtypes, from M1 to M5. (C) The expression level of
miR-10a in different WHO subtypes of AML patients compared with the
normal controls. (D) The expression level of miR-10a in AML
patients with mutated and wild-type NPM1 compared with the normal
controls. (E) The expression level of miR-10b in PBMNCs derived
from 89 AML patients and 65 healthy donors. (F) The expression
level of miR-10b in different AML subtypes, from M1 to M5. (G) The
expression level of miR-10b in different WHO subtypes of AML
patients compared with the normal controls. (H) The expression
level of miR-10b in AML patients with mutated and wild-type NPM1
compared with the normal controls. *P<0.05, **P<0.01,
***P<0.001, ****P<0.0001; ns, not significant. AML, acute
myeloid leukemia; miR, microRNA; NPM1, nucleophosmin; PBMNC,
peripheral blood mononuclear cell; WHO, World Health Organization;
NPM1+, NPM1 mutated; NPM1-, wild-type NPM1. |
Compared with the normal controls, the expression of
miR-10a increased significantly in AML cases (P<0.001; Fig. 1A). These AML cases that were analyzed
included major FAB subtypes from M1 to M5, including 13 cases of
M1, 28 cases of M2, 21 cases of M3, 10 cases of M4 and 17 cases of
M5. The expression level of miR-10a was also compared in the
different AML subtypes, and it was indicated that the expression of
miR-10a in M1, M2 and M3 AML subtypes was increased compared with
M4, M5 and normal controls. The expression of miR-10a in M4 and M5
subtypes was also higher compared with normal controls, but this
difference was not statistically significant (Fig. 1B).
In addition, the expression of miR-10a and miR-10b
based on cytogenetic karyotypes was also compared in 57 patients
with AML, whose cytogenetic karyotype and gene mutation had been
detected. The results showed that miR-10a was significantly
upregulated in AML patients with t(8;21) and t(9;11), but not in
patients with AML with t(15;17) and t(10;16) when compared with the
normal controls (Fig. 1C). In
addition, the expression level of miR-10a was also upregulated in
patients with NPM1-mutated AML compared with normal controls, and
patients with AML and wild-type NPM1 (Fig. 1D), which was consistent with previous
studies (29,30,32).
Similar to miR-10a expression, the expression of
miR-10b also increased significantly in AML cases compared with the
normal controls (P<0.001; Fig.
1E). The expression of miR-10b in M1, M2 and M3 AML subtypes
was higher compared with the expression in M4, M5 and normal
controls (Fig. 1F). Additionally, the
expression in M4 and M5 AML subtypes was also a little but not
significant higher compared with the expression in normal controls
(Fig. 1F). The expression level of
miR-10b expression was increased in patients with AML with t(8;21)
and t(9;11) compared with the expression in normal controls
(Fig. 1G). Additionally, the level of
miR-10b was significantly upregulated in patients with NPM1-mutated
AML compared with normal controls, and patients with AML and
wild-type NPM1 (Fig. 1H). These
results indicated the involvement of miR-10a and miR-10b in
leukemogenesis.
miR-10a and miR-10b are gradually
decreased during granulocytic and monocytic differentiation of
human leukemia cells
To examine the possible role of miR-10a and miR-10b
in granulocytic and monocytic differentiation and leukemogenesis,
the expression level of the two miRNAs was detected during
granulocytic and monocytic differentiation of human leukemia HL-60
cells. HL-60 was induced to undergo granulocytic or monocytic
differentiation using ATRA or PMA, respectively, as previously
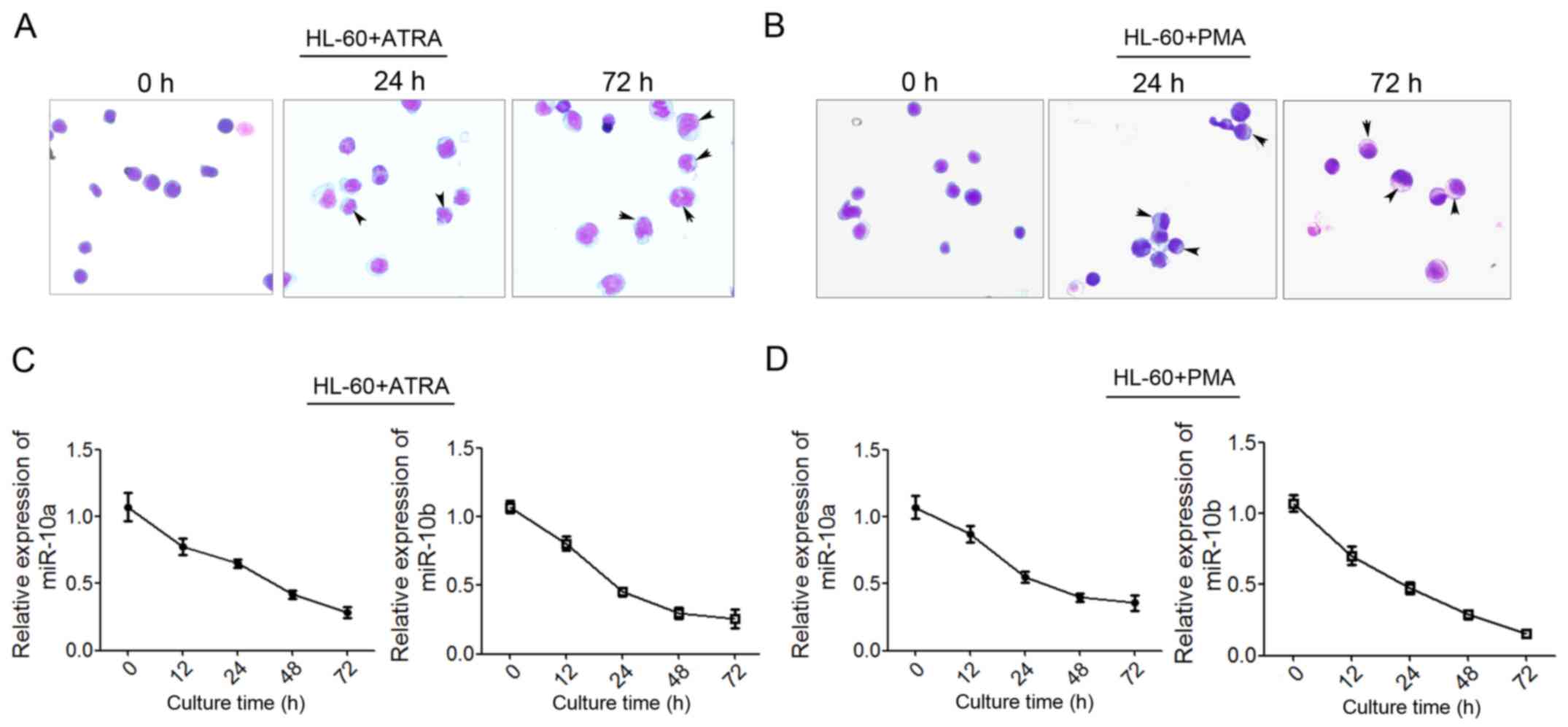
reported (33). As Giemsa staining
shown, more mature granulocytic cells show polylobular nuclei, more
mature monocytes show bluish-gray cytoplasm and a saddle-shaped
nucleus (Fig. 2A and B) qPCR was used
to detect the level of miR-10a and miR-10b expression at 12, 24, 48
and 72 h post-induction. The results showed that miR-10a/b
decreased significantly during ATRA-induced granulocytic
differentiation (Fig. 2C) and
PMA-induced monocytic differentiation (Fig. 2D). These findings suggested that
miR-10a and miR-10b may perform critical roles in
granulocytic/monocytic differentiation.
miR-10a promotes proliferation of
human leukemia cells and inhibits differentiation of human leukemia
cells to granulocytes and monocytes
Given the decrease in miR-10a and miR-10b expression
during granulocytic/monocytic differentiation of HL-60 cells, it
was questioned whether miR-10a and miR-10b regulate
granulocytic/monocytic differentiation or proliferation of leukemia
cells. miR-10a mimics were transfected into HL-60 cells. Cell
growth rate and differentiation were then examined. miR-10a was
successfully overexpressed in HL-60 cells as confirmed by qPCR
(Fig. 3A). Cell proliferation rates
were determined at days 1, 2, 3 and 4 post-transfection, and
overexpression of miR-10a in HL-60 cells was observed to be able to
significantly promote the proliferation of leukemia cells compared
with the scramble control (Fig.
3B).
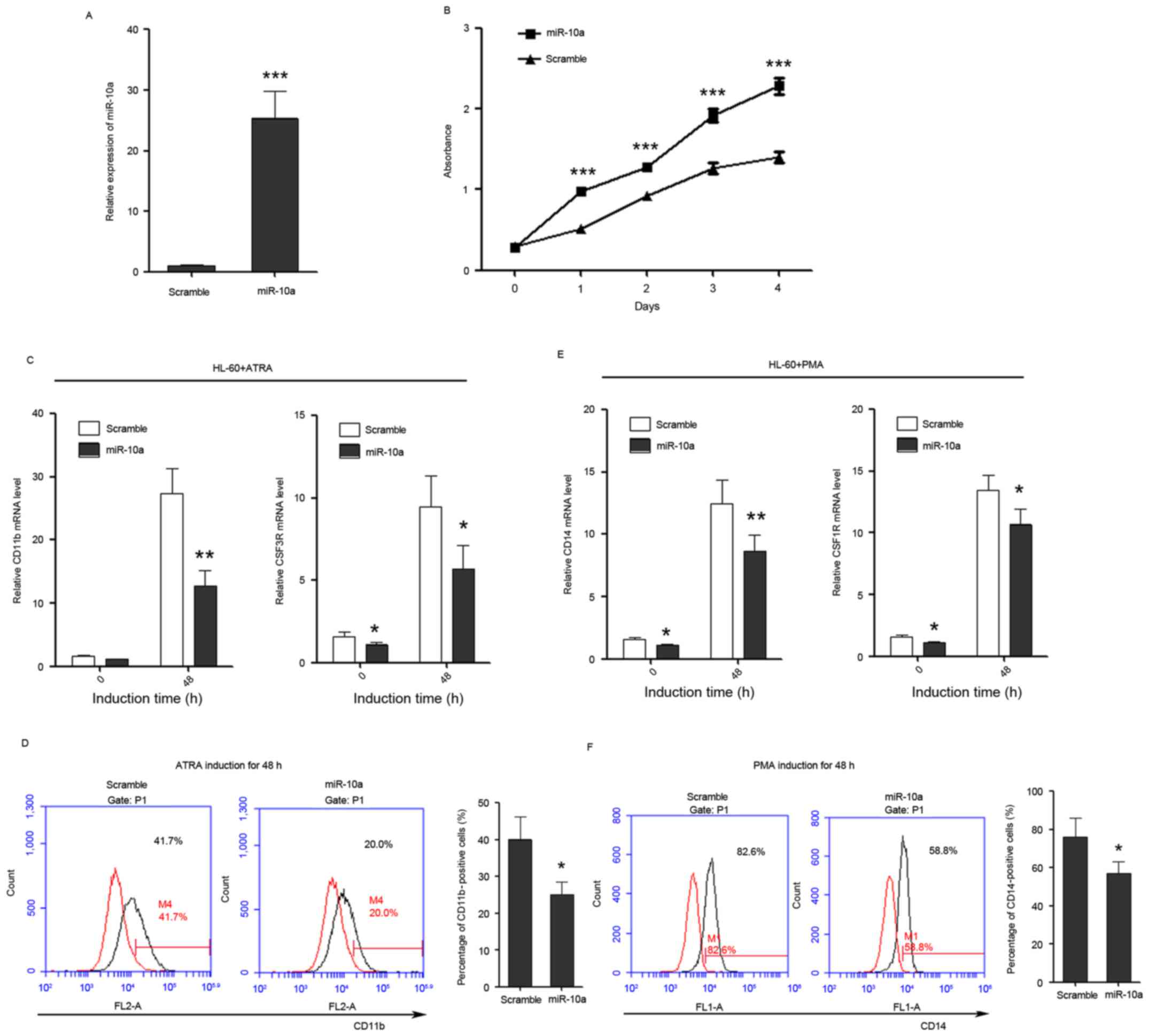 | Figure 3.miR-10a promotes the proliferation of
HL-60 cells, while inhibits the differentiation of HL-60 cells to
granulocytes and monocytes. (A) miR-10a is overexpressed in HL-60
cells as confirmed by quantitative polymerase chain reaction. (B)
The growth of HL-60 cells at day 0, 1, 2, 3 and 4
post-transfection, which was detected by Cell Counting Kit-8 assay.
(C) The expression level of granulocytic marker CD11b and CSF3R in
untreated and ATRA-treated (duration, 48 h) cells that overexpress
miR-10a. (D) CD11b FACS analysis showed that overexpression of
miR-10a was able to delay ATRA-induced granulocytic differentiation
of HL-60 cells. The percentage of CD11b-positive cells was
calculated using the black traces referring to cells stained with
CD11b antibody against the red traces referring to the same cells
not stained with CD11b antibody. (E) The relative expression of
monocytic marker CD14 and CSF1R in untreated and PMA-treated
(duration, 48 h) cells that overexpress miR-10a. (F) CD14 FACS
analysis showed that overexpression of miR-10a delayed PMA-induced
monocytic differentiation of HL-60 cells. The percentage of
CD14-positive cells was calculated in the same way. Data are
presented as the mean ± standard deviation (n=3). All the
comparisons are made between scramble and miR-10a groups.
*P<0.05; **P<0.01; ***P<0.001. miR, microRNA; CSF3R,
colony-stimulating factor 3 receptor; ATRA, all-trans-retinoic
acid; PMA, phorbol myristate acetate; CD, cluster of
differentiation; CSF1R, colony-stimulating factor 1 receptor; FACS,
fluorescence-activated cell sorting. |
In addition, the effect of miR-10a on the ability of
leukemia cells to differentiate into granulocyte and monocytes was
investigated. Transfected HL-60 cells were induced to undergo
granulocytic/monocytic differentiation, and the expression of CD11b
and CSF3 receptor (CSF3R) were used to estimate the progression of
granulocytic differentiation, while the expression of CD14 and CSF1
receptor (CSF1R) were used to estimate the progression of monocytic
differentiation. FACS analysis of CD11b and CD14 was also used to
examine the granulocytic/monocytic differentiation of HL-60 cells.
The results demonstrated that ectopic expression of miR-10a was
able to suppress the upregulation of CD11b and CSF3R during
ATRA-induced granulocytic differentiation (Fig. 3C) and also was able to decrease the
percentage of CD11b-positive cells (Fig.
3D). Ectopic expression of miR-10a also significantly inhibited
the upregulation of CD14 and CSF1R during PMA-induced monocytic
differentiation (Fig. 3E) and
decreased the percentage of CD14-positive cells (Fig. 3F), indicating that miR-10a performs a
negative role in granulocytic/monocytic differentiation of HL-60
cells.
miR-10b promotes cell proliferation
and suppresses granulocytic and monocytic differentiation
The role of miR-10b in regulating
granulocytic/monocytic differentiation and cell proliferation of
leukemia cells was also detected, in the same way as miR-10a.
miR-10b was successfully overexpressed in HL-60 cells as confirmed
by qPCR (Fig. 4A). Cell proliferation
rates were determined at days 1, 2, 3 and 4 post-transfection, and
ectopic expression of miR-10b was observed to promote the
proliferation of leukemia cells (Fig.
4B). In addition, it was revealed that overexpression of
miR-10b suppressed the upregulation of CD11b and CSF3R during
ATRA-induced granulocytic differentiation (Fig. 4C) and also decreased the percentage of
CD11b-positive cells (Fig. 4D).
Furthermore, the overexpression of miR-10b also significantly
inhibited the upregulation of CD14 and CSF1R during PMA-induced
monocytic differentiation (Fig. 4E)
and decreased the percentage of CD14-positive cells (Fig. 4F), indicating that miR-10b has a
negative role in granulocytic/monocytic differentiation of HL-60
cells.
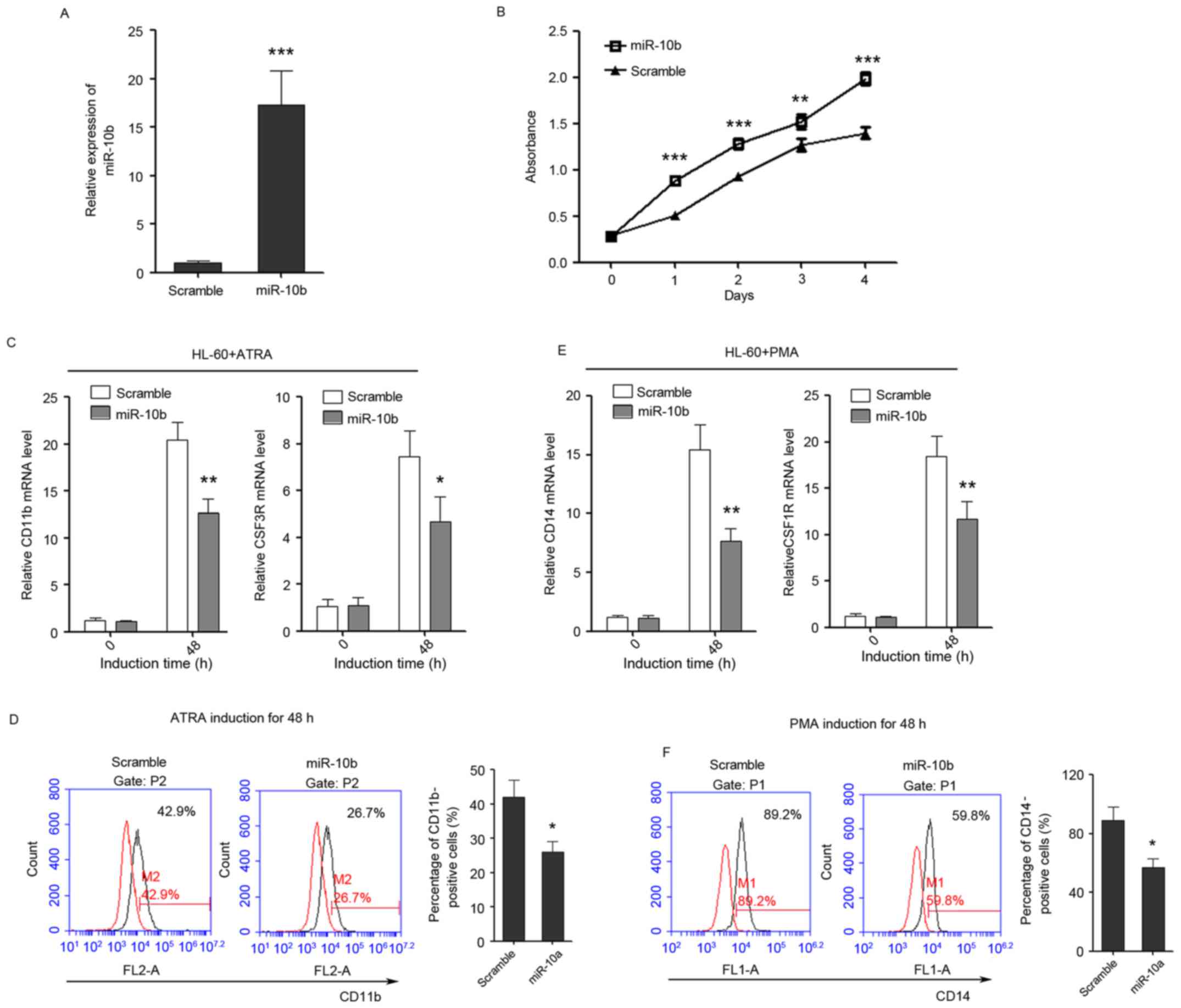 | Figure 4.miR-10b promotes the proliferation of
HL-60 cells, while inhibits the differentiation of HL-60 cells to
granulocytes and monocytes. (A) The overexpression of miR-10b in
HL-60 cells as confirmed by quantitative polymerase chain reaction.
(B) The growth of HL-60 cells at day 0, 1, 2, 3 and 4
post-transfection as detected by Cell Counting Kit-8 assay. (C) The
expression level of granulocytic marker CD11b and CSF3R in
untreated and ATRA-treated (duration, 48 h) cells that overexpress
miR-10b. (D) CD11b FACS analysis showed that enforced expression of
miR-10b delayed ATRA-induced granulocytic differentiation of HL-60
cells. (E) The relative expression of monocytic marker CD14 and
CSF1R in untreated and PMA-treated (duration, 48 h) cells that
overexpress miR-10b. (F) CD14 FACS analysis showed that enforced
expression of miR-10b delayed PMA-induced monocytic differentiation
of HL-60 cells. Data are presented as the mean ± standard deviation
(n=3); *P<0.05; **P<0.01; ***P<0.001. All the comparisons
are made between scramble and miR-10a groups. miR, microRNA; CSF3R,
colony-stimulating factor 3 receptor; CD, cluster of
differentiation; CSF1R, colony-stimulating factor 1 receptor; ATRA,
al-trans-retinoic acid; PMA, phorbol myristate acetate; FACS,
fluorescence-activated cell sorting. |
Discussion
Several studies have investigated the association
between abnormal miR-10a/b expression and the risk of developing
various types of cancer, but the results are inconsistent (19,21,23,26).
Accumulating evidence suggests that miR-10a and miR-10b may behave
as novel oncogenes in human cancer (26–28,34,35).
The detection of upregulation of miR-10a expression may serve as
potential biomarkers of aggressive progression and poor prognosis
in cervical cancer (34) miR-10b has
also been reported to promote cell invasion by targeting homeobox
(HOX) D10 in gastric cancer (35).
However, debate exists on whether miR-10a/b acts as a tumor
suppressor or oncogene in human cancer (34–37). Kim
et al (36) reported that
miR-10b may act as a tumor suppressive gene in gastric
carcinogenesis. The loss of miR-10a was also reported to activate
lactoperoxidase to induce intestinal neoplasia through cooperating
with activated Wnt signaling in female mice (37). However, the aberrant expression and
the potential role of miR-10a/b in AML are largely unknown, with
the exception that miR-10a was reported to be expressed at high
levels in NPM1-mutated AML (38) and
that the level of serum miR-10a was also investigated as a
prognostic biomarker for AML (39).
In the present study, the expression of miR-10a and
miR-10b was examined in a number of patients with AML and healthy
controls. Additionally, the potential application of the levels of
miR-10a and miR-10b expression in AML diagnosis was investigated.
The results indicated that miR-10a and miR-10b were significantly
upregulated in AML samples. The upregulation of miR-10a and miR-10b
in AML indicated that miR-10a/b may serve as potential biomarkers
for diagnosis of AML. Several innate properties of miRNAs make them
attractive as potential biomarkers. miRNAs are small and stable
against degradation and can be detected easily by specific and
sensitive RT-qPCR in small amount samples. In addition, miRNAs are
also detectable in bodily fluids, including serum, plasma, saliva,
urine and tears (40,41). Furthermore, expression profiles of
miRNAs in the plasma and/or serum of cancer patients may reflect
the change in miRNA expression in tumor cells (42). Circulating miRNAs may be a novel class
of non-invasive biomarkers for cancer diagnostic and prognostic
information (43,44). The differential expression of
miR-10a/b in different AML subtypes should be validated in more
clinical samples to develop a novel method for subtyping AML
according to miRNA expression.
In addition, to the best of our knowledge, there has
not been any study that investigated the role of miR-10a and
miR-10b in the progression of AML. The present study revealed that
the exogenous expression of miR-10a and miR-10b in HL-60 cells
decreased the maturation of HL-60 cells to granulocytes and
monocytes, as well as the expression of the granulocytic and
monocytic differentiation markers. The overexpression of miR-10a
and miR-10b was able to promote the proliferation of leukemia
cells. It was hypothesized that the abnormal upregulation of
miR-10a/b in blood progenitor cells would result in the
overproduction and accumulation of immature white blood cells, as
well as the inhibition of granulocyte and monocyte maturation,
which results in leukemogenesis. The oncogenic role of miR-10a/b in
the progression of AML will be investigated in animal models.
In humans, miR-10 is co-expressed with a set of Hox
genes and has been demonstrated to regulate the translation of Hox
transcripts (45,46). Hox genes perform crucial roles during
development and are also involved in the tumorigenesis of various
types of cancer (46). HOXA1 has been
identified as a direct target of miR-10a in gastric cancer,
megakaryocytopoiesis and pancreatic cancer (47). The miR-10 family has also been
reported to facilitate cancer by regulating ribosome biogenesis and
consequently global protein production (48). Therefore, it was hypothesized that
miR-10a and miR-10b modulate granulocytic/monocytic differentiation
and AML carcinogenesis through regulating Hox gene expression or
ribosome biogenesis. The possible mechanism by which miR-10a and
miR-10b regulate granulocytic and monocytic differentiation and AML
carcinogenesis requires additional investigation.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81100355
and 81172613).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
ZS and LB conceived and designed the study. LB
performed the majority of experiments. LS and ZJ collected the
clinical samples. SZ helped to perform the cell culture
experiments. LB and ZS wrote the manuscript. All authors have read
and approved this manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committees of The First Affiliated Hospital of Wenzhou Medical
University and written informed consent was obtained from all
patients.
Consent for publication
Written informed consent was obtained from all
examined patients for the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Birbrair A and Frenette PS: Niche
heterogeneity in the bone marrow. Ann N Y Acad Sci. 1370:82–96.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Morrison SJ and Kimble J: Asymmetric and
symmetric stem-cell divisions in development and cancer. Nature.
441:1068–1074. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Falini B, Tiacci E, Martelli MP, Ascani S
and Pileri SA: New classification of acute myeloid leukemia and
precursor-related neoplasms: Changes and unsolved issues. Discov
Med. 10:281–292. 2010.PubMed/NCBI
|
|
4
|
Jemal A, Thomas A, Murray T and Thun M:
Cancer statistics, 2002. CA Cancer J Clin. 52:23–47. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Döhner H, Weisdorf DJ and Bloomfield CD:
Acute myeloid leukemia. N Engl J Med. 373:1136–1152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
GBD 2015 Disease and Injury Incidence and
Prevalence Collaborators, . Global, regional, and national
incidence, prevalence, and years lived with disability for 310
diseases and injuries, 1990–2015: A systematic analysis for the
global burden of disease study 2015. Lancet. 388:1545–1602. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Das A, Sinha M, Datta S, Abas M, Chaffee
S, Sen CK and Roy S: Monocyte and macrophage plasticity in tissue
repair and regeneration. Am J Pathol. 185:2596–2606. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rua R and McGavern DB: Elucidation of
monocyte/macrophage dynamics and function by intravital imaging. J
Leukoc Biol. 98:319–332. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bennett JM, Catovsky D, Daniel MT,
Flandrin G, Galton DA, Gralnick HR and Sultan C: Proposals for the
classification of the acute leukaemias. French-american-british
(FAB) co-operative group. Br J Haematol. 33:451–458. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Garzon R, Volinia S, Liu CG,
Fernandez-Cymering C, Palumbo T, Pichiorri F, Fabbri M, Coombes K,
Alder H, Nakamura T, et al: MicroRNA signatures associated with
cytogenetics and prognosis in acute myeloid leukemia. Blood.
111:3183–3189. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rosenbauer F and Tenen DG: Transcription
factors in myeloid development: Balancing differentiation with
transformation. Nat Rev Immunol. 7:105–117. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Egeland T, Steen R, Quarsten H, Gaudernack
G, Yang YC and Thorsby E: Myeloid differentiation of purified CD34+
cells after stimulation with recombinant human granulocyte-monocyte
colony-stimulating factor (CSF), granulocyte-CSF, monocyte-CSF, and
interleukin-3. Blood. 78:3192–3199. 1991.PubMed/NCBI
|
|
13
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pulikkan JA, Dengler V, Peramangalam PS,
Peer Zada AA, Müller-Tidow C, Bohlander SK, Tenen DG and Behre G:
Cell-cycle regulator E2F1 and microRNA-223 comprise an
autoregulatory negative feedback loop in acute myeloid leukemia.
Blood. 115:1768–1778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang XS, Gong JN, Yu J, Wang F, Zhang XH,
Yin XL, Tan ZQ, Luo ZM, Yang GH, Shen C and Zhang JW: MicroRNA-29A
and microRNA-142-3p are regulators of myeloid differentiation and
acute myeloid leukemia. Blood. 119:4992–5004. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su R, Lin HS, Zhang XH, Yin XL, Ning HM,
Liu B, Zhai PF, Gong JN, Shen C, Song L, et al: miR-181 family:
Regulators of myeloid differentiation and acute myeloid leukemia as
well as potential therapeutic targets. Oncogene. 34:3226–3239.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Havelange V, Ranganathan P, Geyer S,
Nicolet D, Huang X, Yu X, Volinia S, Kornblau SM, Andreeff M, Croce
CM, et al: Implications of the miR-10 family in chemotherapy
response of NPM1-mutated AML. Blood. 123:2412–5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma L: Role of miR-10b in breast cancer
metastasis. Breast Cancer Res. 12:2102010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sasayama T, Nishihara M, Kondoh T, Hosoda
K and Kohmura E: MicroRNA-10b is overexpressed in malignant glioma
and associated with tumor invasive factors, uPAR and RhoC. Int J
Cancer. 125:1407–1413. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Teplyuk NM, Mollenhauer B, Gabriely G,
Giese A, Kim E, Smolsky M, Kim RY, Saria MG, Pastorino S, Kesari S
and Krichevsky AM: MicroRNAs in cerebrospinal fluid identify
glioblastoma and metastatic brain cancers and reflect disease
activity. Neuro-Oncol. 14:689–700. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chai G, Liu N, Ma J, Li H, Oblinger JL,
Prahalad AK, Gong M, Chang LS, Wallace M, Muir D, et al:
MicroRNA-10b regulates tumorigenesis in neurofibromatosis type 1.
Cancer Sci. 101:1997–2004. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian Y, Luo A, Cai Y, Su Q, Ding F, Chen H
and Liu Z: MicroRNA-10b promotes migration and invasion through
KLF4 in human esophageal cancer cell lines. J Biol Chem.
285:7986–7994. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakata K, Ohuchida K, Mizumoto K,
Kayashima T, Ikenaga N, Sakai H, Lin C, Fujita H, Otsuka T, Aishima
S, et al: MicroRNA-10b is overexpressed in pancreatic cancer,
promotes its invasiveness, and correlates with a poor prognosis.
Surgery. 150:916–922. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li G, Wu Z, Peng Y, Liu X, Lu J, Wang L,
Pan Q, He ML and Li XP: MicroRNA-10b induced by epstein–barr
virus-encoded latent membrane protein-1 promotes the metastasis of
human nasopharyngeal carcinoma cells. Cancer Lett. 299:29–36. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ladeiro Y, Couchy G, Balabaud C,
Bioulac-Sage P, Pelletier L, Rebouissou S and Zucman-Rossi J:
MicroRNA profiling in hepatocellular tumors is associated with
clinical features and oncogene/tumor suppressor gene mutations.
Hepatology. 47:1955–1963. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yamamoto H, Adachi Y, Taniguchi H,
Kunimoto H, Nosho K, Suzuki H and Shinomura Y: Interrelationship
between microsatellite instability and microRNA in gastrointestinal
cancer. World J Gastroenterol. 18:2745–2755. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jongen-Lavrencic M, Sun SM, Dijkstra MK,
Valk PJ and Löwenberg B: MicroRNA expression profiling in relation
to the genetic heterogeneity of acute myeloid leukemia. Blood.
111:5078–5085. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Garzon R, Garofalo M, Martelli MP,
Briesewitz R, Wang L, Fernandez-Cymering C, Volinia S, Liu CG,
Schnittger S, Haferlach T, et al: Distinctive microRNA signature of
acute myeloid leukemia bearing cytoplasmic mutated nucleophosmin.
Proc Natl Acad Sci U S A. 105:pp. 3945–3950. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bryant A, Palma CA, Jayaswal V, Yang YW,
Lutherborrow M and Ma DD: miR-10a is aberrantly overexpressed in
nucleophosmin1 mutated acute myeloid leukaemia and its suppression
induces cell death. Mol Cancer. 11:82012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Capitani S, Marchisio M, Neri LM, Brugnoli
F, Gonelli A and Bertagnolo V: Phosphoinositide 3-kinase is
associated to the nucleus of HL-60 cells and is involved in their
ATRA-induced granulocytic differentiation. Eur J Histochem.
44:61–65. 2000.PubMed/NCBI
|
|
34
|
Safari A, Seifoleslami M, Yahaghi E,
Sedaghati F and Khameneie MK: Retracted article: Upregulation of
miR-20a and miR-10a expression levels act as potential biomarkers
of aggressive progression and poor prognosis in cervical cancer.
Tumour Biol. 2015.
|
|
35
|
Liu Z, Zhu J, Cao H, Ren H and Fang X:
miR-10b promotes cell invasion through RhoC-AKT signaling pathway
by targeting HOXD10 in gastric cancer. Int J Oncol. 40:1553–1560.
2012.PubMed/NCBI
|
|
36
|
Kim K, Lee HC, Park JL, Kim M, Kim SY, Noh
SM, Song KS, Kim JC and Kim YS: Epigenetic regulation of
microRNA-10b and targeting of oncogenic MAPRE1 in gastric cancer.
Epigenetics. 6:740–751. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stadthagen G, Tehler D, Høyland-Kroghsbo
NM, Wen J, Krogh A, Jensen KT, Santoni-Rugiu E, Engelholm LH and
Lund AH: Loss of miR-10a activates lpo and collaborates with
activated Wnt signaling in inducing intestinal neoplasia in female
mice. PLoS Genet. 9:e10039132013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Havelange V, Ranganathan P, Geyer S,
Nicolet D, Huang X, Yu X, Volinia S, Kornblau SM, Andreeff M, Croce
CM, et al: Implications of the miR-10 family in chemotherapy
response of NPM1-mutated AML. Blood. 123:2412–2415. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhi Y, Xie X, Wang R, Wang B, Gu W, Ling
Y, Dong W, Zhi F and Liu Y: Serum level of miR-10-5p as a
prognostic biomarker for acute myeloid leukemia. Int J Hematol.
102:296–303. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kosaka N, Iguchi H and Ochiya T:
Circulating microRNA in body fluid: A new potential biomarker for
cancer diagnosis and prognosis. Cancer Sci. 101:2087–2092. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cortez MA, Bueso-Ramos C, Ferdin J,
Lopez-Berestein G, Sood AK and Calin GA: MicroRNAs in body fluids
the mix of hormones and biomarkers. Nat Rev Clin Oncol. 8:467–477.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cherradi N: microRNAs as potential
biomarkers in adrenocortical cancer: Progress and challenges. Front
Endocrinol (Lausanne). 6:1952016.PubMed/NCBI
|
|
44
|
Saplacan RM, Mircea PA, Balacescu L and
Balacescu O: MicroRNAs as non-invasive screening biomarkers of
colorectal cancer. Clujul Med. 88:453–456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu H, Lindsay J, Feng ZP, Frankenberg S,
Hu Y, Carone D, Shaw G, Pask AJ, O'Neill R, Papenfuss AT and
Renfree MB: Evolution of coding and non-coding genes in HOX
clusters of a marsupial. BMC Genomics. 13:2512012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pearson JC, Lemons D and McGinnis W:
Modulating Hox gene functions during animal body patterning. Nat
Rev Genet. 6:893–904. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jia H, Zhang Z, Zou D, Wang B, Yan Y, Luo
M, Dong L, Yin H, Gong B, Li Z, et al: MicroRNA-10a is
down-regulated by DNA methylation and functions as a tumor
suppressor in gastric cancer cells. PLoS One. 9:e880572014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ørom UA, Nielsen FC and Lund AH:
MicroRNA-10a binds the 5′UTR of ribosomal protein mRNAs and
enhances their translation. Mol Cell. 30:460–471. 2008. View Article : Google Scholar : PubMed/NCBI
|