Introduction
Malignant glioma is the most common type of
intracranial primary tumor, with a high mortality rate and poor
treatment outcomes (1,2). The mean survival time of patients
following surgery, combined with chemotherapy and/or radiotherapy,
is still <1 year (2,3). Glucocorticoids are a type of steroid
hormone with extensive physiological and pharmacological effects
(4). One function is an
anti-proliferative effect that may be observed in a variety of
tissues and cells, including lymphoid tissue, fibroblast tissue and
epithelial cells, through inducing cell cycle arrest in the
G1 phase, or through apoptosis (5–7).
Furthermore, previous studies have demonstrated that
glucocorticoids have an inhibitory effect on the growth of various
tumor cell types, and have been clinically used in the treatment of
a number of malignant tumors, including chronic lymphocytic
leukemia and prostate cancer (8–10).
Dexamethasone (Dex) is frequently used as a clinical synthetic
glucocorticoid, as it has a stronger efficacy than natural
glucocorticoids (11,12). It is now a commonly used agent for
clinical hormone therapy; however, the mechanism underlying Dex is
still unclear (13,14).
Aquaporin-1 (AQP1) is a highly conserved
membrane-bound protein with a molecular size of 28 kDa. This
protein serves an important role in the specific transmembrane
transport of water molecules (15).
The gene is located on the human chromosome 7p14 (15). In the central nervous system (CNS),
AQP1 is mainly expressed in choroid plexus epithelial cells,
fulfilling a dual role as a water channel and in the regulation of
c-GMP ion channels, which are involved in the formation of
cerebrospinal fluid (16). Malignant
gliomas originate in the neural epithelium and are the most common
primary CNS tumors. According to the histological origin, glioma
can be divided into astrocytoma, oligodendroglioma,
oligodendrocytoma and ependymoma (17). The majority of low-grade astrocytomas
can develop into malignant gliomas (17). The malignant glioma phenotype is
characterized by rapid tumor growth, high glucose consumption,
tumor necrosis, hypoxia, an increase in microvessel density, and
the destruction of the blood-brain barrier (17). Prior studies have demonstrated that
AQP1 is expressed in astrocytoma cells and vascular endothelial
cells, and is increased in parallel with the histological grade of
gliomas (18,19). Further studies revealed that AQP1 is
closely associated with tumor angiogenesis, tumor-associated brain
edema and tumor metastasis (20,21). AQP1
expression in the gliosarcoma cell line is induced by Dex,
platelet-derived growth factors, sodium chloride, hypoxia,
D-glucose and fructose (22). Based
on the expression of AQP1 in gliomas, and on the existing studies
regarding its function, the hypothesis investigated in the present
study was that AQP1 may participate in tumor cell proliferation,
apoptosis, migration and invasion.
The present study aimed to investigate the function
of AQP1 and its mechanism using C6 cells. C6 cells are rat brain
glioma cells obtained following N-nitrosomethylurea-induced glioma
in Wistar rats (23,24). These cells were subcultured in
vitro and in vivo, are stable in growth and express
glioma specific markers, including glial fibrillary acidic protein
and the S-100 protein; therefore, C6 cells are widely used in in
experimental studies for the treatment of glioma (25–27). The
roles of AQP1 and Dex in cell proliferation, apoptosis, migration
and invasion in C6 cells were investigated, to determine whether
AQP1 can be used as a novel therapeutic target to inhibit the
proliferation and metastasis of gliomas.
Materials and methods
Cell culture
Rat glioma C6 cells (iCell Bioscience Inc.,
Shanghai, China) were cultured in Dulbecco's modified Eagle's
medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 100 ml/l fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 kU/l penicillin and 100 mg/l chloramphenicol
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in a humidified
5/95% CO2/air incubator at 37°C. The cell growth state
was observed under an inverted light microscope (×100
magnification; Olympus Corporation, Tokyo, Japan). To detect the
role of AQP1, small interfering RNA (siRNA) was transfected into
the C6 cells causing the knockdown of AQP1; the transfected cells
were used for subsequent studies with 1 µM Dex treatment.
MTT test
Cell proliferation was detected using an MTT assay
(Beyotime Institute of Biotechnology, Haimen, China). A total of
100 µl DMEM with 10% FBS containing a concentration of
1×105 cells/ml C6 cells were added to 96-well plates and
were cultured for 24 h; following this, the cells were treated with
Dex (0, 0.01, 0.1 and 1 µM; Sigma-Aldrich; Merck KGaA) for 24 or 48
h in a humidified 5/95% CO2/air incubator at 37°C. A
total of 20 µl MTT solution (0.5 mg/ml) was added into each well
and incubated for 4 h at 37°C, followed by a 150-µl
dimethylsulfoxide culture on an oscillator for 15 min, to dissolve
the formazan crystals. Finally, absorbance of the solution was
measured at 570 nm using an ST-360 microplate reader (Shanghai
Kehua Bio-Engineering Co., Ltd., China) and the rates of cell
proliferation inhibition were calculated. The formula used was as
follows: The inhibition rate of cell proliferation
(%)=(1-absorbance value of the experimental group/that of the
control group) ×100%.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.), homogenized using a
pipette, and the RNA concentrations were measured with a NanoDrop™
2000 spectrophotometer (Thermo Fisher Scientific, Inc.). Following
this, the extracted RNA was transcribed into cDNA using a
PrimeScript™ RT-PCR kit (RR014A; Takara Bio, Inc., Otsu, Japan) and
the RT-qPCR reactions were performed on a T100™qPCR system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) using TransStart®
Top Green qPCR SuperMix AQ131 (TransGen Biotech, Inc., Beijing,
China). Pre-denaturation consisted of 1 cycle at 95°C and 5 sec;
and RT-qPCR reactions consisted of 35 cycles: 95°C for 30 sec,
followed by 55°C for 30 sec, 72°C for 1 sec, 72°C for 5 sec, and
16°C for 30 sec. The primers used were as follows: Forward
5′-CTGTGGTGGCTGAGTTCCTG-3′ and reverse 5′-ACCTCGGCCAAGTGAGTTCTC-3′
for AQP1; and forward 5′-ACCACAGTCCATGCCATCAC-3′ and reverse
5′-TCCACCACCCTGTTGCTGTA-3′ for GAPDH. Gene expression was
normalized to the level of GAPDH within each sample using the
relative 2−∆∆Cq method (28). Gene expression is presented as
relative to the control, and the data are representative of three
independent experiments.
Transwell assay for migration and
invasion
After being pretreated with Dex (0, 0.01, 0.1 and 1
µM) for 48 h, the C6 cells were digested with 0.25% EDTA trypsin,
then collected and C6 cells were resuspended in DMEM with 10% FBS
at a density of 5×105 cells/ml. Following this, C6 cells
(5×105) were seeded into the upper chambers of a
Transwell plate (200 µl/well). Subsequently, 600 µl DMEM containing
20% FBS was added into each lower chamber. After being cultured for
24 h in a humidified 5/95% CO2/air incubator at 37°C,
the cells were fixed in 4% paraformaldehyde for 10 min, washed
twice with PBS and then stained with 0.05% crystal violet for 10
min at 37°C. After removing the cells that had not migrated, the
cells on the lower surfaces were quantified and images were
obtained. To study invasion, 50 µl of 1 g/l Matrigel was used to
coat the upper chambers of the Transwell plate at 37°C for 1 h.
Apoptosis rate
The rate of C6 cell apoptosis was detected using the
terminal deoxynucleotidyl-transferase-mediated dUTP nick-end
labeling (TUNEL) method with In Situ Cell Death Detection Kit (cat
no. 11684795910, Roche Applied Science, Penzberg, Germany)
according to the manufacture's instruction. C6 cells at a
concentration of 1×105/ml were inoculated into 24-well
plates prior to culturing with Dex (0, 0.01, 0.1 and 1 µM) for 24 h
in a 5% CO2 saturated humidity incubator at 37°C.
Following 2 washes with PBS, cells were fixed using 4%
paraformaldehyde in PBS (pH 7.4) for 1 h at 22–25°C.
Permeabilisation solution with 0.1% Triton X-100 in 0.1% sodium
citrate were used for incubation on ice for 2 min. Cell samples
were resuspended in 50 µl/well TUNEL reaction mixture containing
enzyme solution with terminal deoxynucleotidyl transferase from
calf thymus in storage buffer and label solution with nucleotide
mixture in reaction buffer (cat no. 11684795910, Roche Applied
Science, Penzberg, Germany) mixed at a ratio of 1:9. Samples were
incubated for 60 min at 37°C in the dark and washed twice using
PBS. 50 µl of DAPI Staining Solution WH1150 (Biotechwell, Shanghai,
China) were added into every well, incubation for 10 min at room
temperature. After 3 washes with PBS, 50 µl of fluoroshield (F6182,
Sigma) were added to preserve fluorescence of cell smears. The
green fluorescence at 520±20 nm and the blue DAPI staining at 460
nm were observed via fluorescence microscopy using a fluorescein
isothiocyanate (FITC)-labeled TUNEL Apoptosis Detection kit (Roche
Applied Science, Penzberg, Germany). Three different fields of view
were randomly photographed per sample. DAPI can stain apoptotic and
non-apoptotic cells blue, and the green fluorescence of
FITC-12-dUTP is incorporated and localized in the apoptotic nuclei.
Cell apoptosis was confirmed by Hoechst staining.
Western blot analysis
After pretreatment with benzo[a]pyrene
(0,5,10,50,100 and 200 µM) for various lengths of time, C6 cells
were lysed in RIPA lysis buffer (Beyotime Institute of
Biotechnology) and centrifuged at 12,000 × g for 10 min at 4°C.
Protein concentration was measured using a BCA assay (Pierce;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. A total of 30 µg protein from each sample was
resolved via 10% SDS-PAGE. The proteins were then transferred to a
nitrocellulose membrane at 100 V for 120 min. After blocking with
5% non-fat dry milk for 1 h at room temperature, the membranes were
incubated with anti-AQP1 (cat no. ab122821; 1:100; Abcam,
Cambridge, MA, USA) and anti-GAPDH (cat no. ab8245; 1:1,000; Abcam)
antibodies overnight at 4°C. After 3 washes with PBS, the membranes
were incubated with goat anti-rabbit (cat no. 32460; 1:1,000;
Invitrogen; Thermo Fisher Scientific, Inc.) or goat anti-mouse IgG
(cat no. A0216; 1:1,000; Beyotime Institute of Biotechnology)
secondary antibodies conjugated with horseradish peroxidase for 1 h
at room temperature. Detection was performed using the ChemiDoc™
XRS+ enhanced chemiluminescent system (Bio-Rad Laboratories,
Inc.).
Statistical analysis
Data are presented as the mean ± standard deviation.
A value of P<0.05 was considered to indicate statistical
significance, using one-way analysis of variance with least
significant difference post-hoc test in SPSS version 12.0 package
(SPSS, Inc. Chicago, IL, USA). Images were prepared using GraphPad
Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Results
Effects of Dex on C6 cell
proliferation, apoptosis, migration and invasion
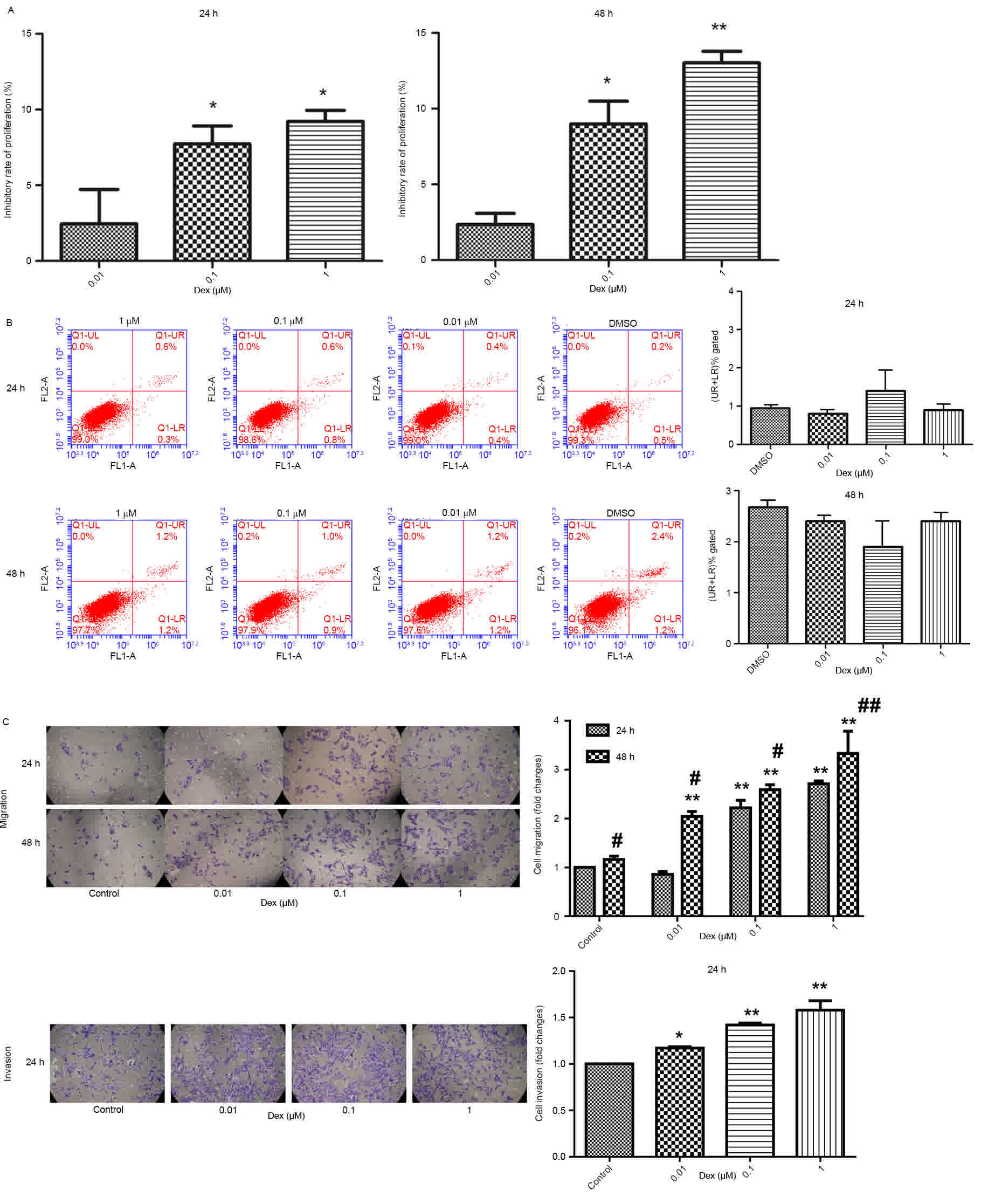
The inhibitory effect of Dex on C6 cell
proliferation was studied (Fig. 1A).
The results indicated that Dex had a significant dose-dependent
inhibitory effect on the proliferation of C6 cells at 24 and 48 h
(P<0.05).
There were no significant changes in cell apoptosis
with various concentrations of Dex (0, 0.01, 0.1 and 1 µM) for 24
and 48 h (P<0.05; Fig. 1B). Cell
migration and invasion were also studied (Fig. 1C). At 24 and 48 h, the migration of C6
cells was significantly increased (P<0.05), in a dose-dependent
manner. In addition, cell migration at 48 h was greater than cell
migration at 24 h, demonstrating that the promotion of cell
migration by Dex was dose- and time-dependent; similarly, Dex
promoted C6 cell invasion.
Dex upregulates the expression of
AQP1
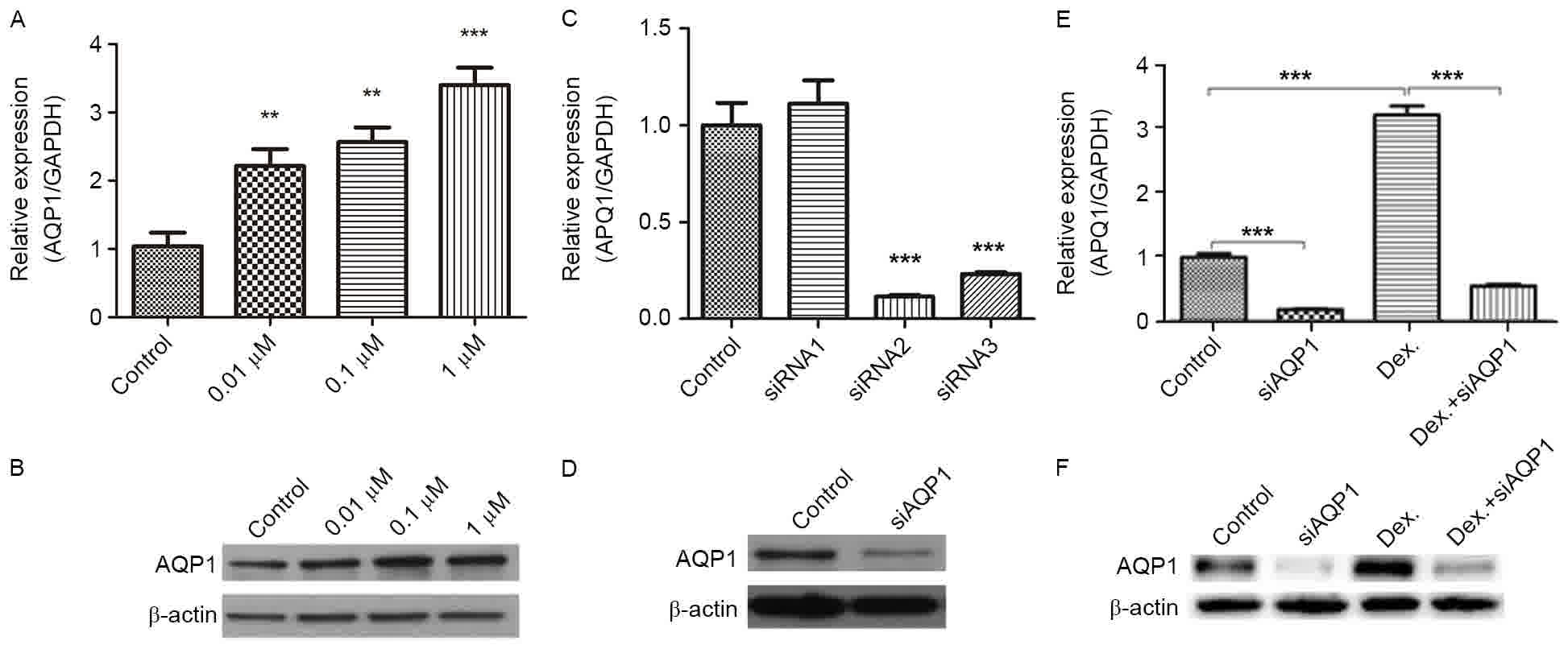
The expression level of AQP1 induced by Dex was
studied (Fig. 2A and B). The results
indicated that the transcription and translation of AQP1 gene could
be enhanced by treatment with Dex for 24 h. Regarding mRNA and
protein levels, the expression levels of AQP1 significantly
increased along with the Dex dose, demonstrating that the changes
in AQP1 expression were dose-dependent. Due to 1 µM Dex having the
greatest significance in inhibiting cell proliferation (P<0.05),
enhancing the expression of AQP1 and promoting cell migration and
invasion, 1 µM Dex was used to detect the roles of AQP1 in the
proliferation, migration and invasion of C6 cells.
To further study the role of AQP1, three siRNAs used
to silence AQP1 in C6 cells were designed (Fig. 2C and D). The results identified that
siRNA-2 and −3 could significantly reduce the transcription level
of AQP1 in C6 cells (Fig. 2C)
(P<0.05). Additionally, siRNA-2 was demonstrated to
significantly reduce the levels of AQP1 protein expression
(P<0.05); therefore, siRNA-2 was used in subsequent studies.
In the C6 cells transfected with AQP1 siRNA
(siAQP1), 1 µM Dex did not increase the expression of AQP1
(Fig. 2E and F); thus, AQP1 siRNA
antagonized the Dex-induced upregulation of AQP1.
Effects of siAQP1 on Dex-induced C6
cell proliferation, migration and invasion
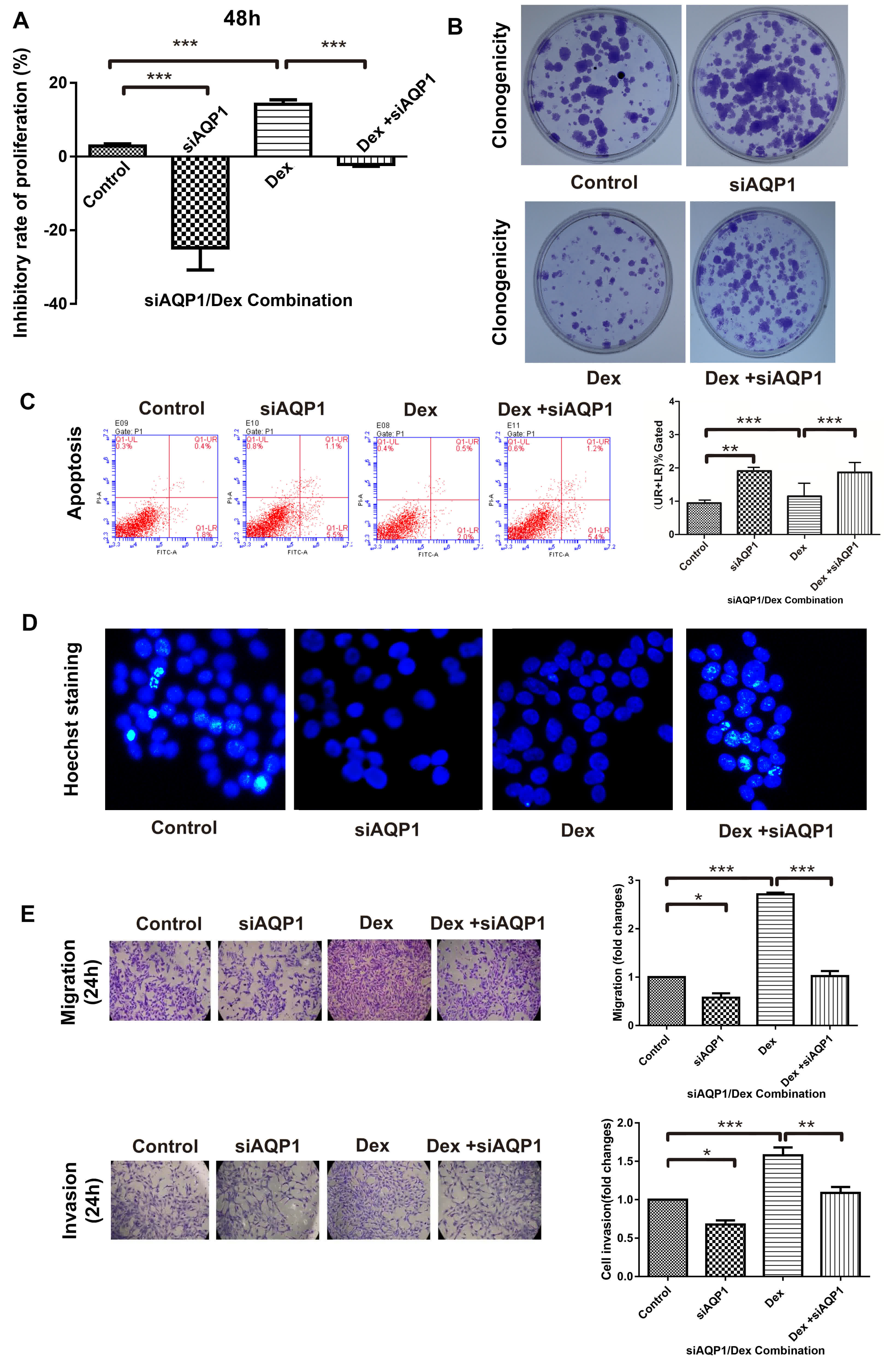
Dex inhibited the proliferation of cells, whereas
siAQP1 promoted proliferation (Fig.
3A), indicating that Dex-inhibited cell proliferation is
mediated by AQP1. The proliferation of cells caused by Dex was
confirmed by a clonogenicity assay (Fig.
3B). siAQP1 increased clonal formation, which was inhibited by
Dex, and Dex-inhibited cell proliferation.
The results for flow cytometry (Fig. 3C) and Hoechst staining (Fig. 3D) demonstrated that cell apoptosis was
promoted by siAQP1, but treatment with Dex did not further enhance
cell apoptosis; this confirmed the effect on cell apoptosis was not
Dex dose-dependent.
The results revealed that the migration of C6 cells
increased after treatment with Dex (Fig.
3E). siAQP1, having the ability to inhibit Dex-induced
migration, demonstrated that Dex facilitates cell migration through
the upregulation of AQP1 expression levels. Similar to cell
migration, siAQP1 inhibited Dex-induced invasion, which indicates
that Dex also facilitates cell invasion through the upregulation of
AQP1 expression levels.
Discussion
Dex inhibited the proliferation of C6 cells, and
promoted the migration and invasion of C6 cells. In addition, Dex
promoted the expression of AQP1. With the downregulation of AQP1 by
using siRNA, the inhibition of cell proliferation and promotion of
cell migration and invasion were reversed; therefore, Dex inhibited
the proliferation of C6 cells and promoted the migration and
invasion of C6 cells via AQP1.
Studies have indicated that Dex can inhibit the
proliferation of a variety of cancer cell lines (29,30). Dex
also inhibits the proliferation of Chinese hamster ovary cells,
HeLa S3 cervical cancer cells, and the human ovarian cancer cell
line 3AO (8). The results of the
present study demonstrated that Dex had a significant inhibitory
effect on the proliferation of C6 cells at 24 and 48 h. The
inhibitory rate was dose-dependent. Cell apoptosis and cell cycle
progression are closely associated (31). When the DNA is damaged or replication
is incomplete, the cell cycle will remain in a certain phase, until
DNA repair or replication is complete, prior to progressing onto
the next phase (31,32). If the DNA cannot be repaired or cannot
be completely replicated, apoptosis is initiated (33,34).
Recent studies on fibroblasts, lung cancer, bladder cancer and
hepatocarcinoma cells revealed that glucocorticoids can induce an
increase in p21/WAF1 levels, causing the cell cycle to become
arrested in the G1 phase (11,35). It
was demonstrated that high-dose Dex also caused C6 cell apoptosis
(36,37). In the present study, it was
demonstrated that 0–1 µM Dex did not induce apoptosis of C6 cells.
Clinically, Dex is only used as an adjuvant drug for the treatment
of brain glioma, to reduce the toxicity of anti-tumor drugs and to
reduce the inflammatory reaction. Migration and invasion of cells
are important markers for metastasis (38,39). Dex
also inhibits the sodium nitroprusside-induced invasion of U87MG
cells (40). The addition of
carmustine and Dex inhibited invasion in the C6, U251, U373 and
A172 glioma cell lines (41). It was
also reported that Dex administered orally to patients with glioma
resulted in positive outcomes (42);
however, whether Dex inhibited glioma cell invasion without the
involvement of other factors was previously unclear (22). The sensitivity of different glioma
cells to Dex may differ; it was demonstrated that 1 µM Dex
significantly inhibited the invasion and migration of U373MG cells,
and that 10 µM Dex inhibited cell proliferation (43). The present findings indicated that Dex
could be used to inhibit the proliferation and promote the
metastasis of the C6 glioma cell line. The difference between the
results for U373MG and C6 cells may be due to the cell type. The
results of the present study demonstrated that Dex functions as an
enhancer for metastasis in C6 cells; therefore, the use of Dex
needs further consideration due to it inhibiting cell
proliferation, but also inducing cell migration and invasion.
AQP1 expression was detected in astrocytomas,
ependymomas and oligodendrogliomas (20,44). AQP1
expression increases with tumor grade. For low-grade astrocytomas,
AQP1 is only located in the tumor cell membrane, whereas for
high-grade astrocytoma, AQP1 is also distributed in the cytoplasm
(18,19). Compared with cells in the necrotic
area of the tumor center, the expression of AQP1 in cells near the
tumor periphery increased (21). By
contrast, in the low-grade glioma, AQP1 expression in the non-tumor
adjacent tissue was greater compared with that in the high-grade
astrocytoma (20). AQP1 is expressed
in reactive astrocytes in the paracancerous tissues of
astrocytomas, indicating a distribution of microvascular colonies
(20,21). In AQP1-knockout mice, tumor
angiogenesis and endothelial cell migration was significantly
reduced; therefore, aquaporin-dependent cell migration may be a
common phenomenon in tumors. Regarding the migration process,
through rapid formation and contraction, cells constantly adjust
their own volume to adapt to the external small space and promote
the forward movement of cells (45).
AQP1 binds to Lin7/β-catenin, causing actin recombination to form
plasma membrane processes and mediating rapid transport of water
molecules into pseudopodia, by promoting the rapid renewal of
plasma membrane processes and increasing water permeability, which
eventually leads to promotion of tumor cell migration (46,47).
Through the knockdown of AQP1 in glioma cells, cell migration and
invasion decreased significantly, indicating that AQP1 mediated the
migration and invasion of glioma cells in the brain, which is
consistent with previous studies (22,48).
Hayashi et al (36) tested the
effect of Dex (10−3-10 µM) on the expression of AQP1 in
rat 9L cells and determined that the expression of AQP1 induced by
10 µM Dex was lower than that recorded for 0.01–1 µM Dex. The
presence of AQP1 in glioma cell lines is closely associated with
enhanced cell migration and invasion (21,48).
Consistently, the present results further indicated that Dex
inhibits proliferation, and promotes the migration and invasion of
C6 cells via the induction of AQP1. Therefore, Dex can inhibit
proliferation and induce apoptosis in cultured C6 glioma cells via
upregulating the expression of AQP1, thus laying a foundation for
the treatment of glioma. In the present study, only C6 cells were
used; as a result, more and varied cell lines should be used for
further investigations.
Acknowledgements
The authors would like to thank Haikou People's
Hospital Center Laboratory and Hainan Medical University First
Affiliated Hospital Central Laboratory for their technical
assistance. The authors would also like to thank Xiangying Li
(Department of Radiology, Haikou People's Hospital, Haikou, China)
and Xiangjun Han (Department of Radiology, KangYa Hospital, Yiyang,
China) for finding literature and guiding experiments.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81360228) and Haikou
key project (grant no. 2014-079).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JC and HL designed the experiments, YG carried out
all the experiments, analyzed the data and wrote the paper. YZ
interpreted the results and contributed to writing of the paper.
All authors have read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AQP1
|
aquaporin-1
|
|
CNS
|
central nervous system
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
siRNA
|
small interfering RNA
|
References
|
1
|
Marshall GM, Carter DR, Cheung BB, Liu T,
Mateos MK, Meyerowitz JG and Weiss WA: The prenatal origins of
cancer. Nat Rev Cancer. 14:277–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rich JN and Bigner DD: Development of
novel targeted therapies in the treatment of malignant glioma. Nat
Rev Drug Discov. 3:430–446. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vandewalle J, Luypaert A, De Bosscher K
and Libert C: Therapeutic mechanisms of glucocorticoids. Trends
Endocrinol Metab. 29:42–54. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Funakoshi Y, Shiono H, Inoue M, Kadota Y,
Ohta M, Matsuda H, Okumura M and Eimoto T: Glucocorticoids induce
G1 cell cycle arrest in human neoplastic thymic epithelial cells. J
Cancer Res Clin Oncol. 131:314–322. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cabrelle A, Maschio N, Carraro S, Frezzato
F, Binotto G, Gattazzo C, Miorin M, Agostini C, Zambello R,
Pandolfi F, et al: Apoptotic effect of cyclosporin a and
dexamethasone in malignant cells of patients with B-chronic
lymphocytic leukemia. J Biol Regul Homeost Agents. 23:239–250.
2009.PubMed/NCBI
|
|
7
|
Bucak YY, Erdurmus M, Terzi EH, Kükner A
and Celebi S: Inhibitory effects of topical cyclosporine A 0.05% on
immune-mediated corneal neovascularization in rabbits. Graefes Arch
Clin Exp Ophthalmol. 251:2555–2561. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Smolej L, Doubek M, Panovská A, Simkovič
M, Brychtová Y, Belada D, Motyčková M and Mayer J: Rituximab in
combination with high-dose dexamethasone for the treatment of
relapsed/refractory chronic lymphocytic leukemia. Leuk Res.
36:1278–1282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Šimkovič M, Motyčková M, Belada D, Vodárek
P, Kapoor R, Jaffar H, Vrbacký F, Žák P and Smolej L: Five years of
experience with rituximab plus high-dose dexamethasone for
relapsed/refractory chronic lymphocytic leukemia. Arch Med Sci.
12:421–427. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fuse H, Nozaki T, Fujiuchi Y, Mizuno I,
Nagakawa O and Okumura A: Treatment with prednisolone of
hormone-refractory prostate cancer. Arch Androl. 52:35–38. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miura N, Numata K, Kusuhara Y, Shirato A,
Hashine K and Sumiyoshi Y: Docetaxel-prednisolone combination
therapy for Japanese patients with hormone-refractory prostate
cancer: A single institution experience. Jpn J Clin Oncol.
40:1092–1098. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ladoire S, Eymard JC, Zanetta S, Mignot G,
Martin E, Kermarrec I, Mourey E, Michel F, Cormier L and
Ghiringhelli F: Metronomic oral cyclophosphamide prednisolone
chemotherapy is an effective treatment for metastatic
hormone-refractory prostate cancer after docetaxel failure.
Anticancer Res. 30:4317–4323. 2010.PubMed/NCBI
|
|
13
|
Juszczak GR and Stankiewicz AM:
Glucocorticoids, genes and brain function. Prog
Neuropsychopharmacol Biol Psychiatry. 82:136–168. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meng J and Li L: The efficiency and safety
of dexamethasone for pain control in total joint arthroplasty: A
meta-analysis of randomized controlled trials. Medicine
(Baltimore). 96:e71262017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deen PM, Weghuis DO, Geurs van Kessel A,
Wieringa B and van Os CH: The human gene for water channel
aquaporin 1 (AQP1) is localized on chromosome 7p15->p14.
Cytogenet Cell Genet. 65:243–246. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boassa D, Stamer WD and Yool AJ: Ion
channel function of aquaporin-1 natively expressed in choroid
plexus. J Neurosci. 26:7811–7819. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cohen AL and Colman H: Glioma biology and
molecular markers. Cancer Treat Res. 163:15–30. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Oshio K, Binder DK, Liang Y, Bollen A,
Feuerstein B, Berger MS and Manley GT: Expression of the
aquaporin-1 water channel in human glial tumors. Neurosurgery.
56:375–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saadoun S, Papadopoulos MC, Davies DC,
Bell BA and Krishna S: Increased aquaporin 1 water channel
expression in human brain tumours. Br J Cancer. 87:621–623. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deb P, Pal S, Dutta V, Boruah D, Chandran
VM and Bhatoe HS: Correlation of expression pattern of aquaporin-1
in primary central nervous system tumors with tumor type, grade,
proliferation, microvessel density, contrast-enhancement and
perilesional edema. J Cancer Res Ther. 8:571–577. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
El Hindy N, Bankfalvi A, Herring A,
Adamzik M, Lambertz N, Zhu Y, Siffert W, Sure U and Sandalcioglu
IE: Correlation of aquaporin-1 water channel protein expression
with tumor angiogenesis in human astrocytoma. Anticancer Res.
33:609–613. 2013.
|
|
22
|
Liao ZQ, Ye M, Yu PG, Xiao C and Lin FY:
Glioma-associated oncogene homolog1 (Gli1)-Aquaporin1 pathway
promotes glioma cell metastasis. BMB Rep. 49:394–399. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peterson DL, Sheridan PJ and Brown WE Jr:
Animal models for brain tumors: Historical perspectives and future
directions. J Neurosurg. 80:865–876. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mattei TA, Ramina R, Miura FK, Aguiar PH
and Valiengo Lda C: Genetic therapy in gliomas: Historical analysis
and future perspectives. Neurol India. 53:17–26. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim SJ, Hwang E, Yi SS, Song KD, Lee HK,
Heo TH, Park SK, Jung YJ and Jun HS: Sea buckthorn leaf extract
inhibits glioma cell growth by reducing reactive oxygen species and
promoting apoptosis. Appl Biochem Biotechnol. 182:1663–1674. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Whittle IR, Macarthur DC, Malcolm GP, Li
M, Washington K and Ironside JW: Can experimental models of rodent
implantation glioma be improved? A study of pure and mixed glioma
cell line tumours. J Neurooncol. 36:231–242. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mead C and Pentreath VW: Evaluation of
toxicity indicators in rat primary astrocytes, C6 glioma and human
1321N1 astrocytoma cells: Can gliotoxicity be distinguished from
cytotoxicity? Arch Toxicol. 72:372–380. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kubota H, Fukuta K, Yamada K, Hirose M,
Naruyama H, Yanai Y, Yamada Y, Watase H, Kawai N, Tozawa K and
Yasui T: Feasibility of metronomic chemotherapy with
tegafur-uracil, cisplatin, and dexamethasone for
docetaxel-refractory prostate cancer. J Rural Med. 12:112–119.
2017. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wong J, Tran LT, Lynch KA and Wood LJ:
Dexamethasone exacerbates cytotoxic chemotherapy induced lethargy
and weight loss in female tumor free mice. Cancer Biol Ther.
19:87–96. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Branzei D and Foiani M: Regulation of DNA
repair throughout the cell cycle. Nat Rev Mol Cell Biol. 9:297–308.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chiarugi A, Dölle C, Felici R and Ziegler
M: The NAD metabolome-a key determinant of cancer cell biology. Nat
Rev Cancer. 12:741–752. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Eto I: Upstream molecular signaling
pathways of p27(Kip1) expression: Effects of 4-hydroxytamoxifen,
dexamethasone, and retinoic acids. Cancer Cell Int. 10:32010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang JK, Li CJ, Liao HJ, Wang CK, Wang GJ
and Ho ML: Anti-inflammatory drugs suppress proliferation and
induce apoptosis through altering expressions of cell cycle
regulators and pro-apoptotic factors in cultured human osteoblasts.
Toxicology. 258:148–156. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jing Y, Qian Y and Li ZJ: Sialylation
enhancement of CTLA4-Ig fusion protein in Chinese hamster ovary
cells by dexamethasone. Biotechnol Bioeng. 107:488–496. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hayashi Y, Edwards NA, Proescholdt MA,
Oldfield EH and Merrill MJ: Regulation and function of aquaporin-1
in glioma cells. Neoplasia. 9:777–787. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Moroz MA, Huang R, Kochetkov T, Shi W,
Thaler H, de Stanchina E, Gamez I, Ryan RP and Blasberg RG:
Comparison of corticotropin-releasing factor, dexamethasone, and
temozolomide: Treatment efficacy and toxicity in U87 and C6
intracranial gliomas. Clin Cancer Res. 17:3282–3292. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zeng Y, Yao X, Chen L, Yan Z, Liu J, Zhang
Y, Feng T, Wu J and Liu X: Sphingosine-1-phosphate induced
epithelial-mesenchymal transition of hepatocellular carcinoma via
an MMP-7/syndecan-1/TGF-β autocrine loop. Oncotarget.
7:63324–63337. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zeng YE, Yao XH, Yan ZP, Liu JX and Liu
XH: Potential signaling pathway involved in
sphingosine-1-phosphate-induced epithelial-mesenchymal transition
in cancer. Oncol Lett. 12:379–382. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lin YM, Jan HJ, Lee CC, Tao HY, Shih YL,
Wei HW and Lee HM: Dexamethasone reduced invasiveness of human
malignant glioblastoma cells through a MAPK phosphatase-1 (MKP-1)
dependent mechanism. Eur J Pharmacol. 593:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bauman GS, MacDonald W, Moore E, Ramsey
DA, Fisher BJ, Amberger VR and Del Maestro RM: Effects of radiation
on a model of malignant glioma invasion. J Neurooncol. 44:223–231.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Marcus HJ, Carpenter KL, Price SJ and
Hutchinson PJ: In vivo assessment of high-grade glioma biochemistry
using microdialysis: A study of energy-related molecules, growth
factors and cytokines. J Neurooncol. 97:11–23. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Piette C, Deprez M, Roger T, Noël A,
Foidart JM and Munaut C: The dexamethasone-induced inhibition of
proliferation, migration, and invasion in glioma cell lines is
antagonized by macrophage migration inhibitory factor (MIF) and can
be enhanced by specific MIF inhibitors. J Biol Chem.
284:32483–32492. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang D and Owler BK: Expression of AQP1
and AQP4 in paediatric brain tumours. J Clin Neurosci. 18:122–127.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zeng Y, Shen Y, Huang XL, Liu XJ and Liu
XH: Roles of mechanical force and CXCR1/CXCR2 in
shear-stress-induced endothelial cell migration. Eur Biophys J.
41:13–25. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Saadoun S, Papadopoulos MC, Hara-Chikuma M
and Verkman AS: Impairment of angiogenesis and cell migration by
targeted aquaporin-1 gene disruption. Nature. 434:786–792. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Noell S, Fallier-Becker P, Mack AF,
Hoffmeister M, Beschorner R and Ritz R: Water channels aquaporin 4
and −1 expression in subependymoma depends on the localization of
the tumors. PLoS One. 10:e01313672015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
McCoy E and Sontheimer H: Expression and
function of water channels (aquaporins) in migrating malignant
astrocytes. Glia. 55:1034–1043. 2007. View Article : Google Scholar : PubMed/NCBI
|