Introduction
Breast cancer is a type of malignant tumor that
occurs in the epithelial tissue of the mammary gland, its incidence
and mortality rates are one of the highest among malignant tumors
in females worldwide (1). In China,
the incidence of breast cancer is increasing and occurs at younger
ages compared with previously (2).
The causal factors and mechanisms of breast cancer involve numerous
factors, the proliferation and apoptosis of breast cancer cells has
become a hotspot for correlative research (3,4).
Nonsteroidal anti-inflammatory drugs (NSAIDs) are a
chemically diverse drug type commonly used to treat inflammatory
conditions and pain. The long-term use of NSAIDs has been
investigated to reduce the risk of mortality from certain types of
cancer (5). In addition, sulindac is
widely used in clinical anti-infection medicine. Previously, a
large number of studies have demonstrated that sulindac sulfide can
effectively inhibit the proliferation and induce the apoptosis of
various cancer cells (6–8). Although its use for patients with
malignant disease has not been well investigated, numerous evidence
suggests that it has potent cancer chemopreventive efficacy:
Several publications have reported that sulindac sulfide can
inhibit the invasion of colorectal, gastric and pancreatic cancer
cells (9–12), but the mechanism remains unclear. In
the present study, the effects of sulindac sulfide on the
proliferation and apoptosis of human breast cancer cells MCF-7
treated with different concentrations, and durations were
investigated. In addition, its potential molecular mechanism was
explored. The current study may provide the experimental basis for
clinical rational administration of sulindac sulfide.
Materials and methods
Materials
The human breast cancer MCF-7 cell line was
purchased from the Type Culture Collection of the Chinese Academy
of Sciences (Shanghai, China). Sulindac sulfide with 98% purity was
obtained from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany), which
was dissolved in DMSO at 80 µmol/l and stored at −20°C. The cell
counting kit (CCK)-8 and Hoechst 33258 kits were obtained from
Beyotime Institute of Biotechnology (Shanghai, China). The annexin
V-FITC/propidium (PI) apoptosis kit was obtained from BD
Biosciences (Franklin Lakes, NJ, USA). Antibodies directed against
apoptosis regulator Bax (Bax; cat. no., 5023; dilution 1:1,000),
apoptosis regulator Bcl-2 (Bcl-2; cat. no., 2870; dilution 1:1,000)
and caspase-3 (cat. no., 9665; dilution 1:1,000) were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA). Secondary
rabbit (cat. no., DC20L; dilution 1:5,000), mouse (cat. no.,
074-1809; dilution 1:5,000) and actin antibodies were obtained from
Sigma-Aldrich (Merck KGaA). An automatic microplate reader was
obtained from Thermo Fisher Scientific, Inc. (Waltham, MA, USA). A
flow cytometer was purchase from (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Cell culture
Human breast cancer MCF-7 cells were cultured in
RPMI-1640 medium containing 10% fetal bovine serum (Invitrogen;
Thermo Fisher Scientific, Inc.), non-essential amino acids, 100
µg/ml streptomycin and 100 U/ml penicillin and maintained at 37°C
in a humidified incubator with 5% CO2. Cells were
collected in the growth phase for subsequent experiments.
Measurement of cell proliferation
Growth phase MCF-7 cells were seeded into 96-well
plates at a density of 1×105 cells/200 µl/well in
triplicates. Cells were incubated with different concentrations of
sulindac sulfide (20, 40 or 80 µmol/l) and the control group was
incubated in complete RPMI-1640 medium without sulindac sulfide.
Following incubation times of 24, 48 and 72 h, 20 µl CCK-8 agent
was added to each well and incubated in a humidified incubator at
37°C for 1 h. The optical density (OD) value was detected using an
automatic microplate reader at 450 nm. Three independent
experiments were performed. The ratio of proliferation was
calculated as follows: (1-OD/control OD) ×100%.
Cell cycle analysis
Analysis of the cell cycle was performed as
described previously (13). Growth
phase MCF-7 cells were seeded into 24-well plates at
1×105 cells/800 µl/well. These cells were incubated with
different concentrations of sulindac sulfide for 48 h, collected
and fixed with 70% ethanol overnight at 4°C. Prior to measuring,
cells were rinsed thoroughly with PBS liquid centrifugal cast
fixation fluid, and 1 ml 100 mg/l RNA enzyme was added to each tube
for cellular staining for 30 min at 37°C. Then, PI tag was added to
each tube and cells were incubated at room temperature in the dark
for 30 min, followed by flow cytometry. Three independent
experiments were performed. Experience data were analyzed using
FlowJo ModFit 3.2. (Tree Star, Inc., Ashland, OR, USA).
Hoechst 33258 staining
Growth phase MCF-7 cells were seeded into 24-well
plates at a density of 1×105 cells 200 µl/well in
triplicates. The cells incubated with different concentrations of
sulindac sulfide (20, 40 or 80 µmol/l) and the control group was
incubated in RPMI-1640 complete medium. Following the incubation
for 24, 48 or 72 h, 10 µl Hoechst 33258 was added to each sample.
Samples were then fixed with 4% paraformaldehyde for 25 min at room
temperature, washed twice with PBS with 0.1% of Triton X-100.
Images of samples of ~200 cell were captured using a fluorescent
microscope at magnification, ×400.
Apoptosis analysis
Apoptosis was measured using the Annexin V-FITC
apoptosis detection kit according to the manufacturer's protocol
(14). Growth phase MCF-7 cells were
seeded into 6-well plates. The cells were incubated with different
concentrations of sulindac sulfide for 72 h, collected, washed
twice in cold PBS, and incubated with 10 µl of Annexin V-FITC and
PI for 30 min. Samples were measured using flow cytometry. Three
independent experiments were performed. Experimental data were
analyzed using ModFit LT (version 3.2; Verity Software House, Inc.,
Topsham, ME, USA).
Western blot analysis
Analysis of western blotting was performed as
described previously (15). Cells
(2×106) were seeded into 6-well plates at a density of
2,000 µl/well and were treated with different concentrations of
sulindac sulfide or the control. Cells were washed with PBS, then
total protein was extracted using a Total protein extraction kit
(Keygentec, Inc., Nanjing, China). Proteins were quantified using a
BCA assay. Samples (50 µg) were resolved using 10% SDS-PAGE and
transferred onto polyvinylidene difluoride (PVDF) membranes.
Membranes were blocked in 5% milk for 1 h at room temperature, and
subsequently stored overnight at 4°C on a shaker with the desired
primary antibodies. Membranes were washed with TBS-Tween 20(T) and
incubated for 1 h with the secondary antibody at room temperature.
Following several washes with TBST, membranes were visualized using
the Electro-Chemi-Luminescence reagent (Keygentec, Inc.). Each PVDF
membrane was reported using β-actin as the loading control. The
relative protein concentration was analyzed using Quantity One
software (version 4.4.0; Bio-Rad Laboratories, Inc.).
Statistical analysis
Differences among groups were analyzed using one-way
analysis of variance followed by Fisher's Least Significant
Difference test. Results are presented as the mean ± standard
deviation. P<0.05 was considered to indicate a statistically
significant difference. SPSS software (version 17.0; SPSS, Inc.,
Chicago, IL, USA) was used for statistical analysis.
Results
Inhibition ratio of cell
proliferation
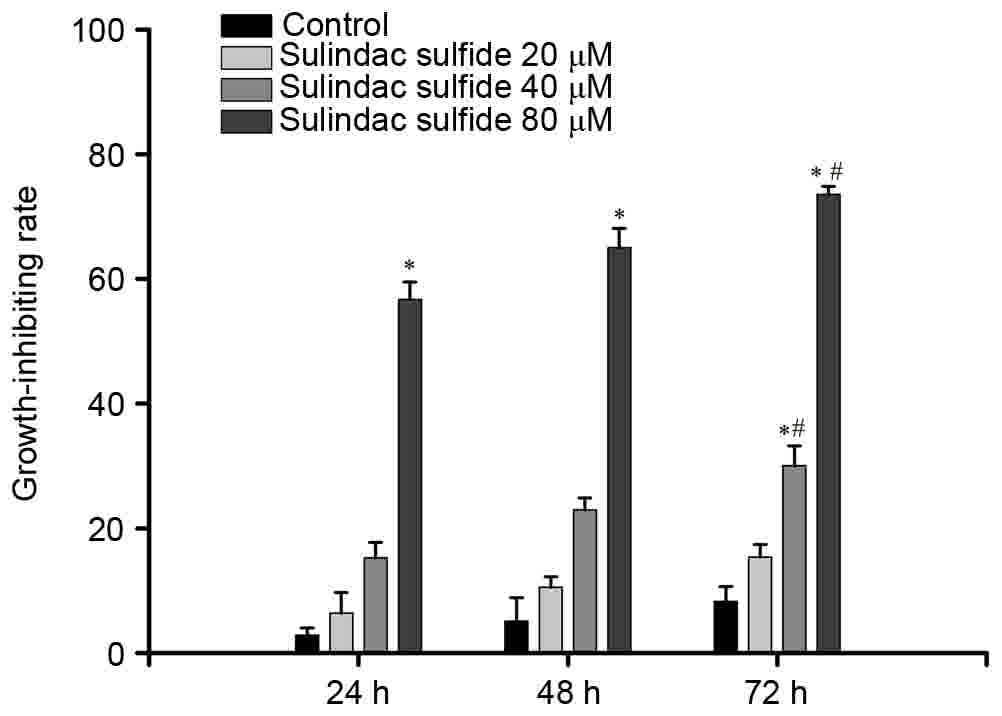
The effect of sulindac sulfide on MCF-7 cell
proliferation was analyzed. The cells were incubated with sulindac
sulfide (20, 40 and 80 µmol/l) for 24, 48 or 72 h. The data
demonstrated that addition of varying doses of sulindac sulfide
produced dose- and time-dependent increases in the inhibition ratio
of cell proliferation (Fig. 1). The
absolute values of inhibiting ratio are presented in Table I.
 | Table I.Growth-inhibiting effects of sulindac
sulfide on MCF-7 cells (n=3). |
Table I.
Growth-inhibiting effects of sulindac
sulfide on MCF-7 cells (n=3).
|
| Time, h |
|---|
|
|
|
|---|
| Group | 24 | 48 | 72 |
|---|
| Control | 2.83±1.2 | 5.07±3.8 | 8.25±2.4 |
| 20 µmol/l | 6.41±3.3 | 10.55±1.7 | 15.37±2.0 |
| 40 µmol/l | 15.24±2.5 | 22.96±1.9 |
30.01±3.2a,b |
| 80 µmol/l |
56.72±2.8a |
65.01±3.1a |
73.54±1.3a,b |
Effects of sulindac sulfide on cell
cycle progression
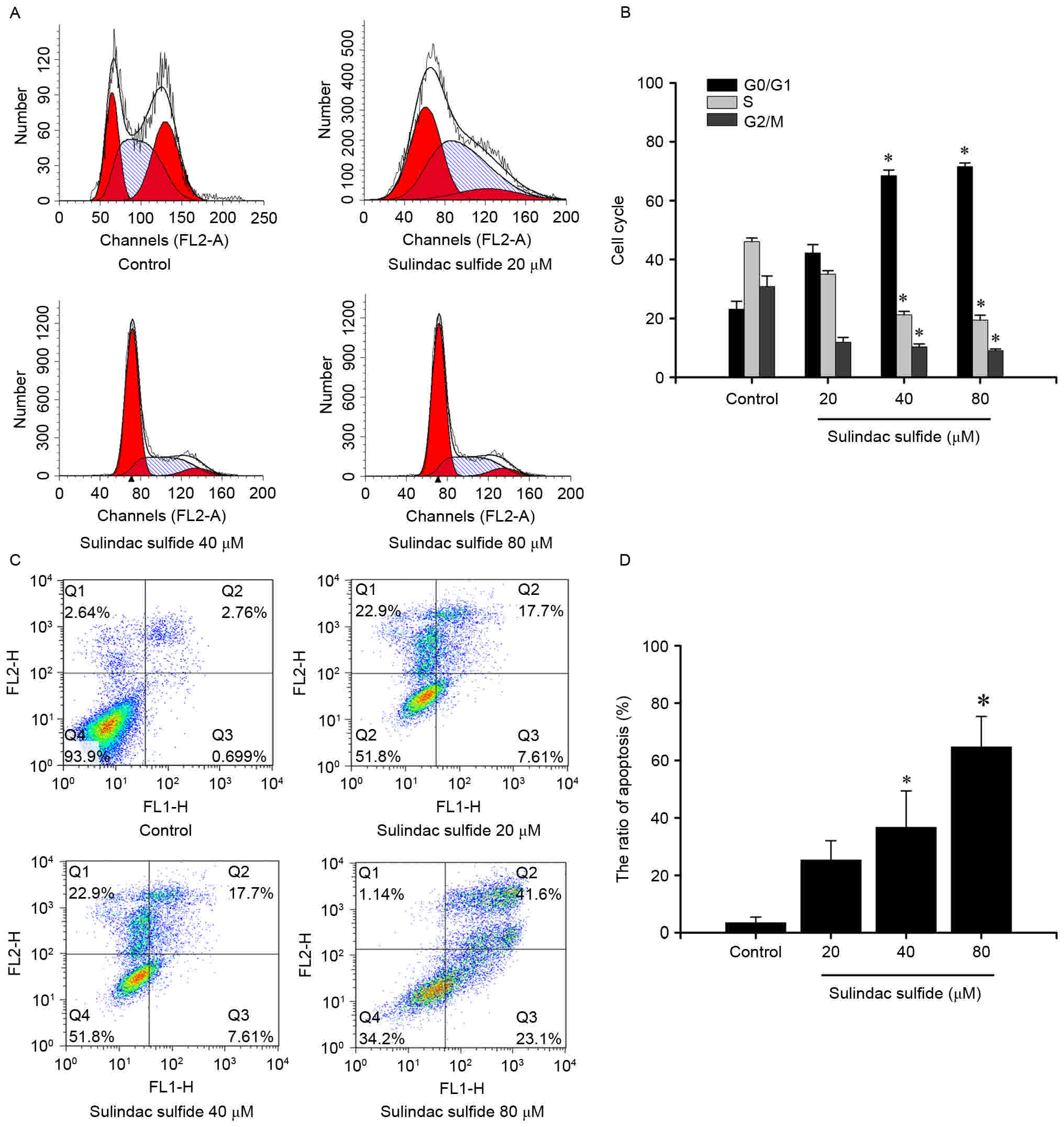
Flow cytometric analysis demonstrated that there was
no significant statistical significance between the 20 µmol/l group
and the control in each cell cycle phase (Fig. 2A and B). However, there was a
significant increase in G0/G1 phase cells
with a simultaneous decrease in S phase and G2/M phase
cells in the 40, and 80 µmol/l groups compared with that of the
control group (Fig. 2A and B;
P<0.05; Table II).
| Table II.Effects of sulindac sulfide on cell
cycle of MCF-7 cells using flow cytometry (n=3). |
Table II.
Effects of sulindac sulfide on cell
cycle of MCF-7 cells using flow cytometry (n=3).
|
| Cell cycle, % |
|---|
|
|
|
|---|
| Group |
G0/G1 | S | G2/M |
|---|
| Control | 23.1±2.7 | 46.1±1.2 | 30.8±3.6 |
| 20 µmol/l | 42.2±2.9 | 35.0±1.2 | 11.9±1.6 |
| 40 µmol/l | 68.5±1.9a | 21.2±1.2a | 10.3±1.0a |
| 80 µmol/l | 71.6±1.2a | 19.4±1.7a | 9.0±0.6a |
Effects of sulindac sulfide on the
apoptosis of MCF-7 cells
Annexin V-FITC/PI staining was used to measure the
percentage of apoptotic cells in response to sulindac sulfide
treatments. Following 72 h of exposure to 20, 40 or 80 µmol/l
sulindac sulfide, the apoptotic rates were 25.31±6.75, 36.7±12.71,
and 64.7±10.61%, respectively, compared with 3.46±1.95% of control
cells (Fig. 2C and D). These results
suggest that sulindac sulfide can induce the apoptosis of MCF-7
cells in a dose-dependent.
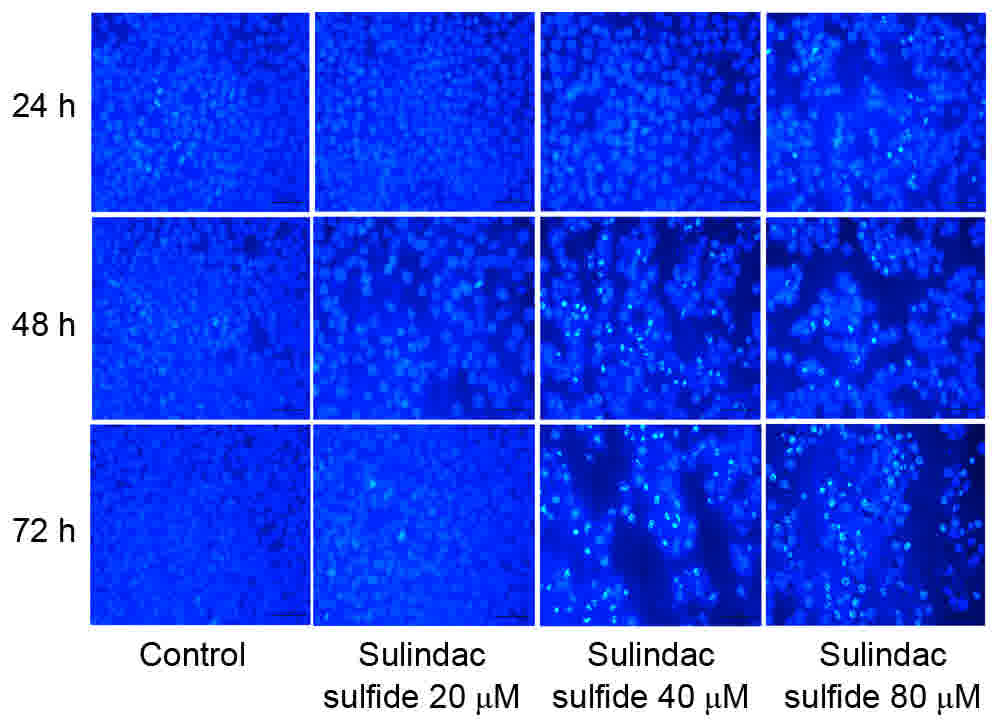
Hoechst 33258 staining
The results of Hoechst 33258 staining demonstrated
that sulindac sulfide could markedly induce the apoptosis of MCF-7
cells in a dose- and time-dependent manner (Fig. 3).
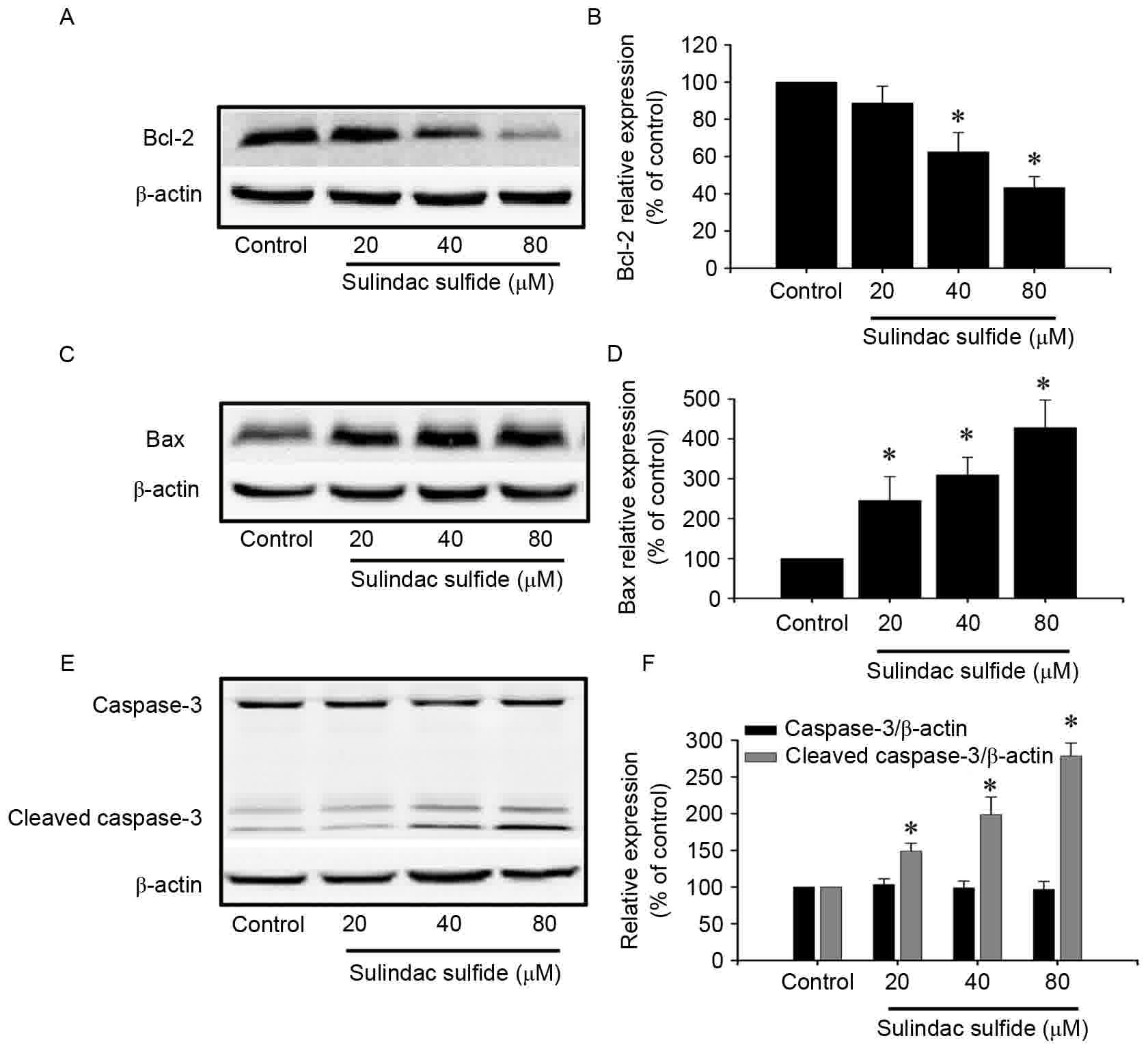
Effects of sulindac sulfide on protein
expression
Cells were treated with different concentrations of
sulindac sulfide or the control for 24 h. The protein expression of
Bcl-2 was downregulated, caspase-3 exhibited no significant change,
and Bax and cleaved caspase-3 were increased with increasing
concentrations of sulindac sulfide (Fig.
4).
Discussion
Breast cancer is a malignant tumor that often occurs
in females, its incidence is one of the highest among the female
malignant tumor types. Therefore, early prevention, detection and
treatment of breast cancer are of great value, and significance
(16). The imbalance between
proliferation and apoptosis of tumor cells is associated with tumor
formation. Furthermore, loss of cell cycle regulation can lead to
excessive cell proliferation and inhibition of cell apoptosis.
Therefore, inhibiting tumor cell proliferation and inducing its
apoptosis is an effective method for the treatment of tumors
(17).
Sulindac sulfide can not only inhibit a variety of
carcinogenic substances, but can also inhibit the growth of cancer
cells and induce its apoptosis; however, the underlying mechanism
remains unclear (4,18–20). A
study performed by Kim et al (21) demonstrated that the combined treatment
of sulindac and simvastatin augmented their anti-apoptotic
potential in lung cancer cells through protein kinase B
signaling-dependent downregulation of survivin. Furthermore, Fink
et al (22) suggested that
sulindac may be the most effective agent for colon cancer
prevention in humans with low 15-hydroxyprostaglandin dehydrogenase
levels, but may also be associated with inflammatory lesions in the
colon. The results from a study by Katoumas et al (23) revealed that treatment with sulindac
appears to delays the progression of oral premalignant lesions to
oral squamous cell carcinoma, resulting in smaller and better
differentiated tumors. These in vivo antineoplastic effects
may be associated with its ability to decrease cell proliferation
and to prevent survivin expression (23).
The results of the present study demonstrated that
sulindac sulfide can significantly inhibit the proliferation of
MCF-7 breast cancer cells, and the inhibitory effect was observed
to be concentration- and time-dependent. The proliferation of
cancer cells is regulated and controlled by key check points
G1/S and G2/M in the cell cycle (24). In each cell cycle phase, along with
the increasing concentrations of sulindac sulfide, the proportion
of G0/G1 phase cells was increased
significantly, and the proportion of S and G2/M phase
cells was reduced, demonstrated that sulindac sulfide inhibited the
transition from G0/G1 to S phase in breast
cancer cells. Apoptosis is influenced by multiple gene regulatory
proteins, the caspase family serve an important role in the process
of apoptosis, among them caspase-3 is a primary apoptosis-mediating
protease, which serves an essential role in the initiation of
apoptosis through various factors. Caspase-3 can cause a cascade
reaction following activation by upstream signaling factors Bax and
Bcl-2, subsequently leading to cell apoptosis. The present study
demonstrated that sulindac sulfide can activate the
apoptosis-promoting Bax gene and inhibit Bcl-2, triggering
downstream cascade activation of caspase-3, eventually leading to
apoptosis. In the current study, the expression of Bcl-2 was
decreased with increasing concentrations of sulindac sulfide,
caspase-3 exhibited no significant change. In addition, the
expression of Bax and cleaved caspase-3 was increased following
treatment with sulindac sulfide, suggesting that sulindac sulfide
induces cell apoptosis.
The results of the present study demonstrated that
sulindac sulfide can inhibit the proliferation and induce the
apoptosis of MCF-7 cells, using a mechanism associated with
alterations to the cell cycle and inhibition of Bcl-2 expression,
thus activating caspase-3. However, the specific mechanism of
sulindac sulfide remains unclear, as well as whether it can be
effectively applied for the clinical treatment of breast cancer.
The concentration and time course of sulindac sulfide in inhibiting
proliferation, and inducing apoptosis of human breast cancer
warrants further investigation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation Project of CQ (grant no. CSTC, 2010BB5111) and
the Project Foundation of Chongqing Municipal Education Committee
(grant no. KJ100308).
Availability of data and materials
The datasets used and analyzed in the present study
are available from the corresponding author on reasonable
request.
Author's contributions
HHS participated in the majority of experiments,
performed the data analyses for these experiments and wrote the
manuscript. YJZ planned the majority of experiments, analyzed the
results and wrote parts of the manuscript. HW participated in the
coordination of the study and reviewed the manuscript. LL and MC
completed the cell proliferation assay and cell culture, revised
the manuscript and helped perform data analysis. JJH designed the
work that led to the submission and approved the final version. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Span PN, Pollakis G, Paxton WA, Sweep FC,
Foekens JA, Martens JW, Sieuwerts AM and van Laarhoven HW: Improved
metastasis-free survival in nonadjuvantly treated postmenopausal
breast cancer patients with chemokine receptor 5 del32 frameshift
mutations. Int J Cancer. 136:91–97. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fu J, Xu X, Kang L, Zhou L, Wang S, Lu J,
Cheng L, Fan Z, Yuan B, Tian P, et al: miR-30a suppresses breast
cancer cell proliferation and migration by targeting Eya2. Biochem
Biophys Res Commun. 445:314–319. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Diller M, Schüler S, Buchholz S, Lattrich
C, Treeck O and Ortmann O: Effects of estriol on growth, gene
expression and estrogen response element activation in human breast
cancer cell lines. Maturitas. 77:336–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lenik J: Preparation and characterization
of a sulindac sensor based on PVC/TOA-SUL membrane. Mater Sci Eng C
Mater Biol Appl. 37:383–389. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Giardiello FM, Hamilton SR, Krush AJ,
Piantadosi S, Hylind LM, Celano P, Booker SV, Robinson CR and
Offerhaus GJ: Treatment of colonic and rectal adenomas with
sulindac in familial adenomatous polyposis. N Engl J Med.
328:1313–1316. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Beazer-Barclay Y, Levy DB, Moser AR, Dove
WF, Hamilton SR, Vogelstein B and Kinzler KW: Sulindac suppresses
tumorigenesis in the Min mouse. Carcinogenesis. 17:1757–1760. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mahmoud NN, Boolbol SK, Dannenberg AJ,
Mestre JR, Bilinski RT, Martucci C, Newmark HL, Chadburn A and
Bertagnolli MM: The sulfide metabolite of sulindac prevents tumors
and restores enterocyte apoptosis in a murine model of familial
adenomatous polyposis. Carcinogenesis. 19:87–91. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Piazza GA, Alberts DS, Hixson LJ, Paranka
NS, Li H, Finn T, Bogert C, Guillen JM, Brendel K, Gross PH, et al:
Sulindac sulfone inhibits azoxymethane-induced colon carcinogenesis
in rats without reducing prostaglandin levels. Cancer Res.
57:2909–2915. 1997.PubMed/NCBI
|
|
9
|
Thompson HJ, Jiang C, Lu J, Mehta RG,
Piazza GA, Paranka NS, Pamukcu R and Ahnen DJ: Sulfone metabolite
of sulindac inhibits mammary carcinogenesis. Cancer Res.
57:267–271. 1997.PubMed/NCBI
|
|
10
|
Altuvia Y, Landgraf P, Lithwick G, Elefant
N, Pfeffer S, Aravin A, Brownstein MJ, Tuschl T and Margalit H:
Clustering and conservation patterns of human microRNAs. Nucleic
Acids Res. 33:2697–2706. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee YS, Choi D, Kim NY, Yang S, Jung E,
Hong M, Yang D, Lenz HJ and Hong YK: CXCR2 inhibition enhances
sulindac-mediated suppression of colon cancer development. Int J
Cancer. 135:232–237. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Verma R, Brahmankar M, Kushwah L and
Suresh B: Evaluating the inhibitory potential of sulindac against
the bleomycin-induced pulmonary fibrosis in wistar rats. Environ
Toxicol Pharmacol. 36:769–778. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liggett JL, Min KW, Smolensky D and Baek
SJ: A novel COX-independent mechanism of sulindac sulfide involves
cleavage of epithelial cell adhesion molecule protein. Exp Cell
Res. 326:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang WW, Ko SW, Tsai HY, Chung JG, Chiang
JH, Chen KT, Chen YC, Chen HY, Chen YF and Yang JS: Cantharidin
induces G2/M phase arrest and apoptosis in human colorectal cancer
cells through inhibition of CDK1 activity and caspase-dependent
signaling pathways. Int J Oncol. 38:1067–1073. 2011.PubMed/NCBI
|
|
15
|
Zhang YT, Ouyang DY, Xu LH, Ji Y, Zha Q,
Cai J and He X: Cucurbitacin B induces rapid depletion of the
G-actin pool through reactive oxygen species-dependent actin
aggregation in melanoma cells. Acta Biochim Biophys Sin (Shanghai).
43:556–567. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Q, Ouyang DY, Geng M, Xu LH, Zhang
YT, Wang FP and He XH: Valproic acid exhibits biphasic effects on
apoptotic cell death of activated lymphocytes through differential
modulation of multiple signaling pathways. J Immunotoxicol.
8:210–218. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Greening DW, Ji H, Kapp EA and Simpson RJ:
Sulindac modulates secreted protein expression from LIM1215 colon
carcinoma cells prior to apoptosis. Biochim Biophy Acta.
1834:2293–2307. 2013. View Article : Google Scholar
|
|
18
|
Corbex M, Bouzbid S and Boffetta P:
Features of breast cancer in developing countries, examples from
North-Africa. Eur J Cancer. 50:1808–1818. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li H, Yang AL, Chung YT, Zhang W, Liao J
and Yang GY: Sulindac inhibits pancreatic carcinogenesis in
LSL-Kras G12D-LSL-Trp53R172H-Pdx-1-Cre mice via suppressing
aldo-keto reductase family 1B10 (AKR1B10). Carcinogenesis.
34:2090–2098. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sinicrope FA and Penington RC: Sulindac
sulfide-induced apoptosis is enhanced by a small-molecule Bcl-2
inhibitor and by TRAIL in human colon cancer cells overexpressing
Bcl-2. Mol Cancer Ther. 4:1475–1483. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim YS, Seol CH, Jung JW, Oh SJ, Hwang KE,
Kim HJ, Jeong ET and Kim HR: Synergistic effect of sulindac and
simvastatin on apoptosis in lung cancer A549 cells through
AKT-dependent downregulation of survivin. Cancer Res Treat.
47:90–100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fink SP, Dawson DM, Zhang Y, Kresak A,
Lawrence EG, Yang P, Chen Y, Barnholtz-Sloan JS, Willis JE,
Kopelovich L and Markowitz SD: Sulindac reversal of
15-PGDH-mediated resistance to colon tumor chemo-prevention with
NSAIDs. Carcinogenesis. 36:291–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Katoumas KM, Nikitakis N, Perrea D, Dontas
I and Sklavounou A: In vivo antineoplastic effects of the NSAID
sulindac in an oral carcinogenesis model. Cancer Prev Res (Phila).
8:642–649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Smalley W, Ray WA, Daugherty J and Griffin
MR: Use of nonsteroidal anti-inflammatory drugs and incidence of
colorectal cancer: A population-based study. Arch Intern Med.
15:161–166. 1999. View Article : Google Scholar
|