Introduction
Hepatoma is a malignant tumor that occurs more
frequently in children than adults; it often cannot be completely
excised and the 5-year survival rate was ~40% in China (1). Multiple factors are involved in the
carcinogenesis of hepatoma, including genetic susceptibility and
environmental factors (2,3). Currently, although multiple therapeutic
methods, including surgery and chemotherapy, are available for the
treatment of hepatoma, the prognosis is often not satisfactory
owing to the rates of recurrence and metastasis (4,5). Thus, the
development of novel drugs with high efficacy and low toxicity for
the treatment of hepatoma is required.
Traditional Chinese medicines (TCMs) have been used
to treat cancer in China for thousands of years owing to their high
efficacy and low toxicity (6,7). Certain natural products have been widely
used for the treatment of cancer, including hepatoma (8,9).
Zerumbone, a monocyclic sesquiterpene derived from Zingiber
zerumbet (L.) Smith, is reported to produce a variety of
pharmacological effects, including antioxidant, antiviral,
anti-inflammatory antibacterial activities (10). Notably, zerumbone has been reported to
have anticancer efficacy in certain cancer cell lines, including
those of breast, bladder and brain cancer (11,12);
however, there are limitations to the antitumor mechanism.
The present study aimed to investigate the effect of
zerumbone on the proliferation, cell cycle distribution and
apoptosis of hepatoma HepG2 cells, and assess its possible
mechanism in vitro. These data demonstrated that zerumbone
could inhibit HepG2 cell proliferation and metastasis of hepatoma
by inducing apoptosis. Furthermore, it was also demonstrate that
zerumbone induced apoptosis through the mitogen-activated protein
kinase (MAPK)-extracellular signal-regulated kinase (ERK) pathways.
In conclusion, the data of the present study indicated that
zerumbone may be a useful plant product for the treatment of
hepatoma.
Materials and methods
Cell culture
The human hepatoma HepG2 cell line was purchased
from the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Cells were maintained in Dulbecco's Modified
Eagle's Medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) in a humidified incubator
under 5% CO2 at 37°C.
MTT assay for cell viability
The cytotoxicity of zerumbone was detected by MTT
assay. Briefly, HepG2 cells were seeded in 96-well culture plates
at a density of 4×104 cells per well at 37°C for 24 h.
Next, cells were incubated with different concentrations (0, 2, 5,
10, 20, 50, 100 µM) of zerumbone in culture medium for 48 h.
Following this, 20 µl/well of MTT solution (dissolved in PBS) was
added and the plates were incubated at 37°C for 4 h. The absorbance
was measured at 490 nm with a microplate reader to assess the
optical density (OD), from which cell counts were determined. The
inhibition rate of zerumbone on these cells was calculated as
follows: Inhibition rate (%)=(1-experimental group OD/control group
OD) ×100.
Flow cytometry analysis
Cells were treated with 0.1% DMSO control or
zerumbone (10 or 30 µM) for 48 h, and then fixed overnight with 70%
ethanol in ice-cold PBS at −20°C, and then resuspended with 50
µg/ml of propidium iodide (PI) (Sigma-Alrich; Merck KgaA,
Darmstadt, Germany) and 50 µg/ml of RNAse A (Sangon Biotech Co.,
Ltd., Shanghai, China) and incubated in the dark for 15 min at room
temperature. The DNA contents of samples were analyzed using a
FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA).
Apoptosis was determined using Annexin V-FITC. Following treatment
of HepG2 cells with different concentrations (10 or 30 µM) of
zerumbone for 48 h. Cells were washed twice with PBS at 4°C and the
supernatant was discarded. Add 100 µl pre-cooled binding buffer
(EMD Millipore, Billerica, MA, USA) to re-suspend the cells for ice
bath. Next, 10 µl Annexin V-fluorescein isothiocyanate (FITC; EMD
Millipore) and 10 µl PI were added to the cell suspension and
gently mixed. The calls were then incubated at room temperature for
15 min dark staining, after which another 400 µl binding buffer was
added to re-suspend the cells. Next, Multimode Plate Reader
(Varioskan LUX; Thermo Fisher Scientific Inc., Waltham, MA, USA)
was used to detect the percentage of apoptotic cells. The
experiment was repeated three times.
Cell adhesion assay
A cell adhesion assay were performed, as previously
described (3). Briefly, each well of
a 96-well plate was coated with 10 µl fibronectin (R&D Systems,
Inc., Minneapolis, MN, USA), and the plates were incubated at 37°C
for 2 days and washed twice with DMEM. HepG2 cells were pretreated
with 0, 10 or 30 µM zerumbone for 48 h at 37°C, cells were
harvested and then seeded into the 96-well fibreonectin-coated
plate at a density of 5×105 cells/ml for 100 µl. After 2
h, the plates were washed and 10 µl MTT (5 mg/ml) solution
(dissolved in PBS) was added to the adhered cells. Absorbance was
detected at 570 nm using the Multilabel counter after a 4-h
incubation. Values reported are from 3 independent experiments.
Cell migration assay
HepG2 cells were pretreated with 0, 10 or 30 µM
zerumbone for 48 h at 37°C. Cells were collected and adjusted to a
single-cell suspension of 5×105/ml cells. Next, 100-µl
cell suspension was placed in each insert upper chamber without
matrigel (Corning Incorporated, Corning, NY, USA) containing 200 µl
FBS free DMEM, whereas 600 µl DMEM in the lower chamber contained
10% FBS. After incubating for 6 h, the chamber was washed twice
with PBS, and the cells on the apical side of each insert were
scraped off using cotton buds. The cells that had migrated and
attached to the lower surface of the insert were fixed with 4%
formaldehyde 10 min and stained with 0.1% crystal violet for 20 min
at room temperature. After washing with PBS and air-drying, the
number of cells that had migrated through the membrane was counted
randomly in five fields under a light microscope (magnification,
×400). The migration rate was calculated using the following
formula: Migration rate (%)=number of migrated cell/number of
inoculated cells ×100.
Cell invasion assay
For the cell invasion assay, transwell filters
(Corning Incorporated) were coated with 100 µl Matrigel (300 µg/ml)
on the upper surface of a polycarbonic membrane. The chambers were
placed into a 37°C incubator for 2 h prior to use. Next,
5×104 cells in 200 µl low serum (1% FBS) DMEM containing
0.1% DMSO (control), 10 or 30 µM zerumbone was added to the upper
chamber, and 600 µl medium containing 10% FBS was added to the
lower chambers. Cells were cultured at 37°C in a humidified
incubator with 5% CO2 for 24 h. After incubation,
invaded cells on the lower side of the membrane were fixed with 4%
paraformaldehyde for 20 min at room temperature, and then washed
with PBS three times before being stained with 0.1% crystal violet
for another 10 min. The stained cells were visualized under an
inverted microscope (magnification, ×200) and counted in five
random fields. The results were averaged for three independent
experiments.
Western blot analysis
HepG2 cells were harvested, rinsed twice with
ice-cold PBS and re-suspended in radioimmunoprecipitation assay
lysis buffer (Sigma-Aldrich; Merck KGaA) and 50 ng protein samples
were quantified using a bicinchoninic acid assay Sigma-Aldrick;
Merck KGaA) and separated by 12% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes. The membranes were dried,
pre-blocked with 5% non-fat milk in PBS-Tween (0.1%) overnight at
room temperature, then incubated with the following specific
antibodies overnight at 4°C: Cyclin dependent kinase inhibitor 1B
(CDKN1B, also known as p27) (1:2,000), cyclin-dependent kinase 1
(CDK1, also known as CDC2) (1:2,000), cyclin B1 (1:2,000), B-cell
lymphoma-2 (Bcl-2) (1:2,000), Bcl-2-associated X (Bax) (1:2,000),
cytochrome c (1:2,000), cleaved caspase-3 (1:1,000), cleaved
caspase-9 (1:1,000), focal adhesion kinase (FAK) (1:2,000), Ras
homolog gene family, member A (RhoA) (1:2,000), Rho-associated
protein kinase-1 (ROCK-1) (1:2,000), matrix metalloproteinase-2
(MMP-2) (1:2,000), MMP-9 (1:2,000), phosphorylated (p)-p38 MAPK
(1:1,000), p38 MAPK (1:1,000), p-ERK1/2 (1:1,000), ERK1/2
(1:1,000), p-c-Jun N-terminal kinase (JNK) (1:1,000), JNK (1:1,000)
and β-actin (1:5,000) at 4°C overnight followed by incubated with
goat anti-mouse horseradish peroxidase (HRP)-conjugated secondary
antibody (1:2,000; cat no. ab7068; Abcam, Cambridge, UK) at room
temperature for 1 h. Immune complexes were visualized using the
Pierce ECL Western Blotting kit (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The expression levels of Bcl-2, Bax, cytochrome c,
FAK, RhoA, ROCK-1, MMP-2 and MMP-9 were detected by RT-qPCR using
ABI Prism 7900 sequence detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). Total RNA was extracted from the
HepG2 cells which were treated by 10 or 30 µM zerumbone 48 h using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Subsequently, cDNA was
synthesized using M-MLV First Strand cDNA Synthesis kit
(Invitrogen; Thermo Fisher Scientific, Inc.). qPCR reactions were
performed using a SYBR Premix Ex Taq (Takara Biotechnology Co.,
Ltd., Dalian, China) according to the manufacturer's protocol.
Thermocycling conditions were as follows: Denaturation for 30 sec
at 95°C, 20 sec at 95°C, 45 sec at 68°C for 30 cycles, 5 min at
68°C for final extension, then 4°C for holding. Specific primers
for each gene were designed as follows: The PCR primer sets were
following: p27 forward, 5′-CTGCCCTCCCCAGTCTCTCT-3′ and reverse,
5′-CAAGCACCTCGGATTTT-3′; CDC2 forward,
5′-GCGGCGGGGTACCCCCTGCAGTAAGTGCAGAAATCT-3′ and reverse,
5′-CGCCGGAGGATCTTCGCAGCGGCAGCTACAACAAC-3′; cyclin B1 forward,
5′-GCAAATGACAAAGCAAATGGGG-3′ and reverse,
5′-ACAACCAGCAGAAACCAACAGC-3′; Bcl-2 forward,
5′-CGTACAGTTCCACAAAGGCA-3′ and reverse, 5′-ATGTGTGTGGAGAGCGTCAA-3′;
Bax forward, 5′-CCCGAGAGGTCTTTTTCCGAG-3′ and reverse,
5′-CCAGCCCATGATGGTTCTGAT-3′; cytochrome c forward,
5′-CTTTGGGCGGAAGACAGGTC-3′ and reverse 5′-TTATTGGCGGCTGTGTAAGAG-3′;
FAK forward, 5′-GCAATTTCCTGGTCCACTTG-3′ and reverse,
5′-CGTTATTTGCCAAAAGGATTTC-3′; RhoA forward,
5′-GGAAAGCAGGTAGAGTTGGCT-3′ and reverse,
5′-GGCTGTCGATGGAAAAACACAT-3′; ROCK-1 forward,
5′-GGCAGGAAAATCCAAATCAT-3′ and reverse, 5′-GGGGACAGTTTTGAGACTCG-3′;
MMP2 forward, 5′-AAGAAGTAGCTGTGACCGCC-3′ and reverse,
5′-TTGCTGGAGACAAATTCTGG-3′; MMP9 forward,
5′-GCACTGCAGGATGTCATAGG-3′ and reverse, 5′-ACGACGTCTTCCAGTACCGA-3′;
and GAPDH forward, 5′-CCACATCGCTCAGACACCAT-3′ and reverse,
5′-ACCAGGCGCCCAATACG-3′. Relative mRNA expression was calculated
using the 2−ΔΔCq method (13), using GAPDH as an endogenous control
and untreated samples as the calibrator.
Statistical analysis
The experimental data were analyzed using SPSS 11.0
(SPSS, Inc., Chicago, IL, USA). Data are expressed and the mean ±
standard deviation. The quantitative ratios of different groups
were compared using one-way analysis of variance with tukey's post
test. P<0.05 was considered to indicate a statistically
significant difference. All statistical tests were two sided.
Results
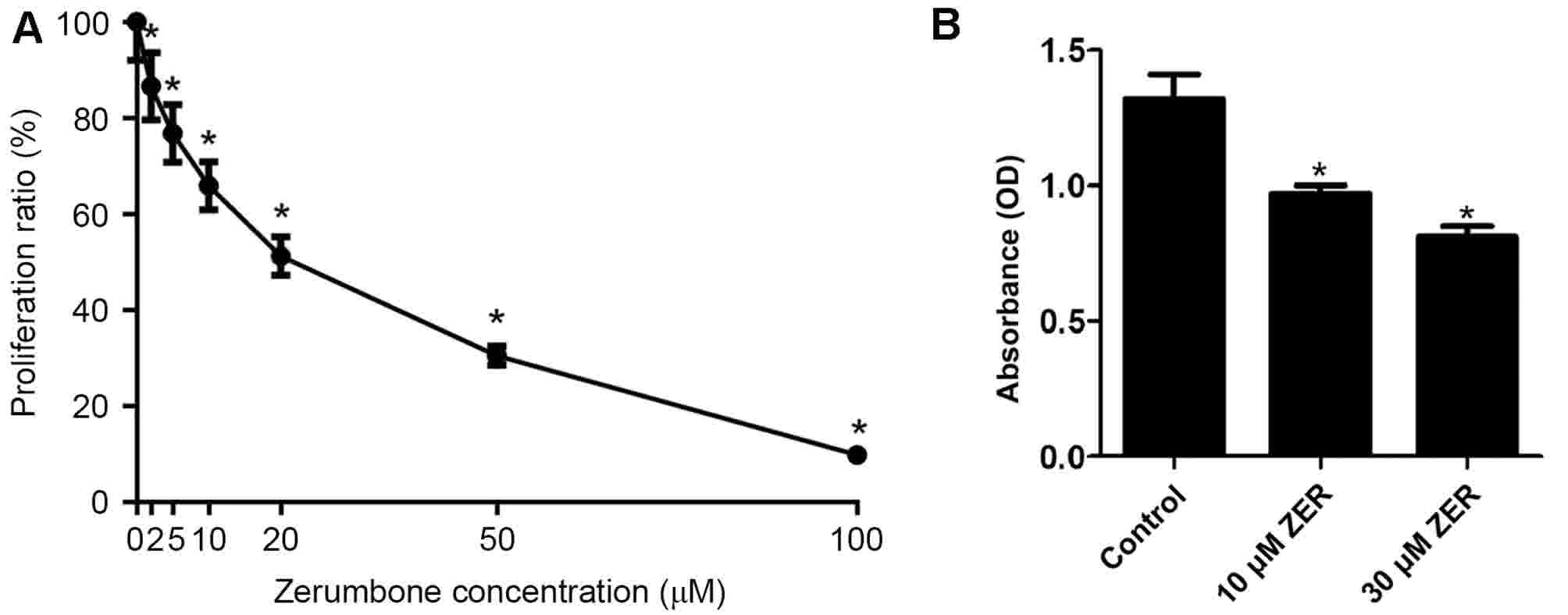
Zerumbone inhibits the proliferation
of HepG2 cells
After cells were treated with different
concentrations (0, 2, 5, 10, 20, 50 or 100 µM, n=5) of zerumbone
for 48 h, the cell growth inhibition for the HepG2 cells is
presented in Fig. 1. The results of
the MTT assay demonstrated that zerumbone inhibited the growth of
HepG2 cells in a concentration-dependent manner (Fig. 1). The half-maximal inhibitory
concentration (IC50) value for zerumbone was 23.64±1.23
µM. Thus, doses higher and lower than the IC50 (10 and
30 µM) were used in subsequent experiments.
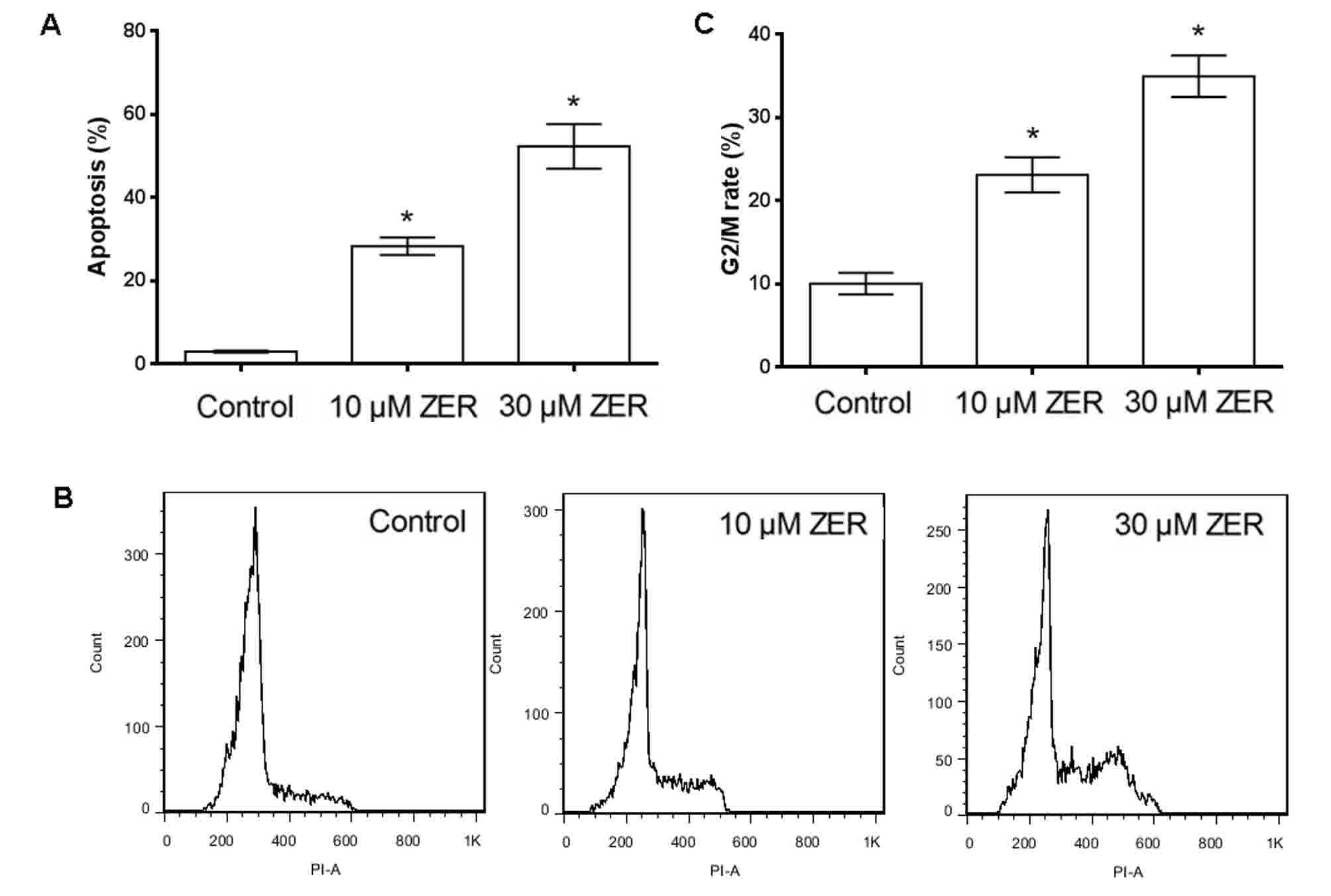
Zerumbone induces apoptosis in HepG2
cells
HepG2 cells were cultured in 10 or 30 µM zerumbone
for 48 h and the proportion of apoptotic HepG2 cells was then
determined using annexin V-FITC staining and flow cytometry.
Following treatment of HepG2 cells with 10 or 30 µM zerumbone for
48 h, the percentage of apoptotic cells increased from 3.00 to
28.28 or 52.30% following 10 or 30 µM zerumbone treatment,
respectively (Fig. 2A). These results
indicated that zerumbone treatment could induce apoptosis in HepG2
cells in a concentration-dependent manner.
Zerumbone induced cell cycle arrest at
G2/M phase
Cell cycle analysis by flow cytometry showed
treatment with 10 or 30 µM zerumbone for 48 h increased the
proportion of cells in G2/M phase (from 10.05±0.13 to
23.1±0.21 or 34.96%±0.25 following 10 or 30 µM zerumbone treatment,
respectively) (Fig. 2B and C). These
results indicated that zerumbone induced cell cycle arrest at
G2/M in HepG2 cells in a concentration-dependent
manner.
Zerumbone reduces the adherence
abilities of HepG2 cells
To test the effect of zerumbone on the capacity of
cancer cells to bind to the extracellular matrix, a cell adhesion
assay was performed in HepG2 cells. As shown in Fig. 2, the adherence abilities of HepG2
cells were significantly reduced following pre-treatment with
zerumbone. These results indicated that zerumbone effectively
inhibited the adherence ability of HepG2 cells.
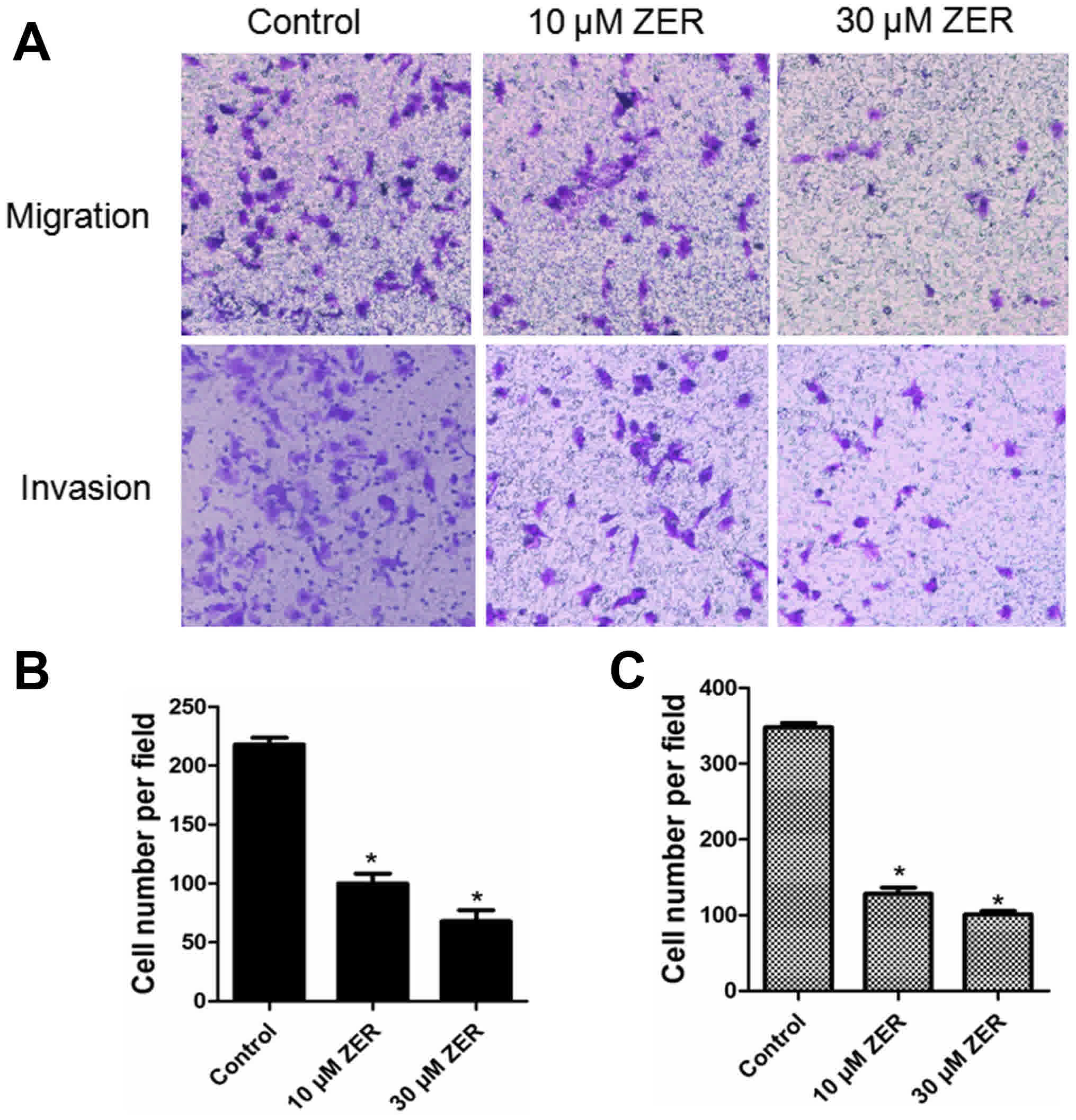
Zerumbone inhibits cancer cell
migration
Cellular migration was analyzed using a transwell
migration assay. As shown in Fig. 3A,
compared with the control group, the migratory ability of HepG2
cells was significantly inhibited by zerumbone treatment in a
dose-dependent manner (P<0.05). The results indicated that
zerumbone treatment significantly inhibited the migration of the
HepG2 cells.
Zerumbone inhibits cancer cell
invasion
The effect of zerumbone on cancer cell invasion by
was investigated using a transwell invasion assay. As shown in
Fig. 3B, the cell invasion rate in
the zerumbone-treated groups was significantly lower than those in
the control group.
Zerumbone induces activation of
caspase-3 and-9 and apoptosis and cell cycle-associated genes in
HepG2 cells
The caspase cascade triggered by caspase family
members, among them the frequently activated death proteases
caspase-3 and-9, serves a key role in apoptosis (14). To investigate whether caspase
activation was involved in zerumbone-induced apoptosis, activation
of caspase-3 and-9 was detected by western blotting. As shown in
Fig. 4A, HepG2 cells treated with
zerumbone markedly increased the expression level of cleaved
caspase-3 and-9, compared with the control cells. Other
apoptosis-and cell cycle-associated genes in HepG2 cells were also
investigated by western blotting and RT-qPCR. As shown in Fig. 4, compared to the control, zerumbone
treatment significantly increased the expression level of p27, Bax,
and cytochrome-c, but decreased the expression level of CDC2,
cyclinB1 and Bcl-2. These results indicated that zerumbone induced
the activation of caspase-3, caspase-9, cytochrome-c and Bax, and
decreased the expression level of Bcl-2, eventually leading to the
apoptosis in HepG2 cells.
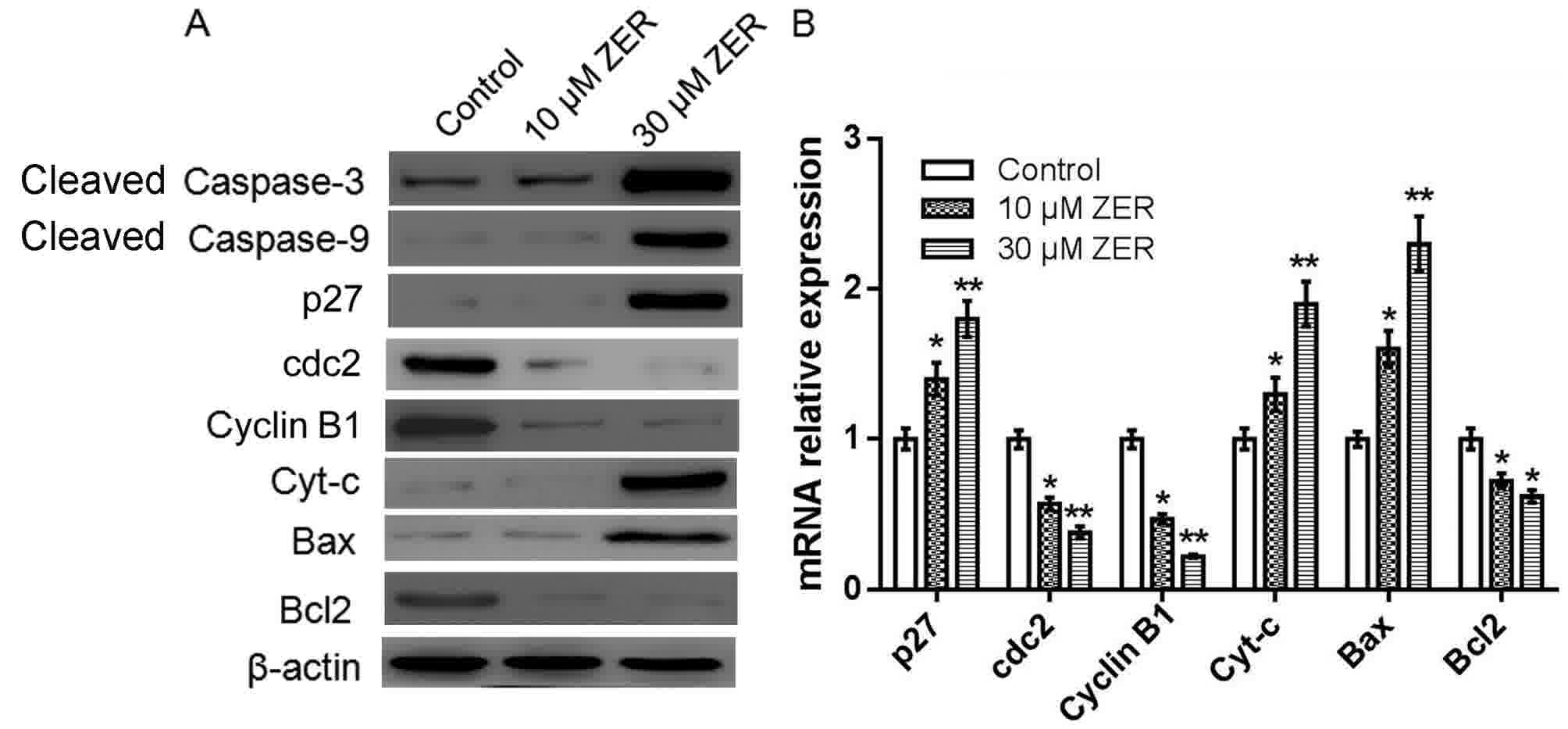 | Figure 4.Zerumbone induces activation of
caspase-3 and-9 and other apoptosis-associated genes in HepG2
cells. (A) The protein expression of cleaved caspase-3, cleaved
caspase-9, p27, CDC2, cyclinB, cytochrome c, Bax and Bcl-2 were
measured by western blot analysis. β-actin was used as an internal
control. (B) The mRNA expression of Cyt-c, Bax and Bcl-2 were
measured by reverse transcription-quantitative polymerase chain
reaction analysis. All experiments were performed in triplicate.
*P<0.05, **P<0.01 vs. control. p27, cyclin-dependent kinase
inhibitor 1B; CDC2, cyclin-dependent kinase 1; Cyt-c, cytochrome c;
Bcl-2, B-cell lymphoma-2; Bax, Bcl-2-associated X; ZER,
zerumbone. |
Zerumbone treatment downregulates the
expression levels of FAK, RhoA, ROCK-1, MMP-2 and MMP-9 in HepG2
cells
FAK, RhoA, ROCK-1, MMP-2 and MMP-9 are
metastasis-associated proteins. The present study determined the
effect of zerumbone on the expression of FAK, RhoA, ROCK-1, MMP-2
and MMP-9 by western blotting and RT-qPCR. As depicted in Fig. 5, treatment with zerumbone reduced the
mRNA and protein expression of FAK, RhoA, ROCK-1, MMP-2 and MMP-9
in HepG2 cells, compared with the control group.
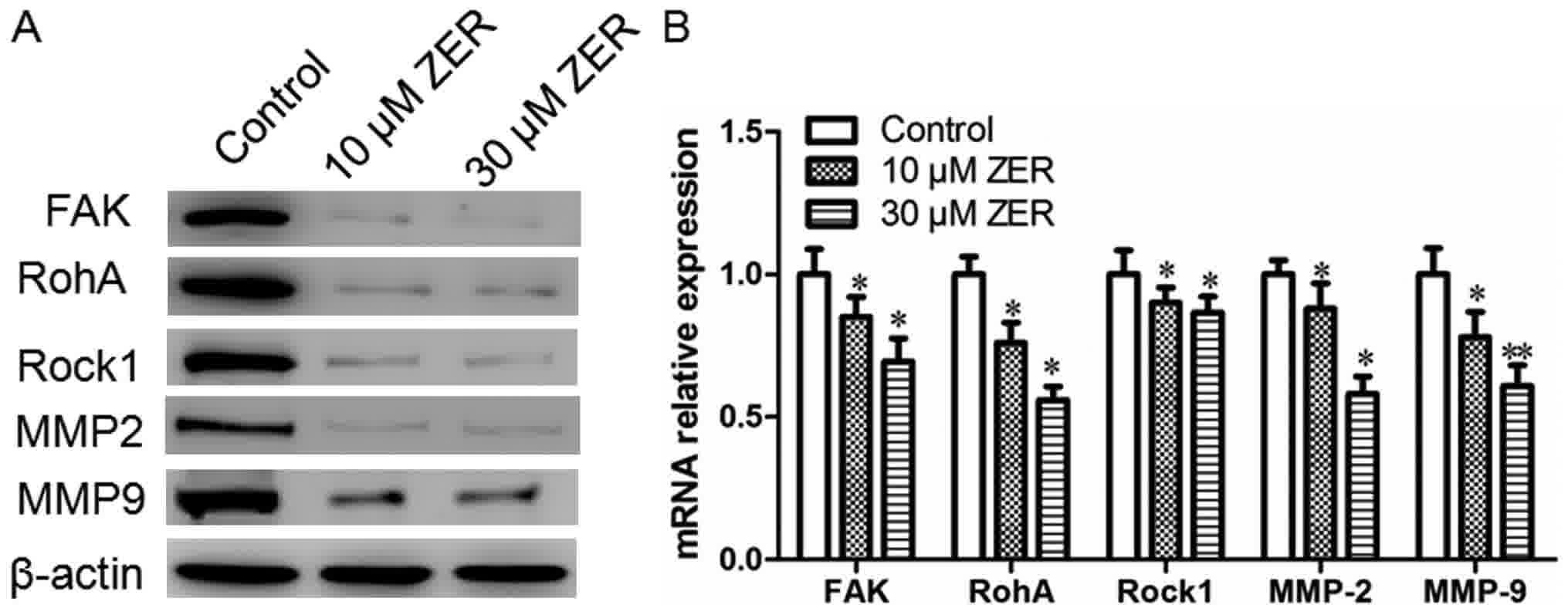 | Figure 5.Zerumbone downregulates the expression
levels of FAK, RhoA, ROCK-1, MMP2 and MMP9 in HepG2 cells. (A) The
protein expression of FAK, RhoA, ROCK-1, MMP2 and MMP9 were
measured by western blot analysis. β-actin was used as an internal
control. (B) The mRNA expression of FAK, RhoA, ROCK-1, MMP2 and
MMP9 were measured by reverse transcription-quantitative polymerase
chain reaction analysis. All experiments were performed in
triplicate, *P<0.05, **P<0.01 vs. control. FAK, focal
adhesion kinase; RhoA, Ras homolog gene family, member A; ROCK-1,
Rho-associated protein kinase-1; MMP2, matrix metalloproteinase-2;
ZER, zerumbone. |
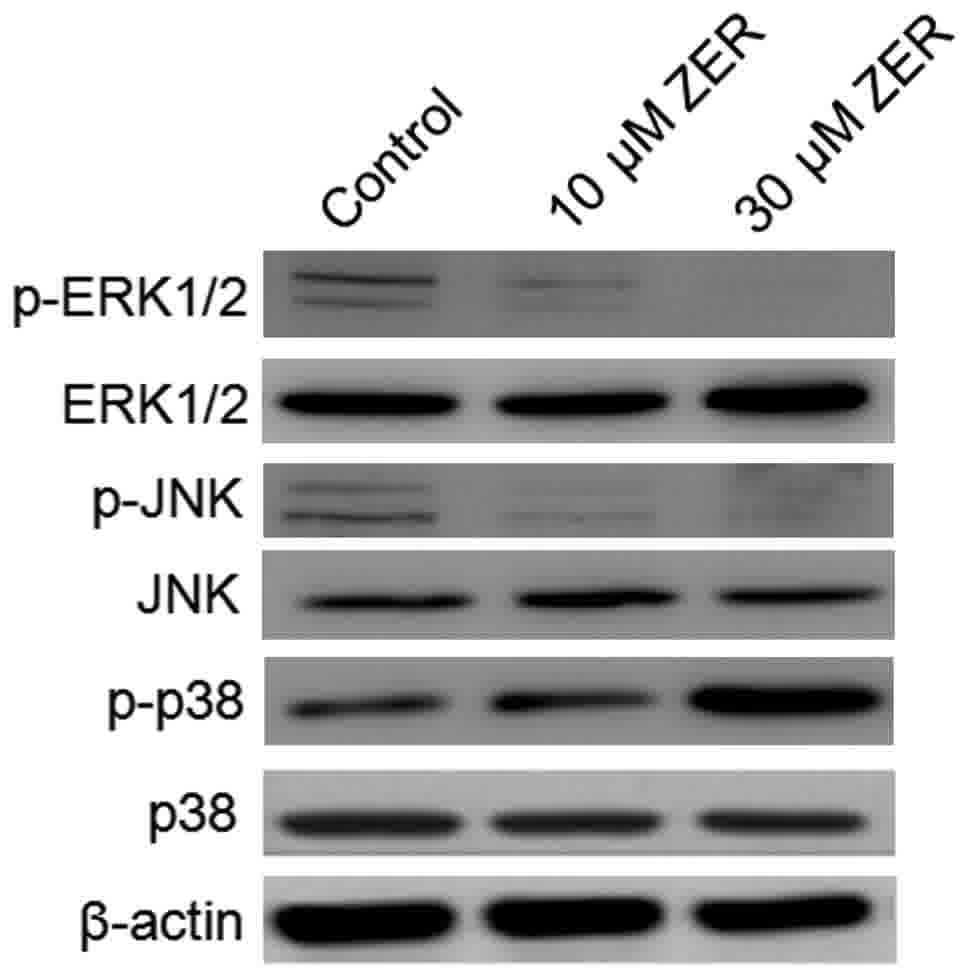
Zerumbone regulates MAPK and ERK
phosphorylation in HepG2 cells
The MAPK signaling pathway serves a notable role in
the action of chemotherapeutic drugs. Zerumbone inactivated ERK and
activated p38 MAPK in a time-dependent manner (Fig. 6). No effect of zerumbone treatment was
observed on the regulation of JNK. These findings indicate that
inactivation of ERK and activation of p38 MAPK may serve a crucial
upstream role in mediating zerumbone activity and inhibiting
metastasis in HepG2 cells.
Discussion
Hepatoma has a high incidence in China and the
prevalence was 30–40% at 5 years postoperatively globally (15). It is difficult to detect during its
early stages owing to a lack of symptoms (16). Prognosis for patients with hepatoma is
poor owing to the high rate of recurrence and metastasis. Thus, it
is essential to investigate the molecular mechanisms driving
recurrence and metastasis for improving patient prognosis, and to
develop a novel strategy for treating patients with hepatoma by
inhibiting specific targets. The present study evaluated the
ability of zerumbone to suppress the growth of hepatoma cancer
cells in vitro. Zerumbone could significantly inhibit
adhesion, migration and invasion in HepG2 cells, and could induce
apoptosis. Western blot analysis revealed that zerumbone could
induce activation of caspase-3, caspase-9 and and Bax, decrease the
expression level of Bcl-2, suppress the expression of MMP-2 and
MMP-9, the activity of JNK and ERK was increased, and increase the
expression of p-p38 MAPK. These results indicated that zerumbone
could inhibit hepatoma growth and metastasis via the MAPK-ERK
signal pathway.
To elucidate the mechanism of zerumbone action in
hepatoma cells, the expression of cell cycle-and
apoptosis-associated genes was examined in HepG2 cells. The release
of cytochrome c from the mitochondria into cytoplasm has been
considered to be an important event in the apoptotic process
mediated by mitochondria (17). In
the present study, the data revealed that zerumbone could cause the
release of cytochrome c. Induction of apoptosis was associated with
the regulation of anti-and pro-apoptotic proteins. Bcl-2 (a notable
apoptosis-inhibiting protein) and Bax (an apoptosis-promoting
protein) serve key roles in the apoptotic process (18). Induction of Bax expression results in
a downstream program of mitochondrial dysfunction, leading to
caspase-9 activation and the subsequent activation of caspase-3
(19). The present study revealed
that zerumbone treatment of HepG2 cells resulted in a
dose-dependent decrease in Bcl-2 levels and an increase in Bax.
Caspase-3 and-9 were also activated in a dose-dependent manner
following zerumbone treatment. These results demonstrated that
zerumbone induced mitochondrial activation-mediated cell apoptosis
and HepG2 cell death.
The development and progression of tumor metastasis
involves a complex multistep process, including cancer cell
adhesion, migration and invasion (20). A previous study demonstrated that
changes in cytogenetic characteristics are responsible for the
invasion and metastasis of cancer cells (21). RhoA, ROCK-1, FAK, MMP-2 and MMP-9 are
classical metastasis-associated genes; the effect of zerumbone on
these molecules was assessed to investigate the underlying
mechanism of zerumbone in vitro (22). RhoA, is a small GTPase protein
belonging to the Rho family; it is primarily associated with
cytoskeletal regulation, mediating actin stress fiber formation and
actomyosin contractility (23).
ROCK-1 is a serine/threonine kinase that belongs to the Rho family
and acts to indirectly diminish the activity of upstream RhoA by
stimulating Rac1 activity (24). FAK
is involved in cellular adhesion and mediates a key notable early
step in cell migration (25). MMP-2
and MMP-9 serve a pivotal role in mediating the malignant behavior
of cancer cells, including invasion and metastasis, by degrading
the extracellular matrix (26). In
the present study, zerumbone was demonstrated to decrease the
migration and invasion of hepatoma cells. In addition, the mRNA and
protein expression of RhoA, ROCK-1, FAK, MMP-2 and MMP-9 was
inhibited following zerumbone treatment in HepG2 cells. These
results indicated that suppressing tumor metastasis could be
achieved through regulating reorganization of the actin
cytoskeleton via Rho GTPase signaling pathways.
The MAPK pathways serve a notable role in mediating
the survival of mammalian cells and tumor metastasis (27). The MAPK family includes JNK, p38 MAPK
and ERK. p38 MAPK phosphorylation has been implicated to serve a
notable role in cell apoptosis, and activation of p38 MAPK
decreases ERK1/2 activity (28). The
results of the present study demonstrated that the phosphorylation
of P38 MAPK was upregulated and phosphorylation of ERK1/2 was
downregulated following zerumbone treatment of HepG2 cells but did
not exhibit a significant influence on total JNK protein
expression. These results indicated that regulation of ERK1/2 and
p38 MAPK serves a critical role in restraining cancer cells from
invasion and metastasis and inducing apoptosis and cell cycle
arrest in response to zerumbone treatment in HepG2 cells.
In conclusion, considering the results of the
present study, we hypothesize that zerumbone effectively inhibits
the proliferation, and invasion and migration of hepatoma cells
in vitro. We hypothesize that the inactivation of ERK1/2 and
activation of p38 MAPK are important initiating signals of the
mitochondrial-mediated apoptosis induced and invasion and
metastasis restrained by zerumbone. These results indicated that
zerumbone might be a potential anticancer agent for the treatment
of hepatoma. However, the present study preliminarily investigated
several molecules involved in the MAPK pathways, and rescue
experiments would be required to confirm the findings and further
demonstrate how zerumbone regulates hepatoma invasiveness.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TL designed the study, WZ performed the experiment
and XH performed the analysis and wrote the paper.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gong Q, Qin Z and Hou F: Improved
treatment of early small hepatocellular carcinoma using sorafenib
in combination with radiofrequency ablation. Oncol Lett.
14:7045–7048. 2017.PubMed/NCBI
|
|
2
|
Qiu X, Huang Y, Zhou Y and Zheng F:
Aberrant methylation of TRIM58 in hepatocellular carcinoma and its
potential clinical implication. Oncol Rep. 36:811–818. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang Y, Chen SW, Fan CC, Ting LL, Kuo CC
and Chiou JF: Clinical parameters for predicting radiation-induced
liver disease after intrahepatic reirradiation for hepatocellular
carcinoma. Radiat Oncol. 11:892016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xiao Y, Tian Q, He J, Huang M, Yang C and
Gong L: MiR-503 inhibits hepatocellular carcinoma cell growth via
inhibition of insulin-like growth factor 1 receptor. Onco Targets
Ther. 9:3535–3544. 2016.PubMed/NCBI
|
|
5
|
Yoo T, Lee KW, Yi NJ, Choi YR, Kim H, Suh
SW, Jeong JH, Lee JM and Suh KS: Peri-transplant change in AFP
level: A useful predictor of hepatocellular carcinoma recurrence
following liver transplantation. J Korean Med Sci. 31:1049–1054.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun J, Liu BR, Hu WJ, Yu LX and Qian XP:
In vitro anticancer activity of aqueous extracts and ethanol
extracts of fifteen traditional Chinese medicines on human
digestive tumor cell lines. Phytother Res. 21:1102–1104. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hsiao WL and Liu L: The role of
traditional Chinese herbal medicines in cancer therapy-from TCM
theory to mechanistic insights. Planta Med. 76:1118–1131. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun YW, Qiu HC, Ou MC, Chen RL and Liang
G: Saponins isolated from Schizocapsa plantaginea inhibit human
hepatocellular carcinoma cell growth in vivo and in vitro via
mitogen-activated protein kinase signaling. Chin J Nat Med.
16:29–40. 2018.PubMed/NCBI
|
|
9
|
Pan JX, Chen G, Li JJ, Zhu QD, Li JJ, Chen
ZJ, Yu ZP and Ye LY: Isocorydine suppresses doxorubicin-induced
epithelial-mesenchymal transition via inhibition of ERK signaling
pathways in hepatocellular carcinoma. Am J Cancer Res. 8:154–164.
2018.PubMed/NCBI
|
|
10
|
Kitayama T: Attractive reactivity of a
natural product, zerumbone. Biosci Biotechnol Biochem. 75:199–207.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rahman HS, Rasedee A, Yeap SK, Othman HH,
Chartrand MS, Namvar F, Abdul AB and How CW: Biomedical properties
of a natural dietary plant metabolite, zerumbone, in cancer therapy
and chemoprevention trials. Biomed Res Int. 2014:9207422014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kirana C, McIntosh GH, Record IR and Jones
GP: Antitumor activity of extract of Zingiber aromaticum and its
bioactive sesquiterpenoid zerumbone. Nutr Cancer. 45:218–225. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sobenin IA, Bobryshev YV, Korobov GA,
Borodachev EN, Postnov AY and Orekhov AN: Quantitative analysis of
the expression of caspase 3 and caspase 9 in different types of
atherosclerotic lesions in the human aorta. Exp Mol Pathol. 99:1–6.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen L, Liu H, Liu J, Zhu Y, Xu L, He H,
Zhang H, Wang S, Wu Q, Liu W, et al: Klotho endows hepatoma cells
with resistance to anoikis via VEGFR2/PAK1 activation in
hepatocellular carcinoma. PLoS One. 8:e584132013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Poortahmasebi V, Poorebrahim M, Najafi S,
Jazayeri SM, Alavian SM, Arab SS, Ghavami S, Alavian SE, Moghadam
Rezaei A and Amiri M: How hepatitis C virus leads to hepatocellular
carcinoma: A network-based study. Hepat Mon. 16:e360052016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garrido C, Galluzzi L, Brunet M, Puig PE,
Didelot C and Kroemer G: Mechanisms of cytochrome c release from
mitochondria. Cell Death Differ. 13:1423–1433. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Antonsson B: Mitochondria and the Bcl-2
family proteins in apoptosis signaling pathways. Mol Cell Biochem.
256–257:141–155. 2004. View Article : Google Scholar
|
|
19
|
Deng Y, Lin Y and Wu X: TRAIL-induced
apoptosis requires Bax-dependent mitochondrial release of
Smac/DIABLO. Genes Dev. 16:33–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stetler-Stevenson WG, Aznavoorian S and
Liotta LA: Tumor cell interactions with the extracellular matrix
during invasion and metastasis. Annu Rev Cell Biol. 9:541–573.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chang CT, Weng WH, Chou AS, Chuang CK,
Porwit-McDonald A, Pang ST, Larsson C and Liao SK: Immunophenotypic
and molecular cytogenetic features of the cell line UP-LN1
established from a lymph node metastasis of a poorly-differentiated
carcinoma. Anticancer Res. 25:683–691. 2005.PubMed/NCBI
|
|
22
|
Lin HJ, Su CC, Lu HF, Yang JS, Hsu SC, Ip
SW, Wu JJ, Li YC, Ho CC, Wu CC and Chung JG: Curcumin blocks
migration and invasion of mouse-rat hybrid retina ganglion cells
(N18) through the inhibition of MMP-2,-9, FAK, Rho A and Rock-1
gene expression. Oncol Rep. 23:665–670. 2010.PubMed/NCBI
|
|
23
|
Ren XD, Kiosses WB and Schwartz MA:
Regulation of the small GTP-binding protein Rho by cell adhesion
and the cytoskeleton. EMBO J. 18:578–585. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi H, Cheng Y, Ye J, Cai P, Zhang J, Li
R, Yang Y, Wang Z, Zhang H, Lin C, et al: bFGF promotes the
migration of human dermal fibroblasts under diabetic conditions
through reactive oxygen species production via the
PI3K/Akt-Rac1-JNK pathways. Int J Biol Sci. 11:845–859. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu Y, Zhang X, Qi L, Cai Y, Yang P, Xuan
G and Jiang Y: HULC long noncoding RNA silencing suppresses
angiogenesis by regulating ESM-1 via the PI3K/Akt/mTOR signaling
pathway in human gliomas. Oncotarget. 7:14429–14440.
2016.PubMed/NCBI
|
|
26
|
Bauvois B: New facets of matrix
metalloproteinases MMP-2 and MMP-9 as cell surface transducers:
Outside-in signaling and relationship to tumor progression. Biochim
Biophys Acta. 1825:29–36. 2012.PubMed/NCBI
|
|
27
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi L, Lin Q, Yang T, Nie Y, Li X, Liu B,
Shen J, Liang Y, Tang Y and Luo F: Oral administration of Lentinus
edodes β-glucans ameliorates DSS-induced ulcerative colitis in mice
via MAPK-Elk-1 and MAPK-PPARγ pathways. Food Funct. 7:4614–4627.
2016. View Article : Google Scholar : PubMed/NCBI
|