Introduction
Epstein-Barr virus (EBV) belongs to the γ-herpes
virus subfamily, which was identified and isolated from Burkitt's
lymphoma cells. The virus is classified as an oncogenic virus
associated with malignant tumors including nasopharyngeal carcinoma
(NPC), gastric cancer (GC), Burkitt's lymphoma and Hodgkin's
lymphoma (1,2). Therefore, the presence of EBV in GC is
occurs in 10% of case worldwide (3).
The EBV-positive GC is termed as EBV-associated GC (EBVaGC)
(4). EBV self-replicates in an
episomal form extrachromosomally and expresses latent genes
(5). According to the different host
cell types, the appearance of viral latent genes are varied and
categorized into three patterns, consisting of Latency I, II and
III (6). EBVaGC belongs to the
Latency I pattern, with latent gene products including EBV-encoded
small RNAs (EBERs), EBV-determined nuclear antigen I
(EBNA1) and BamHI-A rightward transcripts (BARTs,
BARF0 and BARF1), not including LMP1 and
EBNA2, which are usually expressed in NPC. The tumorigenic
effect of BARF1 has been demonstrated in numerous types of
cells, which are associated with stimulating cell proliferation and
transformation (7,8). In addition, BARF1 can be detected
in almost all cases of EBVaGC, but not LMP1 (7). Therefore, BARF1 is the only viral
oncogene identified in EBVaGC at present and may have an important
role in the development of EBVaGC. Nevertheless, the exact
mechanism through which BARF1 induces GC remains
elusive.
BARF1 has also been reported to have
anti-apoptotic function. For instance, N-terminal domain of
BARF1 gene is able to activate anti-apoptotic protein B-cell
lymphoma-2 (Bcl-2) expression in rodent fibroblasts (9). Bcl-2 activation has also been observed
in EBV-negative human B lymphoma cell line Akata transfected by
BARF1 (10). These findings
indicate that BARF1 can resist apoptosis by activating Bcl-2
expression. Transcriptional activator protein-1 (AP-1) is a
heterodimer complex mainly composed of c-Fos and c-Jun, wherein
c-Jun is the most important transcription factor. As an oncogenic
protein, c-Jun has a key role in the regulation of cell
proliferation, differentiation, invasion and apoptosis. Wang et
al (3) analyzed and confirmed
several transcription-associated genes including c-jun and
c-fos were upregulated in the BARF1 transfectants.
The expression of the anti-apoptotic protein Bcl-2 was upregulated,
while the expression of pro-apoptotic caspases and Bax protein was
reduced.
In multicellular organisms, apoptosis is one of the
most important forms of cell death, and usually presents disorders
in human tumors (11). The abnormal
activation or inhibition of signaling pathways is an important
cause of apoptosis dysregulation, and the mitogen-activated protein
kinase (MAPK) signaling pathway is one such pathway. When
stimulated by various extracellular signals, c-Jun N-terminal
kinase (JNK) is fully activated through phosphorylation of Thr183
and Tyr185, which can be mediated by the upstream MAPK kinase
kinase 4 or MAPK kinase 7 and then has enzyme catalytic activity
(12). Activation of JNK can enable
the transcription factor c-Jun phosphorylation in ser63 and ser73,
which improves the transcriptional activity. Additionally,
activated c-Jun involved in the formation of AP-1 proteins and then
to regulate a variety of proteins expression into the nucleus while
formative AP-1 proteins are also capable of binding c-Jun promoter
site itself to form positive feedback. Excessive activation of the
extracellular regulated protein kinase 1/2 (ERK1/2) signaling
pathway has also been found in a variety of tumors, and is closely
associated with the development of a variety of tumors. Regulation
of apoptosis proteins is one of the main mechanisms that
ERK1/2-MAPK act to resist apoptosis (13). In addition, another important member
of MAPK, p38, which can also be activated in the cytoplasm or
transfer to the nucleus to further regulate downstream substrates,
such as the protein kinases Prak, MSK1/2 and transcription factor
p53, ATF-2 and AP-1 to control cell proliferation, apoptosis and
metastasis (14,15).
Overall, the present study investigates whether
BARF1 is capable of regulating bcl-2 expression
through the JNK-, p38- and ERK-MAPK/c-Jun signaling pathway in
gastric carcinoma cells.
Materials and methods
Cell culture and treatment
The immortalized normal human embryo gastric
epithelial GES1 cell line and EBV-negative human gastric carcinoma
MKN28, which has been reported to be contaminated with the
moderately-differentiated MKN74 gastric carcinoma cell line
(16), SGC7901 and BGC823 cell lines
were purchased from the Chinese Academy of Sciences Cell Bank
(Shanghai, China). The pSG5 empty vector and pSG5-BARF1
stable transfectants, GES1-pSG5 (GES-SG), MKN28-pSG5 (MKN-SG),
SGC7901-pSG5 (SGC-SG), BGC823-pSG5 (BGC-SG), and GES1-BARF1
(GES-BARF1), MKN28-BARF1 (MKN-BARF1),
SGC7901-BARF1 (SGC-BARF1), BGC823-BARF1
(BGC-BARF1) (three cell clones of each), were constructed
and preserved by the Key Laboratory of Laboratory Medicine of
Jiangsu Province, School of Medicine (Jiangsu University,
Zhenjiang, China). It was then identified by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) that
the expression of BARF1 was normal. Total RNA was extracted
from cells using TRIzol (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and reverse transcription was performed
using the Prime Script RT kit (Takara Bio, Inc., Otsu, Japan)
according to the manufacturer's protocol. The levels of target mRNA
in cells were analyzed by qPCR using SYBR Green I dye (Takara Bio,
Inc.) detection. The PCR reactions were run at 95°C for 5 min
followed by 30 cycles of 95°C for 30 sec, 55°C for 30 sec and
finally 72°C for 5 min.
Cells were maintained in Dulbecco's modified Eagle's
medium (Invitrogen; Thermo Fisher Scientific, Inc.) growth medium
plus 10% fetal bovine serum (Zhejiang Tianhang Biotechnology Co.,
Ltd., Zhejiang, China), 100 U/ml penicillin and 100 µg/ml
streptomycin at 37°C in a humidified 5% CO2 atmosphere.
To study the effect of the JNK1/2/3, p38, and ERK1/2-MAPK/c-Jun
signaling axes on the expression and phosphorylation of Bcl-2 and
Bcl-xL, the cells were separately treated with 25 µmol/l specific
inhibitors of the above three signaling pathways, anthrapyrazolone
(SP600125; targeted at the JNK1/2/3 pathway),
4-(4-fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4-pyridyl)-1H-imidazole
(SB203580; targeted at the p38 pathways) and
1,4-diamino-2,3-dicyano-1,4-bis (o-aminophenylmercapto) butadiene
(U0126; targeted at the ERK1/2/MAPK/c-JUN pathway) (Beyotime
Institute of Biotechnology, Haimen, China) at 37°C for 8 h, and the
same concentration of dimethyl sulfoxide (DMSO) (Sinopharm Chemical
Reagent Co., Ltd., Shanghai, China) was used as a control.
Western blot analysis
Cells were cultured to a density of ~75%, and the
inhibition tests were added with 50 mol/l SP600125, SB203580, U0126
or DMSO to culture for 8 h. Cells were lysed for total protein
extraction in radioimmunoprecipitation assay buffer (50 mM
Tris-HCl, pH 7.4; 150 mM NaCl; 1% NP-40; 0.25% Na-deoxycholate; 1
mM EDTA; 1 mM NaF) together with a protease inhibitor cocktail
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Cell lysate was
collected, transferred into a 1.5-ml Eppendorf tube and clarified
by centrifugation at 14,000 × g for 5 min at 4°C. The supernatant
was collected and the protein concentration was determined using a
BCA protein assay kit (Beyotime Institute of Biotechnology)
according to the manufacturer's protocols.
Proteins (20 µg/lane) were separated by 10% SDS-PAGE
and transferred to polyvinylidene difluoride membranes (Merck
KGaA). After blocking with 5% skim milk in 0.01 mol/l PBS with
Tween-20 (0.05%) (PBST) for 1 h at room temperature, each membrane
was incubated with the following primary antibodies in 5% skim milk
overnight at 4°C in a humidified chamber: Rabbit anti-c-Jun
(9165T), anti-phosphorylated (p-) c-Jun (3270T), anti-ERK1/2
(4695T) and anti-p-ERK1/2 (4370T) (dilution, 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA); anti-JNK1/2/3
(BS9939M), anti-p-JNK1/2/3 (BS4322), anti-p38 (BS9851M), anti-p-p38
(BS4635) (dilution, 1:1,000); anti-Bcl-2 (BS70205); and anti-Bcl-xL
(BS1032) (dilution, 1:2,000; Bioworld Technology, Shanghai, China).
The membranes were then incubated with horseradish peroxidase
(HRP)-linked goat anti-rabbit IgG (BA1054; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) for 1 h at room temperature
subsequent to washing with PBST. The membranes were stripped and
re-probed with a mouse anti-β-actin antibody (BM0627; dilution,
1:3,000; Wuhan Boster Biological Technology, Ltd.) at 4°C overnight
and then incubated with an HRP-linked goat anti-mouse
immunoglobulin G antibody (BA1050; dilution, 1:3,000; Wuhan Boster
Biological Technology, Ltd.) at room temperature for 1 h.
Immunoreactive proteins were detected by using an enhanced
chemiluminescence system (GE Healthcare ImageQuant; GE Healthcare,
Chicago, IL, USA). Relative protein expression levels were
quantified by comparing the density of each band relative to the
β-actin reference gene using Quantity One analysis software 4.62
(Bio-Rad Laboratories, Inc., Waltham, MA, USA). All experiments
were independently performed three times at least.
Statistical analysis
Values were presented as the mean ± standard
deviation. Student's t-test was performed to evaluate differences
between the GEC-BARF1 and GEC-SG groups. One-way analysis of
variance was used to assess differences among the groups. Tukey's
honest significant difference was used for post hoc analysis of the
means. SPSS (version 19.0) software (IBM Corp., Armonk, NY, USA)
was used for statistical analyses. Linear regression and linear
correlation analysis were applied to trend analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
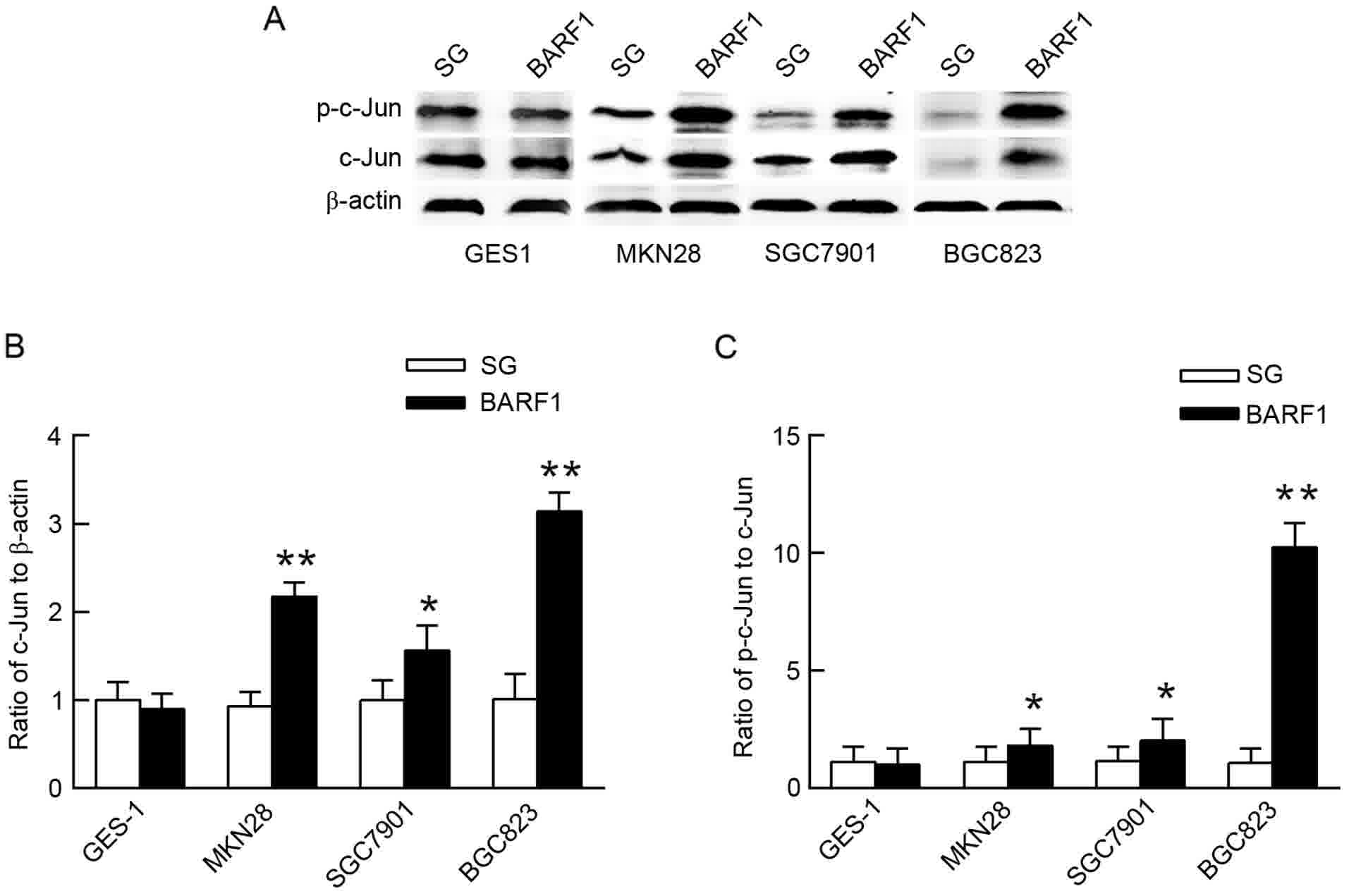
Expression and phosphorylation levels
of the c-Jun protein are increased in GCC-BARF1 cells
In order to analyze the activity of c-Jun in
GEC-BARF1, gastric epithelial cells were transfected with
pSG5 empty vector (GEC-SG) as the control. The expression and
phosphorylation levels of the c-Jun protein were then analyzed by
western blot analysis in GEC-BARF1 cells. The results showed
that the c-Jun protein expression and phosphorylation levels were
significantly increased in GCC-BARF1 cells compared with
GCC-SG cells (MKN-BARF1 vs. MKN-SG, P<0.01 and P<0.05
for c-Jun expression and phosphorylation levels, respectively;
SGC-BARF1 vs. SGC-SG, P<0.01 and P<0.05 for c-Jun
expression and phosphorylation levels, respectively,
BGC-BARF1 vs. BGC-SG, P<0.05 and P<0.01 for c-Jun
expression and phosphorylation levels, respectively), but not
significantly changed in GES-BARF1 cells compared with
GES-SG cells (P>0.05). In addition, the c-Jun protein expression
and phosphorylation levels in the GCC-BARF1 group were
increased compared with the GES-BARF1 group
(MKN-BARF1 vs. GES-BARF1, P<0.05; SGC-BARF1
vs. GES-BARF1, P<0.05; BGC-BARF1 vs.
GES-BARF1, P<0.01; Fig.
1).
JNK1/2/3, p38 and ERK1/2-MAPK/c-Jun
cascade pathways are activated in GCC-BARF1 cells
To investigate whether the activation of c-Jun was
associated with the JNK-MAPK signaling pathway, western blot
analysis was used to analyze the changes in the phosphorylation
level of JNK1/2/3 proteins in the GEC-BARF1 group with
GEC-SG as controls. The results showed that there was no
significant difference in the phosphorylation level of JNK1/2/3
protein between GES-BARF1 cells and GES-SG cells
(P>0.05), while the phosphorylation level of JNK1/2/3 proteins
was significantly increased in GCC-BARF1 cells compared with
GCC-SG cells (MKN-BARF1 vs. MKN-SG, P<0.01;
SGC-BARF1 vs. SGC-SG, P<0.05; BGC-BARF1 vs.
BGC-SG, P<0.05). Additionally, the phosphorylation levels of
JNK1/2/3 protein in the GCC-BARF1 group was increased
compared with that in the GES-BARF1 group (MKN-BARF1
vs. GES-BARF1, P<0.01; SGC-BARF1 vs.
GES-BARF1, P<0.01; BGC-BARF1 vs. GES-BARF1,
P<0.05), and the phosphorylation level of JNK1/2/3 protein in
MKN-BARF1 had the highest increase in the GCC-BARF1
group (P<0.01; Fig. 2A-a and b).
The expression and phosphorylation levels of c-Jun protein were
significantly decreased in SGC-BARF1 cells subsequent to
treatment with the specific inhibitor of JNK1/2/3 SP600125
(P<0.05; Fig. 2A-c and d).
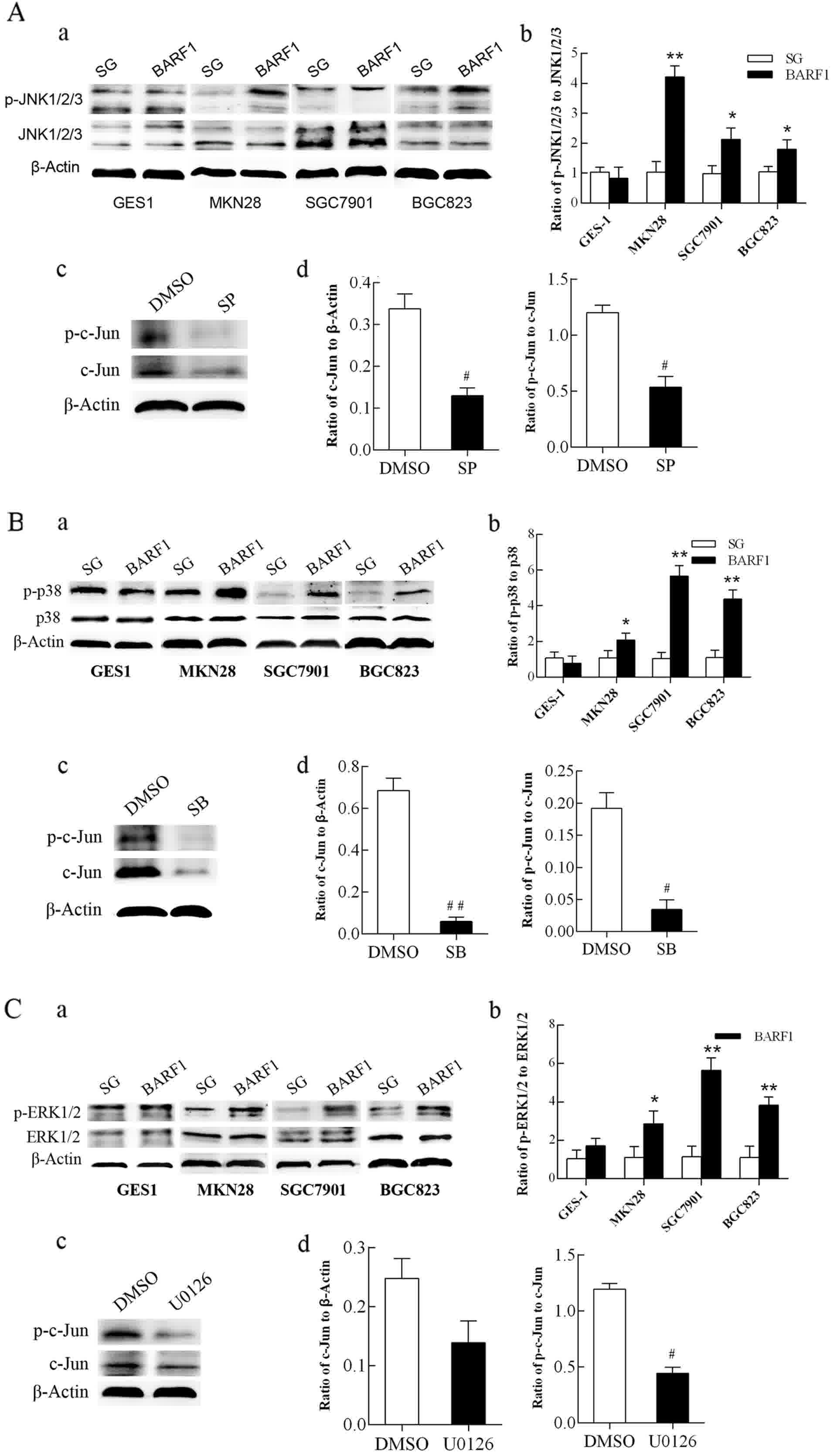 | Figure 2.Analysis of the activity of JNK1/2/3,
p38 and ERK1/2-MAPK/c-Jun signaling pathways in GEC-BARF1
cells. (A-a, B-a and C-a) Western blotting and (A-b, B-b and C-b)
quantitative analysis of JNK1/2/3, p38 and ERK1/2 protein
phosphorylation, respectively, in GEC cells. (A-c, B-c and C-c)
Western blotting and (A-d, B-d and C-d) quantitative analysis of
c-Jun expression and phosphorylation in SGC-BARF1 treated
with SP600125, SB203580 and U0126 compared with DMSO, respectively.
Data were expressed as the mean ± standard deviation from three
independent experiments. *P<0.05, **P<0.01 vs. GEC-SG.
#P<0.05, ##P<0.01 vs. DMSO control.
DMSO, dimethyl sulfoxide. |
Western blotting showed that the phosphorylation
level of the p38 protein was not significantly different
(P>0.05) between GES-BARF1 and GES-SG groups, while there
was a significant increase in GCC-BARF1 compared with GCC-SG
(MKN-BARF1 vs. MKN-SG, P<0.05; SGC-BARF1 vs.
SGC-SG, P<0.01; BGC-BARF1 vs. BGC-SG, P<0.01). The
phosphorylation level of the p38 protein in the GCC-BARF1
group was significantly increased compared with the
GES-BARF1 group (full P<0.01), and the level in the
SGC-BARF1 group was significantly increased compared with
that in the MKN-BARF1 group (P<0.01) (Fig. 2B-a and b). Subsequent to treatment
with the specific inhibitor of p38, SB203580, the expression and
phosphorylation levels of c-Jun protein were significantly
decreased (P<0.01 and P<0.05, respectively; Fig. 2B-c and d).
Western blot analysis showed that the
phosphorylation level of ERK1/2 was not significantly different
between GES-BARF1 and GES-SG groups (P>0.05), while a
significant increase occurred in the GCC-BARF1 group
(MKN-BARF1 vs. MKN-SG, P<0.05; SGC-BARF1 vs.
SGC-SG, P<0.01; BGC-BARF1 vs. BGC-SG, P<0.01).
Additionally, the phosphorylation level of ERK1/2 in the
GCC-BARF1 group was increased compared with that in the
GES-BARF1 group except MKN-BARF1 (MKN-BARF1
vs. GES-BARF1, P>0.05; SGC-BARF1 vs.
GES-BARF1, P<0.01; BGC-BARF1 vs. GES-BARF1,
P<0.05), and SGC-BARF1 cells had the highest
phosphorylation level in the GEC-BARF1 group (P<0.05;
Fig. 2C-a and b). Subsequent to
treatment with the specific inhibitor of ERK1/2, U0126, the
phosphorylation level of the c-Jun protein in SGC-BARF1 was
significantly decreased (P<0.05), while the expression of c-Jun
was not evidently decreased (P>0.05; Fig. 2C-c and d).
JNK1/2/3, p38 and ERK1/2-MAPK/c-Jun
signaling pathways mediated upregulation of the anti-apoptotic
proteins Bcl-2 and Bcl-xL
To further investigate whether the role of these
pathways mediated BARF1-induced apoptosis inhibition in GC
cells, GEC-SG was used as a control and the expression of
anti-apoptotic proteins Bcl-2 and Bcl-xL was analyzed in
GEC-BARF1 using immunoblotting. The results showed that
there were no significant difference between GES-BARF1 and
GES-SG (P>0.05), but the expression levels of Bcl-2 and Bcl-xL
proteins in the GCC-BARF1 group were significantly increased
compared with the GCC-SG group (MKN-BARF1 vs. MKN-SG,
P<0.05; SGC-BARF1 vs. SGC-SG, P<0.01; BGC-BARF1
vs. BGC-SG, P<0.01), and the GES-BARF1 groups
(MKN-BARF1 vs. GES-BARF1; SGC-BARF1 vs.
GES-BARF1, BGC-BARF1 vs. GES-BARF1; P<0.05,
P<0.01, P<0.01, and all P<0.01; Fig. 3A).
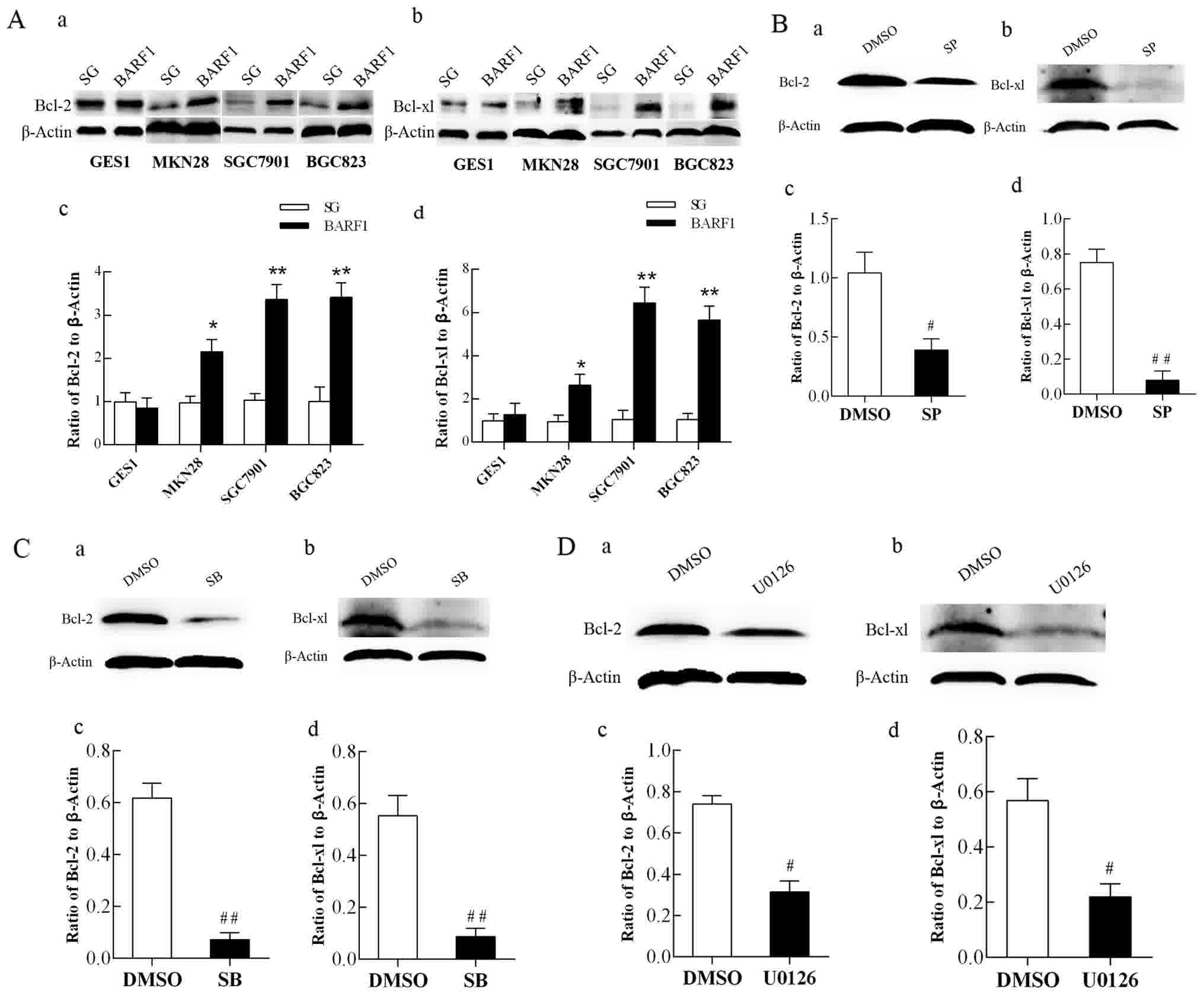 | Figure 3.Analysis of Bcl-2 and Bcl-xL protein
expression in GEC-BARF1 cells. Western blotting of (A-a)
Bcl-2 and (A-b) Bcl-xL expression, and quantitative analysis of
(A-c) Bcl-2 and (A-d) Bcl-xL expression in GEC-BARF1 cells
compared with GEC-SG cells. (B-a, B-b, C-a, C-b, D-a and D-b)
Western blotting and quantitative analysis of (B-c, C-c and D-c)
Bcl-2 and (B-d, C-d and D-d) Bcl-xL protein expression levels in
SGC-BARF1 treated with SP600125, SB203580 and U0126 compared
with DMSO, respectively. Data were expressed as the mean ± standard
deviation from three independent experiments. *P<0.05,
**P<0.01 vs. GEC-SG. #P<0.05,
##P<0.01 vs. DMSO control. DMSO, dimethyl sulfoxide;
Bcl, B-cell lymphoma. |
To further understand the association between the
anti-apoptotic proteins Bcl-2 and Bcl-xL, and the JNK1/2/3, p38 and
ERK1/2-MAPK/c-Jun signaling pathways, western blotting was
performed to analyze the protein expression levels of Bcl-2 and
Bcl-xL in SGC-BARF1 subsequent to treatment with SP600125,
SB203580 and U0126, respectively. The results then showed that the
protein expression levels of Bcl-2 and Bcl-xL were significantly
decreased subsequent to treatment with 3 inhibitors
(SGC-BARF1-SP vs. SGC-BARF1-DMSO P<0.05 and
P<0.01 for Bcl-2 and Bcl-xL, respectively; SGC-BARF1-SB
vs. SGC-DMSO, P<0.01 for Bcl-2 and Bcl-xL; SGC-BARF1-U
vs. SGC-BARF1-DMSO, P<0.05 for Bcl-2 and Bcl-xL; Fig. 3B-D).
Discussion
To the best of our knowledge, the present study
revealed for the first time that JNK1/2/3, p38 and
ERK1/2-MAPK/c-Jun cascade signaling pathways participate to mediate
the upregulation of anti-apoptotic protein Bcl-2 and Bcl-xL induced
by EBV-encoded BARF1 in 3 gastric carcinoma cell lines in
comparison with the normal gastric epithelium GES1 cells. It has
been confirmed in a number of studies that BARF1 can inhibit
cell apoptosis by activating anti-apoptotic protein Bcl-2
expression (3,9,10).
However, the precise mechanism by which BARF1 regulates
Bcl-2 expression remains unclear. In order to analyze the
mechanism, the immortalized normal human embryo gastric epithelial
GES1 cell line, and three gastric carcinoma cell lines in different
differentiation stages, consisting of well, moderately and
poorly-differentiated cells (MKN28, SGC7901 and BGC823,
respectively), were used in the present study.
AP-1 is a complex composed of homologous or
heterologous dimers of Jun and Fos family members. The AP-1 protein
activates target gene transcription activity by binding to its DNA,
and then contributes to a series of pathophysiological processes,
including cell proliferation, apoptosis and differentiation
(17). c-Jun is the fundamental
member of the AP-1 family, and is also the most powerful
transcriptional activator. As an oncoprotein, the c-Jun/AP-1
protein shows overexpression in breast cancer, ovarian cancer,
endometrial cancer, colon cancer and lung cancer (17–19), and
can also promote skin cancer and liver cancer development (20,21). For
example, in terms of cell apoptosis, Eferl et al (22) showed that c-Jun-deficient mouse
embryos can promote the apoptosis of fetal rat hepatocytes.
Additionally, Eferl et al (20) demonstrated that c-Jun/AP-1 resisted
TNF-α-induced liver cell apoptosis by inhibiting the expression of
the apoptotic protein p53. In the present study, it was found that
BARF1 promotes the phosphorylation of the c-Jun protein in
gastric carcinoma cells. The phosphorylation of c-Jun first
triggers activation of AP-1. In turn, activated c-Jun/AP-1
interacts with the c-Jun promoter to promote the synthesis of
c-Jun/AP-1 to form the positive feedback regulation of the Jun
gene, which is important in signal transduction into long-term
effects on cellular gene expression (16). Previously, it has been found that
several transcription associated genes, including c-jun and
c-fos, were upregulated in the BARF1 transfectant
(BGC823-BARF1) (3). Similar to
the aforementioned research, the present study found that the
increase of c-Jun at the protein level by western blotting. As a
transcription factor, c-Jun may be adjusted both at the protein
expression and the phosphorylation levels, and improvement of any
level is able to promote its downstream target gene expression.
Activity of c-Jun is regulated by a variety of
upstream factors. Among these factors, JNK is a member of the MAPK
family, and is a highly-conserved serine/threonine protein kinase.
In mammals, JNK and p38-MAPK are described as stress activated
protein kinases (SAPKs) that promote cell death (23,24) and
mediate inflammatory response (25,26). The
JNK protein kinase is encoded by 3 homologous genes, consisting of
JNK1, JNK2 and JNK3. The JNK/SAPK signaling pathway has two roles
in the regulation of cell apoptosis (27). For example, activation of the
JNK/c-Jun signaling pathway can promote apoptosis of neural cells.
However, occasionally, c-Jun does not participate in the
JNK-mediated apoptosis effect. Hilberg et al (28) showed that in Jnk1−/−
Jnk2-/− mice, the neuronal cells exhibited impaired
apoptosis, while nerve cells did not appear to have impaired
apoptosis in c-Jun-/-mice. This indicates that JNK1 and JNK2
induced the apoptosis of neuronal cells is not always mediated by
c-Jun, another factor may be involved in this process.
Subsequently, Raitano et al (29) found that the bcr-abl leukemia
oncogene activates the JNK/SAPK pathway of hematopoietic stem cells
and then increase the capacity of cell transformation by activating
downstream c-Jun protein. When c-Jun or JNK is repressed, the
transformation function of bcr-abl is also inhibited. Hess
et al (30) showed that
bcr-abl can further up-regulate bcl-2 expression by
activating the JNK/SAPK pathway and result in enhanced
transformation ability of B lymphoblastoid cells accompany with
diminished apoptosis. Consequently, it can be speculated that
bcr-abl-induced activation of JNK/c-Jun may promote
bcl-2 expression and mediate cell transformation ability,
accompanied by anti-apoptosis function.
In the present study, the phosphorylation level of
JNK1/2/3, and c-Jun expression and phosphorylation levels were
found to be increased in the GCC-BARF1 group, and
significantly decreased in the SGC-BARF1 group subsequent to
treatment with the specific inhibitor of JNK1/2/3, suggesting that
BARF1 promoted c-Jun activity at both protein expression and
phosphorylation levels by activating JNK1/2/3-MAPK signaling, and
consequentially regulated the expression of downstream target
genes.
As a regulator of cell death, p38-MAPK plays a dual
role in mediating cell death or cell survival, and the function of
p38-MAPK is determined by the type of stimulus or a specific cell
type. Activated p38 can act on a variety of transcription factors,
such as STAT1, NF-κB and AP-1, and then regulates cell
proliferation, apoptosis, metastasis and differentiation. p38-MAPK
can enhance the anti-apoptosis ability of cells that are mostly
associated with drug resistance. Previously, Milone et al
(31) described that the activation
of the p38-MAPK signaling pathway in prostate cancer cells leads to
resistance of zoledronic acid-induced apoptosis and overexpression
of the anti-apoptotic protein Bcl-2. It was also found that the
p38-MAPK signaling pathway augments the expressions of associated
factors, such as MMP-2, VEGF and IL-2, and cause
epithelial-to-mesenchymal transition. It shows that p38-MAPK signal
pathway may promote drug resistance in numerous aspects, such as
transfer, proliferation and anti-apoptosis. Consistent with the
aforementioned studies, in the present study, it was found that
when the p38-MAPK signaling pathway is specifically suppressed by
SB203580, Bcl-2 and Bcl-xL protein expression, as well as the
expression and phosphorylation levels of c-Jun, was significantly
decreased, indicating that BARF1 performs its anti-apoptotic
effect by activating the p38-MAPK/c-Jun pathway in gastric
carcinoma cells.
Zelivianski et al (32) found that the activated ERK pathway
mediates an anti-apoptotic response to the treatment with certain
chemotherapeutic agents. Nishinaka et al (33) found that phorbol ester,
12-O-tetradecanoyl phorbol 13-acetate can induce c-Jun expression
in an ERK-dependent manner in human lung cancer cells. A previous
study has indicated that BARF1 may upregulate Bcl-2
expression through the ERK1/2-MAPK/c-Jun signaling pathway in NPC
cells. In the present study, it was observed that ERK1/2 was
activated by BARF1, and c-Jun protein expression and
phosphorylation levels were significantly increased in
GCC-BARF1 cells. After SGC-BARF1 cells were treated
with U0126, the phosphorylation level of the c-Jun protein was
markedly decreased, although there was no significant alteration in
protein expression, indicating that ERK1/2 regulated c-Jun
activation mainly at the phosphorylation level. Accordingly, this
indicated that the ERK signaling pathway may be involved in
BARF1-induced apoptosis inhibition in gastric carcinoma
cells.
c-Jun/AP-1 regulates cell apoptosis by a variety of
mechanisms (34). Li et al
(35) has shown that the c-Jun
protein can enhance the transcription level of bcl-2 by
binding to the bcl-2 promoter in human endometrial glandular
cells. In agreement with a previous study (3), it was observed that the expression of
Bcl-2 was significantly increased in GCC-BARF1 compared with
GCC-SG, and there was a gradual increase in expression that
was associated with the malignant degree of cells and showed a
certain association with c-Jun activity, but this was not
statistically significant. Following treatment with three specific
inhibitors of the MAPK signaling pathway, c-Jun protein and
phosphorylation levels, as well as Bcl-2 and Bcl-xL protein
expression levels, were significantly decreased, indicating
JNK1/2/3, p38 and ERK1/2-MAPK/c-Jun signaling pathway were involved
in the regulation of the Bcl-2 and Bcl-xL proteins in gastric
carcinoma cells.
Furthermore, the phosphorylation levels of p38 and
ERK1/2 proteins in SGC-BARF1 and of JNK1/2/3 proteins in
MKN-BARF1 was significantly increased compared with that in
the other two cells in the GCC-BARF1 group, while the
expression levels of c-Jun, Bcl-2 and Bcl-xL and phosphorylation
levels of c-Jun, JNK1/2/3, p38 and ERK1/2 were not significantly
different in GES-BARF1 cells compared with GES-SG cells,
suggesting that the p38 and ERK1/2-MAPK/c-Jun signaling pathways
may be dominant in the progressive stage, and JNK1/2/3-MAPK/c-Jun
may mainly occur in the early stage of EBVaGC development. However,
the MAPK/c-Jun signaling cascade is unlikely to play an important
role in the initial stage of EBVaGC.
Previously, it been reported that the MKN28 cell
line may be cross-contaminated with MKN74 cells which are
moderately-differentiated gastric carcinoma cell lines (16). Even if MKN74 cross-contaminated MKN28
cell line has been used in the present study, it is unlikely to
affect the conclusions derived from the comparisons between
BARF1 and SG-transfected cells and between the GCC and GES1
groups. However, the reliability of the conclusions about the
functions of the 3 MAPK pathways in the different phases of EBVaGC
evolution may have been affected, by having been derived from the
comparison among 3 differently differentiated gastric carcinoma
cell lines.
Chen et al (36) constructed a transfectant of
B-cell-specific Moloney murine leukemia virus integration site 1
(Bmi-1) stably expressed (GES-Bmi-1) and find that overexpression
of Bmi-1 can promote cell proliferation and inhibit apoptosis, and
also enhance cell migration and invasion ability by upregulating
vimentin and fibronectin via the PI3K/Akt signaling pathway.
Accordingly, whether PI3K/Akt signaling pathway is involved in
BARF1 mediated regulation on apoptosis of GES is worthy of
further study. Furthermore, numerous issues required further
exploration. Signaling networks are complicated, and whether other
signaling pathways are involved in the activation of c-Jun to
participate in the anti-apoptotic effect of BARF1 in gastric
carcinoma requires investigation. Apart from the anti-apoptotic
effect, whether c-Jun is also involved in other biological
behavior, including cell proliferation, migration and invasion also
requires additional investigation. These issues will be further
explored in a following study.
Acknowledgements
The present study was supported by the Professional
Research Foundation for Advanced Talents of Jiangsu University
(grant no. 06JDG011) to Tianji Zhou and Student's Scientific
Research of Jiangsu University to Yuqiong Zhang.
References
|
1
|
Seto E, Yang L, Middeldorp J, Sheen TS,
Chen JY, Fukayama M, Eizuru Y, Ooka T and Takada K: Epstein-Barr
virus (EBV)-encoded BARF1 gene is expressed in nasopharyngeal
carcinoma and EBV-associated gastric carcinoma tissues in the
absence of lytic gene expression. J Med Virol. 76:82–88. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Y, Ohyashiki JH, Takaku T, Shimizu N
and Ohyashiki K: Transcriptional profiling of Epstein-Barr virus
(EBV) genes and host cellular genes in nasal NK/T-cell lymphoma and
chronic active EBV infection. Br J Cancer. 94:599–608. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Q, Tsao SW, Ooka T, Nicholls JM,
Cheung HW, Fu S, Wong YC and Wang X: Anti-apoptotic role of BARF1
in gastric cancer cells. Cancer Lett. 238:90–103. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takada K: Epstein-Barr virus and gastric
carcinoma. Mol Pathol. 53:255–261. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Klein E: The complexity of the
Epstein-Barr virus infection in humans. Pathol Oncol Res. 4:3–7.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rowe M, Lear AL, Croom-Carter D, Davies AH
and Rickinson AB: Three pathways of Epstein-Barr virus gene
activation from EBNA1-positive latency in B lymphocytes. J Virol.
66:122–131. 1992.PubMed/NCBI
|
|
7
|
zur Hausen A, Brink AA, Craanen ME,
Middeldorp JM, Meijer CJ and van den Brule AJ: Unique transcription
pattern of Epstein-Barr virus (EBV) in EBV-carrying gastric
adenocarcinomas: Expression of the transforming BARF1 gene. Cancer
Res. 60:2745–2748. 2000.PubMed/NCBI
|
|
8
|
Wei MX, de Turenne-Tessier M, Decaussin G,
Benet G and Ooka T: Establishment of a monkey kidney epithelial
cell line with the BARF1 open reading frame from Epstein-Barr
virus. Oncogene. 14:3073–3081. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sheng W, Decaussin G, Sumner S and Ooka T:
N-terminal domain of BARF1 gene encoded by Epstein-Barr virus is
essential for malignant transformation of rodent fibroblasts and
activation of BCL-2. Oncogene. 20:1176–1185. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sheng W, Decaussin G, Ligout A, Takada K
and Ooka T: Malignant transformation of Epstein-Barr virus-negative
Akata cells by introduction of the BARF1 gene carried by
Epstein-Barr virus. J Virol. 77:3859–3865. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bai L and Wang S: Targeting apoptosis
pathways for new cancer therapeutics. Annu Rev Med. 65:139–155.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Plotnikov A, Zehorai E, Procaccia S and
Seger R: The MAPK cascades: Signaling components, nuclear roles and
mechanisms of nuclear translocation. Biochim Biophys Acta.
1813:1619–1633. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Balmanno K and Cook SJ: Tumour cell
survival signalling by the ERK1/2 pathway. Cell Death Differ.
16:368–377. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ohashi M, Fogg MH, Orlova N, Quink C and
Wang F: An Epstein-Barr virus encoded inhibitor of colony
stimulating factor-1 signaling is an important determinant for
acute and persistent EBV infection. PLoS Pathog. 8:e10030952012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Iyoda K, Sasaki Y, Horimoto M, Toyama T,
Yakushijin T, Sakakibara M, Takehara T, Fujimoto J, Hori M, Wands
JR and Hayashi N: Involvement of the p38 mitogen-activated protein
kinase cascade in hepatocellular carcinoma. Cancer. 97:3017–3026.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Angel P, Hattori K, Smeal T and Karin M:
The jun proto-oncogene is positively autoregulated by its product,
Jun/AP-1. Cell. 55:875–885. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng Q and Xia Y: c-Jun, at the crossroad
of the signaling network. Protein Cell. 2:889–898. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Neyns B, Katesuwanasing, Vermeij J,
Bourgain C, Vandamme B, Amfo K, Lissens W, DeSutter P,
Hooghe-Peters E and DeGrève J: Expression of the jun family of
genes in human ovarian cancer and normal ovarian surface
epithelium. Oncogene. 12:1247–1257. 1996.PubMed/NCBI
|
|
19
|
Vleugel MM, Greijer AE, Bos R, van der
Wall E and van Diest PJ: c-Jun activation is associated with
proliferation and angiogenesis in invasive breast cancer. Hum
Pathol. 37:668–674. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eferl R, Ricci R, Kenner L, Zenz R, David
JP, Rath M and Wagner EF: Liver tumor development. c-Jun
antagonizes the proapoptotic activity of p53. Cell. 112:181–192.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Young MR, Li JJ, Rincón M, Flavell RA,
Sathyanarayana BK, Hunziker R and Colburn N: Transgenic mice
demonstrate AP-1 (activator protein-1) transactivation is required
for tumor promotion. Proc Natl Acad Sci USA. 96:9827–9832. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eferl R, Sibilia M, Hilberg F,
Fuchsbichler A, Kufferath I, Guertl B, Zenz R, Wagner EF and
Zatloukal K: Functions of c-Jun in liver and heart development. J
Cell Biol. 145:1049–1061. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harper SJ and LoGrasso P: Signalling for
survival and death in neurones: The role of stress-activated
kinases, JNK and p38. Cell Signal. 13:299–310. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rawal N, Parish C, Castelo-Branco G and
Arenas E: Inhibition of JNK increases survival of transplanted
dopamine neurons in Parkinsonian rats. Cell Death Differ.
14:381–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Adhikary G, Sun Y and Pearlman E: C-Jun
NH2 terminal kinase (JNK) is an essential mediator of Toll-like
receptor 2-induced corneal inflammation. J Leukoc Biol. 83:991–997.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ruano D, Revilla E, Gavilán MP, Vizuete
ML, Pintado C, Vitorica J and Castaño A: Role of p38 and inducible
nitric oxide synthase in the in vivo dopaminergic cells'
degeneration induced by inflammatory processes after
lipopolysaccharide injection. Neuroscience. 140:1157–1168. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin A: Activation of the JNK signaling
pathway: Breaking the brake on apoptosis. Bioessays. 25:17–24.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hilberg F, Aguzzi A, Howells N and Wagner
EF: c-jun is essential for normal mouse development and
hepatogenesis. Nature. 365:179–181. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Raitano AB, Halpern JR, Hambuch TM and
Sawyers CL: The Bcr-Abl leukemia oncogene activates Jun kinase and
requires Jun for transformation. Proc Natl Acad Sci USA.
92:11746–11750. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hess P, Pihan G, Sawyers CL, Flavell RA
and Davis RJ: Survival signaling mediated by c-Jun NH (2)-terminal
kinase in transformed B lymphoblasts. Nat Genet. 32:201–205. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Milone MR, Pucci B, Bruzzese F, Carbone C,
Piro G, Costantini S, Capone F, Leone A, Di Gennaro E, Caraglia M
and Budillon A: Acquired resistance to zoledronic acid and the
parallel acquisition of an aggressive phenotype are mediated by
p38-MAP kinase activation in prostate cancer cells. Cell Death Dis.
4:e6412013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zelivianski S, Spellman M, Kellerman M,
Kakitelashvilli V, Zhou XW, Lugo E, Lee MS, Taylor R, Davis TL,
Hauke R and Lin MF: ERK inhibitor PD98059 enhances
docetaxel-induced apoptosis of androgen-independent human prostate
cancer cells. Int J Cancer. 107:478–485. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nishinaka T, Miura T, Sakou M, Hidaka C,
Sasaoka C, Okamura A, Okamoto A and Terada T: Down-regulation of
aldo-keto reductase AKR1B10 gene expression by a phorbol ester via
the ERK/c-Jun signaling pathway. Chem Biol Interact. 234:274–281.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shaulian E and Karin M: AP-1 as a
regulator of cell life and death. Nat Cell Biol. 4:E131–E136. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li ZL, Abe H, Ueki K, Kumagai K, Araki R
and Otsuki Y: Identification of c-Jun as bcl-2 transcription factor
in human uterine endometrium. J Histochem Cytochem. 51:1601–1609.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen Y, Lian G, Zhang Q, Zeng L, Qian C,
Chen S and Huang K: Overexpression of Bmi-1 induces the malignant
transformation of gastric epithelial cells in vitro. Oncol Res.
21:33–41. 2013. View Article : Google Scholar : PubMed/NCBI
|