Introduction
Human colorectal cancer has been ranked the third
most commonly diagnosed malignancy through the world and the
leading cause of cancer-related mortality in Western countries,
exceeded only by lung, live and stomach cancers (1). Over the past decades, despite the
mortality of colorectal cancer has been obviously improved, due to
the rapid development of new drugs (i.e., irinotecan and
oxaliplatin) and target therapies (i.e., bevacizumab, cetuximab,
panitumab, aflibercept and regorafenib), approximately one million
new cases of colorectal cancer diagnosed worldwide and half a
million people died from colorectal cancer annually, which implies
significant impact on public health (2). Moreover, it is now clear that excessive
alcohol use, obesity, older age, chronic intestinal inflammation,
family history, racial and ethnic background, environmental and
genetic factors have been identified as the most important risk
factors (3,4). Among these, genetic factors, mainly
referred to the accumulation of both gene mutations and epigenetic
modifications of the genome in colonic mucosa cells, are considered
as the key components which lead to cell proliferation and
metastasis and ultimately potentiate carcinogenesis of colorectal
cancer (5). For example, the changes
of EGF receptor (EGFR) signaling, phosphatase and tensin homolog
(PTEN)/phosphatidylinositol-3-kinase (P13K) signaling and p53
signaling were implicated in pathogenesis of colorectal cancer
(6). Therefore, in-depth
investigations on the underlying molecular mechanisms of colorectal
cancer occurrence and development and the identification of serum
and tissue markers with prognostic and predictive value involved in
colorectal cancer are the urgent need for the management and
treatment of this disease (7).
Currently, accumulating studies have highlighted that microRNA
(miRNA) is a promising prognostic biomarkers and a potential
therapeutic target in colorectal cancer (8,9). For
instance, miR-181c was determined as a predictive molecule for
recurrence of colorectal cancer patients at stage II (10).
miRNA, a discovered group of endogenous, non-coding
RNA molecules approximately 22 nt in length, directly interacts
with a member of the Argonaute (Ago) protein family and forms
effector complexes that modulate gene expression
post-transcriptionally by binding to complementary sequences in the
3′-untranslated region (UTR) of target messenger RNAs (mRNAs)
(11). Given the tremendous impact of
miRNA-guided gene regulation on almost all aspects of cellular
processes in eukaryotic organisms, such as proliferation, cell fate
determination, apoptosis, signal transduction, organ development,
hematopoietic lineage differentiation and tumorigenesis, it is not
surprising that miRNA deregulation is intimately related to the
molecular mechanisms of various clinical diseases, including cancer
(12). In the past decades, it was
found that miRNA abnormalities play a pivotal role in diverse
cancer subtypes, such as lung cancer, breast cancer, and
glioblastoma, and different cancers have different miRNA profiles,
which could reflect the developmental lineage and differentiation
state of the tumors, thereby studying the specific function of
these aberrant miRNAs in human carcinogenesis might be provide a
powerful tool as novel clinical biomarkers for early cancer
diagnosis, prognosis and targets for therapy (13,14).
Numerous literatures reported that many miRNAs involved in
colorectal cancer initiation and emergence were detected, for
example, reduced expression of miR-143 is responsible for
colorectal cancer development through derepressing Kirsten rat
sarcoma viral oncogene homolog (KRAS) expression and Insulin-like
growth factor 1 receptor (IGF1R) (15,16);
elevated miR-92a levels promoted cell proliferation, invasion,
epithelial-mesenchymal transition (EMT) via targeting phosphatase
and tensin homolog (PTEN) (17).
In previous study, it was revealed that miR-155
might be implicated in formation of colorectal cancer (18), and collagen triple helix repeat
containing 1 (CTHRC1) was also predicted as a potential target of
miR-155 by bioinformatics analysis. Since its discovery, miR-155
was uncovered that it participated in promoting cancers of lung,
liver, pancreas and gastrointestinal tract by repressing different
targets (19). Furthermore, it is
verified that CTHRC1 could suppress tumor growth and metastasis in
colorectal cancer (20). Thus, in
this study, we aimed to confirm the interaction between miR-155 and
CTHRC1 and explore their roles in colorectal cancer, which might
not only help elucidate the molecular mechanism of miR-155 and
CTHRC1 in colorectal cancer, but also offer potential utilization
as innovative therapeutics in colorectal cancer.
Materials and methods
Ethics statement and clinical
samples
The procedures of this study were approved by the
Clinical Ethics Management Committee of Jinshan Hospital Affiliated
to Fudan University. Written informed consent was obtained from 4
participants who have hospitalized in Jinshan Hospital Affiliated
to Fudan University from March, 2016 to May, 2016 and have been
made a definite diagnosis by at least three pathologists (21). The detailed patients' clinical
characteristics were displayed in Table
I. Fresh colorectal cancer tissues and paired adjacent normal
specimens were collected by surgical resection. The samples were
washed with an appropriate amount of cold saline to reduce blood
contamination and then immediately removed and preserved in liquid
nitrogen until they were processed for miR-155 and CTHRC1
expressions detection by quantitative reverse
transcription-polymerase chain reaction (qRT-PCR) assay.
 | Table 1.The basic clinical characteristics of
study participants. |
Table 1.
The basic clinical characteristics of
study participants.
| ID | Gender | Age (Year) | Tumor location | Pathological
type | Differentiated
degree | TNM
classification |
|---|
| 1 | Male | 42 | Ascending colon | Adenocarcinoma | Moderately
differentiated | T3aN2M1 |
| 2 | Male | 46 | Sigmoid colon | Adenocarcinoma | Moderately
differentiated | T3aN0M1 |
| 3 | Female | 39 | Transverse colon | Adenocarcinoma | Moderately
differentiated | T3aN0M1 |
| 4 | Female | 47 | Descending colon | Adenocarcinoma | Moderately
differentiated | T3aN2M1 |
Cell line culture and
transfection
Human colon cancer cells-derived cell lines HCT-8,
Lovo, Colo205, HCT-116 and HT-29 were purchased from American Type
Culture Collection (ATCC) and cultured in RPMI-1640 cell culture
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 2 mM glutamine, 1 mM sodium pyruvate, 100 UI/ml
penicillin and 0.1 mg/ml streptomycin at 37°C in a humidified 5%
CO2/95% air atmosphere until cell lines reached 90%
confluence. Then, the cells washed twice with phosphate-buffered
saline (PBS) and continued to passage with 0.25% trypsin.
Additionally, 293T cells similarly obtained from ATCC were
cultivated in RPMI-1640 cell culture medium, containing 10% FBS, 1%
non-essential amino acids and 1% penicillin/streptomycin under
humidified conditions at 37°C with 5% CO2 for dual
luciferase assay.
For transfection with plasmids, including miR-155
mimics plasmid, miR-155 inhibitor plasmid, negative control (NC)
miRNA plasmid and NC miRNA inhibitor plasmid, cells were seeded in
6-well plates with antibiotic free growth medium at a density of
1×106 cells/well. Next day, when grown to 80%
confluence, cells were transfected with the above plasmid using
Lipofectamine 2000 (Promega Corporation, Madison, WI, USA)
following the manufacturer's guidelines. After 6 h of transfection,
the cell culture medium was replaced with fresh medium and the
cells were harvested at 48 h for the following experiments. The
transfection efficiency was evaluated by qRT-PCR and WB. All of
these plasmids used in this study were designed and purchased from
Guangzhou RiboBio Co., Ltd., Guangzhou, China.
RT-qpcr
Isolation of miRNAs from patient specimens and
colorectal cancer cell lines was carried out using the mirVana™
miRNA Isolation kit (Thermo Fisher Scientific, Inc.) according to
manufacturer's instructions. The quantity and quality of RNA were
evaluated by a BioPhotometer and RNA integrity was determined by
gel analysis. RNA was converted to complementary DNA (cDNA) with a
miR-155-specific stem-loop primer using a PrimeScript RT Reagent
Kit with cDNA Eraser (Takara Biotechnology Co., Ltd., Dalian,
China) in accordance with the manufacturer's manual. Reverse
transcription reaction was conducted at 16°C for 30 min and at 42°C
for 40 min, followed by heat inactivation at 85°C for 5 min.
Subsequently, the cDNA was in turn PCR amplified in a 96-well
optical plate on an ABI PRISM® 7500 Sequence Detection
System (Thermo Fisher Scientific, Inc.) with SYBR Premix Ex Taq TM
II (Takara Biotechnology Co., Ltd.) at 95°C for 30 sec, followed by
40 cycles at 95°C for 15 sec and 60°C for 32 sec and dissociation
at 95°C for 60 sec, 55°C for 30 sec and 95°C for 30 sec. The
expression level of miR-155 and CTHRC1 was normalized to that of
the housekeeping gene U6 snRNA and GAPDH, respectively. The
relative fold change for miR-155 and CTHRC1 was calculated using
the 2−ΔΔCt method. Sequences for miR-155, U6, CTHRC1 and
GAPDH primers used in this study are following: miR-155 stem-loop
primer, 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACCCCTA-3′, miR-155
forward primer, 5′-ACACTCCAGCTTAATGCTAATCGTGATAG-3′, miR-155
reverse primer, 5′-CTCAACTGGTGTCGTGGA-3′; U6 stem-loop primer,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAAAAATATGG-3′, U6 forward
primer, 5′-CTCGCTTCGGCAGCACA-3′, U6 reverse primer,
5′-AACGCTTCACGAATTTGCGT-3′; CTHRC1 forward primer,
5′-ACAATTAATATTCATCGCACT-3′, CTHRC1 reverse primer,
5′-ACAATTAATATTCATCGCACT-3′; GAPDH forward primer,
5′-CTGACTTCAACAGCGACACC-3′, GAPDH reverse primer,
5′-TCTGACTTCAACAGCGACACC-3′. The results were expressed as the mean
of three individual experiments with duplicate samples.
Vector construction and
dual-luciferase reporter assay
The wild-type and mutant 3′-UTR of CTHRC1 containing
the seed sequence were synthesized and ordered from Sangon Biotech
Co., Ltd., Shanghai, China. Then, these fragments were cloned into
psi-CHECK2 Basic luciferase reporter plasmid (Promega Corporation)
between the Xho I and Not I sites and the insertions were verified
by sequencing. To identify the direct target relationship between
miR-155 and the 3′-UTR of CTHRC1 mRNA, the 293T cells in 24-well
plates were cotransfected with the above constructed plasmids
concomitant with control psiCHECK-2 plasmid (that is Blank group),
miR-155 mimics, miR-155 inhibitor, NC plasmid or NC inhibitor with
the final dosage of 100 ng using Lipofectamine 2000 according to
the manufacturer's recommendations. After 6 h, the media of the
transfected cells were switched to fresh media. At last, the cells
were split, and Firefly and Renilla relative luciferase activity
was tested with the Dual-Luciferase Reporter Assay System (Promega
Corporation) in accordance with the manufacturer's instructions at
48 h post transfection. The experiments were performed in
triplicate.
MTT assay
Cell proliferation was determined 48 h after
transfection treatment using a
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
kit (Promega Corporation) in a 96-well plate. The results were
subsequently analyzed at 0, 1, 2 and 3 day. For the analysis,
first, the cell culture medium was aspirated from the 96-well
plate, and the cells were gently washed with PBS twice, followed by
the addition of 20 µl of 5 mg/ml MTT reagent. After 4 h of
incubation, MTT was carefully removed without disturbing the cells
and 150 µl of dimethylsulfoxide (DMSO) solution was added to the
wells, which was followed by continuous shaking at room temperature
for 15 min to solubilise the purple formazan crystals. Finally, the
optical density of each well, including the blank well without
cells, was measured at a wavelength of 490 nm in a multimode plate
reader (Bio-Rad, USA). Each sample was assayed in triplicate.
Flow cytometry
The flow cytometry was performed 48 h after HT-29
cells were transfected with three plasmids, namely NC plasmid,
miR-155 mimics plasmid and miR-155 inhibitor plasmid, for apoptosis
and cell cycle analysis. For apoptosis analysis, the harvested
cells were washed by ice-cold PBS, centrifuged and labeled with
FITC-Annexin V and propidium iodide (PI) in binding buffer for 15
min at room temperature in the dark according to manufacturer's
guidelines. Fluorescence signals were examined within 1 h
post-staining on a FACSCalibur flow cytometer. However, for cell
cycle analysis, the harvested cells were washed by ice-cold PBS,
centrifuged, and fixed in 70% ethanol at −20°C overnight. Then the
cells were washed with PBS again and resuspended in 1 ml of PBS
containing 50 mg/ml PI (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) and 25 mg/ml RNase A (Sigma-Aldrich; Merck KGaA) for 30
min at room temperature in the dark. A total number of
1×104 cells were subject to cell cycle analysis by flow
cytometry using a FACSCalibur flow cytometer.
Transwell assay
HT-29 cells with different treatments were
trypsinized to generate a single-cell suspension, and
1×105 cells/well (for the migration assay) or
2×105 cells/well (for the invasion assay) were plated
into 24-transwell chamber with 8 µm pore size membranes (BD
Biosciences, Franklin Lakes, NJ, USA) pre-coated with or without
matrigel basement membrane matrix (BD Biosciences), respectively.
Meanwhile, 600 µl of culture media with 20% FBS as a
chemoattractant was added to the lower chamber. Approximately 24 h
later, the cells on the apical side of each transwell membranes
were gently removed with the mechanical scraping, while the cells
that pass through the membrane were fixed with 4% paraformaldehyde
at room temperature for 30 min, stained with 0.1% crystal violet
for 10 min, washed with PBS three times and ultimately counted from
4~5 randomly microscopic filed. The migration and invasion assays
were repeated at least three times.
Statistical analysis
Statistical analysis of the study data was performed
using SPSS v18.0 software (IBM Corp., Armonk, NY, USA). All values
are expressed as the mean ± standard deviation (SD) of three
independent biological experiments. Independent sample t-test and
one-way analysis of variance (ANOVA) were applied to comparisons
between two or more than two groups, respectively. P<0.05 was
considered to indicate a statistically significant difference..
Results
Different expression levels of miR-155
and CTHRC1 in patient specimens and multiple colorectal cancer cell
lines
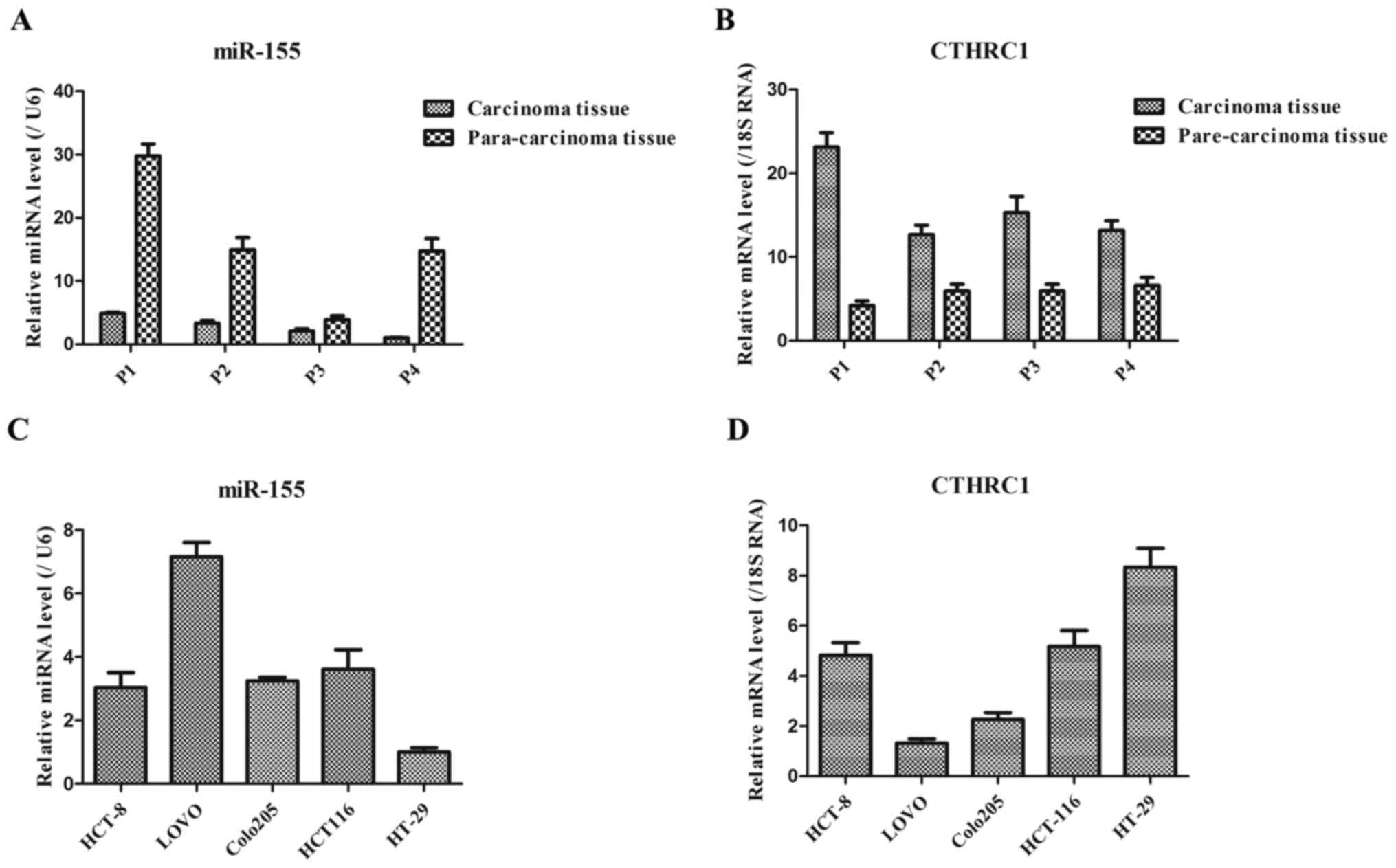
As shown in Fig. 1A,
the expression level of miR-155 in carcinoma tissues of colorectal
cancer patients was generally lower than that in the corresponding
para-carcinoma tissues of these patients. However, the expression
level of CTHRC1 in carcinoma tissues of colorectal cancer patients
was generally higher than that in the corresponding para-carcinoma
tissues of these patients (Fig. 1B).
These results suggested that the down-regulated miR-155 and
up-regulated CTHRC1 might be the key markers in colorectal cancer
patients. Additionally, in five colorectal cancer cell lines, the
expression levels of miR-155 in HCT8, Colo205 and HCT116 cells were
basically consistent, while miR-155 expression presented lowest and
highest in HT-29 and Lovo cells, respectively (Fig. 1C). Moreover, CTHRC1 expression
exhibited the highest in HT-29 cells (Fig. 1D), thereby HT-29 cells were chosen as
following experiment cells in this study.
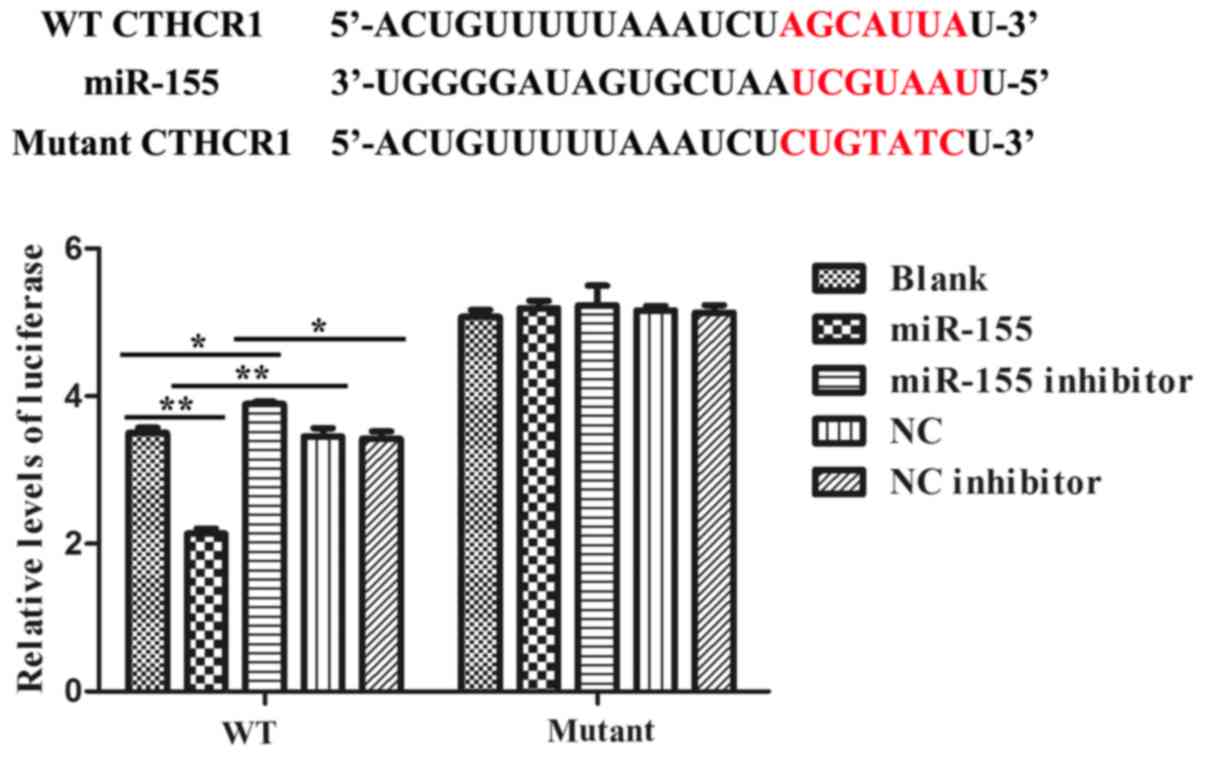
miR-155 directly targeted the 3′-UTR
of CTHRC1
We constructed the dual-luciferase reporter system
to determine the direct interaction between miR-155 and 3′-UTR of
CTHRC1. Alignment between the predicted miR-155 target site and
mutant site of the CTHRC1 3′-UTR region and miR-155 was displayed
in upper part of Fig. 2. And the
results of the dual-luciferase assay revealed that the activity of
luciferase was remarkably decreased after co-transfection with WT
constructive luciferase reporter plasmid harbouring the CTHRC1
3′UTR and miR-155 mimics plasmid. By contrast, the activity of
luciferase was notably increased following co-transfection with WT
constructive luciferase reporter plasmid and miR-155 inhibitor
(Fig. 2). However, there were no
changes in 293T cells co-transfected with the Mutant constructive
luciferase reporter plasmid and other plasmid (P>0.05; Fig. 2). The results have firmly confirmed
that miR-155 specifically binds to the 3′-UTR of CTHRC1.
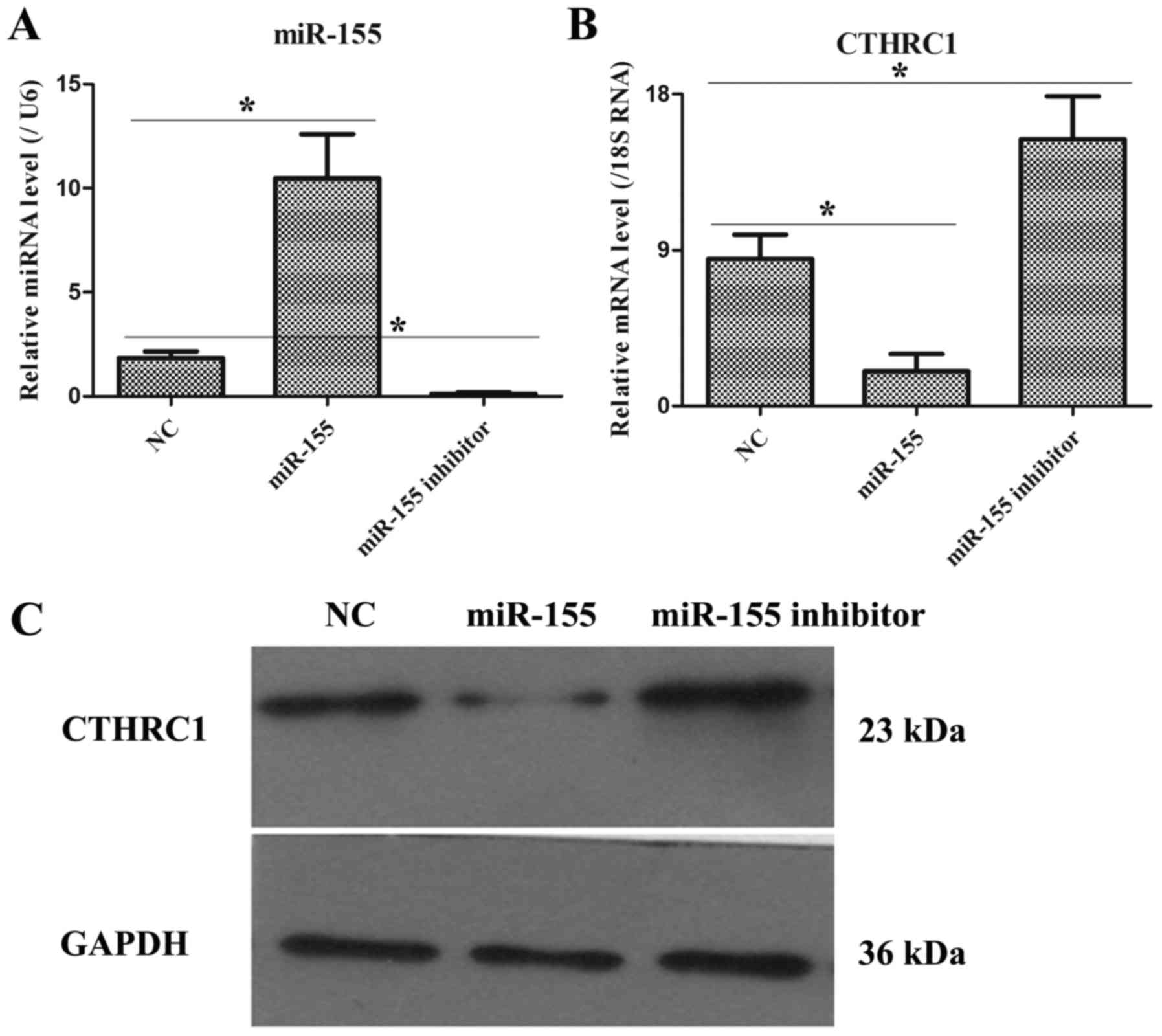
The expression levels of miR-155 and
CTHRC1 in HT-29 cells after transfecting with NC, miR-155 mimic and
miR-155 inhibitor plasmids
As illustrated in Fig.
3A, after transfecting 48 h, miR-155 and CTHRC1 expressions
were remarkably increased and decreased, respectively, in miR-155
group as compared to NC group, but miR-155 and CTHRC1 expressions
were markedly down-regulated and up-regulated, respectively, in
miR-155 inhibitor group compared with NC group (Fig. 3B). Moreover, the protein level of
CTHRC1 was also notably declined and elevated in miR-155 mimic and
miR-155 inhibitor groups, respectively (Fig. 3C). Hence, these data pointed that the
transfection efficiency of miR-155 mimic or miR-155 inhibitor was
obvious.
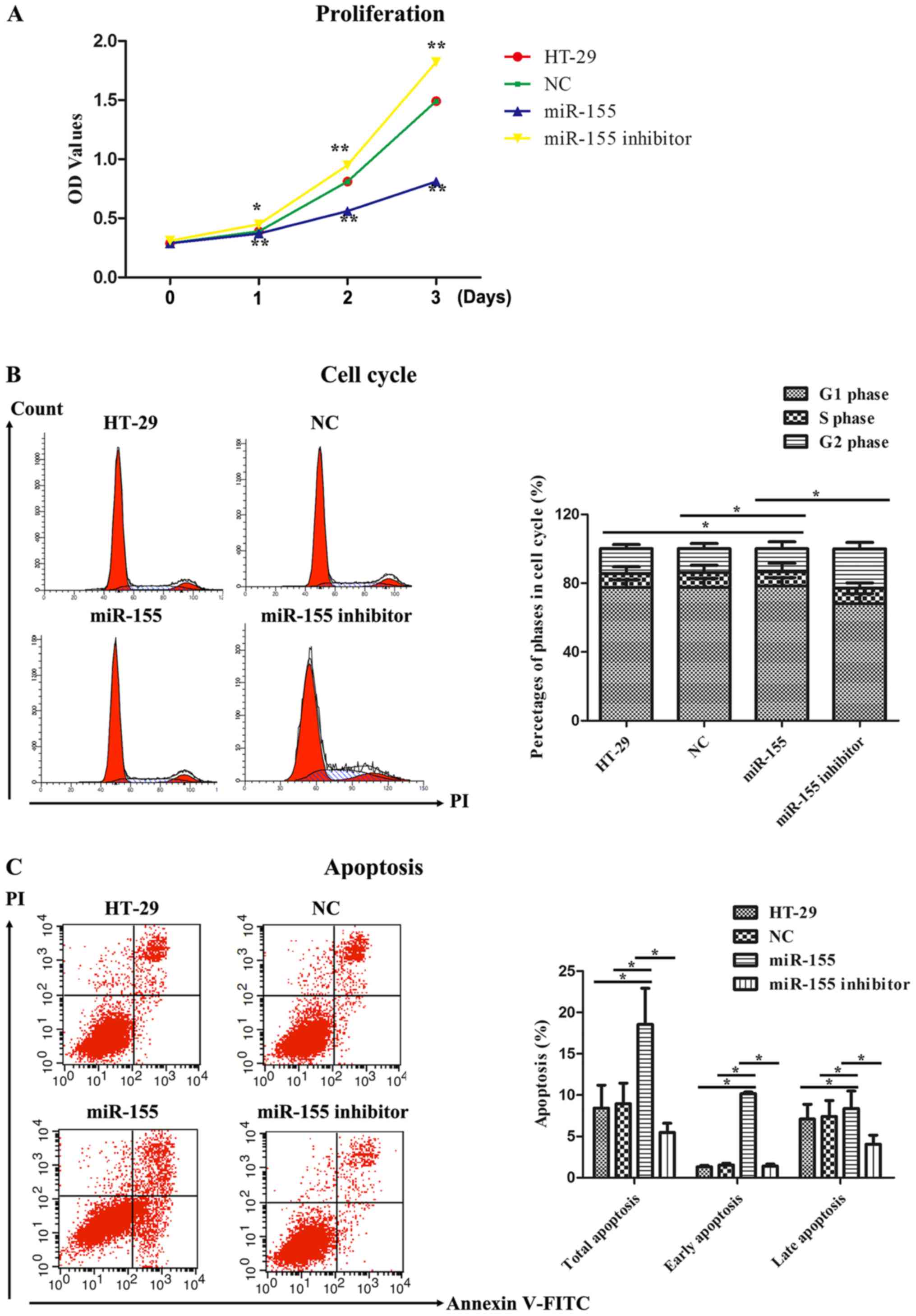
miR-155 affected cell proliferation
activity, cell cycle and cell apoptosis
The proliferation activity of miR-155 on HT-29 cells
was detected by MTT assay. As illustrated in Fig. 4A, the OD values representing cell
proliferation activity in miR-155 group were markedly reduced
compared to that in the control groups, including HT-29 cells group
and NC group, at 1~3 days. Nevertheless, miR-155 inhibitor
obviously reversed the proliferating effect of miR-155 on HT-29
cells. We next examined cell cycle progression in HT-29 cells
treated with miR-155 mimics and miR-155 inhibitor. It was found
that miR-155 induced cell cycle arrest in G0/G1 phase as comparison
with HT-29 cells group and NC group, whereas miR-155 inhibitor
abolished the impacts of cell cycle arrest in G0/G1 phase (Fig. 4B). Finally, cell apoptosis was further
measured by flow cytometry. As displayed in Fig. 4C, Annexin V/PI dual staining was
uncovered that miR-155 enhanced the early and late apoptosis rate
in HT-29 cells. However, miR-155 inhibitor suppressed the early and
late apoptosis rate compared to miR-155. Hence, these data
indicated that miR-155 could inhibit cell proliferation, promote
cell cycle arrest and facilitate cell apoptosis in HT-29 cells.
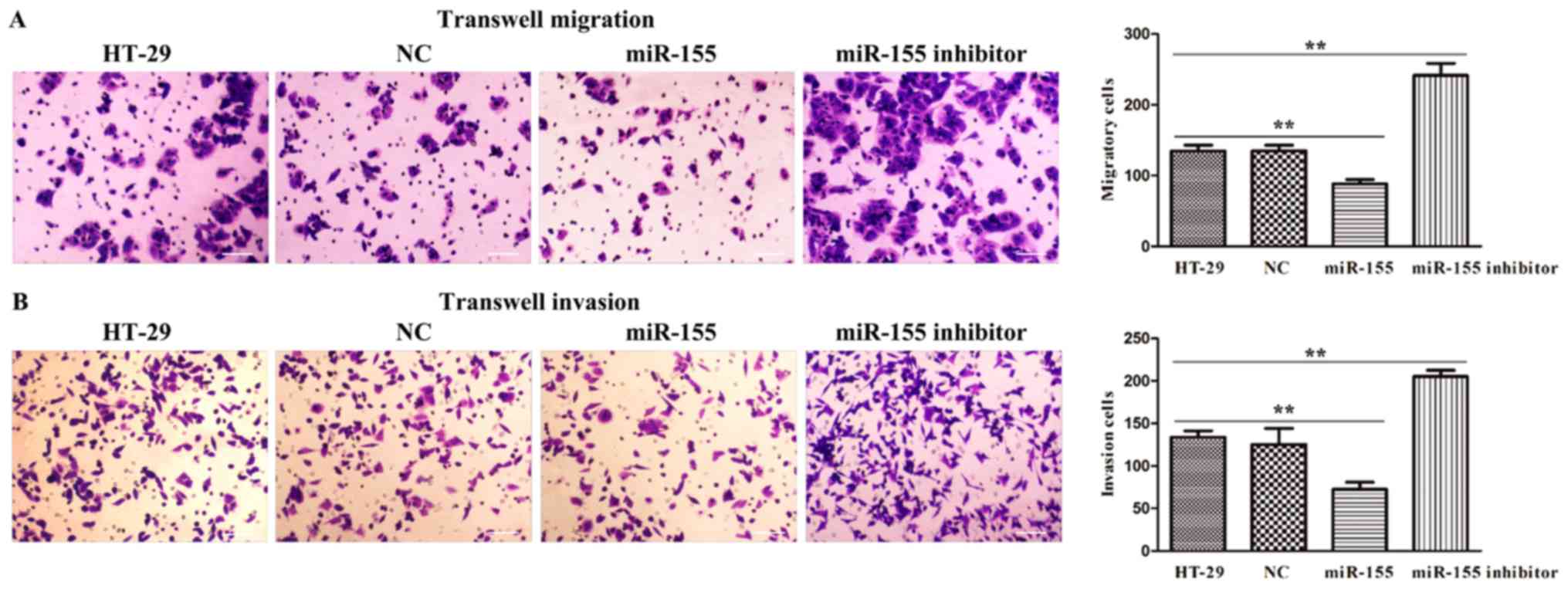
miR-155 regulated cell migration and
invasion in HT-29 cells
To further investigate the biological function of
miR-155 in colorectal cancer, we adopted HT-29 cells transfected
with miR-155 mimics and miR-155 inhibitor plasmids to confirm cell
migration and invasion activity through transwell assays. As
presented in Fig. 5A, it was
discovered that miR-155 was significantly declined the number of
migration and invasion cells, while miR-155 inhibitor was
remarkably elevated the number of migration and invasion cells
(Fig. 5B). Thus, these results
implied that miR-155 negatively regulated cell migratory and
invasive ability of HT-29 cells.
Discussion
Colorectal cancer is known as one of main types of
gastrointestinal cancers which remain an important public health
problem in different populations (22). Although recent advances in medicine
have significantly improved the survival of patients with
early-stage colorectal cancer, patients with advanced colorectal
cancer have poor prognosis mainly due to frequent tumor metastasis
and tumor recurrence after surgical resection (8,23).
Emerging studies show that chromosomal abnormalities, genetic
alterations, epigenetic modifications and unhealthy lifestyle are
decisive factors that initiate and drive the occurrence and
development of colorectal cancer (4,5). Moreover,
miRNAs increasingly reported to be differentially expressed either
as oncogenes (e.g., miR-21, miR-191) or tumor suppressors (e.g.,
let-7, miR-34) play a pivotal role in the pathogenesis and
tumorigenicity of colorectal cancer (24,25).
Otherwise, aberrant miR-155 expression was previously observed in
colorectal cancer (18,26), but the biological function and
regulatory mechanisms of miR-155 in colorectal cancer is still
largely unknown. In this study, we found that expressions of
miR-155 and CTHRC1 in colorectal cancer specimens were remarkably
lower and higher than those in corresponding adjacent tissues,
respectively, suggesting miR-155 and CTHRC1 might exert a crucial
role in colorectal cancer. However, higher expression levels of
miR-155 have been detected in other human malignancies, including
lung cancer, cervical cancer, hematological malignancies and
thyroid carcinoma (18,19). This distinction may be caused by
tissue specificity, ethnic diversity or different tumor stages.
Subsequently, we examined miR-155 and CTHRC1 expressions in five
different colorectal cancer cell lines by qRT-PCR. The result
displayed the lowest expression of miR-155 and the highest
expression of CTHRC1 in HT-29 cells, which was chosen as the
following experiment cell line. Moreover, the opposite expression
trend between miR-155 and CTHRC1 also implied that miR-155 might
target CTHRC1. Functional assays revealed that restoration of
miR-155 notably inhibited cell proliferation, migration, and
invasion, and promoted cell arrested at G0/G1 stage and cell
apoptosis in HT-29 cells. The precise regulation of cell
proliferation, cell cycle and apoptosis is basic premise for
cellular normal growth. Once dysregulation in cell proliferation,
cell cycle and apoptosis, it might lead to cellular abnormal
changes, which ultimately might trigger cell canceration (27). Thus, the alterations of cell
proliferation, cell cycle and apoptosis indicated that miR-155
suppressed the growth and progression of colorectal cancer in
vitro. Additionally, migratory and invasive abilities are
closely associated with tumor metastasis which has been widely
recognized as the important reason for the unsatisfactory prognosis
of colorectal cancer patients (28,29).
Hence, our data implied that miR-155 decelerated the tumor
metastasis of HT-29 cells by inhibiting cell migratory and invasion
behaviors.
CTHRC1, initially screened from differentially
expressed genes in balloon-injured vs. normal rat arteries, is a
well-known regulator of the growth and metastasis of human cancers,
such as lung cancer, pancreas cancer, breast cancer, cervix cancer,
and liver cancer (30). In the
present study, it was confirmed that miR-155 could directly
interact with the 3′-UTR of CTHRC1 using luciferase reporter assay,
suggesting that CTHRC1 is a direct target of miR-155. Furthermore,
inhibition of miR-155 have an opposite effects on these features
related to tumor growth and metastasis compared with miR-155
treatment in HT-29 cells. Based on the targeted interaction between
miR-155 and CTHRC1, as well as the CTHRC1 expression levels after
transfecting with miR-155 mimic and miR-155 inhibitor as shown in
Fig. 3, the function of miR-155
inhibitor should be similar to that of CTHRC1 over-expression,
thereby these results indirectly reflected that CTHRC1 might be an
oncogenic gene in colorectal cancer.
In conclusion, in the current study, we validated
the significance of down-regulated miR-155 expression in colorectal
cancer patients and its tumor suppressive role in HT-29 cells by
attenuating cell growth and metastasis. Additionally, CTHRC1 was
also verified to be downstream target of miR-155 and we adopted an
indirectly method (i.e., utilizing miR-155 inhibitor to replace
CTHCR1 over-expression) to investigate the oncogenic role of CTHRC1
in colorectal cancer in vitro. Taken together, these
findings implicated that miR-155 and its target CTHCR1 might be
used as a prognostic indicator and therapeutic target in colorectal
cancer patients.
Acknowledgements
The present study is supported by Shanghai Emerging
Frontier Technology Joint Research Program (SHDC12016104).
References
|
1
|
Mármol I, Sánchez-de-Diego C, Pradilla
Dieste A, Cerrada E and Rodriguez Yoldi MJ: Colorectal carcinoma: A
general overview and future perspectives in colorectal cancer. Int
J Mol Sci. 18(pii): E1972017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bolocan A, Ion D, Ciocan DN and Paduraru
DN: Prognostic and predictive factors in colorectal cancer.
Chirurgia (Bucur). 107:555–563. 2012.PubMed/NCBI
|
|
3
|
De Rosa M, Rega D, Costabile V, Duraturo
F, Niglio A, Izzo P, Pace U and Delrio P: The biological complexity
of colorectal cancer: Insights into biomarkers for early detection
and personalized care. Therap Adv Gastroenterol. 9:861–886. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fumery M, Dulai PS, Gupta S, Prokop LJ,
Ramamoorthy S, Sandborn WJ and Singh S: Incidence, risk factors,
and outcomes of colorectal cancer in patients with ulcerative
colitis with low-grade dysplasia: A systematic review and
meta-analysis. Clin Gastroenterol Hepatol. 15:665–674.e5. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Coppedè F, Lopomo A, Spisni R and Migliore
L: Genetic and epigenetic biomarkers for diagnosis, prognosis and
treatment of colorectal cancer. World J Gastroenterol. 20:943–956.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang J, Du Y, Liu X, Cho WC and Yang Y:
MicroRNAs as regulator of signaling networks in metastatic colon
cancer. Biomed Res Int. 2015:8236202015.PubMed/NCBI
|
|
7
|
Berretta M, Alessandrini L, De Divitiis C,
Nasti G, Lleshi A, Di Francia R, Facchini G, Cavaliere C, Buonerba
C and Canzonieri V: Serum and tissue markers in colorectal cancer:
State of art. Crit Rev Oncol Hematol. 111:103–116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lech G, Słotwiński R, Słodkowski M and
Krasnodębski IW: Colorectal cancer tumour markers and biomarkers:
Recent therapeutic advances. World J Gastroenterol. 22:1745–1755.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lech G, Slotwinski R and Krasnodebski IW:
The role of tumor markers and biomarkers in colorectal cancer.
Neoplasma. 61:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yamazaki N, Koga Y, Taniguchi H, Kojima M,
Kanemitsu Y, Saito N and Matsumura Y: High expression of miR-181c
as a predictive marker of recurrence in stage II colorectal cancer.
Oncotarget. 8:6970–6983. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer. 96
Suppl:R40–R44. 2007.PubMed/NCBI
|
|
13
|
Ruan K, Fang X and Ouyang G: MicroRNAs:
Novel regulators in the hallmarks of human cancer. Cancer Lett.
285:116–126. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giannakakis A, Coukos G, Hatzigeorgiou A,
Sandaltzopoulos R and Zhang L: miRNA genetic alterations in human
cancers. Expert Opin Biol Ther. 7:1375–1386. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pagliuca A, Valvo C, Fabrizi E, di Martino
S, Biffoni M, Runci D, Forte S, De Maria R and Ricci-Vitiani L:
Analysis of the combined action of miR-143 and miR-145 on oncogenic
pathways in colorectal cancer cells reveals a coordinate program of
gene repression. Oncogene. 32:4806–4813. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Guo X, Zhang H, Xiang Y, Chen J,
Yin Y, Cai X, Wang K, Wang G, Ba Y, et al: Role of miR-143
targeting KRAS in colorectal tumorigenesis. Oncogene. 28:1385–1392.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang G, Zhou H, Xiao H, Liu Z, Tian H and
Zhou T: MicroRNA-92a functions as an oncogene in colorectal cancer
by targeting PTEN. Dig Dis Sci. 59:98–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wan J, Xia L, Xu W and Lu N: Expression
and function of miR-155 in diseases of the gastrointestinal tract.
Int J Mol Sci. 17(pii): E7092016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hou Y, Wang J, Wang X, Shi S, Wang W and
Chen Z: Appraising MicroRNA-155 as a noninvasive diagnostic
biomarker for cancer detection: A meta-analysis. Medicine
(Baltimore). 95:e24502016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan L, Yu J, Tan F, Ye GT, Shen ZY, Liu H,
Zhang Y, Wang JF, Zhu XJ and Li GX: SP1-mediated microRNA-520d-5p
suppresses tumor growth and metastasis in colorectal cancer by
targeting CTHRC1. Am J Cancer Res. 5:1447–1459. 2015.PubMed/NCBI
|
|
21
|
Wang JP: Chinese standard for the
diagnosis and treatment of colorectal cancer (2010). Zhonghua Wei
Chang Wai Ke Za Zhi. 14:1–4. 2011.(In Chinese). PubMed/NCBI
|
|
22
|
Binefa G, Rodriguez-Moranta F, Teule A and
Medina-Hayas M: Colorectal cancer: From prevention to personalized
medicine. World J Gastroenterol. 20:6786–6808. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sideris M and Papagrigoriadis S: Molecular
biomarkers and classification models in the evaluation of the
prognosis of colorectal cancer. Anticancer Res. 34:2061–2068.
2014.PubMed/NCBI
|
|
24
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moridikia A, Mirzaei H, Sahebkar A and
Salimian J: MicroRNAs: Potential candidates for diagnosis and
treatment of colorectal cancer. J Cell Physiol. 233:901–913. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He B, Gao SQ, Huang LD, Huang YH, Zhang
QY, Zhou MT, Shi HQ, Song QT and Shan YF: MicroRNA-155 promotes the
proliferation and invasion abilities of colon cancer cells by
targeting quaking. Mol Med Rep. 11:2355–2359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Zhao F, Wang Z, Song Y, Luo Y, Zhang
X, Jiang L, Sun Z, Miao Z and Xu H: MicroRNA-335 acts as a
metastasis suppressor in gastric cancer by targeting Bcl-w and
specificity protein 1. Oncogene. 31:1398–1407. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He Z, Yu L, Luo S, Li M, Li J, Li Q, Sun Y
and Wang C: miR-296 inhibits the metastasis and
epithelial-mesenchymal transition of colorectal cancer by targeting
S100A4. BMC Cancer. 17:1402017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang L, Dai DL, Su M, Martinka M, Li G and
Zhou Y: Aberrant expression of collagen triple helix repeat
containing 1 in human solid cancers. Clin Cancer Res. 12:3716–3722.
2006. View Article : Google Scholar : PubMed/NCBI
|