Introduction
Colorectal cancer (CRC) is one of the most fatal
types of cancer worldwide (1). A
range of chemotherapeutic drugs and other methods are available for
the treatment of CRC, but the associated side effects hamper their
clinical application and efficacy (2). With the development of
molecular-targeted therapies, progress has been made in cancer
therapy. In recent years, a number of molecules have been
identified as targets for molecular-targeted anticancer drugs,
including growth factors, factors regulating cell survival and
molecules associated with the cell cycle (3). There is a critical need to identify
potential molecular targets and improve the therapy of CRC.
Ubiquitin-specific peptidase 39 (USP39) is a
deubiquitinating enzyme containing a central zinc finger and two
ubiquitin C-terminal hydrolase domains (4). USP39 is critical in the process of
pre-mRNA splicing, but does not exhibit ubiquitin-specific
peptidase activity (5). As a 65 kDa
SR protein of the U4/U6.U5 tri-small nuclear ribonucleoprotein
complex, USP39 also functions in the assembly of the mature
spliceosome complex and is indispensable in maintaining the
integrity of the mitotic spindle checkpoint (6,7). A
previous study indicated that USP39 silencing induced defective
chromosome segregation and cytokinesis in U2OS cells, indicating
USP39 is critical in the regulation of mitosis (7). Furthermore, the pro-growth effect of
USP39 in cancer cells has been widely investigated. Wen et
al (8) demonstrated that the
overexpression of USP39 promoted the proliferation of prostate
cancer cells, which is in accord with a study by Wang et al
(9) in breast cancer cells.
Furthermore, USP39 knockdown has been demonstrated to affect cell
cycle distribution by inducing arrest in the G2/M phase,
and to promote apoptosis, in human hepatocellular carcinoma
(10) and medullary thyroid carcinoma
(11) cells, indicating that USP39
may be a feasible target for the molecular therapy of various types
of cancer. However, limited data regarding the biological function
of USP39 in CRC cells is available.
Therefore, the effect of USP39 in CRC cells was
assessed in the present study. Lentivirus-delivered short hairpin
RNA (shRNA) was used to infect CRC cells to facilitate
loss-of-function analysis. The function of USP39 knockdown on the
proliferation, colony formation ability, cell cycle progression and
apoptosis of CRC cells was investigated. To the best of our
knowledge, this is the first study to demonstrate that USP39 is
associated with CRC cell proliferation.
Materials and methods
Cell lines and cell culture
SW1116 and HCT116 human CRC cells, and 293T cells
were purchased from the Cell Bank of Chinese Academy of Sciences
(Shanghai, China). SW1116 and 293T cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.). HCT116 cells were
cultured in RPMI-1640 (Hyclone; GE Healthcare Life Sciences, Logan,
UT, USA) supplemented with 10% FBS. All cells were maintained at
37°C in a humidified incubator with 5% CO2.
Construction of lentiviral particles
and USP39 knockdown
To silence the expression of USP39, two candidate
shRNAs for human USP39, based on the Gen Bank sequence
NM_001256725.1, were designed (KD-1 and −2). The shRNA sequences
targeting USP39 were as follows: KD-1,
5′-GATTTGGAAGAGGCGAGATAACTCGAGTTATCTCGCCTCTTCCAAATC-3′; KD-2,
5′-CCTTCCAGACAACTATGAGATCTCGAGATCTCATAGTTGTCTGGAAGG-3′. A
non-silencing shRNA with the sequence,
5′-TTCTCCGAACGTGTCACGTCTCGAGACGTGACACGTTCGGAGAA-3′, was used as a
negative control (shCon). The oligos were annealed and inserted
into pFH-L plasmids (Shanghai Holly Lab, Shanghai, China), which
included a green fluorescent protein (GFP) tag. The lentiviral
particles were constructed in 293T cells as described in a previous
study (12). SW1116 and HCT116 cells
were incubated in 6-well plates and infected with KD or shCon for
96 h with a multiplicity of infection of 30. Successful infection
was confirmed by the observation of GFP expression with
fluorescence microscopy (DMI4000B; Leica Microsystems GmbH,
Wetzlar, Germany). The efficiency of knockdown was determined by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blot analyses. All experiments were repeated
in triplicate.
RT-qPCR analysis
Cells were harvested at 5 days after lentiviral
infection. TRIzol reagent (Thermo Fisher Scientific, Inc.) was used
to extract total RNA, according to the manufacturer's instructions.
Single-stranded cDNA was synthesized using Superscript II Reverse
Transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.). The
primers were as follows: USP39 forward, 5′-GCCAGCAGAAGAAAAAGAGC-3′
and reverse, 5′-GCCATTGAACTTAGCCAGGA-3′; β-actin (endogenous
control) forward, 5′-GTGGACATCCGCAAAGAC-3′ and reverse,
5′-AAAGGGTGTAACGCAACTA-3′. The mRNA levels of USP39 were determined
using SYBR Green on the Bio-Rad Connect Real-Time PCR system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The total PCR
reaction volume was 20 µl, including 10 µl 2X SYBR Premix Ex Taq
(Takara Bio, Inc., Otsu, Japan), 0.5 µl primers (2.5 µM), 5 µl cDNA
and 4.5 µl ddH2O. The PCR thermocycling procedure was as
follows: Initial denaturation at 95°C for 1 min, followed by 40
cycles of denaturation at 95°C for 5 sec and extension at 60°C for
20 sec. The relative gene expression levels were calculated and
compared using the 2−ΔΔCq method (13).
Western blot analysis
Cells were harvested and lysed in 2X protein lysis
buffer [10 mM EDTA, 100 mM Tris-HCl (pH 6.8), 4% SDS and 10%
glycine] 5 days after lentiviral infection. Protein lysates were
collected by centrifugation at 12,000 × g for 15 min at 4°C. Equal
amount of protein samples (30 µg) were separated on 10% SDS-PAGE
and transferred to a PVDF membrane at 300 mA for 1.5 h. Then the
membrane was blocked with Tris-buffered saline with 0.1% Tween-20
containing 5% non-fat dried milk for 1 h at room temperature, and
probed with the corresponding primary antibodies. Primary
antibodies included mouse anti-p53 (dilution 1:1,000; sc-126; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA), rabbit anti-p-p53
(dilution 1:500; cat. no. 2528), rabbit anti-caspase-3 (dilution
1:500; cat. no. 9661), rabbit anti-poly(ADP-ribose) polymerase
(PARP; dilution 1:1,000; cat. no. 9542; all from Cell Signaling
Technology, Inc., Danvers, MA, USA) and rabbit anti-GAPDH (dilution
1:100,000; cat. no. 10494-1-AP; Protein Tech Group, Inc., Chicago,
IL, USA). The membranes were immunoblotted with the appropriate
primary antibodies at 4°C overnight, and then incubated with a
HRP-conjugated goat anti-rabbit (dilution 1:5,000; cat. no.
SC-2054) or goat anti-mouse antibody (dilution 1:5,000; cat. no.
SC-2005; both from Santa Cruz Biotechnology, Inc.) as appropriate
for 2 h at room temperature. Signals were detected using Super ECL
Detection Reagent (Applygen Technologies, Inc., Beijing,
China).
MTT assay for cell viability
SW1116 and HCT116 cells were seeded at 2,000
cells/well in 96-well plates at 4 days after infection. MTT
solution (20 µl) was added into each well, and incubated for a
further 4 h at 37°C. The supernatant was removed and 100 µl acidic
isopropanol (10% SDS, 5% isopropanol and 0.01 mol/l HCl) was added
to each well. The absorbance values at 595 nm were determined using
a microplate reader.
Colony formation assay
Stably transfected SW1116 (500 cells/well) and
HCT116 (400 cells/well) cells were seeded into 6-well plates.
Following incubation for 8 days (SW1116) or 7 days (HCT116), cells
were washed in ice-cold PBS and fixed with methanol at the
temperature of 37°C for 15 min. Then crystal violet (Beyotime
Institute of Biotechnology, Haimen, China) staining was performed,
according to the manufacturer's instructions. The total number of
colonies, defined as groups of >50 cells, was counted under
light microscopy. The colony images were analyzed using Metamorph
software version 7.5 (Molecular Devices LLC, Sunnyvale, CA,
USA).
Cell cycle analysis
To investigate the mechanisms underlying the effects
of USP39 on cell growth, cell cycle distribution was examined by
flow cytometry and propidium iodide (PI) staining. SW1116
(7×104 cells/well) and HCT116 (2×105
cells/well) cells were harvested at 7 days and 5 days after
infection, respectively. Cells were fixed in 70% ice-cold ethanol
for 4 h and washed twice with cold PBS. The fixed cells were
stained with PI and detected by a FACSCalibur flow cytometer (BD
Biosciences, San Jose, CA, USA) according to the manufacturer's
protocol. The results were analyzed using FlowJo software version
7.6.2 (FlowJo LLC, Ashland, OR, USA).
Apoptosis analysis
To further confirm whether the effect of
USP39-knockdown on cell proliferation was associated with
apoptosis, flow cytometry with Annexin V-APC/7-AAD double staining
was performed on SW1116 cells. Briefly, at 7 days after infection,
SW1116 cells (8×104 cells/dish) were seeded in 6-cm
dishes and cultured for a further 48 h. Then the cells were
collected, double stained using Annexin V-APC and 7-AAD according
to the manufacturer's protocol (Nanjing Key Gen Biotech Co., Ltd.,
Nanjing, China), and detected by a FACSCalibur flow cytometer.
Statistical analysis
All experiments were repeated in triplicate and the
results were presented as the mean ± standard deviation of three
independent experiments using GraphPad Prism 5 (GraphPad Software,
Inc., La Jolla, CA, USA). Analysis was performed with a one-way
analysis of variance followed by Dunnett's multiple comparisons
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Lentivirus-mediated RNAi suppresses
the expression of USP39
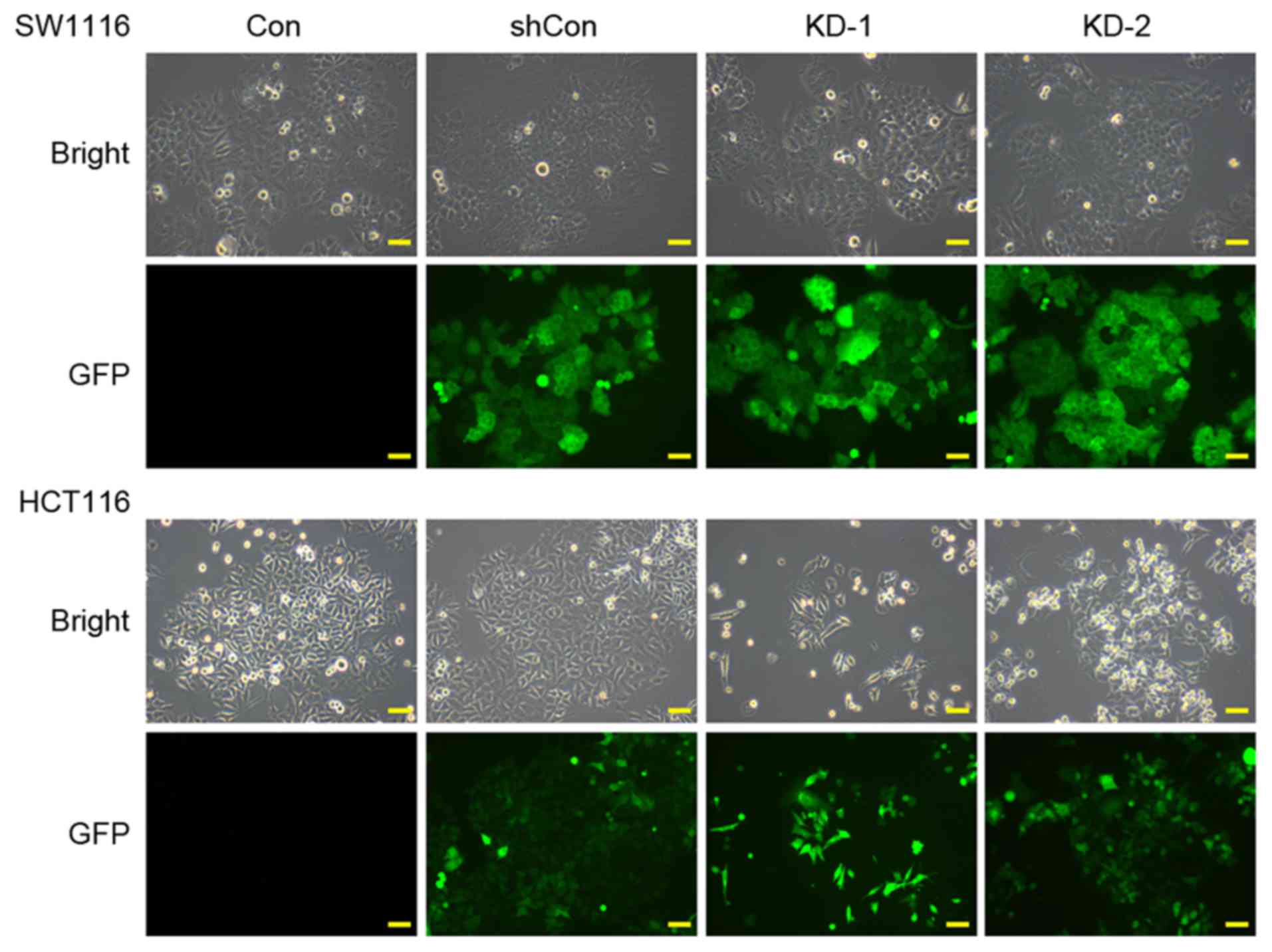
To investigate the effects of USP39 in CRC, the
expression of USP39 was knocked down in SW1116 and HCT116 cells
using lentivirus-mediated transfection. As shown in Fig. 1, >80% of the cells expressed GFP
following infection with shRNA targeting USP39 (KD-1 and −2) or
control shRNA (shCon), suggesting that the recombinant lentivirus
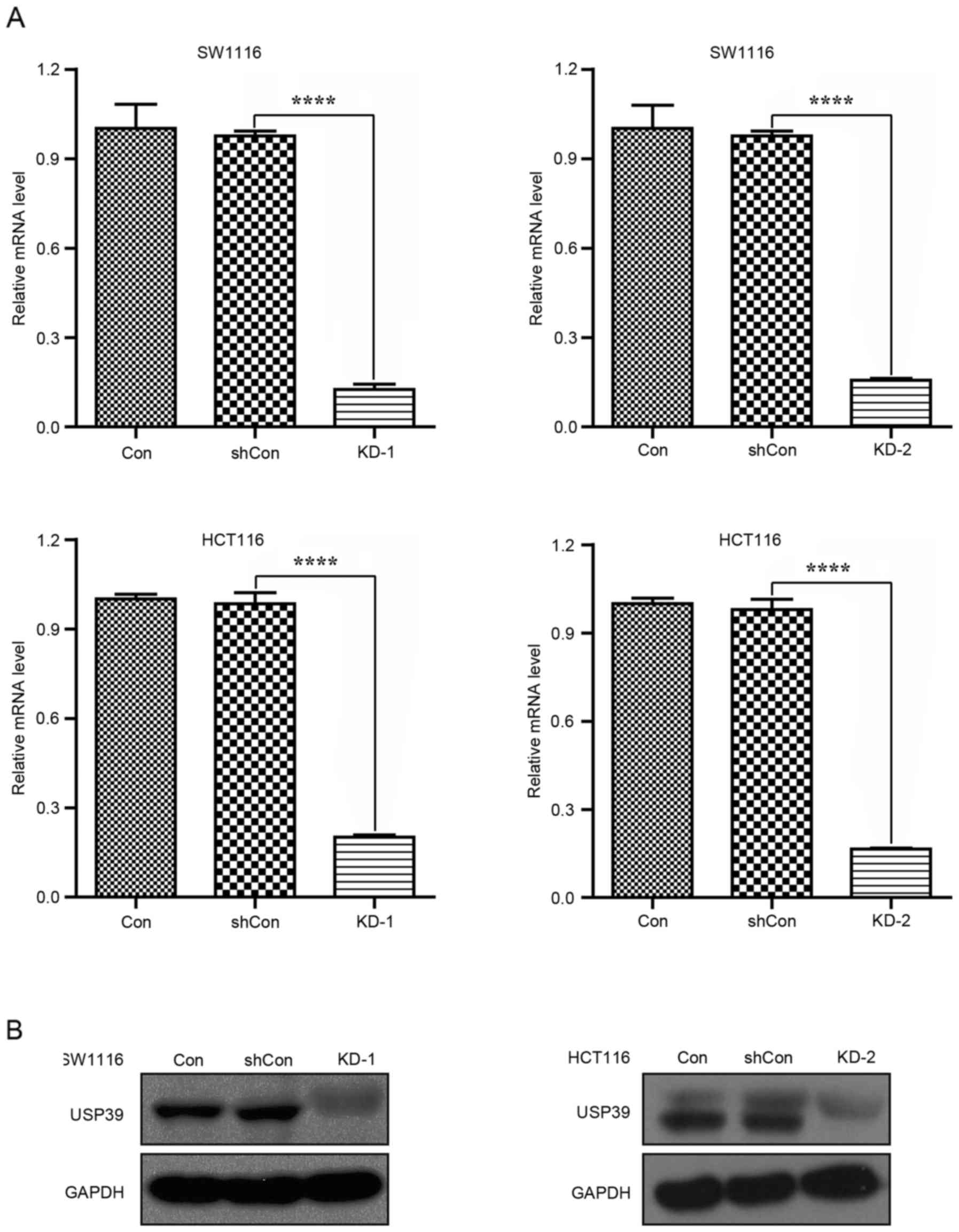
generated high infection efficiency in CRC cells. RT-qPCR analysis
indicated that the USP39 mRNA levels were significantly
downregulated in the KD-1 and 2 groups compared with the shCon and
Con groups of SW1116 and HCT116 cells (Fig. 2A; P<0.0001). Consistent with this,
USP39 protein levels were also downregulated in the KD-1 and −2
groups of SW1116 and HCT116 cells (Fig.
2B). Therefore, it was demonstrated that both shRNAs against
USP39 exerted knockdown effects on USP39 expression.
USP39 knockdown inhibits CRC cell
viability and colony formation
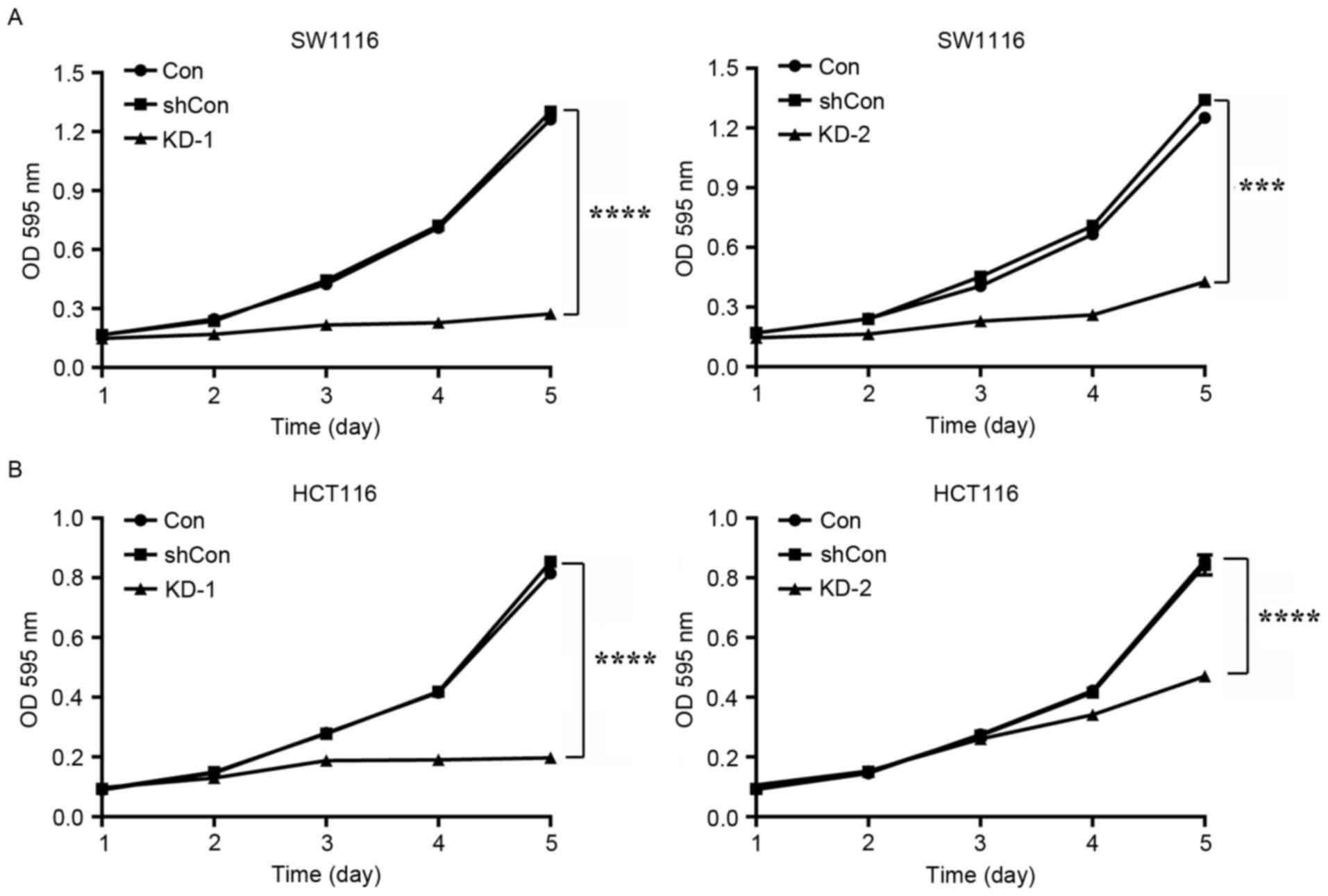
To determine the role of USP39 expression on CRC
cell viability, an MTT assay was performed on SW1116 and HCT116
cells. As included in Fig. 3A, the
viability of cells of SW1116 cells in the KD-1 (P<0.0001) and
KD-2 (P<0.001) groups was significantly decreased compared with
the shCon and Con groups. A similar result was achieved in HCT116
cells (Fig. 3B; both P<0.0001).
KD-1 suppressed cell viability to a greater extent than KD-2 in the
SW1116 and in HCT116 cells, and was therefore selected for use in
further procedures.
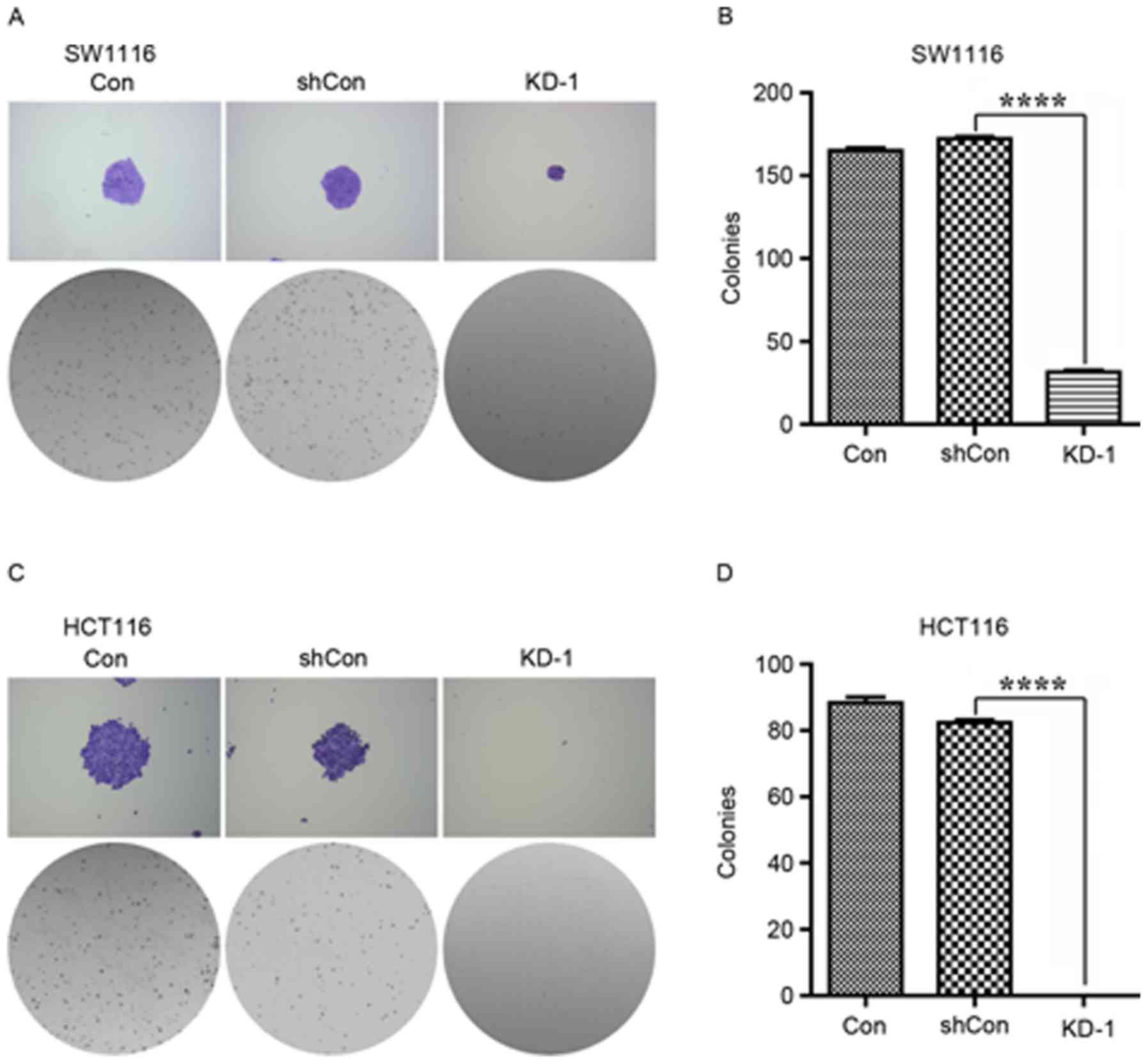
The colony formation capacity of SW1116 and HCT116
cells was then determined for Con, shCon and KD-1 cells.
Representative images of the size of each colony and the number of
colonies per well are provided in Fig.
4. Analysis indicated that the number of colonies was reduced
in the KD-1 group, compared with the Con and shCon groups, for
SW1116 and HCT116 (Fig. 4;
P<0.0001).
USP39 knockdown inhibits cell cycle
progression
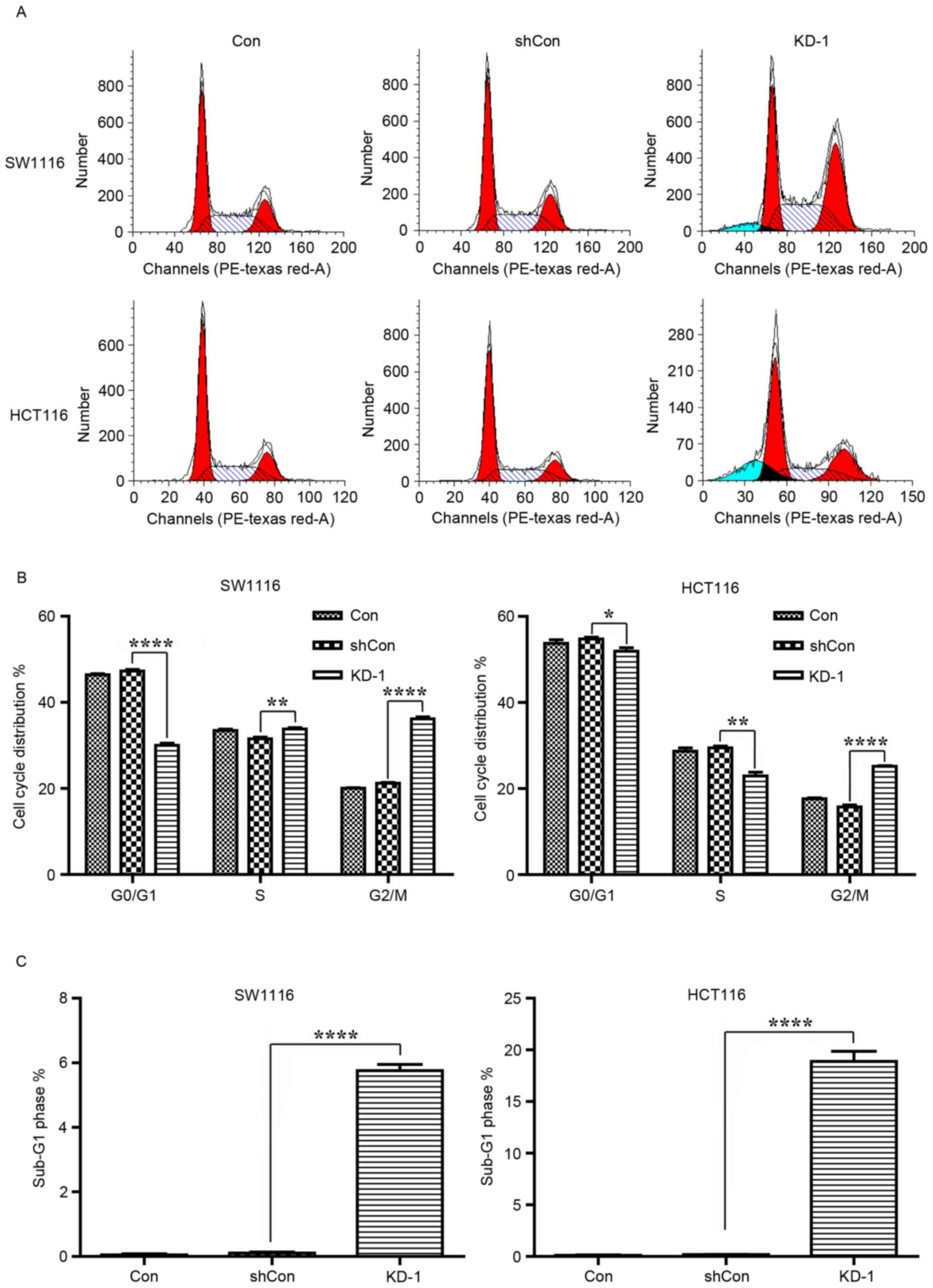
Cell proliferation is controlled by cell cycle
progression. Therefore, cell cycle distribution was analyzed with
FACS in SW1116 and HCT116 cells subsequent to USP39 knockdown
(Fig. 5A). As presented in Fig. 5B, the proportion of SW1116 and HCT116
cells in the G0/G1 (SW1116, P<0.0001;
HCT116, P<0.05) and S (both P<0.01) phases was significantly
reduced, and the proportion in the G2/M phase
significantly increased (both P<0.0001) in the KD-1 group
compared with the Con and shCon groups. These results indicated
that USP39 may have regulated cell proliferation through an effect
on cell cycle distribution. In addition, more cells in the
sub-G1 phase and therefore, undergoing apoptosis, were
detected in SW1116 and HCT116 cells subsequent to USP39 knockdown
(Fig. 5C; both P<0.0001).
USP39 knockdown promotes
apoptosis
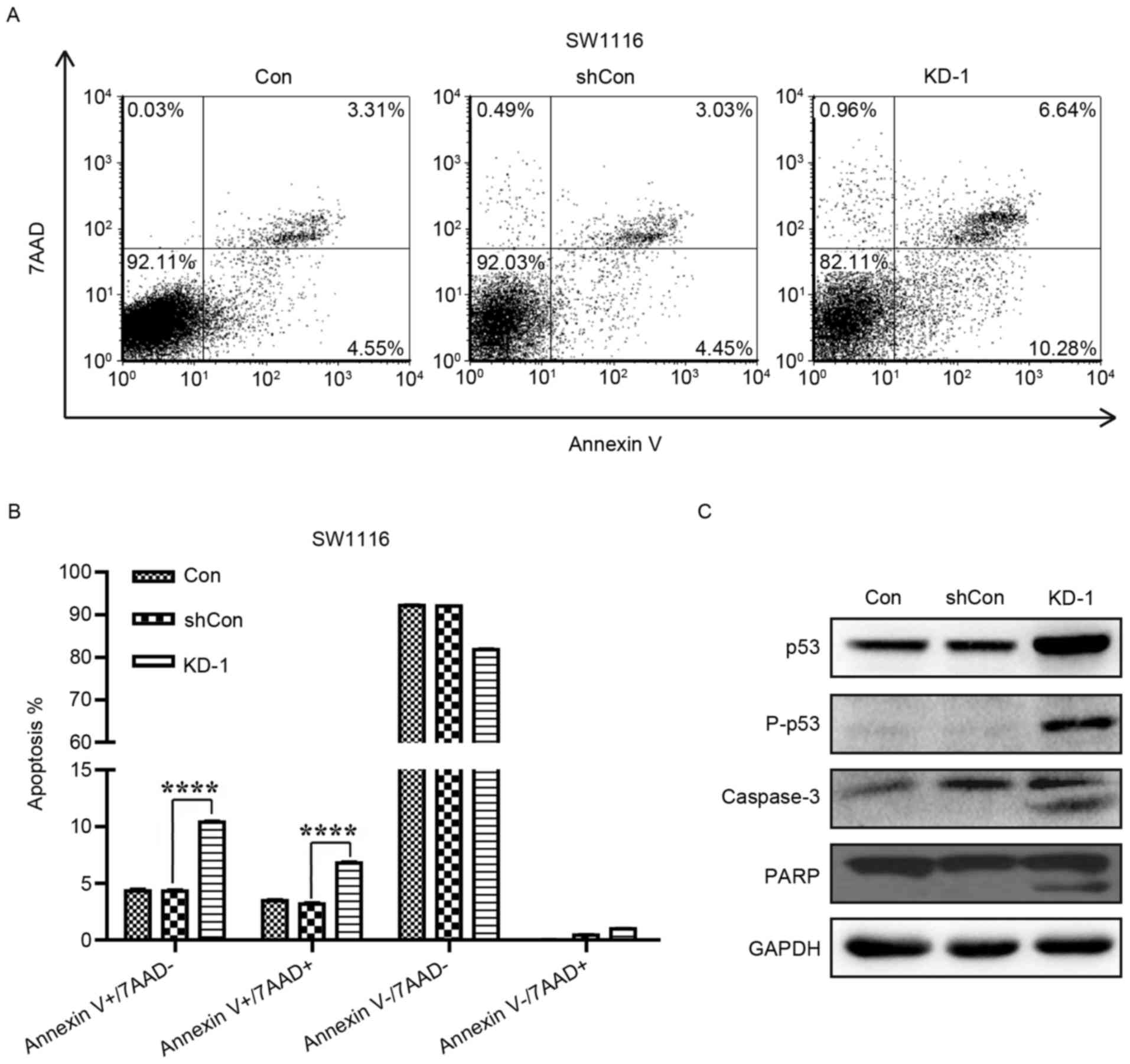
To further investigate the apoptosis-promoting
effects of USP39 silencing in CRC cells, SW1116 cells were analyzed
with Annexin V-APC/7-AAD double staining and flow cytometry
(Fig. 6A). As demonstrated in
Fig. 6B, the knockdown of USP39
increased the proportion of cells at early (Annexin
V+/7-AAD-) and late (Annexin
V+/7-AAD+) apoptotic stages by ~2-fold
compared with the Con and shCon groups (P<0.0001). This data
demonstrated that USP39 silencing induced apoptosis in SW1116
cells. In addition, the expression levels of apoptotic markers,
including PARP, p53 and caspase-3, were also assessed in SW1116
cells. As depicted in Fig. 6C, the
expression levels of p53, p-p53, PARP and caspase-3 were increased
in SW1116 cells following KD-1 infection. Thus, it was demonstrated
that the knockdown of USP39 in SW1116 cells induced apoptosis via
altering the expression of apoptosis-associated proteins.
Discussion
CRC is one of the leading causes of cancer
mortality, and results from uncontrolled cell growth in the colon,
rectum or appendix (14).
Molecular-targeted therapies may present a powerful treatment
option for CRC. Thus, the present study focused on the
identification of an oncogenic target in CRC and investigated the
biological effects of silencing the identified gene.
In the present study, it was demonstrated that the
knockdown of USP39 by USP39-specific shRNA could significantly
suppress the growth and colony formation abilities of the CRC cell
lines SW1116 and HCT116. It was previously identified that
spliceosome factors are associated with cancer development
(15,16). As a confirmed spliceosome factor,
USP39 is critical to maintain the spindle checkpoint and promote
successful cytokinesis through the regulation of Aurora B mRNA
splicing in mammalian cells (7). The
downregulation of Aurora B leads to defects in spindle checkpoint
function and cytokinesis (7). This
indicates that USP39 may affect proliferation via affecting Aurora
B mRNA splicing. In addition, flow cytometry analysis in the
present study identified that the knockdown of USP39 induced cell
cycle arrest in the G2/M phases, which may have induced
the inhibition of proliferation. These results correspond with a
previous study, which indicated that the knockdown of USP39
markedly suppressed the proliferation of TT medullary thyroid
carcinoma cells (11).
It was identified in the present study that cells
accumulated in the sub-G1 phase, indicative of the
induction of apoptosis. Further analysis confirmed that USP39
silencing significantly promoted the apoptosis of SW1116 cells.
Apoptosis is a caspase-dependent form of programmed cell death
(17), which is necessary for the
maintenance of bodily health (18).
Caspase-3 is the key enzyme in the process of apoptosis (19); once activated, it cleaves specific
substrates, including PARP, to mediate apoptosis (20,21). The
transcription factor p53, a tumor suppressor, is phosphorylated and
activated as a reaction to DNA damage, leading to growth arrest and
the induction of cell death (22).
Consistent with the results of the present study, the level of p53
expression and PARP cleavage have been demonstrated to increase
during the cell cycle arrest and apoptosis of CRC cells (23). Taken together, it can be concluded
that the growth inhibition associated with USP39 silencing in CRC
cells may have been induced by the induction of spliceosome
factor-associated apoptosis.
In conclusion, the present study has revealed that
USP39 silencing suppressed CRC cell proliferation via activating
the caspase cascade and upregulating the expression of p53. These
data may provide an experimental basis for the development of USP39
as a potential molecular target against CRC. Further investigation
is required to verify the efficacy of USP39-targeted therapy in
vivo.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZX performed the experiments and drafted the
manuscript. FS and WH participated in the experiments. ZW and XS
participated in the data analysis and figure format. FZ
participated in the research design, the article reviewing and the
data examining.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gansler T, Ganz PA, Grant M, Greene FL,
Johnstone P, Mahoney M, Newman LA, Oh WK, Thomas CR Jr, Thun MJ, et
al: Sixty years of CA: A cancer journal for clinicians. CA Cancer J
Clin. 60:345–350. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li Q, Zhou S, Jing J, Yang T, Duan S, Wang
Z, Mei Q and Liu L: Oligosaccharide from apple induces apoptosis
and cell cycle arrest in HT29 human colon cancer cells. Int J Biol
Macromol. 57:245–254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Allgayer H and Fulda S: An introduction to
molecular targeted therapy of cancer. Adv Med Sci. 53:130–138.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lygerou Z, Christophides G and Seraphin B:
A novel genetic screen for snRNP assembly factors in yeast
identifies a conserved protein, Sad1p, also required for pre-mRNA
splicing. Mol Cell Biol. 19:2008–2020. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hadjivassiliou H, Rosenberg OS and Guthrie
C: The crystal structure of S. cerevisiae Sad1, a catalytically
inactive deubiquitinase that is broadly required for pre-mRNA
splicing. RNA. 20:656–669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Makarova OV, Makarov EM and Luhrmann R:
The 65 and 110 kDa SR-related proteins of the U4/U6.U5 tri-snRNP
are essential for the assembly of mature spliceosomes. EMBO J.
20:2553–2563. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Leuken RJ, Luna-Vargas MP, Sixma TK,
Wolthuis RM and Medema RH: Usp39 is essential for mitotic spindle
checkpoint integrity and controls mRNA-levels of aurora B. Cell
Cycle. 7:2710–2719. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wen D, Xu Z, Xia L, Liu X, Tu Y, Lei H,
Wang W, Wang T, Song L, Ma C, et al: Important role of SUMOylation
of Spliceosome factors in prostate cancer cells. J Proteome Res.
13:3571–3582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang H, Ji X, Liu X, Yao R, Chi J, Liu S,
Wang Y, Cao W and Zhou Q: Lentivirus-mediated inhibition of USP39
suppresses the growth of breast cancer cells in vitro. Oncol Rep.
30:2871–2877. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pan Z, Pan H, Zhang J, Yang Y, Liu H, Yang
Y, Huang G, Ni J, Huang J and Zhou W: Lentivirus mediated silencing
of ubiquitin specific peptidase 39 inhibits cell proliferation of
human hepatocellular carcinoma cells in vitro. Biol Res. 48:182015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
An Y, Yang S, Guo K, Ma B and Wang Y:
Reduced USP39 expression inhibits malignant proliferation of
medullary thyroid carcinoma in vitro. World J Surg Oncol.
13:2552015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun W, Yao L, Jiang B, Guo L and Wang Q:
Spindle and kinetochore-associated protein 1 is overexpressed in
gastric cancer and modulates cell growth. Mol Cell Biochem.
391:167–174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
de-Freitas Junior JC and Morgado-Diaz JA:
The role of N-glycans in colorectal cancer progression: Potential
biomarkers and therapeutic applications. Oncotarget. 7:19395–19413.
2016.PubMed/NCBI
|
|
15
|
Wang L, Lawrence MS, Wan Y, Stojanov P,
Sougnez C, Stevenson K, Werner L, Sivachenko A, DeLuca DS, Zhang L,
et al: SF3B1 and other novel cancer genes in chronic lymphocytic
leukemia. N Engl J Med. 365:2497–2506. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cazzola M, Rossi M and Malcovati L;
Associazione Italiana per la Ricerca sul Cancro Gruppo Italiano
Malattie Mieloproliferative, : Biologic and clinical significance
of somatic mutations of SF3B1 in myeloid and lymphoid neoplasms.
Blood. 121:260–269. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salvesen GS and Dixit VM: Caspases:
Intracellular signaling by proteolysis. Cell. 91:443–446. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shimizu S, Yoshida T, Tsujioka M and
Arakawa S: Autophagic cell death and cancer. Int J Mol Sci.
15:3145–3153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 37:719–727. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ghavami S, Hashemi M, Ande SR, Yeganeh B,
Xiao W, Eshraghi M, Bus CJ, Kadkhoda K, Wiechec E, Halayko AJ and
Los M: Apoptosis and cancer: Mutations within caspase genes. J Med
Genet. 46:497–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matuo R, Sousa FG, Escargueil AE,
Grivicich I, Garcia-Santos D, Chies JA, Saffi J, Larsen AK and
Henriques JA: 5-Fluorouracil and its active metabolite FdUMP cause
DNA damage in human SW620 colon adenocarcinoma cell line. J Appl
Toxicol. 29:308–316. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vousden KH and Lane DP: p53 in health and
disease. Nat Rev Mol Cell Biol. 8:275–283. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thorenoor N, Faltejskova-Vychytilova P,
Hombach S, Mlcochova J, Kretz M, Svoboda M and Slaby O: Long
non-coding RNA ZFAS1 interacts with CDK1 and is involved in
p53-dependent cell cycle control and apoptosis in colorectal
cancer. Oncotarget. 7:622–637. 2016. View Article : Google Scholar : PubMed/NCBI
|