Introduction
Osteosarcoma is the most common malignant tumor of
the bone and remains the second leading cause of cancer-associated
mortality in adolescents globally at present (1). Surgery combined with adjuvant
chemotherapy is currently the standard treatment for osteosarcoma
(1). In recent years, although a
great deal of effort has been made toward improving chemotherapy
regimens, the overall prognosis remains poor and much of this may
be attributed to drug resistance (2).
P-glycoprotein (P-gp), an ATP-binding cassette (ABC) membrane
transporter encoded by multidrug resistance 1 (MDR1), is commonly
located at the plasma membrane and functions as an ATP-dependent
efflux pump for diverse naturally occurring hydrophobic anticancer
drugs, including Adriamycin (ADR) (3). Finding an efficient method of inhibiting
drug resistance may contribute to better therapeutic outcomes.
Vasohibin (VASH)1 was first identified to be a
negative feedback modulator of angiogenesis in vascular endothelial
cells in a previous study (4).
Inhibitory functions of mesenchymal VASH1 in tumor progression have
been reported in different types of tumor (5–7). The
functions of parenchymal VASH1 in tumor development have drawn more
and more attention, but relevant reports remain limited. Liu et
al (2) reported that
overexpression of VASH1 in colon cancer cells was able to induce
apoptosis and senescence, and inhibited cancer cell growth and
colony formation in vitro and tumor growth in vivo.
In addition, knockdown of VASH1 in cancer cells was able to promote
cell growth, adhesion and migration in vitro and enhance
tumorigenesis and metastasis in vivo (8). Takahashi et al (9) reported that VASH1 overexpression in
ovarian cells inhibited ovarian cancer growth and peritoneal
dissemination and prolonged host survival. Thus far, there remains
no report on the functions of VASH1 in osteosarcoma to the best of
our knowledge.
In the present study, it was identified that VASH1
is underexpressed in osteosarcoma cells. It was also revealed that
VASH1 was able to inhibit ADR resistance of osteosarcoma cells
through regulation of the protein kinase B (AKT) signaling pathway.
This suggested that further evaluation of VASH1 may yield a novel
therapeutic approach to the treatment of osteosarcoma.
Materials and methods
Cell culture
The human osteoblast cell line hFOB1.19 and human
osteosarcoma cell lines U-2OS and 143B were purchased from American
Type Culture Collection (Manassas, VA, USA). All cells were
cultured in Dulbecco's modified Eagle's medium (DMEM) (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Invitrogen; Thermo Fisher Scientific,
Inc.), at 37°C in 5% CO2.
Drug resistance assay
U-2OS and 143B cells were counted and plated in
96-well plate at 10,000 cells/well. After 24 h, the culture medium
was replaced with DMEM containing different concentrations (2, 4,
8, 16, 32 µmol/l) of ADR (HarveyBio, Inc., Beijing, China). These
cells served as experimental groups. Cells in medium without ADR
served as the control group. After 48 h, an MTT assay kit (Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China) was
used and the optical density (OD) value was measured at 490 nm
wavelength using an ultraviolet spectrophotometer (Shanghai
Spectrum Instrument Co., Ltd., Shanghai, China) according to the
manufacturer's protocol according to the manufacturer's protocol.
Inhibition rate (IR) was calculated using the following equation:
IR = 1 - OD value of experiment group/OD value of control group
×100%. Half maximal inhibitory concentration (IC50) was
calculated using regression analysis by SPSS 11.0 software (SPSS,
Inc., Chicago, IL, USA). All experiments were repeated 3 times.
Cell transfection
U-2OS and 143B cells were counted and plated in
6-well plates at 2×105 cells/well. After 24 h,
p-GPU6/Neo/VASH1 (Shanghai GenePharma Co. Ltd., Shanghai, China) to
silence VASH1 expression, and pEZM61/VASH1 (Gene Copoeia,
Guangzhou, China) to overexpress VASH1 were transfected using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Empty plasmids were used as control. Reverse
transcription-polymerase chain reaction (RT-PCR) and western
blotting were performed to confirm transfection efficiency.
RT-PCR
RNA was extracted from cells using TRIzol (Life
Sciences; Thermo Fisher Scientific, Inc.) according to the
manufacturers protocol. cDNA was synthesized using a PrimeScript
RT-PCR kit (Takara Biotechnology Co., Ltd., Dalian, China). PCR was
performed using specific primers and Universal PCR Master Mix
(Thermo Fisher Scientific, Inc.). The thermocycling conditions were
as follows; VASH1, 4°C for 5 min, 94°C for 30 sec, 57°C for 30 sec,
72°C for 30 sec for 40 cycles and 72°C for 5 min; P-gp, 4°C for 5
min, 94°C for 30 sec, 54°C for 30 sec, 72°C for 30 sec for 36
cycles and 72°C for 5 min; GAPDH, 4°C for 5 min, 94°C for 30 sec,
57°C for 30 sec, 72°C for 30 sec for 36 cycles and 72°C for 5 min.
PCR products were electrophoretically separated on 1.0% agarose
gel. Results were analyzed using Labwork software (version 4; UVP,
Inc., Upland, CA, USA). All primers were as follows: VASH1 forward,
5′-CCACGCCCTGATTTCTTAAA-3′ and reverse, 5′-CCCTGTCAGAGGTCTGCTCT-3′;
P-gp forward, 5′-CCCATCATTGCAATAGCAGG-3′ and reverse,
5′-GTTCAAACTTCTGCTCCTGA-3′; GAPDH forward,
5′-AGAAGGCTGGGGCTCATTTG-3′ and reverse, 5′-AGGGGCCATCCACAGTCTTC-3′
GAPDH served as an internal control. All experiments were repeated
3 times.
Western blotting
Protein was extracted from cells using
radioimmunoprecipitation assay lysis containing 1% phenylmethane
sulfonyl fluoride (Beyotime Institute of Biotechnology, Haimen,
China) and protein concentration was analyzed using a bicinchoninic
acid assay kit (Pierce; Thermo Fisher Scientific, Inc.). Equal
quantities of protein were loaded per well in 5% acrylamide and
separated using 10% SDS-PAGE, and transferred to nitrocellulose
membrane. The membrane was incubated with primary antibodies
(Table I) at 4°C overnight, and then
in horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (cat. no. ab205718; 1:4,000; Abcam, Cambridge, MA, USA) at
room temperature for 1 h. Signals were detected using enhanced
chemiluminescence reagents (Pierce; Thermo Fisher Scientific, Inc.)
and quantified using Image-Pro software (version 5.1; Media
Cybernetics, Inc., Rockville, MA, USA). All experiments were
repeated 3 times.
 | Table I.Primary antibodies used in western
blotting. |
Table I.
Primary antibodies used in western
blotting.
| Name | Cat. no. | Dilution | Supplier |
|---|
| VASH1 | Ab199732 | 1:300 | Abcam, Cambridge,
UK |
| P-gp | Ab170904 | 1:400 | Abcam, Cambridge,
UK |
| phospho-ERK1/2 | AF1018 | 1:1,000 | Cell Signaling
Technology, Inc., Danvers, MA, USA |
| ERK1/2 | AF1576 | 1:1,000 | Cell Signaling
Technology, Inc., Danvers, MA, USA |
| phospho-AKT | AF887 | 1:1,000 | Cell Signaling
Technology, Inc., Danvers, MA, USA |
| AKT | AF2055 | 1:1,000 | Cell Signaling
Technology, Inc., Danvers, MA, USA |
| GAPDH | Ab9485 | 1:4,000 | Abcam, Cambridge,
UK |
Statistical analysis
SPSS software (version 11.0; SPSS, Inc., Chicago,
IL, USA) was used and data were expressed as mean ± standard
deviation. Differences between groups were analyzed using one-way
analysis of variance with Dunnett's post hoc test. IC50
was calculated using regression analysis. P<0.05 was considered
to indicate a statistically significant difference.
Results
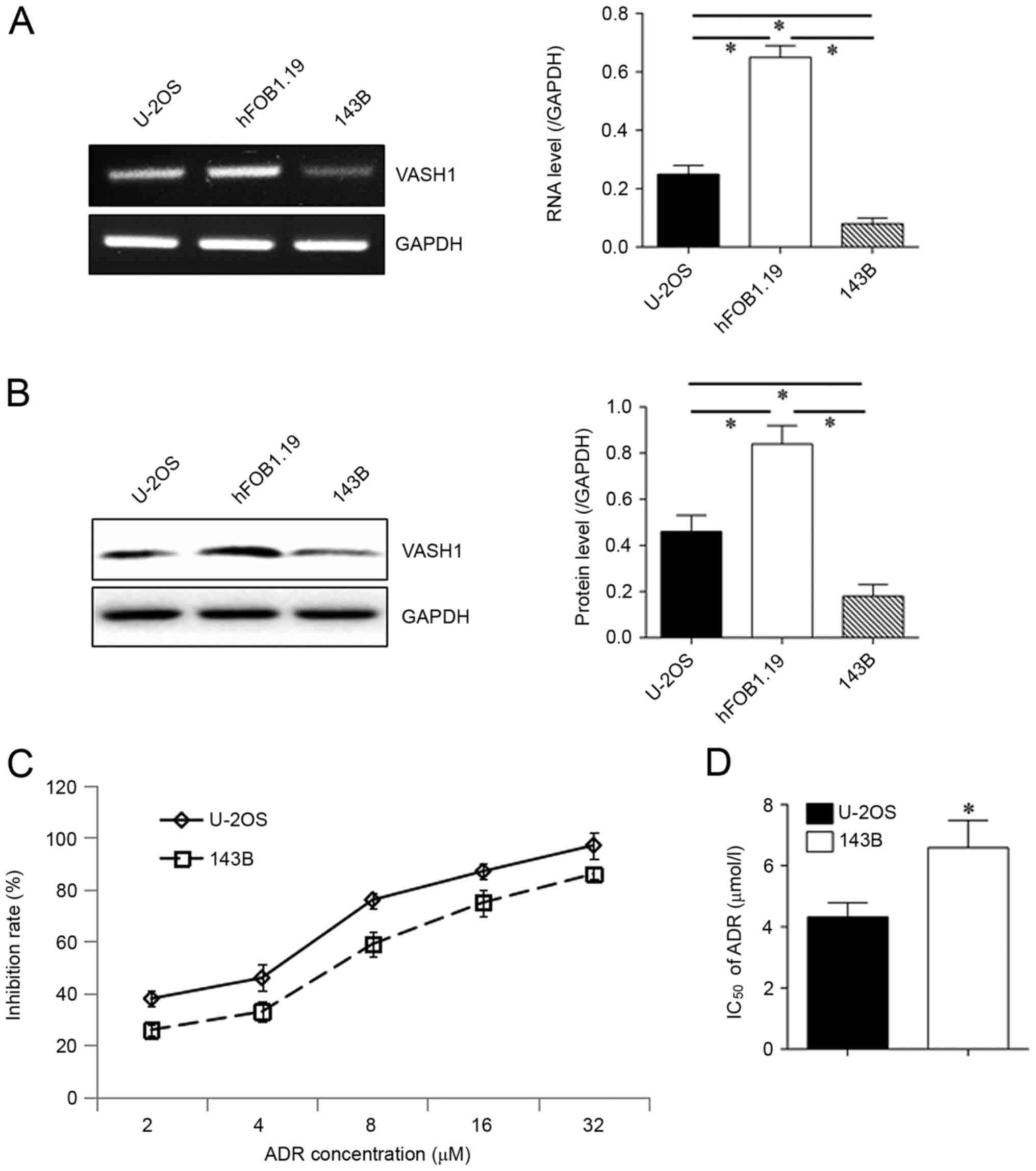
VASH1 is expressed weakly in
osteosarcoma cells
VASH1 expression was exhibited both at RNA (Fig. 1A) and protein (Fig. 1B) levels. Compared with human
osteoblast cell lines hFOB1.19, decreased VASH1 expression was
detected in osteosarcoma cell lines U-2OS and 143B. VASH1
expression was significantly decreased in 143B cells compared with
that in U-2OS cells. A drug resistance assay was subsequently
performed, revealing that the inhibition rate (IR) of 143B cells in
ADR was decreased compared with that of U-2OS (Fig. 1C). The IC50 of 143B cells
(6.59±0.89 µmol/l) was significantly increased compared with that
of U-2OS cells (4.32±0.47 µmol/l; Fig.
1D). All these results indicate possible associations between
VASH1 expression and drug resistance.
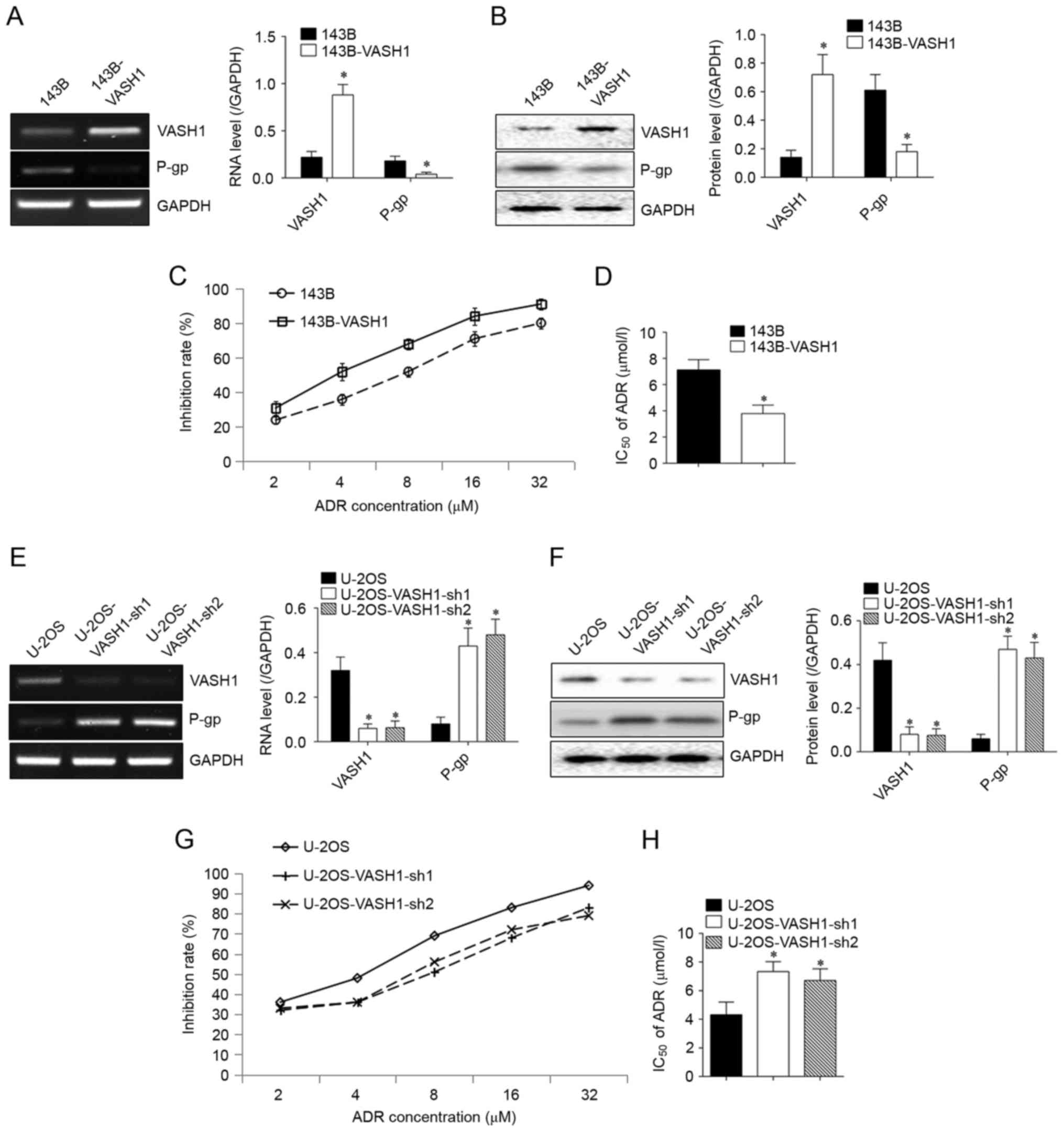
VASH1 inhibits the ADR resistance of
osteosarcoma cells
To confirm whether VASH1 was able to regulate drug
resistance of osteosarcoma cells, VASH1 expression was manipulated
through transfection. Following overexpression of VASH1 in 143B
cells, P-glycoprotein (P-gp) expression was significantly inhibited
at both the RNA (Fig. 2A) and protein
(Fig. 2B) levels. The IR of 143B
cells was increased compared with control cells (Fig. 2C). IC50 declined from
7.14±0.83 to 3.79±0.56 µmol/l (Fig.
2D). Following silencing of VASH1 in U-2OS cells, P-gp
expression was upregulated both at RNA (Fig. 2E) and protein (Fig. 2F) levels. IR of U-2OS cells declined
significantly (Fig. 2G),
IC50 increased from 4.32±0.88 to 7.34±0.69 or 6.71±0.82
µmol/l (Fig. 2H). All results
suggested the inhibitory function of VASH1 in ADR resistance.
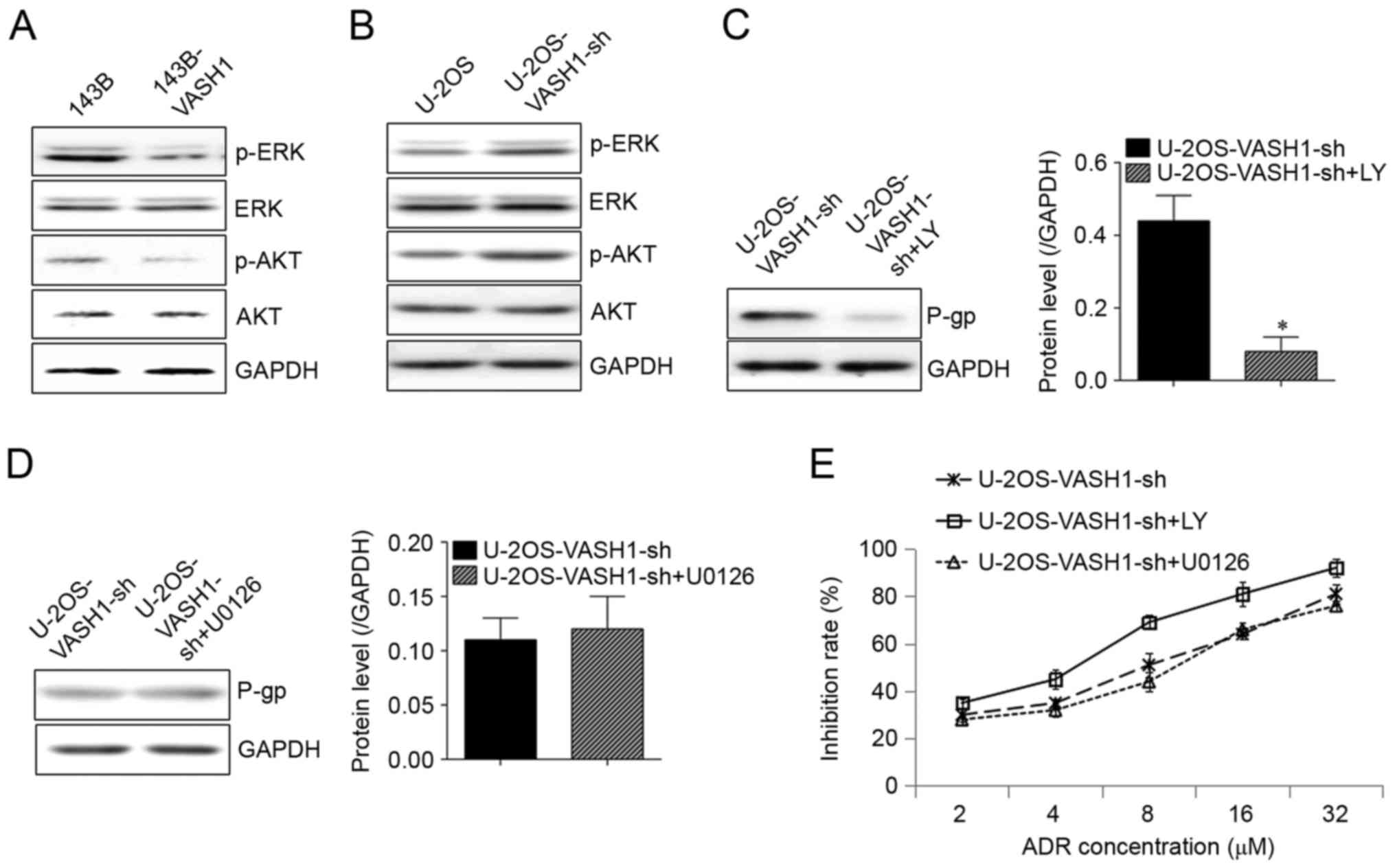
VASH1 regulation of ADR resistance
uses the AKT signaling pathway
As presented in Fig.
3A, following overexpression of VASH1 in 143B cells,
phosphorylation of extracellular signal-related kinase (ERK) and
AKT was inhibited (Fig. 3A).
Conversely, following silencing of VASH1 in U-2OS cells,
phosphorylation of ERK and AKT was upregulated (Fig. 3B). Once AKT inhibitor LY294002 was
added, the increase of P-gp in U-2OS cells induced by silencing
VASH1 was decreased (Fig. 3C).
However, with ERK inhibitor U0126 added, no change was observed in
P-gp expression (Fig. 3D). A drug
resistance assay also revealed that LY294002 could counteract the
decrease of IR of U-2OS cells in ADR induced by silencing VASH1,
but U0126 did not influence declination of IR of U-2OS cells in ADR
induced by silencing VASH1. This suggests that the AKT signaling
pathway may serve a function in ADR resistance regulated by VASH1
(Fig. 3E).
Discussion
A member of the vasohibin family, the human VASH1
gene is located on chromosome 14q24.3. VASH1 protein is composed of
365 amino acids with no glycosylation sites (10,11).
Vasohibin 2 is also a member of the vasohibin family and was
initially known as an angiogenic factor. VASH1 was first noticed
for its ability to inhibit angiogenesis; it is restricted in vessel
endothelial cells and several other types of cell (12). The negative regulation of VASH1 from
tumor cells on tumor progression has been demonstrated in colon
cancer (8), ovarian (9) and renal carcinoma (13). However, in 2014, Kitajima et al
(14) reported that high VASH1 in the
cytoplasm of colorectal cancer (CRC) tissues was positively
associated with tumor progression, and silencing VASH1 inhibited
CRC cell proliferation, migration and invasion, and promoted
anoikis. Thus, the functions of VASH1 in different types of tumor
are not consistent, therefore the effects of VASH1 on osteosarcoma
require further investigation.
Drug resistance is an important characteristic of
malignant tumors and has been an important factor in the failure of
cancer treatment (15). ATP-binding
cassette drug efflux pump P-gp has been proposed to serve crucial
functions for tumor cells acquiring MDR (16,17). ADR
is the first-line chemotherapy drug used to treat osteosarcoma. It
not only inhibits DNA transcription and replication but also
induces breakage of DNA double strands (18). In the present study, data revealed low
expression of VASH1 in osteosarcoma cells at both the RNA and
protein levels. Furthermore, osteosarcoma cells with lower VASH1
levels exhibited more marked ADR resistance. This suggests that
VASH1 may serve negative regulatory functions in osteosarcoma drug
resistance. Through changing VASH1 using transfection, it was
identified that VASH1 was able to inhibit the P-gp expression and
ADR resistance of osteosarcoma cells. This is consistent with the
negative regulatory functions of VASH1 reported by the majority of
works (8,9,13), but
inconsistent with a report from Kitajima et al (14). Different organs of origin of different
tumors may explain this divergence.
The AKT and ERK signaling pathways may be stimulated
in different types of tumors. This activates proliferation and
survival signals that ultimately lead to tumorigenesis and
progression (19). In 2016, Yang
et al (20) reported that the
ERK signal pathway may serve important functions in 5-FU-mediating
of colorectal cancer. Xiao et al (21) identified that Oridonin inhibits
gefitinib-resistant lung cancer cells by suppressing ERK and AKT
signaling pathways. In the present study, it was identified that
VASH1 downregulated P-gp expression by blocking the AKT signal
pathway, thus inhibiting ADR resistance of osteosarcoma cells. In
the present study, no effects of ERK were observed in ADR
resistance. This is not consistent with relevant reports from Yang
et al (20) and Xiao et
al (21). This may be attributed
to differences in the type of tumor and drugs with different
anti-tumor mechanisms.
To conclude, the inhibitory effects of VASH1 on
osteosarcoma drug resistance were confirmed. This has enhanced
understanding of the functions of VASH1 in tumors and supplied a
basis for ongoing studies targeting VASH1. VASH1 may be treated as
an enhancer of chemotherapeutic sensitivity in osteosarcoma cells
to foster better prognosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HL designed this study. WH performed the
experiments. YR analyed the results.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sobhan MR, Forat Yazdi M, Mazaheri M, Zare
Shehneh M and Neamatzadeh H: Association between the DNA repair
gene XRCC3 rs861539 polymorphism and risk of osteosarcoma: A
systematic review and meta-analysis. Asian Pac J Cancer Prev.
18:549–555. 2017.PubMed/NCBI
|
|
2
|
Liu R, Fu C, Sun J, Wang X, Geng S, Wang
X, Zou J, Bi Z and Yang C: A new perspective for osteosarcoma
therapy: Proteasome inhibition by MLN9708/2238 successfully induces
apoptosis and cell cycle arrest and attenuates the invasion ability
of osteosarcoma cells in vitro. Cell Physiol Biochem. 41:451–465.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Colabufo NA, Contino M, Cantore M,
Capparelli E, Perrone MG, Cassano G, Gasparre G, Leopoldo M,
Berardi F and Perrone R: Naphthalenyl derivatives hitting for
P-gp/MRP1/BCRP transporters. Bioorg Med Chem. 21:1324–1332. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coch L, Mejias M, Berzigotti A,
Garcia-Pras E, Gallego J, Bosch J, Mendez R and Fernandez M:
Disruption of negative feedback loop between vasohibin-1 and
vascular endothelial growth factor decreases portal pressure,
angiogenesis, and fibrosis in cirrhotic rats. Hepatology.
60:633–647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Watanabe K, Hasegawa Y, Yamashita H,
Shimizu K, Ding Y, Abe M, Ohta H, Imagawa K, Hojo K, Maki H, et al:
Vasohibin as an endothelium-derived negative feedback regulator of
angiogenesis. J Clin Invest. 114:898–907. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miyashita H, Watanabe T, Hayashi H, Suzuki
Y, Nakamura T, Ito S, Ono M, Hoshikawa Y, Okada Y, Kondo T and Sato
Y: Angiogenesis inhibitor vasohibin-1 enhances stress resistance of
endothelial cells via induction of SOD2 and SIRT1. PLoS One.
7:e464592012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miyashita H, Suzuki H, Ohkuchi A and Sato
Y: Mutual balance between vasohibin-1 and soluble VEGFR-1 in
endothelial cells. Pharmaceuticals. 4:782–793. 2011. View Article : Google Scholar
|
|
8
|
Liu S, Han B, Zhang Q, Dou J, Wang F, Lin
W, Sun Y and Peng G: Vasohibin-1 suppresses colon cancer.
Oncotarget. 6:7880–7898. 2015.PubMed/NCBI
|
|
9
|
Takahashi Y, Saga Y, Koyanagi T, Takei Y,
Machida S, Taneichi A, Mizukami H, Sato Y, Matsubara S and Fujiwara
H: The angiogenesis regulator vasohibin-1 inhibits ovarian cancer
growth and peritoneal dissemination and prolongs host survival. Int
J Oncol. 47:2057–2063. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang T, Yu TT, Zhang DM, Hou XM, Liu XJ,
Zhao D and Shan L: Vasohibin-1 expression detected by
immunohistochemistry correlates with prognosis in non-small cell
lung cancer. Med Oncol. 31:9632014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan Y, Shen Z, Ye Y, Jiang K, Zhang H,
Shen C, Mustonen H, Puolakkainen P and Wang S: A novel molecular
marker of prognosis in colorectal cancer: Vasohibin-1. Med Oncol.
31:8162014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kern J, Steurer M, Gastl G, Gunsilius E
and Untergasser G: Vasohibin inhibits angiogenic sprouting in vitro
and supports vascular maturation processes in vivo. BMC Cancer.
9:2842009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao G, Yang Y, Tang Y, Han R and Sun Y:
Reduced expression of vasohibin-1 is associated with
clinicopathological features in renal cell carcinoma. Med Oncol.
29:3325–3334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kitajima T, Toiyama Y, Tanaka K, Saigusa
S, Kobayashi M, Inoue Y, Mohri Y and Kusunoki M: Vasohibin-1
increases the malignant potential of colorectal cancer and is a
biomarker of poor prognosis. Anticancer Res. 34:5321–5329.
2014.PubMed/NCBI
|
|
15
|
He S, Shen J, Hu N, Xu X and Li J: DKK4
enhances resistance to chemotherapeutics 5-Fu and YN968D1 in
colorectal cancer cells. Oncol Lett. 13:587–592. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tandia M, Mhiri A, Paule B, Saffroy R,
Cailliez V, Noé G, Farinotti R and Bonhomme-Faivre L: Correlation
between clinical response to sorafenib in hepatocellular carcinoma
treatment and polymorphisms of P-glycoprotein (ABCB1) and of breast
cancer resistance protein (ABCG2): Monocentric study. Cancer
Chemother Pharmacol. 79:759–766. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Esser L, Zhou F, Pluchino KM, Shiloach J,
Ma J, Tang WK, Gutierrez C, Zhang A, Shukla S, Madigan JP, et al:
Structures of the multidrug transporter P-glycoprotein reveal
asymmetric ATP binding and the mechanism of polyspecificity. J Biol
Chem. 292:446–461. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao M, Yu S and Zhang M: Differential
expression of multidrug resistance-related proteins in
Adriamycin-resistant (pumc-91/ADM) and parental (pumc-91) human
bladder cancer cell lines. Mol Med Rep. 14:4741–4746. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu YL, Maachani UB, Schweitzer M, Singh R,
Wang M, Chang R and Souweidane MM: Dual inhibition of PI3K/AKT and
MEK/ERK pathways induces synergistic antitumor effects in diffuse
intrinsic pontine glioma cells. Transl Oncol. 10:221–228. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang K, Gao K, Hu G, Wen Y, Lin C and Li
X: CTGF enhances resistance to 5-FU-mediating cell apoptosis
through FAK/MEK/ERK signal pathway in colorectal cancer. Onco
Targets Ther. 9:7285–7295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiao X, He Z, Cao W, Cai F, Zhang L, Huang
Q, Fan C, Duan C, Wang X, Wang J and Liu Y: Oridonin inhibits
gefitinib-resistant lung cancer cells by suppressing
EGFR/ERK/MMP-12 and CIP2A/Akt signaling pathways. Int J Oncol.
48:2608–2618. 2016. View Article : Google Scholar : PubMed/NCBI
|